
Abstract
Staphylococcus aureus, a common cause of bovine mastitis, relies on several virulence factors, with biofilm formation being a key contributor to its pathogenicity. The present study investigated the occurrence of S. aureus as etiological agents in bovine mastitis with a focus on the existence of various virulence factors and antibiotic resistance status. Among 120 milk samples collected from West Bengal and Uttar Pradesh, 36 (30%) S. aureus strains were confirmed by conventional methods and PCR. Phenotypic analysis revealed hemolysin (55.55%) and coagulase production (36.11%), while molecular analysis revealed the presence of leukotoxin (luksF, 19.44%), hemolysin (hlb, 58.33%), coagulase (coa, 63.88%), and toxic shock syndrome toxin (tsst-1, 30.55%) genes. Biofilm production ability was detected in 97.22% (crystal violet assay) and 86.11% (Congo red agar assay) strains. Biofilm-associated genes, namely, icaA (80.55%, 29/36), icaB (75%, 27/36), icaC (69.44%, 25/36), icaD (86.11%, 31/36), and MSCRAMMs genes, namely, clfA (58.33%, 21/36), clfB (75%, 27/36), fnbA (75%, 27/36), fnbB (55.55%, 20/36), bap (38.88%, 14/36), bbp (83.33%, 30/36), ebps (69.44%, 25/36), eno (66.66%, 24/36), fib (41.66%, 15/36), and cna (8.33%, 3/36), were also detected. Antimicrobial resistance was observed in 88.88% isolates, with 72.22% exhibiting multidrug resistance (MDR). Among the isolates, 83.33% were methicillin-resistant S. aureus (MRSA), and mecA, femA, and femB genes were present either singly or in combination in 76.66% of the isolates. Efflux pump protein genes, namely, norA, norB, norC, mdeA, mepA, and sepA, were detected either singly or in combination in S. aureus isolates. 61.53% of MDR-MRSA isolates harbored all six efflux pump genes. According to this study, S. aureus of mastitis origin harbors various virulence, antibiotic resistance, biofilm-forming, and efflux pump genes. Bovine mastitis-derived MDR S. aureus isolates can pose a significant public health risk and need urgent attention to formulate strategies for their control and preventing transfer to the human food chain.
Significance and Impact of the Study
The study provides a glimpse of antibiotic resistance pattern, biofilm-producing ability, and the virulence potential of Staphylococcus aureus strains involved in bovine mastitis. The majority of the tested isolates were found to be resistant to multiple classes of antimicrobial agents. High incidence of MRSA among bovine mastitic milk represents one of the major public health threats due to the possibility of transmission to human beings. Most of the isolates had biofilm-production capability, harbored biofilm-related and efflux pump protein-encoding genes, indicating potential virulent bacteria that can enter and persist in the human food chain.
Introduction
Staphylococcus aureus is a major foodborne pathogen that can affect both humans and animals and is associated with a range of diseases. Studies have reported its occurrence in meat and dairy products (Hassan et al., 2022). In animals, S. aureus is a leading cause of bovine mastitis, which has a significant effect on the quantity and quality of milk production. Mastitis is an inflammatory infection of the mammary glands posing serious economic losses to the dairy industry (Akkou et al., 2024; Campos et al., 2022). Clinical and subclinical mastitis enhances antimicrobial usage, significantly impacting the emergence of antimicrobial resistance.
S. aureus is a Gram-positive bacterium that resides on the skin and mucosa of humans and animals. However, pathogenic S. aureus strains carry different virulence factors and cause clinical or subclinical mastitis (Jesse et al., 2023). Several secreted and cell surface-associated virulence factors produced by S. aureus promote adhesion and invasion to host cells, biofilm formation, and immune evasion (Pugazhendhi et al., 2022). S. aureus is considered the third most important cause of foodborne disease in the world, caused by contaminated milk and milk products (Cardamone et al., 2024; Patel et al., 2021; Santos et al., 2020). In recent years, the emergence and spread of antimicrobial-resistant S. aureus, especially multidrug-resistant (MDR) and methicillin-resistant S. aureus (MRSA) strains, has been an important public health risk (Silva et al., 2023). The extrusion of multiple substrates through the efflux pump is a predominant mechanism of antibacterial resistance.
Previous studies have demonstrated that this bacterium can induce different types of mastitis, including clinical (subacute, acute, and chronic) and subclinical (Fazal et al., 2023; Sharifi et al., 2023). Effective attachment and colonization of S. aureus are important steps in the development of mastitis. By expressing various adhesion proteins, S. aureus can adhere to and colonize the mammary gland tissues, leading to the establishment of infection and subsequent mastitis (Campos et al., 2022; Graber et al., 2013).
Bacteria may form biofilms, which are organized microbial communities encased in a self-produced extracellular matrix, once they firmly adhere to and colonize a surface (O’Toole et al., 2000). Biofilm formation in S. aureus occurs through polysaccharide intercellular adhesion (PIA) and microbial surface components recognizing adhesive matrix molecules (MSCRAMMs) (Foster, 2005; Mirzaee et al., 2015; O’Gara, 2007; Periasamy et al., 2012). These structures mediate the S. aureus initial attachment to both host tissues and biomaterials (Atshan et al., 2012). Biofilm formation interferes with bacterial recognition and killing mechanisms by the innate immune system and also prevent access of antibiotics to bacterial population (Foster, 2005). PIA is a crucial factor in S. aureus biofilm formation. PIA is a cationic, partially deacetylated homopolymer of N-acetylglucosamine, and its biosynthesis is governed by the ica locus (encoded by icaADBC genes). PIA plays a key role in the adhesion and aggregation stages of biofilm development, facilitating cell-to-cell and cell-to-surface interactions. S. aureus strains may differ in the prevalent constellations of MSCRAMMs and also can predispose the host to certain kinds of infections through binding to molecules such as collagen, fibronectin, and fibrinogen (Archer et al., 2011; Foster and Höök, 1998; Kwon et al., 2008). S. aureus can express up to 20 different adhesive MSCRAMMs that are covalently anchored by sortase to peptidoglycan via the C-terminal LPXTG motif (Archer et al., 2011). These adhesion proteins include the ClfA and ClfB (clumping factors A and B), FnbA and FnbB (fibronectin-binding proteins A and B), Fib (fibrinogen-binding protein), Cna (collagen-binding protein), Eno (laminin-binding protein), Ebp (elastin-binding protein), Bap (biofilm-associated protein), and Bbp (bone sialoprotein-binding protein). Biofilms can develop on various surfaces, including medical devices, tissues, and biological surfaces, such as the mammary gland in the case of bovine mastitis. Biofilms provide numerous advantages to bacteria, including increased resistance to antimicrobial compounds compared with their planktonic or free‐floating counterparts. Accordingly, combating the biofilm form of bacteria is crucial in controlling infectious diseases (Sharifi et al., 2018; 2021).
To determine possible hazards of S. aureus strains causing mastitis, it is crucial to know variations in phenotype and genotype among S. aureus isolates in terms of virulence potential, resistance to antibiotics, and zoonotic potential (particularly concerning MRSA) (Patel et al., 2021). This information shall help in formulating suitable management strategies at the herd level. This study was aimed at investigating antibiotic resistance profiles, virulence, biofilm, and efflux pump-related genes in S. aureus strains recovered from intramammary infections of dairy herds to support herd-level disease management.
Materials and Methods
Sample collection and processing
Dairy farms of West Bengal and Uttar Pradesh, and Teaching Veterinary Clinical Complex, ICAR-IVRI were enrolled in the study from February to November 2023. Ten milliliters of milk samples were collected from clinical and subclinical mastitis cases as assessed by field veterinarian in a sterile sample container containing brain heart infusion (BHI) semisolid media (0.3% agar) (HiMedia, India). Samples were transported to the Bacteriology Laboratory, Division of Biological Products, ICAR-IVRI under cold chain and incubated aerobically at 37°C for 24 h.
Isolation and identification of S. aureus
Samples were inoculated into mannitol salt broth (HiMedia, India) and incubated for 24 h at 37°C, and subsequently plated on mannitol salt agar and Baird Parker agar (BPA) (HiMedia, India) plates. The plates were incubated overnight at 37°C and observed for colony characteristics. Gram’s staining was carried out to observe the characteristic microscopic morphology. The S. aureus isolates were identified using conventional biochemical tests, namely, catalase, oxidase and various other tests performed using Hi-Staph test strips (HiMedia, India) as described previously (Mishra, 2018).
DNA extraction and PCR confirmation
An isolated colony of bacteria suspected to be S. aureus, identified by morphological and biochemical tests, was inoculated in Luria Bertani broth (HiMedia, India) and incubated overnight at 37°C. Genomic DNA was extracted using the HiPurA Bacterial Genomic DNA Purification Kit (HiMedia, India) following the manufacturer’s protocol. DNA purity was assessed by UV spectrophotometry at OD 260/280 using EPOCH 2.0 (BioTek, USA), and the extracted DNA was stored at −20°C until further use.
The isolates were further confirmed genotypically as S. aureus by polymerase chain reaction (PCR) amplification of the species-specific thermonuclease (nuc) gene, following the methodology of Mishra (2018). The 25 µL PCR reaction contained 2.5 µL of 10× PCR buffer, 2.5 µL of 25 mM MgCl2, 0.5 µL of 10 mM dNTPs, 0.5 µL each of forward and reverse primers (10 pM/µL), 1 µL of template DNA (∼50 ng), and nuclease-free water. S. aureus ATCC 43300 was used as the positive control, and a no-template reaction served as the negative control. PCR conditions included initial denaturation at 95°C for 5 min, followed by 36 cycles of denaturation at 95°C for 45 sec, annealing at 63°C for 45 sec, and extension at 72°C for 45 sec, with a final extension at 72°C for 10 min. The amplified nuc gene products were loaded on 1.5% agarose gel containing ethidium bromide (0.5 µg/mL), electrophoresed at 100 V for 1 h, and visualized using a gel imaging system (AlphaImager EC, UK) (Dai et al., 2019). Primer details are provided in Supplementary Table S1.
Detection of hemolysin and coagulase production
S. aureus isolates were cultured on blood agar medium (HiMedia, India) enriched with 5% defibrinated sheep blood to determine the hemolytic activity of the isolates. In brief, a colony of S. aureus from the BPA plate was streaked on the blood agar plate and incubated for 24 h at 37°C. The complete or partial zone of hemolysis was observed after completion of incubation. To detect coagulase production by tube test, 0.1 mL S. aureus broth culture was inoculated in tubes containing 0.5 mL of reconstituted rabbit plasma (HiMedia, India), incubated at 37°C, and observed for clumping at every 4 h time interval up to 24 h.
Antimicrobial susceptibility test
The Kirby–Bauer disc diffusion method was used to assess the susceptibility of bacterial strains to various antibiotics (HiMedia, India). Briefly, 100 µL of test bacterial culture in BHI broth (0.5 McFarland standard) was spread on Mueller–Hinton agar plates, and antibiotic discs were evenly placed on the plates. The plates were incubated at 37°C overnight. To assess the susceptible, resistant, or intermediate profiles of isolates, the zone of inhibition was measured (in mm) and compared with Clinical and Laboratory Standards Institute criteria (CLSI, 2024; Ersanli et al., 2023). Amoxicillin-clavulanic acid (20/10 µg), cefepime (30 µg), erythromycin (15 µg), ceftriaxone (10 µg), oxacillin (5 µg), cloxacillin (5 µg), chloramphenicol (30 µg), cefotaxime (30 µg), tetracycline (10 µg), enrofloxacin (10 µg), ampicillin (10 µg) and amikacin (30 µg) discs were used in this study.
Multiple antibiotic resistance index
The multiple antibiotic resistance (MAR) index is a practical and dependable method for figuring out antimicrobial resistance pattern. It is computed as the ratio of the number of antimicrobials to which an isolate exhibits resistance to the total number of antimicrobials to which the organism is exposed (Almuhayawi et al., 2023). MAR index of more than 0.2 suggests that antimicrobials are frequently used at the point of infection (Almuhayawi et al., 2023).
Phenotypic biofilm production assay
The biofilm-producing capacity of S. aureus isolates was assessed phenotypically using Congo red agar (CRA) assay and modified crystal violet assay (Manandhar et al., 2021).
CRA assay
The CRA plates were prepared with BHI agar (HiMedia, India) containing sucrose (SRL, India). Congo red dye (SRL, India) was added to the autoclaved media at 55°C. After solidification, agar plates were kept in an incubator for sterility check and stored at 4°C for further use. Fresh overnight-grown pure cultures of S. aureus were streaked on CRA plates and incubated for 24 h at 37°C. Strong, moderate, and non-biofilm-forming bacteria produced black, brown, and red colonies, respectively, on CRA plates.
Crystal violet assay
For the crystal violet assay (CVA), 200 µL of overnight-grown bacterial culture in BHI broth supplemented with 1% glucose was seeded into 96-well cell culture plates (CoStar, USA) and incubated at 37°C for 24 h. Planktonic cells were removed, and wells were washed three times with 1× phosphate buffer saline (PBS), air-dried, heat-fixed at 80°C for 30 min, and stained with 0.2% crystal violet for 30 min. Excess stain was removed by washing and tapped to dry. The bound dye was solubilized with 200 µL of absolute alcohol for 30 min. Absorbance was measured at 595 nm. The assay was performed in triplicate. Isolates were classified as biofilm producers if optical density (OD570) values exceeded the ODc (at least three times higher than the standard deviation [SD] around the mean cutoff value) and categorized as weak (ODc < OD ≤ 2ODc), moderate (2ODc < OD ≤ 4ODc), or strong (OD > 4ODc) biofilm formers based on standard ODc criteria (Dai et al., 2019).
Detection of methicillin-resistance genes by PCR
S. aureus isolates were subjected to the detection of various genes associated with methicillin-resistance, that is, mecA, femA, and femB genes. Three single-plex PCRs were used to investigate the occurrence of mecA, femA, and femB genes (Supplementary Table S1).
Detection of virulence genes by PCR
S. aureus isolates were subjected to the detection of the toxin-encoding genes, viz. toxic shock syndrome toxin-1 (tsst-1), coagulase (coaG), leukotoxin (Luks), α-hemolysin (hla), and β-hemolysin (hlb) by PCR using specific primers. Three single-plex PCRs for tsst-1, coaG, and Luks and one duplex PCR for hla and hlb were used to investigate the occurrence of virulence genes (Supplementary Table S1).
Detection of biofilm-associated genes by PCR
For detection of adhesion (clfA, clfB, fnbA, fnbB, fib, and cna) and biofilm-related (icaA, icaB, icaD, icaC, epbs, bap, bbp, and eno) genes, PCR was performed following the methodology used in a previous study (Mishra, 2018). Four single-plex PCR for icaA, fnbA, bap, bbp, and three multiplex PCR (icaB, icaC, icaD; fib, ebps, eno, cna; and clfA, clfB, fnbB) were used to investigate the occurrence of biofilm-producing genes (Supplementary Table S1).
Detection of efflux pump genes by PCR
The presence of chromosomally encoded efflux pump genes was examined in the isolated S. aureus strains. Specific primers for norA, norB, norC, mepA, sepA, and mdeA were used for the amplification (Suma et al., 2023). Three duplex PCR (norA, norC; norB, mepA, and sepA, mdeA) were used to investigate the diversity of efflux pump genes (Supplementary Table S1).
Statistical analysis
For the prevalence estimate, 95% confidence intervals (CIs) were calculated using the Wilson score method without continuity correction. Prevalence values and their corresponding 95% CIs were expressed as percentages. The association between methicillin resistance and MDR phenotype was assessed using Fisher’s exact test because of the small sample size. Agreement between phenotypic oxacillin resistance and mecA gene detection was evaluated using Cohen’s kappa coefficient. The strength of agreement was interpreted according to the kappa interpretation scale. For the crystal violet assay, the OD values of isolates within each biofilm category were expressed as mean ± SD. The association between MDR phenotype and the presence of each efflux pump gene was assessed using Fisher’s exact test. A p value below 0.05 was considered statistically significant.
Results
Isolation and identification of S. aureus from mastitic milk samples
Out of 120 clinical and subclinical mastitic milk samples, 36 isolates were confirmed as S. aureus (30%, 95% CI: 22.53–38.72) by culture, biochemical tests, and nuc gene PCR (670 bp), of which 24 were from subclinical cases and 12 from clinical cases (Supplementary Figs. S1 and S2, Supplementary Table S2). All isolates were able to ferment mannitol. A total of 20 (55.55%) isolates showed hemolysin production on sheep blood agar (Table 1), and 13 (36.11%) isolates showed coagulase production (Table 1), suggesting the pathogenic potential of S. aureus (Supplementary Figs. S3 and S4).
Phenotypic Hemolytic and Coagulase Production Ability of Staphylococcus Aureus Field Isolates

Note: Values represent the number of positive isolates. Percentages were calculated using the total number of confirmed S. aureus isolates as the denominator (n = 36).
Antibiotic resistance profile of field isolates of S. aureus and determination of MAR index
In the present study, the Kirby–Bauer disc diffusion method was used to assess the antimicrobial sensitivity of the obtained clinical isolates to identify drug-resistant S. aureus. The clinical isolates of S. aureus were evaluated for their susceptibility towards 12 different antibiotics (Table 2). A higher number of S. aureus showed resistance to oxacillin (83.33%), followed by cloxacillin (75%), amikacin (69.44%), cefotaxime (66.66%), ampicillin (63.88%), tetracycline (61.11%), and cefepime, ceftriaxone, erythromycin (55.55%), amoxyclav (52.77%), and enrofloxacin (50%). All the isolates were susceptible to chloramphenicol (100%). The class-wise phenotypic resistance profiles of the S. aureus isolates are as follows: penicillin, 83.33%; cephalosporin, 66.66%; tetracycline, 61.11%; aminoglycocycline, 69.44%; macrolides, 55.55%; and fluoroquinolones, 50%. A total of 26 (72.22%, 95% CI: 56.01–84.15) isolates were found to be resistant to three or more classes of antibiotics and therefore classified as MDR. 33.33% (12/36) of hemolytic isolates and 16.66% (6/36) of coagulase-positive isolates were MDR in nature. Four (11.11%) isolates were found susceptible to all the antimicrobial agents tested, whereas four (11.11%) isolates were resistant to two classes of antibiotics and two (5.55%) isolates were resistant to one class of antibiotics. MAR index revealed 30 isolates (83.33%) with MAR index more than or equal to 0.2 and 6 (16.67%) isolates with MAR index less than 0.2. However, none of the isolates had a MAR index of 1, meaning resistant to all the antimicrobials tested (Supplementary Table S3).
Antimicrobial Susceptibility Profile of Staphylococcus aureus Field Isolates
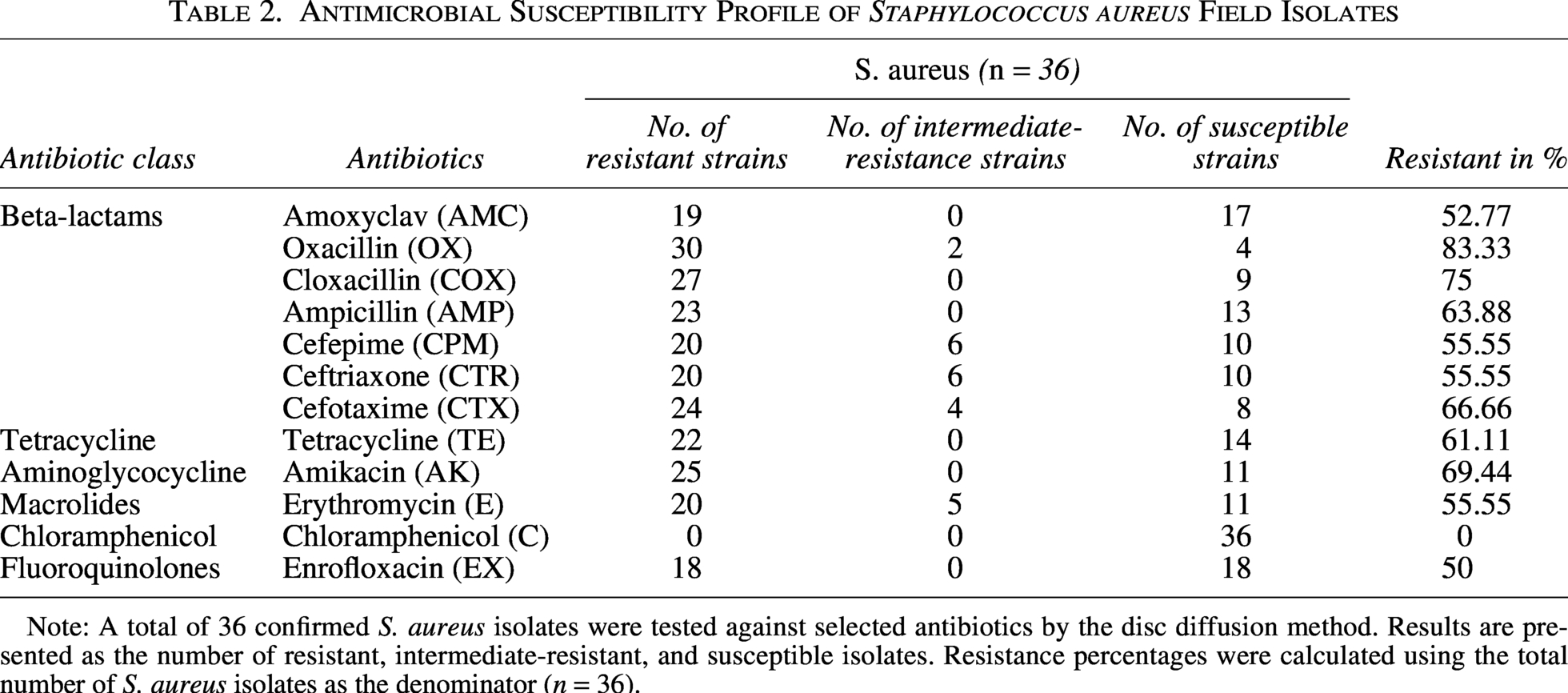
Note: A total of 36 confirmed S. aureus isolates were tested against selected antibiotics by the disc diffusion method. Results are presented as the number of resistant, intermediate-resistant, and susceptible isolates. Resistance percentages were calculated using the total number of S. aureus isolates as the denominator (n = 36).
Methicillin resistance in field isolates of S. aureus
Phenotypically, 30 of 36 isolates (83.33%, 95% CI: 68.11–92.13) were resistant to oxacillin by the disc diffusion method (Table 3). PCR targeting the mecA (hallmark of MRSA), femA, and femB genes of S. aureus was used to identify MRSA. Among the S. aureus isolates, 22.22% (8/36, 95% CI: 11.72–38.09) carried mecA gene, 36.11% (13/36) were positive for the femA gene, and 72.22% (26/36) were positive for femB gene (Fig. 1). None of the methicillin-sensitive S. aureus (n = 6) isolates showed MDR characteristics, whereas 26 MRSA isolates were MDR (Table 3). Fisher’s exact test showed a statistically significant association between MRSA and MDR phenotype (p = 0.0001). Out of 30 MRSA isolates, 23 (76.66%) harbored the mecA, femA, or femB gene singly or in combinations. All mecA-positive isolates were phenotypically resistant to oxacillin. However, 22 oxacillin-resistant isolates were negative for mecA. The agreement between phenotypic oxacillin resistance and mecA detection was 38.89% (using Cohen’s kappa coefficient), and Cohen’s kappa value was 0.11, indicating only slight agreement between the two methods. Among 26 MDR-MRSA isolates, 76.92% (20/26) harbored at least one gene singly or in combinations, while six strains were negative for all three genes.
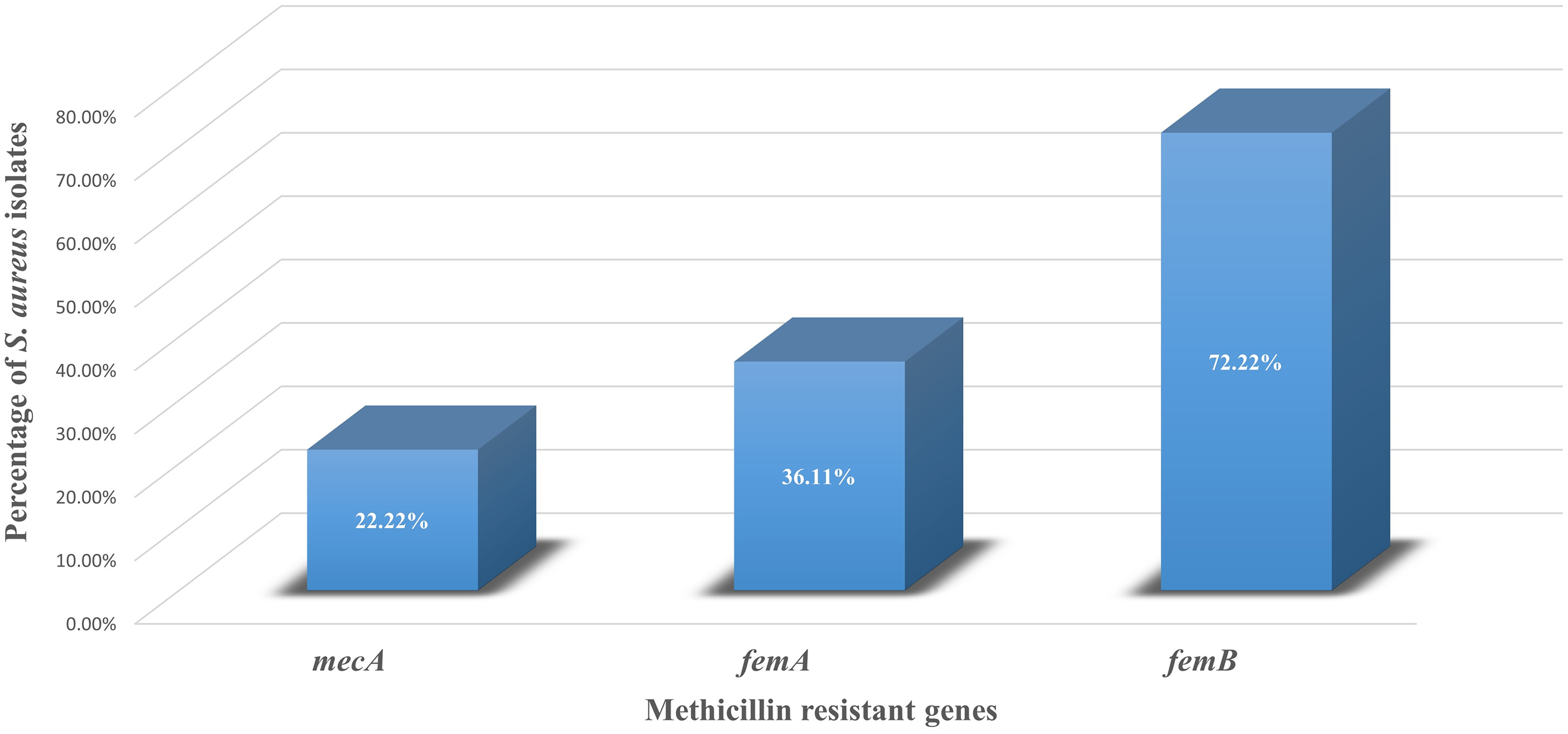
PCR-based detection of methicillin-resistance-associated genes in mastitis-associated Staphylococcus aureus isolates. S. aureus isolates (n = 36) were screened by PCR for mecA, femA, and femB genes. Results are expressed as percentage of gene-positive isolates. mecA, femA, and femB were detected in 22.22%, 36.11%, and 72.22% of isolates, respectively.
Classification of Different MRSA Types Among Field Isolates of Staphylococcus aureus
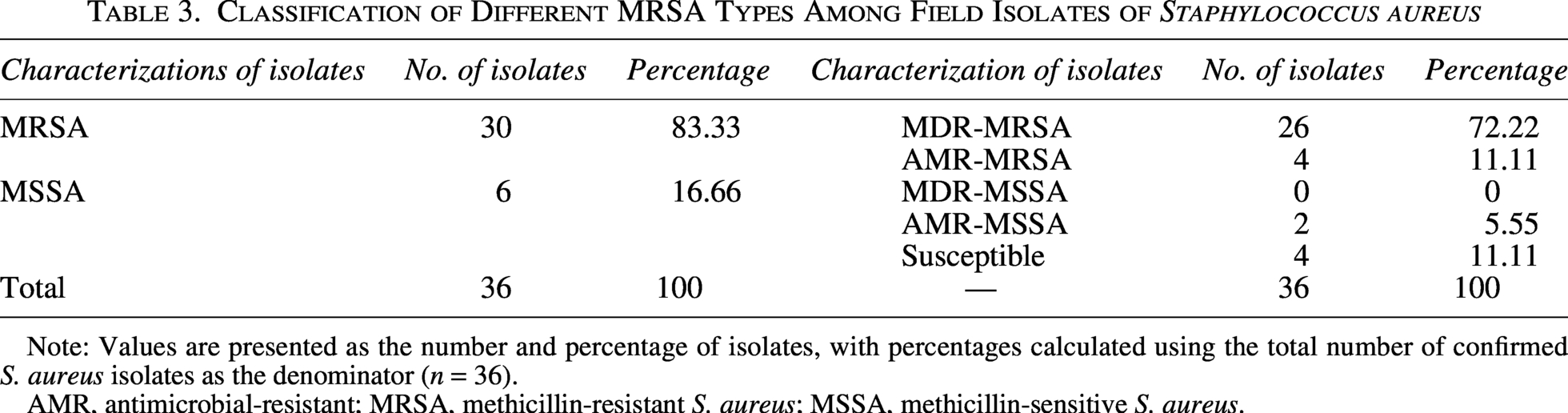
Note: Values are presented as the number and percentage of isolates, with percentages calculated using the total number of confirmed S. aureus isolates as the denominator (n = 36).
AMR, antimicrobial-resistant; MRSA, methicillin-resistant S. aureus; MSSA, methicillin-sensitive S. aureus.
Screening for in vitro biofilm production and biofilm-related genes in field isolates of S. aureus
All 36 S. aureus isolates were screened for biofilm formation by the CRA and CVA methods (Supplementary Table S4). Of these isolates, 86.11% (31/36, 95% CI: 71.34–93.92) formed black or brown colonies showing biofilm formation, whereas 13.88% (5/36) were nonformers (red/pink colonies) by CRA method. Among biofilm producers, 38.88% (14/36), 44.44% (16/36), and 2.77% (1/36) were strong, moderate, and weak biofilm producers by CRA method, respectively (Supplementary Fig. S5). The CVA results showed that 97.22% (35/36; 95% CI: 85.83–99.51) of the isolates were biofilm producers. Based on OD cutoff criteria, 80.55% (29/36) were classified as strong biofilm formers, with a mean OD value of 2.514 ± 0.710, 13.89% (5/36) as moderate biofilm formers, with a mean OD value of 1.494 ± 0.323, and 2.78% (1/36) as weak biofilm former, with an OD value of 0.475 ± 0.171 (Supplementary Fig. S6).
Biofilm-associated genes, that is, icaA (80.55%, 29/36), icaB (75%, 27/36), icaC (69.44%, 25/36), and icaD (86.11%, 31/36) and MSCRAMM genes, that is, clfA (58.33%, 21/36), clfB (75%, 27/36), fnbA (75%, 27/36), fnbB (55.55%, 20/36), bap (38.88%, 14/36), bbp (83.33%, 30/36), ebps (69.44%, 25/36), eno (66.66%, 24/36), fib (41.66%, 15/36), and cna (8.33%, 3/36), were found in S. aureus isolates (Supplementary Table S4; Fig. 2).
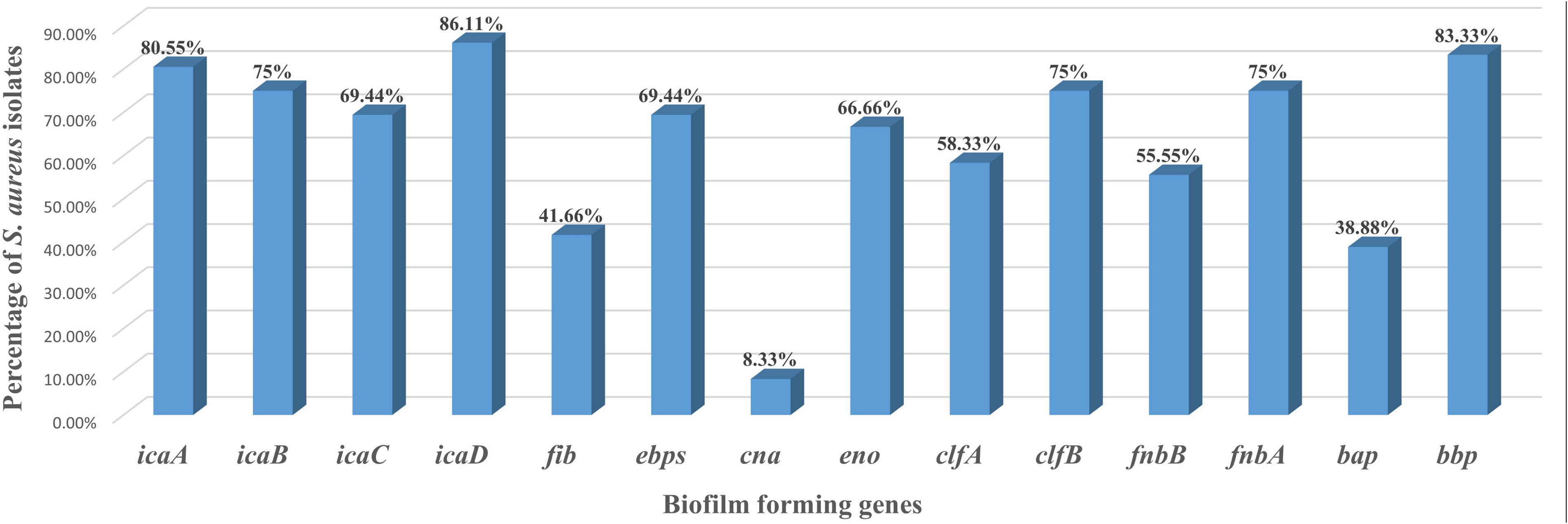
PCR detection of biofilm-associated and MSCRAMM genes in Staphylococcus aureus field isolates. Mastitis-associated S. aureus isolates (n = 36) were screened by PCR for biofilm-associated genes (icaA, icaB, icaC, icaD, and bap) and MSCRAMM genes (fib, ebps, cna, eno, clfA, clfB, fnbA, fnbB, and bbp). Results are expressed as the percentage of gene-positive isolates, calculated using the total number of S. aureus isolates as denominator. MSCRAMM, microbial surface components recognizing adhesive matrix molecule.
Occurrence of different virulence genes in the field isolates of S. aureus
The pyrogenic toxin gene tsst‐1 was found in 30.55% (11/36) of the S. aureus isolates, 21 isolates (58.33%) harbored hlb gene, 7 isolates (19.44%) possessed leukotoxins (luksF) gene, whereas hla gene was detected. The coagulase gene (coa) was positive in 23 S. aureus (63.88%) isolates (Fig. 3).
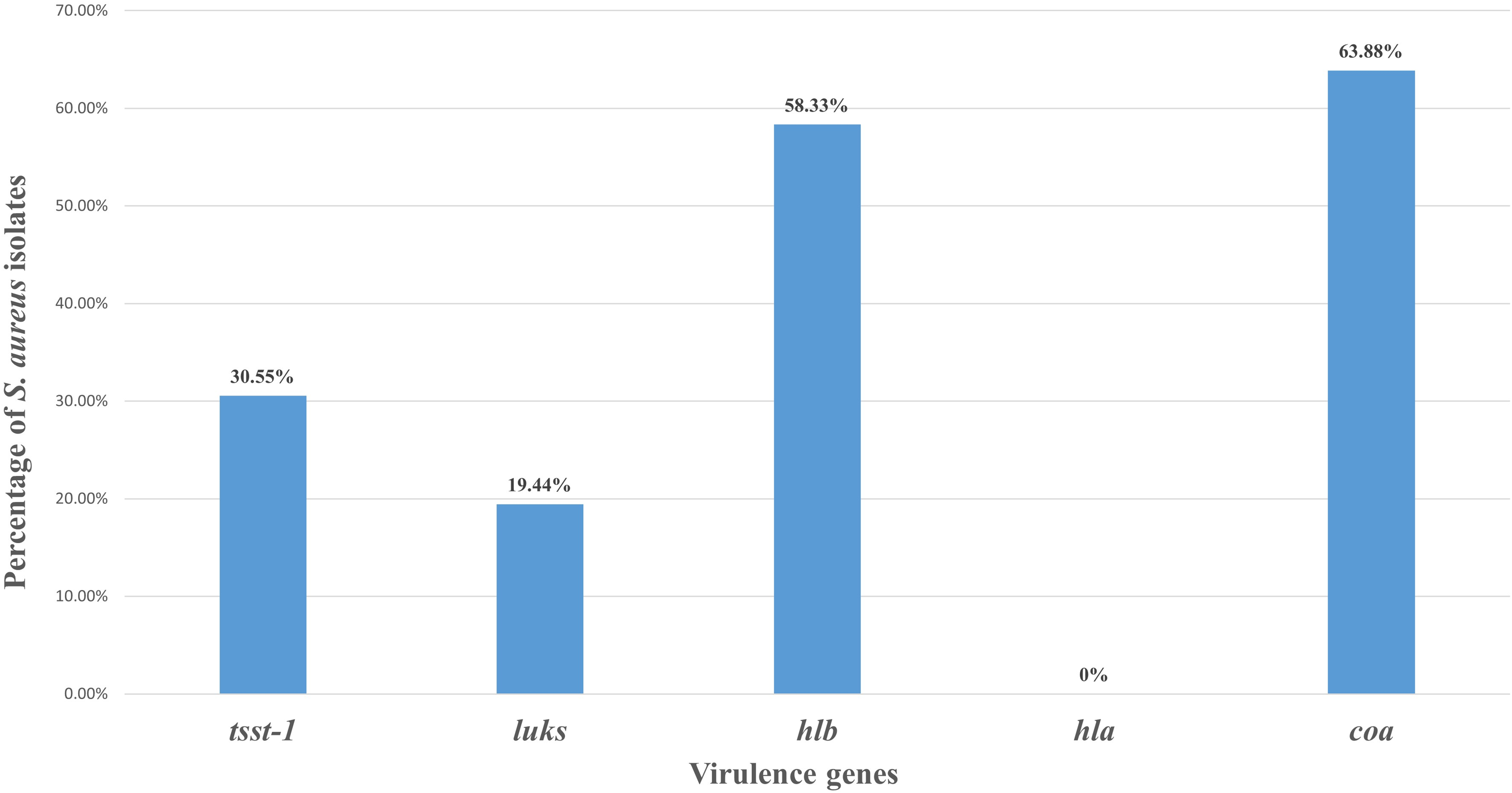
PCR detection of toxin and virulence genes in Staphylococcus aureus field isolates. Mastitis-associated S. aureus isolates (n = 36) were screened by PCR for selected virulence genes (tsst-1, lukS-F, hlb, hla, and coa). Results are expressed as the percentage of gene-positive isolates, calculated using total S. aureus isolates as denominator. hla was not detected in any of the S. aureus isolate.
Occurrence of different efflux pump genes in the field isolates of S. aureus
Efflux pump genes were identified in all S. aureus field isolates (n = 36). An extremely high prevalence of chromosomally encoded genes, including norA (83.33%, 30/36), norB (100%, 36/36), norC (86.11%, 31/36), mepA (72.22%, 26/36), sepA (83.33%, 30/36), and mdeA (94.44%, 34/36), was observed (Fig. 4). All six genes were present in 69.44% (25/36) of S. aureus isolates.
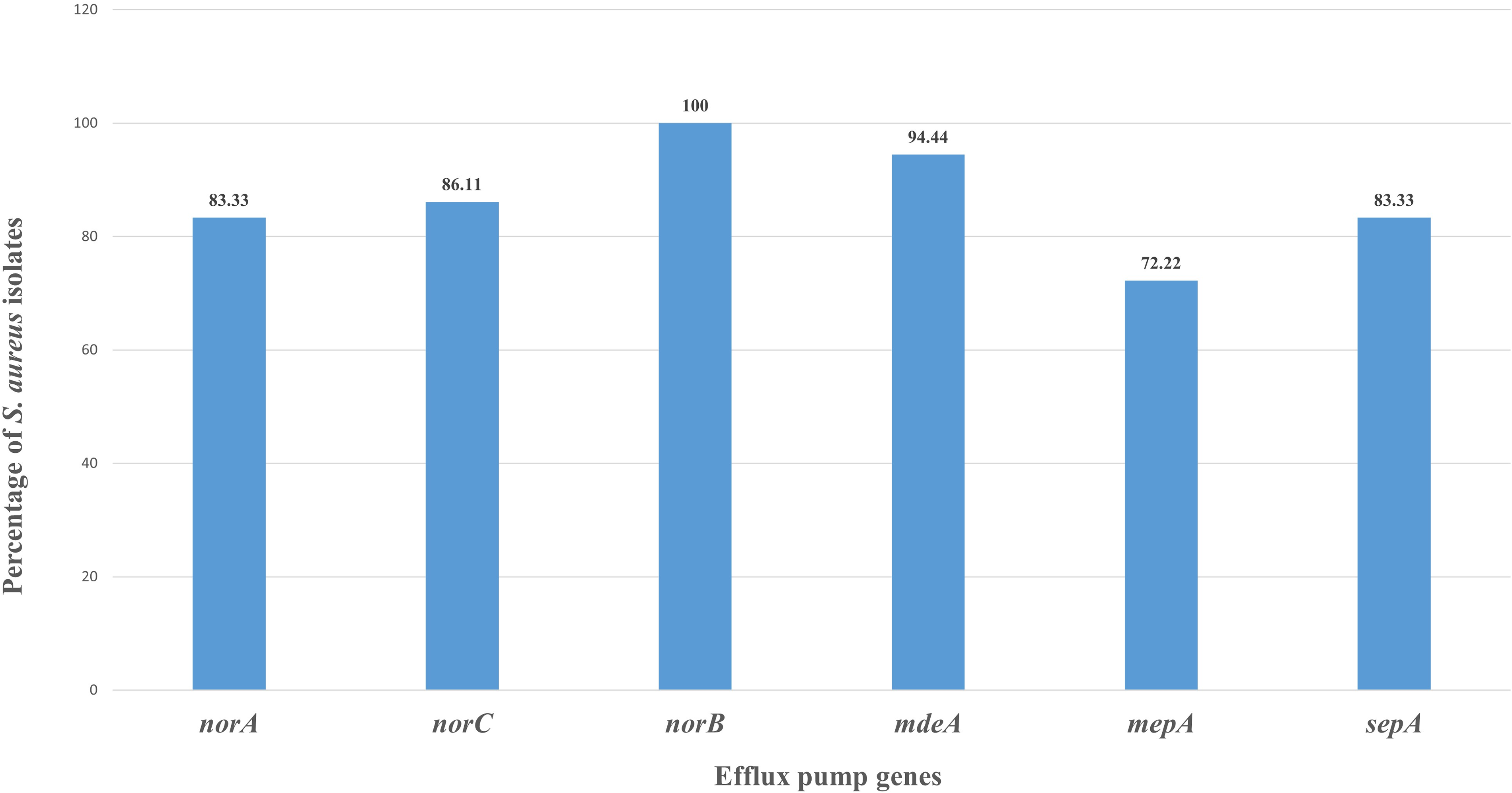
PCR detection of efflux pump genes in Staphylococcus aureus field isolates. Mastitis-associated S. aureus isolates (n = 36) were screened by PCR for efflux pump genes (norA, norB, norC, mepA, sepA, and mdeA). Results are expressed as the percentage of gene-positive isolates, calculated using total S. aureus isolates as denominator.
MDR profile associated with efflux pump gene combinations in field isolates of S. aureus
In the present study, the antibiotic resistance profile and efflux gene combinations in S. aureus isolates were evaluated. Six efflux pump-encoded genes were found in 25 antibiotic-resistant isolates (Fig. 5). Among the 26 MDR isolates, all carried norB, while 24/26 carried mdeA, 21/26 carried norC, 20/26 carried norA, 20/26 carried sepA, and 17/26 carried mepA gene. Sixteen MDR isolates carried all six efflux pump genes. All MDR isolates (26/36) carried norB gene and were resistant to the penicillin group of antibiotics. Among these, 16 isolates (61.53%) harbored all six efflux pump genes (norA, norB, norC, mdeA, mepA, sepA), and they were resistant to at least 3 classes of antibiotics. These 16 efflux pump gene-positive MDR isolates were resistant to penicillin, 13 isolates (81.25%) were resistant to cephalosporin, 11 isolates (68.75%) were resistant to tetracycline group, 13 isolates (81.25%) were resistant to aminoglycocyclines, 12 isolates (75%) were resistant to macrolides, and 10 isolates (62.5%) were resistant to fluoroquinolone antibiotics. However, Fisher’s exact test showed no statistically significant association between MDR status and any individual efflux pump gene or the presence of all six genes.
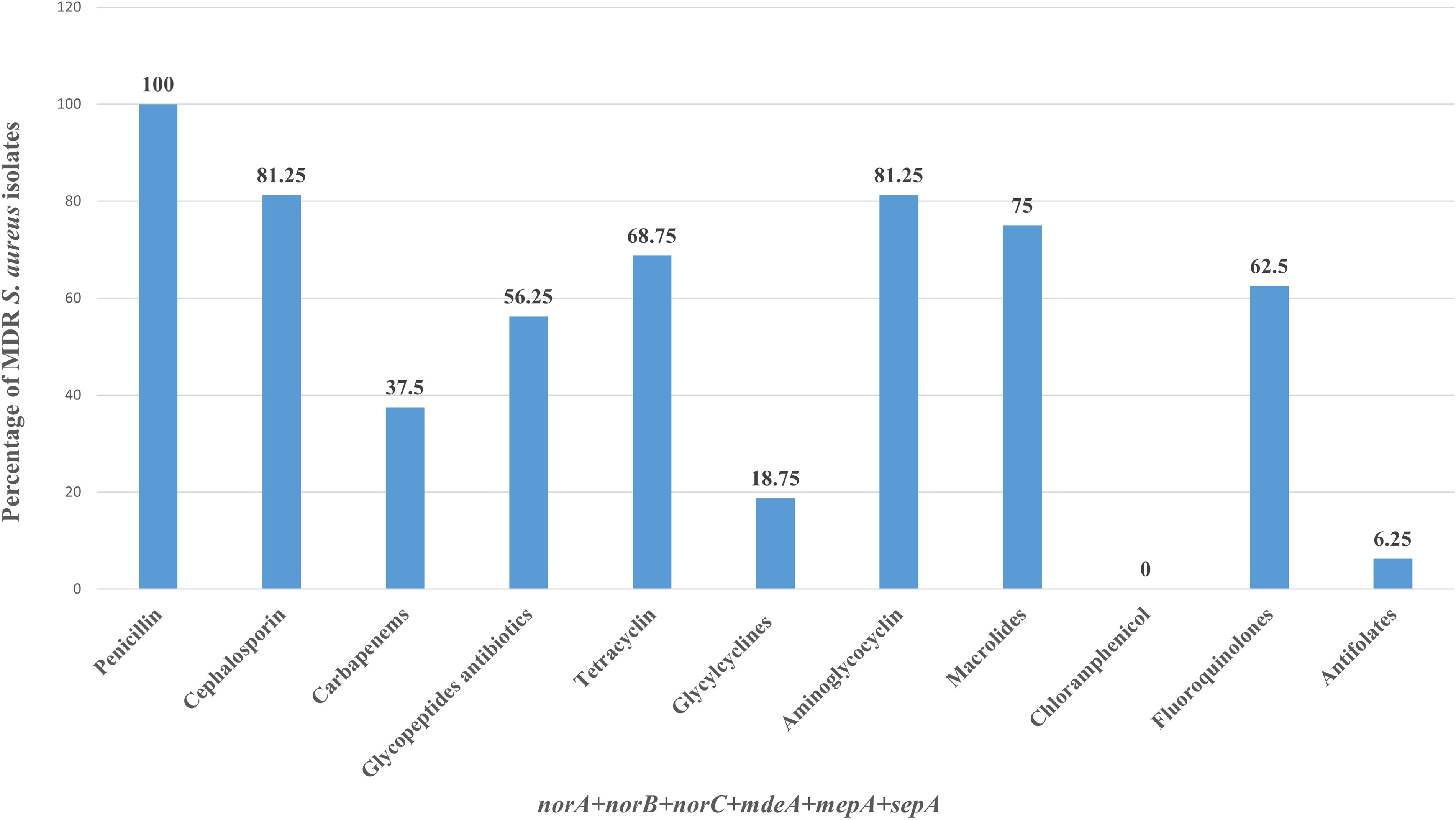
Antibiotic class resistance profile of MDR Staphylococcus aureus isolates carrying efflux pump genes. Mastitis-associated multidrug-resistant S. aureus isolates (n = 26) were analyzed for resistance to different classes of antibiotics and efflux pump gene carriage. Results are expressed as the percentage of MDR isolates showing resistance to different classes of antibiotics. MDR, multidrug-resistant.
Discussion
In the present study, 30% S. aureus isolates were recovered from milk samples collected in West Bengal and Uttar Pradesh from bovine mastitis cases and identified as one of the most important bacterial causes of bovine mastitis. The recovery of S. aureus from clinical and subclinical cases further highlights their relevance in persistent intramammary infection, leading to huge economic losses (Paramasivam et al., 2023). The occurrence of S. aureus is in concordance with earlier reports from India, although some variation across studies reflects differences in the geographical area, herd management, mastitis status, sampling strategy, and diagnostic methods used (Bhattacharyya et al., 2016; Hamid et al., 2017; Kutar et al., 2015; Roshan et al., 2022).
The isolates exhibited varying phenotypic and genotypic expression of virulence-associated characteristics. Coagulase activity (36.11%), hemolysis (55.55%), and the detection of genes for coa (63.88%), hlb (58.33%), tsst-1 (30.55%), and lukS/F (19.44%) indicate the pathogenic potential of the isolates. However, partial agreement between phenotype and presence of their genetic determinants suggests that virulence expression may be influenced by strain variation, gene regulation, and assay conditions. Previous research on S. aureus associated with bovine mastitis has also documented such differences or similarities (Azara et al., 2017; Elsayed et al., 2015; Fitzgerald et al., 2000; Momtaz et al., 2011; Neelam et al., 2022; Pérez et al., 2020; Vaughn et al., 2020; Monistero et al., 2020). Superantigenic toxins (tsst-1) located on pathogenicity island SaPI of S. aureus facilitate immunosuppression and promote the persistence of S. aureus (Kumar et al., 2020). The presence of hemolysin increases the adherence and proliferation of S. aureus to mammary epithelial cells, which contributes to biofilm formation, suggesting a synergistic action of these virulence mechanisms (Pérez et al., 2020). Therefore, rather than providing clear proof of the severity of the disease, the virulence profile should be interpreted as a sign of pathogenic potential.
The majority of the isolates in this study demonstrated the ability to form biofilms using the CRA assay (86.11%) and crystal violet assay (97.22%). The ica locus and MSCRAMMs, consisting of the adhesion genes, have been reported to have a potential role in biofilm production (Kumar et al., 2020). The frequent detection of biofilm-associated genes in the present study, particularly icaA (80.55%), icaD (86.11%), and adhesion-related genes, supports the role of biofilm formation. Our findings were compatible with the previous reports of Ahmed et al. (2023) and Singh et al. (2023). S. aureus that produces biofilms may be less susceptible to antimicrobial therapy and may survive better in the environment of the mammary gland. However, phenotypic biofilm assays have limitations as results can vary with inoculum size, incubation conditions, and the type of assay.
In the present study, antibiotic resistance was found to be common among the field isolates, with high resistance to oxacillin (83.88%), amikacin (69.44%), ampicillin (63.88%), and tetracycline (61.11%). A large proportion of isolates were MDR (72.22%), which is consistent with previous reports of increasing antimicrobial resistance among mastitis-associated S. aureus (Brahma et al., 2022; Hamid et al., 2017; Haq et al., 2024; Neelam et al., 2022; Ramasamy et al., 2020; Shrestha et al., 2021) but contradicts the study of Ramandinianto et al. (2020) and Tripathi et al. (2023). The high MDR frequency is clinically significant because it may reduce therapy options and make mastitis control more challenging. It may also indicate repetitive or inappropriate antibiotic exposure in field-based mastitis treatment (Roshan et al., 2022).
Methicillin resistance in S. aureus is an important concern as MRSA infections are generally associated with limited therapeutic options and poor clinical outcomes. In our study, 83.33% S. aureus isolates were resistant to oxacillin, and all MDR strains (72.22%) were oxacillin-resistant in nature. Our result is consistent with the reports of Neelam et al. (2022). One methodological limitation of the present study is that oxacillin disk diffusion was used for phenotypic screening of methicillin resistance, whereas cefoxitin disk diffusion and MIC-based methods are preferred for more reliable MRSA confirmation in S. aureus (Fang and Yeung, 2026). Therefore, the oxacillin resistance results should be interpreted as phenotypic screening findings rather than definitive MRSA confirmation. It has been reported that the majority of MRSA isolates were resistant to at least three β-lactam antibiotics with coresistance to fluoroquinolones and aminoglycosides. In this study, 43.33% MRSA isolates showed resistance to fluoroquinolones and aminoglycosides, posing a significant challenge in treatment. According to the WHO, from One Health point of view, MRSA infections are associated with increased rates of septic shock, prolonged hospital stays, worse clinical outcomes, and higher mortality rates in humans (Pal et al., 2024). Although phenotypic oxacillin resistance was observed in the present study, only 22.22% carried the mecA gene, which is the hallmark determinant of methicillin resistance, while femA and femB are auxiliary genes associated with cell wall synthesis and may contribute to the expression of resistance (Tabassum et al., 2025). The variation in the phenotypic and genotypic detection of MRSA may be due to a divergent mecA gene, referred to as mecC. Moreover, oxacillin resistance can occur even without detectable mecA due to heterogeneous resistance expression, increased β-lactamase production, or changes in native penicillin-binding proteins. Genotypic detection may also be affected by technical factors such as primer mismatch, low gene copy number, or screening only a limited set of resistance genes (Tabassum et al., 2025). In the present study, mecA detection was low and showed only slight agreement with phenotypic oxacillin resistance (Cohen’s kappa value of 0.11). These findings suggest that oxacillin-resistant bovine mastitis isolates may carry broader antimicrobial resistance traits, relevant to veterinary treatment, but should not be interpreted as direct evidence of zoonotic or foodborne risk.
Efflux pump genes were found to be widely distributed among the isolates in this study, with all isolates carrying at least one efflux-associated gene and all MDR isolates carrying the norB gene. Among the MDR isolates, 61.53% were harboring all six efflux pump-encoded genes and were also resistant to the methicillin group of antibiotics. A review of Hassanzadeh et al. (2020) showed 75% prevalence of norA efflux pump genes on S. aureus in Asia. Suma et al. (2023) also reported the high prevalence of efflux pump genes, aligning with our result. Efflux pump genes should be regarded as potential contributors to antimicrobial resistance rather than independent predictors of the MDR phenotype.
Overall, the present study demonstrates the presence of virulence traits, biofilm-forming ability, antimicrobial resistance, and efflux pump genes among bovine mastitis-associated S. aureus. The detection of MDR traits and mecA-positivity highlights the need for continued surveillance in dairy animals within a One Health perspective. However, the findings of this study do not provide direct evidence of animal-to-human transmission, food-associated spread, or entry of these isolates into the human food chain. The global spread of MRSA is concerning because these strains resist β-lactam antibiotics. WHO reports indicate that MRSA infections are associated with 64% higher risk of death than drug-sensitive S. aureus infections (Pal et al., 2024). Further studies with larger sample sizes and a greater number of isolates are needed to establish the association between antimicrobial resistance and virulence factors in S. aureus isolated from bovine mastitis, and their public health significance. Cefoxitin disk-based screening, MIC testing, mecC gene detection, and molecular typing would further help in the confirmation of methicillin resistance more precisely and evaluate their veterinary and public health relevance.
Conclusion
Systematic records regarding the occurrence of mastitis in dairy animals, including status of infection and antibiogram results, provide useful information to the producers, farmers, and veterinarians for effective management of the disease. The current study provides a detailed picture of antibiotic resistance patterns and virulence potential, including biofilm production ability of S. aureus strains from Uttar Pradesh and West Bengal states of India. It was found that the majority of the tested isolates were resistant to multiple antimicrobial agents. Routine surveillance for antibiotic resistance pattern of isolates from bovine mastitis cases is an important measure for the detection of the emergence and spread of such resistance. Most of the isolates had biofilm-production capability, indicating that these potentially virulent bacteria could persist in the food-contact environment. From a One Health view, this can increase the chance of spread among animals, to the farm environment, and people. Hence, further study is needed to determine the association between efflux proteins and the waning sensitivity of S. aureus to different classes of antibiotics.
Authors’ Contributions
R.S.: Conducted experiments and writing of article draft; R.K.A.: Conceptualization, funding, and article draft and editing; K.B.: Conducted experiments; J.S.: Conducted experiments and article draft; K.K.: Conducted experiments; M.S.: Conducted experiments; D.R.: Conducted experiments; G.K.S.: Conducted experiments; V.S.: Conducted experiments; M.C.: Conducted experiments and data analysis; B.R.S.: Article editing and data analysis.
Data Availability
All data are presented within the article.
Footnotes
Acknowledgments
The authors duly acknowledge the Director-cum-Vice Chancellor, ICAR-IVRI, Izatnagar, and the Joint Director of Research, ICAR-IVRI, for providing the necessary infrastructural facilities to conduct the research work.
Disclosure Statement
All authors declare no conflict of interest. The authors have no relevant financial or nonfinancial interests to disclose. The funders had no role in the design of the study, in the collection, analyses, or interpretation of data; in the writing of the article; or in the decision to publish the results.
Funding Information
The financial support provided under the DST-SERB CRG project (
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.