
Abstract
Porphyra tenera (PT), an edible red alga, is consumed as a health-promoting seaweed. Human norovirus (HuNV), the major cause of foodborne disease outbreaks worldwide, binds to histo-blood group antigens (HBGAs) for cell entry. We investigated the inhibitory activity of fucose-containing polysaccharides in PT on HuNV binding to the HBGAs. With saliva as a source of HBGAs and recombinant P domains of HuNV GII.4 and GII.17 purified as NV antigens, the inhibition and binding affinity of PT toward P domains were evaluated using enzyme-linked immunosorbent assay and bio-layer interferometry, respectively. The PT polysaccharide extract and its 3–10 kDa fraction (F3-10) among molecular weight fractions inhibited the P domain binding to saliva significantly, compared with that of a commercial fucoidan. F3-10 bound directly to the P domains with submicromolar affinities (KD = 5.6 × 10−7 M for GII.4 and 8.7 × 10−7 M for GII.17). When applied to virus inhibition assays, F3-10 significantly reduced murine NV titers in a dose-dependent manner; specifically, it showed a 1.8 log10 plaque forming unit (PFU)/mL reduction at 1 mg/mL in RAW 264.7 cells (p < 0.05) and a 1.4 log10 PFU/mL reduction at 5 mg/mL under simulated human digestion conditions involving sequential incubation in simulated saliva, gastric, and intestinal fluids (p < 0.01). F3-10 contained galactose, glucose, fucose, and xylose as its constituent monosaccharides and also sulfate groups. The PT fraction F3-10 is a promising candidate for further study aimed at inhibiting HuNV binding to host cell HBGAs.
Keywords
Introduction
Noroviruses (NVs) are highly contagious pathogens causing foodborne diseases (Lindesmith et al., 2003; Bányai et al., 2018). NV outbreaks impose a substantial burden on global health and economy (Bartsch et al., 2016) and frequently occur during winter seasons, referred to as winter vomiting disease, primarily transmitted via the fecal-to-oral route. NVs belong to the Caliciviridae family with 10 genogroups, including genogroups infecting humans, GI, GII, GIV, GVIII, and GIX (Chhabra et al., 2019). The GII.4 genotype has been the predominant pathogen for the past decades, and GII.17 was confirmed in sporadic outbreaks worldwide during the 2023/2024 season (Chhabra et al., 2024; Du et al., 2021).
The NV genome contains three open reading frames (ORFs): ORF1 encodes nonstructural polyproteins; ORF2 and ORF3 encode VP1 and VP2, the major and minor capsid proteins, respectively. VP1 is composed of protruding (P) and shell (S) domains. The P-domain of human NV (HuNV) is known to interact with histo-blood group antigens (HBGAs), as attachment factors for NV adhesion of GI.1, GI.3, GII.4, and GII.17 to the cell surface, though in a strain-specific manner (Koromyslova et al., 2017; Prasad et al., 2025; Ayyar et al., 2025). Some GII.4 NV strains induce capsid reorganization through binding to HBGAs for viral entry (Ayyar et al., 2025). HuNV interacts with both human HBGAs in saliva and human milk oligosaccharides (Huang et al., 2005; Kubota et al., 2012). GII P domains interact with the fucose moiety of the HBGAs and human milk oligosaccharides, 2′-fucosyllactose and 3-fucosyllactose, which inhibit HuNV–HBGA interactions (Koromyslova et al., 2015a; Weichert et al., 2016).
HuNV can be cultivated in B cells and human intestinal enteroid-derived organoids, which is limited by the lack of indefinite serial viral passaging and the requirement for converting 3D organoids into 2D monolayers (Prasad et al., 2025). Despite CD300lf being its specific receptor, murine NV (MNV) is a valuable surrogate for studying HuNV infection due to its ability to propagate in cell lines and genetic and biochemical similarities (Graziano et al., 2020; Song et al., 2020).
Porphyra tenera (PT; laver), known as gim or nori, is an edible red alga, and its global production reached ∼8% of the total algae production (FAO, 2021). PT contains ∼46% carbohydrate, with porphyran, a sulfated polysaccharide, accounting for 10% of the total content (Yim et al., 2021). We hypothesized that PT polysaccharides containing ∼5% of fucose (Yim et al., 2021; Park et al., 2022) inhibit HuNV binding. In the absence of approved antivirals or vaccines against HuNV (Ayyar et al., 2025), this study describes the inhibition of NV by PT.
Materials and Methods
Preparation and fractionation of PT extract
The PT polysaccharide extract (hereafter PT extract) was prepared as previously described (Takahashi et al., 2000). Dried PT (80 g) was ground using a blender, suspended in 1600 mL distilled water, autoclaved at 120°C for 1 h, and filtered through a cheesecloth, which was centrifuged at 8000 × g at 4°C for 1 h. Two volumes of 95% ethanol were added to the supernatant to precipitate polysaccharides, which were centrifuged at 12,000 × g for 30 min. The precipitate was lyophilized and stored at −80°C, and the final yield was 27.8%(w/w). The PT extract was fractionated using ultrafiltration membranes of molecular weight cutoff of 3 and 10 kDa (Millipore Corp., MA, USA) and lyophilized: ≤3 kDa, 3–10 kDa (F3-10), and ≥ 10 kDa fractions. A schematic diagram of the preparation and characterization of the F3-10 from PT is shown in Supplementary Figure S1.
Expression and purification of HuNV P domains
The P domains from HuNV GII.4 and GII.17 were expressed and purified (Kim et al., 2020). DNA fragments encoding HuNV GII.4 and GII.17 P domains (GenBank accession numbers BAI49908.1 and LC037415.1, respectively) with an N-terminal His-tag were synthesized by Macrogen (Seoul, Korea). Each DNA fragment was cloned into the pET14b vector (Novagen, Madison, USA) and transformed into Escherichia coli BL21 (DE3) (Novagen). The cells were sonicated and centrifuged at 12,000 × g for 20 min. The supernatant containing the P domain was purified by a nickel–nitrilotriacetic acid (Ni–NTA) affinity chromatography (Qiagen, Hilden, Germany) and size exclusion chromatography with a Superdex 200 increase column (Cytiva, Marlborough, MA, USA). The size of the purified P domain was verified by sodium dodecylsulfate-polyacrylamide gel electrophoresis (SDS-PAGE).
Enzyme-linked immunosorbent assay
The inhibition of binding of P domains to A- and O-type saliva by the PT extract or F3-10 was examined (Kim et al., 2020). Saliva samples from volunteers at Duksung Women’s University were pooled by blood group types and centrifuged after heat treatment. Supernatants were coated onto Maxisorp 96-well plates at 4°C overnight, blocked with 5% nonfat dried milk for 1 h at 37°C. Recombinant P domains were preincubated with PT extract at 1 mg/mL or F3-10 at 0.125–1 mg/mL at 4°C overnight. The mixtures were incubated in saliva-coated plates for 2 h at 37°C. The bound P domains were determined by GII.4 or GII.17 rabbit polyclonal antibody and horseradish peroxidase-conjugated goat anti-rabbit antibody. A commercial fucoidan from Undaria pinnatifida (≥95% purity, Sigma-Aldrich, St. Louis, MO, USA) with a significant inhibitory activity against P domain binding was used as a positive control (Kim et al., 2020). The inhibition (%) was calculated as: (1 − OD450 [with PT sample]/OD450[without PT sample]) × 100, where the absorbance at 450 nm without PT sample was taken as 100% binding to the saliva. The experiment was performed in triplicate.
Bio-layer interferometry
The binding affinities between the P domain and F3-10 were measured as previously reported (Lee et al., 2023). The P domains and F3-10 were used at 0.2 mg/mL and 1.25 − 40 mg/mL, respectively. The binding affinities were analyzed by bio-layer interferometry (BLI) using a GatorPrime (Gator Bio, Palo Alto, CA, USA) at ambient temperature with shaking at 1000 rpm. The association and dissociation, via the Ni–NTA biosensors, with immobilized P domain with F3-10 were measured for 300 s each. The plots were fitted using the GatorPrime analysis software (version 12.0).
Viruses and cells
RAW 264.7 cells (American Type Culture Collection, Manassas, VA, USA) were grown in Dulbecco’s modified Eagle’s medium (DMEM, Gibco BRL, Karlsruhe, Germany) supplemented with 10% fetal bovine serum (FBS, Sigma-Aldrich) and 1% penicillin–streptomycin (PS, Invitrogen, Grand Island, NY, USA) at 37°C in 5% CO2. MNV-1 (MNV; provided by Dr. Herbert Virgin, USA) was propagated in RAW 264.7 cells at 37°C for 1 h and incubated in DMEM (10% FBS, 1% PS) for 72 h. Infected cells underwent three freeze–thaw cycles, and the lysates were centrifuged and filtered (0.22 μm). The filtrate was ultracentrifuged at 96,589 × g through a 30% sucrose cushion for 3 h at 4°C. The purified MNV (4 × 7 log10 plaque-forming unit [PFU]/mL) were resuspended in 20% glycerol and stored at −80°C.
Cytotoxicity and plaque assay
For cytotoxicity screening, RAW 264.7 cells were seeded in 96-well plates in DMEM-FBS-PS. After 24 h incubation at 37°C and 5% CO2, 90 µL of DMEM-FBS-PS and 10 µL of F3-10 were incubated in the cells at 37°C and 5% CO2 for 24 h. The 3-(4,5-dimethylthiazol-2-yl)−2,5-diphenyltetrazolium bromide and dimethyl sulfoxide (Sigma-Aldrich) were added to the plate. The cell viability was calculated using the equation: the cell viability (%) = (treated OD570/control OD570) × 100. The experiment was conducted in triplicate.
A time-of-addition plaque assay was performed to focus on whether F3-10 directly interacts with MNV particles. For pretreatment of viruses by F3-10, MNV suspension was treated with F3-10 at a 1:1 ratio at room temperature for 3 h (a final titer of 6 log10 PFU/mL). The pretreated viral suspension was serially diluted 10-fold in DMEM and inoculated onto cell monolayers at 37°C and 5% CO2 for 1 h. The inocula were discarded, and 1 mL DMEM containing 1% agarose, 5% FBS, and 0.5% PS were added. After 48 h incubation at 37°C and 5% CO2, the cells were fixed in 4% formaldehyde for 1 h at room temperature. The agarose overlay was discarded, 0.5% crystal violet was added, and the number of plaques was counted. For co-treatment, the same experimental procedure as the pretreatment of the MNV was conducted, except that confluent cell monolayers were infected simultaneously with MNV (6 log10 PFU/mL) and F3-10 and incubated at 37°C in 5% CO2 for 1 h. For the post-treatment, MNV (6 log10 PFU/mL) was adsorbed onto the cells, which were incubated with F3-10 for 1 h after aspiration. The remaining procedure was the same as described for the pretreatment of the MNV. The experiment was performed in triplicate.
Antiviral effects of F3-10 in a simulated human digestion model
Simulated saliva fluid (SSF), simulated gastric fluid (SGF), and simulated intestinal fluid (SIF) were prepared according to the in vitro digestion method (Minekus et al., 2014). MNV suspension (5 mL of 6–7 log10 PFU/mL) was mixed with 5 mL of SSF containing human α-amylase (Sigma-Aldrich) at 5 mg/mL of F3-10 and incubated for 10 min at pH 7.0. SGF (10 mL) containing porcine gastric pepsin (Sigma-Aldrich) was added (at 2.5 mg/mL of F3-10) and incubated for 2 h at pH 3.0. Finally, 20 mL SIF containing porcine pancreatin and bile (Sigma-Aldrich) was added to the mixture and incubated for 2 h at pH 7.0 (at 1.25 mg/mL of F3-10). All incubation procedures were performed at 37°C in a shaking water bath at 120 strokes/min. The mixture in DMEM (10% FBS) was inoculated onto confluent monolayers of RAW 264.7 cells with a 2-fold serial dilution. The remaining steps of the plaque assay were described above. The control experiment was performed without F3-10 under the same conditions. The experiment was performed in triplicate.
Monosaccharide composition and sulfate content of the F3-10
Monosaccharide composition of the F3-10 was analyzed as previously described (Manns et al., 2014). F3-10 and 2 M trifluoroacetic acid (TFA, Sigma-Aldrich) were mixed and heated at 121°C for 2 h. Hydrolysates were dried under N2. For HPLC analyses, the dried F3-10 sample was dissolved in distilled water, neutralized, and filtered through a 0.2 µm filter. The monosaccharide content was analyzed using a Dionex ICS-5000 ion chromatography (Thermo Scientific Dionex; Waltham, MA, USA) with a CarboPac SA 10 (4 × 250 mm, Dionex; Sunnyvale, CA, USA) column. The flow rate was 0.5 mL/min, and the mobile phase was water and 200 mM NaOH at a 99.2:0.8 ratio (v/v).
The sulfate content of the F3-10 was determined as described previously (Tabatabai, 1974; Kim et al., 2020). The barium–gelatin reagent (50 µL) and 250 µL of TFA-hydrolyzed F3-10 sample were added to microtiter plates and stirred for 1 h, and absorbance was measured at 450 nm. The standard curve was generated using anhydrous Na2SO4 at 0–10 mM.
Statistical analysis
IBM SPSS Statistics (version 26, IBM Corp, New York, USA) was used, in which the data were analyzed by analysis of variance, and the mean values were compared with a Tukey’s test at the 5% significance level. A t-test was performed to compare two different groups; p-values < 0.01 were considered significant.
Results
Expression and purification of recombinant P domains of HuNV
The recombinant P domains of HuNV GII.4 and GII.17 were expressed in E. coli BL21 (DE3) and purified using affinity and size exclusion chromatography. Size exclusion chromatographic elution profiles of P domains of HuNV GII.4 and GII.17 are shown in Figure 1A and B. The molecular weights of purified P domains were estimated to be ∼35 kDa by SDS-PAGE.
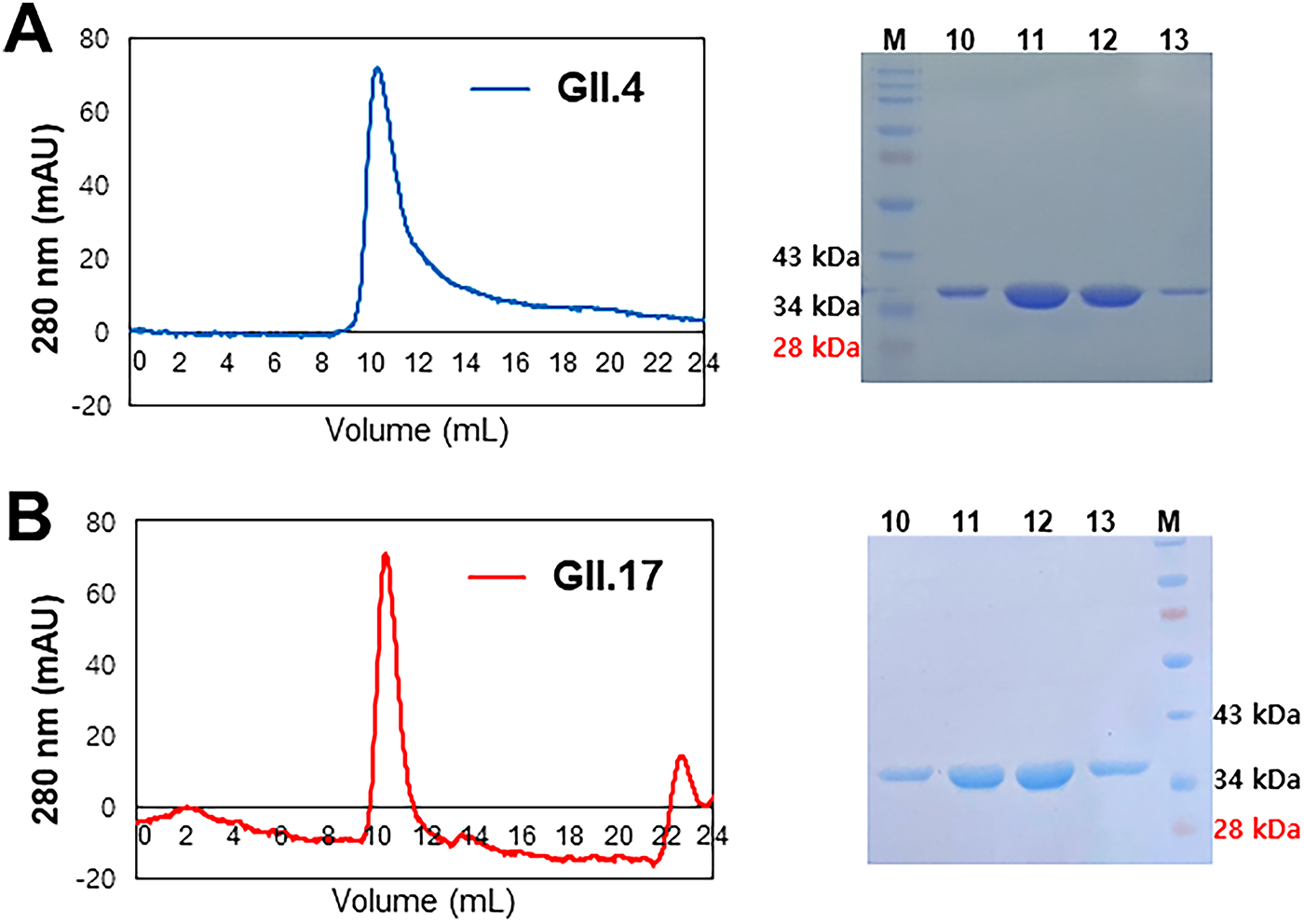
Chromatographic elution profiles of P domains of HuNV GII.4 and GII.17 at the final size exclusion steps. The HuNV
Inhibition of P domain binding to receptors by PT extract and fractions
PT extract at 1 mg/mL showed 93% and 85% inhibition against the binding of the HuNV GII.4 P domain to A- and O-type saliva, respectively (Fig. 2A). The extract revealed 70% and 51% inhibition against the binding of the GII.17 P domain to the two types of saliva at the same concentration, respectively (Fig. 2B). Fucoidan at 1 mg/mL, a positive control, showed 19–37% inhibition on the binding of the P domains to saliva. The extract was ultrafiltered into three fractions using the molecular weight cutoff membrane filters (≤3 kDa, 3–10 kDa, and ≥ 10 kDa). The 3–10 kDa fraction (F3-10) showed the most significant inhibitory effect, reaching 71–82% binding inhibition comparable to that of PT extract (Fig. 2A). F3-10 also inhibited the binding of the GII.17 P domain to saliva significantly by 49–61% (Fig. 2B).
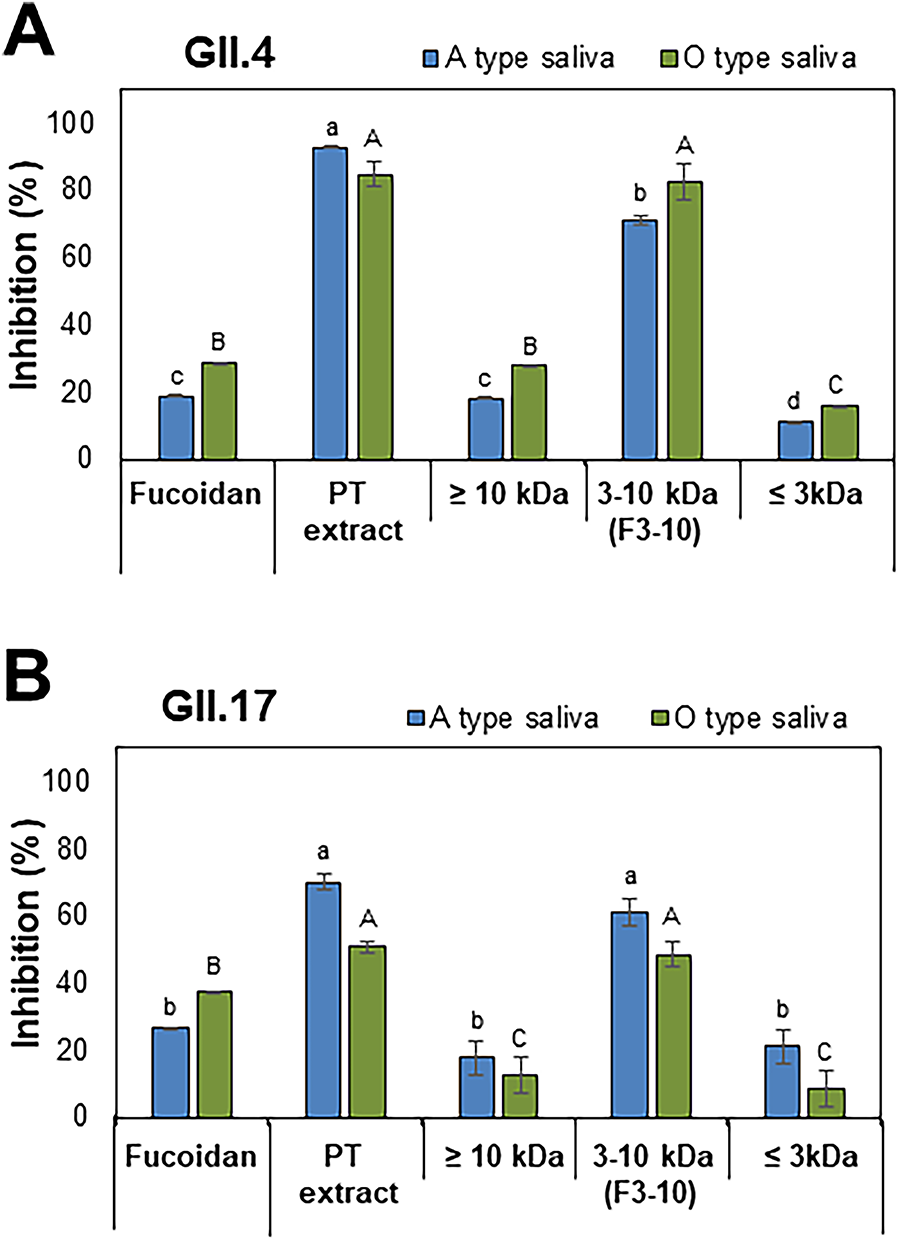
Inhibitory effects of the PT polysaccharide extract (PT extract) and its fractions on the binding of the P domains of HuNV to saliva.
The effect of F3-10 on blocking the binding of the P domains to saliva was dose-dependent at concentrations from 0.125 to 1 mg/mL (Fig. 3A,B). BLI analysis showed strong affinities of F3-10 to the GII.4 and GII.17 P domains, with dissociation constants (KD) of 5.6 × 10−7 M and 8.7 × 10−7 M, respectively (Fig. 4A,B).
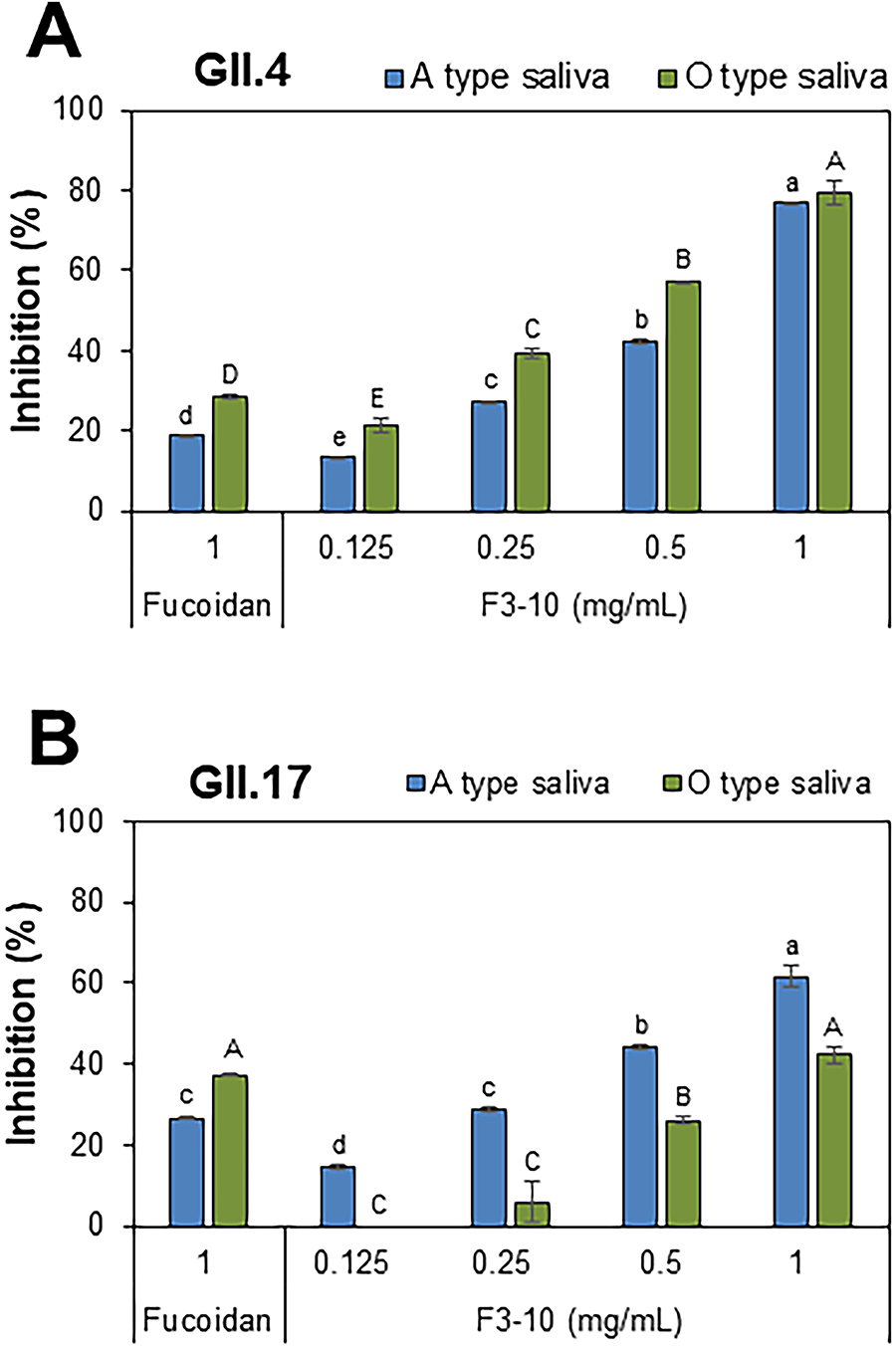
Dose-dependent inhibition of F3-10 on binding of the HuNV P domains to saliva.
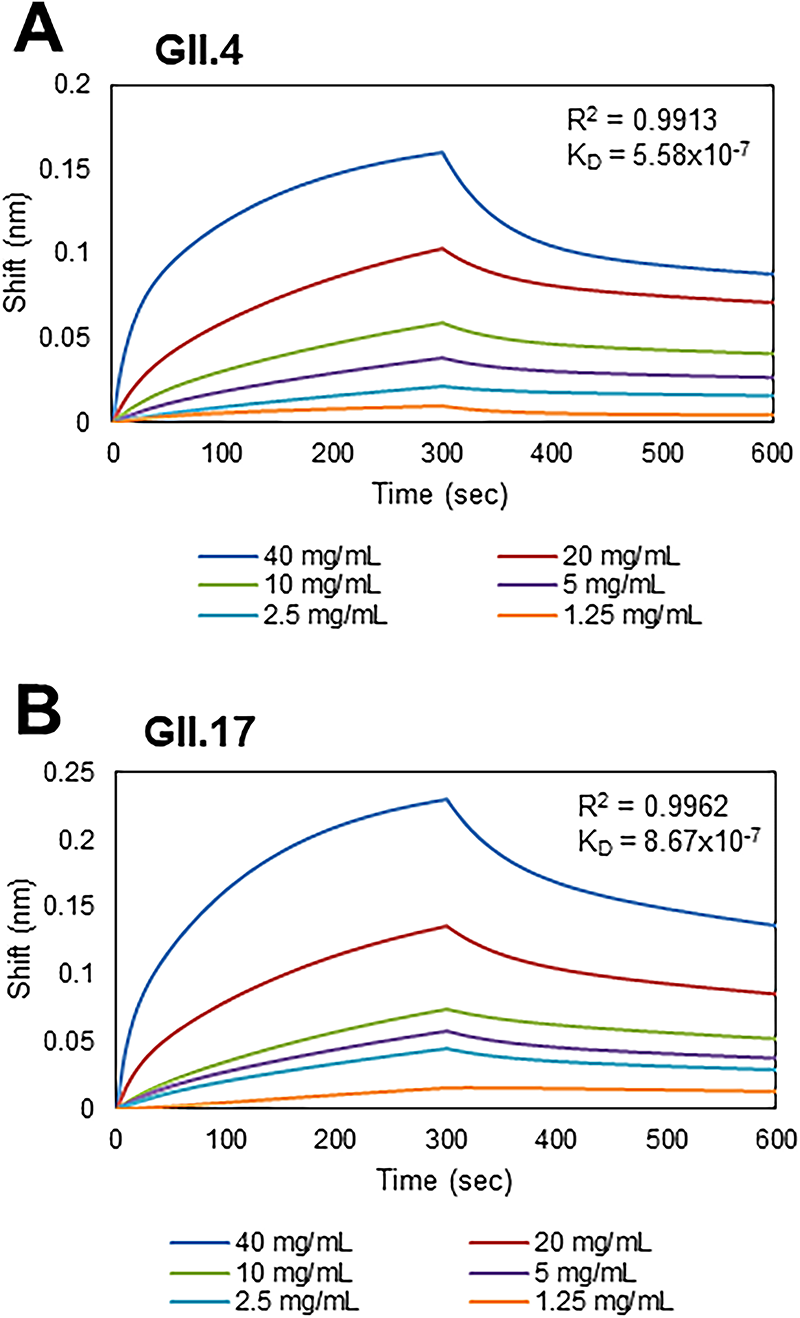
Affinities of F3-10 on binding the P domains of HuNV to saliva. The binding of
Viral titer reduction by F3-10 in RAW 264.7 cells and simulated human digestion models
The inhibitory effects of F3-10 were further evaluated by plaque assays against MNV, a surrogate for HuNV, in RAW 264.7 cells (cell viability was 99% at 1 mg/mL of F3-10). The treatment of the virus with F3-10 significantly reduced MNV titer at 0.0625 to 1 mg/mL in a dose-dependent manner (p < 0.05), and the reduction titer was 1.8 log10 PFU/mL at 1 mg/mL (Fig. 5A).
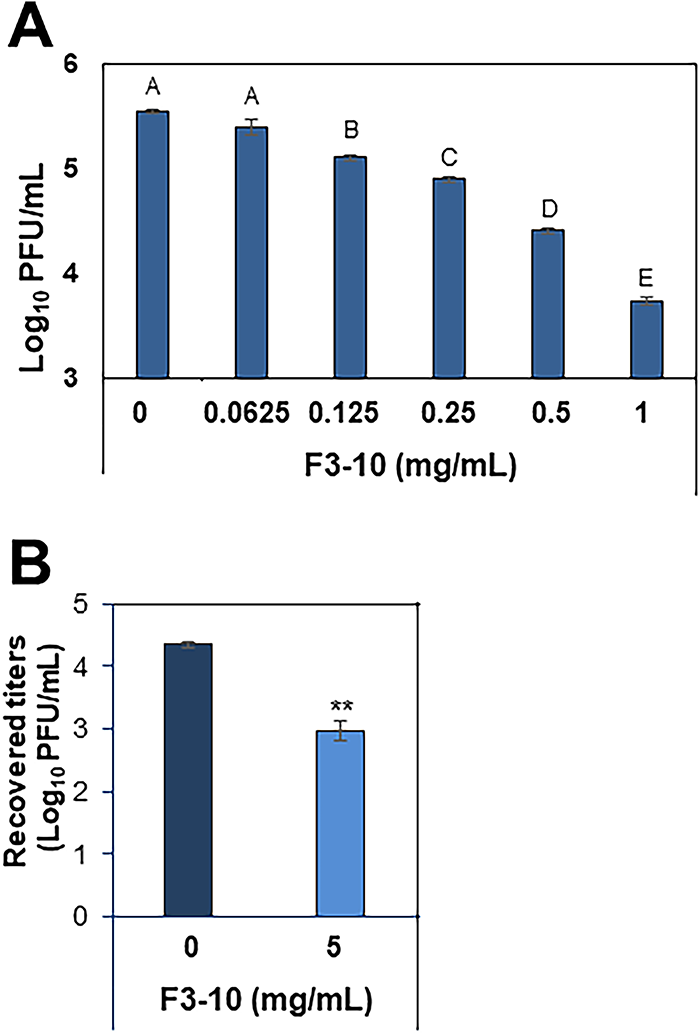
Plaque assay-based antiviral effects of F3-10 against MNV, a HuNV surrogate.
The antiviral activity of F3-10 was evaluated in an in vitro digestive system simulating human digestion. The experiment utilized simulated digestive fluids, including saliva, gastric, and intestinal fluid. At 5 mg/mL, F3-10 significantly reduced MNV titers by 1.4 log10 PFU/mL (p < 0.01), compared with MNV alone in simulated digestive fluids (Fig. 5B).
Monosaccharide composition of F3-10
The monosaccharide composition and sulfate content of F3-10 were examined by HPLC and the barium-gelatin method, respectively. The predominant monosaccharide was galactose (17.0%), followed by glucose (16.5%), fucose (1.5%), and xylose (1.5%) (Table 1). The sulfate content of F3-10 was 45.2%. These data suggest that F3-10 is a fucose-containing sulfated polysaccharide.
Monosaccharide and Sulfate Profiles of the F3-10

Discussion
Laver (Porphyra tenera), typically consumed as a side dish or seasoned snack, is a nutritious, low-calorie, and convenient ready-to-eat food. The main carbohydrate of laver is porphyran, a complex polysaccharide composed of repeating units of galactose esterified with sulfate groups, but the number of sulfate groups per unit varies. It exhibits significant bioactivities, including antiallergenic and anticancer effects (Ishihara et al., 2005; Liu et al., 2019).
The GII.4 genotype has become endemic in ongoing outbreaks, and the GII.17 genotype has also been increasingly detected in infection (Chen et al., 2020; Chhabra et al., 2024; Du et al., 2021). It was reported that extracts of Chinese gall and pomegranate, as well as tannic acid, a common compound in both extracts, inhibited GII.4 P protein binding to types A and B saliva (Zhang et al., 2012). Citrate from lemon juice was bound to the HBGA binding pocket of the GII. 10 P domain, showing a potential as a competitive inhibitor against HBGA binding (Hansman et al., 2012; Koromyslova et al., 2015b).
In this study, the extract and its fraction F3-10 were prepared from PT, which strongly inhibited the binding of GII.4 and GII.17 P domains to A- or O-type saliva. Among the three molecular weight fractions from the extract, F3-10 (3–10 kDa fraction) showed the most significant and dose-dependent inhibition to the binding of the P domains to saliva (Fig. 2A,B). The inhibition was much greater than that of a commercial fucoidan, a complex sulfated galactofucan (Zhao et al., 2018). Many studies have shown that NV interactions with HBGAs are crucial for the initial binding of HuNV to host cells (Huang et al., 2005; Weichert et al., 2016; Ayyar et al., 2025). Studies also show that fucose, as part of HBGAs or Lewis-like antigens, plays an essential role in NV binding and its structural arrangement and exposure as a nonreducing terminal residue are critical (Koromyslova et al., 2015a; Hanisch et al., 2021; Hanisch, 2023). The F3-10 from PT consists primarily of galactose (17.0%), glucose (16.5%) and fucose (1.5%), which is likely to possess a galactan-type core with short oligofucose ends. Fucoidan from Undaria pinnatifida is known to be composed of fucose (50.9%) and galactose (44.6%) (Zhao et al., 2018), and its processed and low molecular weight fraction, which has a β-6-galactane backbone with oligo-fucose branches, shows a high inhibitory effect on virus-like particle binding. In this context, the structural arrangement and exposure of fucose at the nonreducing terminal ends are critical for NV binding, rather than the total fucose content.
F3-10 binds to the P domains derived from GII.4 and GII.17 with KD values of 5.6 × 10−7 M and 8.7 × 10−7 M, respectively. The binding affinities to P domain are several hundred times greater than those of the HBGA-like trisaccharide, fucose, or citrate (KD values of 390–460 µM) (Hansman et al., 2012). Our results demonstrate that F3-10 exhibited submicromolar affinities to the GII.4 and GII.17 P domains, which, together with the dose-dependent inhibition of F3-10 to P domain binding to saliva, suggest direct inhibition of the binding of the HuNV particles to cell receptors.
The application of F3-10 to the antiviral effect on MNV particles showed a 1.8-log10 PFU/mL reduction in MNV titer in the pretreatment and much less reduction in the co- and post-treatment (Supplementary Fig. S2). F3-10 may block the binding of MNV to host cell receptors or inhibit the viral internalization into RAW 264.7 cells. MNV and HuNV use different host receptors, CD300lf and HBGAs, respectively, although the exact primary receptor for HuNVs is still investigated (Graziano et al., 2020). We postulate that F3-10 may have a common element of action against the P domain of HuNV or MNV, preventing infection by blocking their ability to bind to receptors, or that multiple components in F3-10 may block binding of the HuNV P domains to receptors with submicromolar KDs, a critical step for HuNV infection, highlighting the potential of F3-10 as an antiviral material. We are currently purifying a single compound from F3-10 using HPLC and BLI. Further structural analysis is needed to understand how a potential component in F3-10 interacts with the HBGA receptor. In conclusion, PT extract and F3-10, a galactose-containing polysaccharide, strongly inhibit the binding of HuNV GII.4 and GII.17 P domains to saliva and showed strong submicromolar affinities to the P domains.
Authors’ Contributions
H.K.: Investigation (lead), methodology (equal), visualization, and data curation (equal). J.A.L. and Y.L.: Investigation (supporting) and data curation (equal). K.H.K.: Conceptualization (equal), methodology (equal), data curation (equal), resources (equal), and writing—review and editing (equal). M.S.C.: Conceptualization (equal), data curation (equal), resources (equal), writing—original draft, writing—review and editing (equal), and funding acquisition. All authors have read and agreed to the published version of the article.
Footnotes
Funding Information
This work was supported by grants of the National Research Foundation (2021R1A2C2009539, M.S.C.).
Disclosure Statement
No competing financial interests exist.
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.