
Abstract
This study evaluated the effects of a phytogenic-based liquid anti-mycotoxin agent (grape and olive extracts) on the redox status, health, and performance of broilers. The experimental broilers were divided into two groups: (a) group C (control group) with 22,950 broilers and (b) group T (treatment group) with 23,256 broilers, including administration of 500 mL/ton of the tested agent in drinking water on the 9th–11th, and on the 23rd–25th and 31st–33rd day. A liquid chromatography tandem mass spectrometry with a quadrupole time of flight analyzer was performed with fungal metabolites on liver samples. Thiobarbituric acid reactive substances (TBARSs), protein carbonyls (CARBs), total antioxidant capacity (TAC), total protein, and liver enzymes (alanine aminotransferase, aspartate aminotransferase, alkaline phosphatase) were determined in plasma. The relative abundance of the tenuazonic acid (TeA) was calculated for both groups, showing that the exposure of TeA was 83% lower in group T. TBARS and CARBs, as well as the levels of liver enzymes and total proteins, were significantly reduced, while TAC was increased in group T. No differences in mortality rate and a potential improvement in the growth performance of group T were noticed. In conclusion, administering a phytogenic-based liquid anti-mycotoxin agent to broiler positively affects redox status and mycotoxin management.
Keywords
Introduction
Mycotoxins are among the world’s major toxins and important risk factors for human and animal health (Schatzmayr and Streit, 2013; Pitt and Miller, 2017; Warth et al., 2016; Alshannaq and Yu, 2017; Elliott et al., 2020; Eskola et al., 2020). In addition to the direct consumption of mycotoxins from contaminated grains and other plant-derived feed materials, animal products such as meat, milk, and eggs contaminated with mycotoxins may also pose a risk to human health (Alshannaq and Yu, 2017). In poultry, ingestion of low levels of mycotoxins is more commonly associated with subclinical effects, including reduced growth performance, impaired feed efficiency, and consequent economic losses for producers (Rodrigues and Naehrer, 2012; Murugesan et al., 2015). In general, mycotoxins can affect multiple organs and systems, such as the gastrointestinal tract, liver, and kidneys, leading to health disorders, reduced performance, and increased mortality in poultry (Devegowda and Murthy, 2005; Murugesan et al., 2015).
The most frequently detected mycotoxins or metabolites are aflatoxins (AF), zearalenone (ZEN), fumonisins (FUM), deoxynivalenol (DON), ochratoxin A (OTA), and toxin T2/HT-2 (T2HT2) (Binder, 2007; Streit et al., 2013; Kolawole et al., 2020). Apart from the mycotoxins already mentioned, there are increasing concerns about emerging mycotoxins (EMs) that are neither routinely determined nor regulated (Gruber-Dorninger et al., 2017). Several in vitro and in vivo studies have reported serious health effects of these EMs on livestock, such as reproductive disorders, organ toxicity (e.g., gut, liver), and increased susceptibility to diseases and infections due to immune system dysfunction (Gerez et al., 2015; Bertero et al., 2020; Novak et al., 2021; Chiminelli et al., 2022; Garofalo et al., 2023). Tenuazonic acid (TeA) is one of the most frequently reported acutely toxic EMs among Alternaria toxins, and it is highly prevalent in cereals, which are key raw materials in poultry diets (Bottalico and Logrieco, 1998; Gross et al., 2011; Lopez et al., 2016; Babič et al., 2021). However, limited information is available on the toxic properties, toxicological mechanisms, bioavailability, and stability of Alternaria mycotoxins in the gastrointestinal tract (Babič et al., 2021). TeA toxicity has been observed in chick embryos and various animal species (e.g., pigs, mice, rabbits) due to the potential impairment of protein biosynthesis (Ostry, 2008; EFSA Panel on Contaminants in the Food Chain [CONTAM] et al., 2011; Escrivá et al., 2017). Notably, chronic exposure of broiler chickens to TeA may lead to reduced growth performance (Giambrone et al., 1978).
Due to the challenges of regulation and the detection of EMs, nutritional approaches have gained interest as preventive measures. Specifically, phytogenics, sourced from plant-based products, such as essential oils, herbs, and spices, are increasingly recognized for their antifungal, anti-inflammatory, and antioxidant properties. They also play a key role in inhibiting the growth of mycotoxin-producing fungi and reducing feed contamination (Steiner and Syed, 2015; Loi et al., 2020). They have been reported to reduce inflammation in organs commonly affected by mycotoxins, such as liver, kidneys, and gastrointestinal tract, thereby minimizing tissue damage and supporting overall organ function (Steiner and Syed, 2015; Basiouni et al., 2023). Phenolic compounds (polyphenols) are well-known for their anti-mycotoxin and antioxidant properties in animals (Abdel-Moneim et al., 2020; Papatsiros et al., 2022, 2023, 2024a, b; Papakonstantinou et al., 2023). For example, grape and olive extracts as agricultural byproducts have been studied for their potential as anti-mycotoxin agents and growth promoters (Guerra et al., 2005; Sorrenti et al., 2012). Previous studies reported anti-mycotoxin properties of olive pomace, olive leaves and branches, and olive mill wastewater against various mycotoxins, such as AF, ZEN, T-2 and OTA, due to their compounds such as hydroxytyrosol and quercetin (Bavaro et al., 2016; Abdel-Razek et al., 2017; Greco et al., 2019; Deng et al., 2019; Pereira et al., 2019; Crupi et al., 2020). Moreover, previous studies reported that grape seed extracts (Vitis vinifera), which are rich in polyphenols, exhibit protective effects against free radicals, oxidative stress, tissue damage, and inflammation. Grape pomace has been reported to possess significant biofungicidal and anti-mycotoxin properties against key mycotoxigenic species. In broilers, numerous studies have highlighted the potential of grape seed extract as an effective anti-mycotoxin, antioxidant, and growth-promoting agent, particularly against AFB1 (Baker et al., 2010; Lawal and Bolu, 2014; Ali Rajput et al., 2017; EFSA Panel on Contaminants in the Food Chain [CONTAM] et al., 2018; Almusawi et al., 2019; Khalil et al., 2022; Hassan et al., 2023).
Mycotoxin exposure presents significant challenges to the poultry industry, causing metabolic, physiological, and immunological disruptions, that ultimately reduce the productivity (Bryden, 2012). Several studies reported the positive effect of anti-mycotoxin agents in preventing losses associated with mycotoxicosis in poultry (Tsiouris et al., 2021; Raj et al., 2023; Stefanović et al., 2023; Jannah et al., 2024). However, a key limitation in controlling EMs with binders underscores for new mitigation strategies, including the development of novel anti-mycotoxin agents for this important group of mycotoxins (Siri-anusornsak et al., 2023). This study aimed to investigate the effects of a liquid anti-mycotoxin agent based on grape and olive extracts on the redox status, health, and performance of broiler chickens. The innovation of our study lies in the application of an anti-mycotoxin phytogenic formula under commercial broiler production conditions, with emphasis on oxidative stress, liver function, and hepatic exposure of TeA. We hypothesized that the application of the phytogenic product would reduce oxidative stress and hepatic exposure of TeA. Specifically, our research objectives were (i) quantification of mycotoxin contamination in feed and liver samples; (ii) investigation of the effects of the product on plasma redox biomarkers and liver function; and (iii) assessment of mortality rates and body weight measurements in broiler flocks.
Materials and Methods
Description of the trial farm/management and diets
The present study was carried out on a broiler farm in Greece, which featured two poultry houses of equal size. Broiler chickens (Ross 308) were placed in these houses and raised under standard field conditions until slaughter at 42 d of age. Specifically, 23,256 broiler chickens were housed in the treatment house, while 22,950 chickens were placed in the control house.
The management program followed official guidelines for broiler chickens to ensure proper health and welfare (Ross Broiler Management Handbook, 2018). Wood shavings and sawdust were used as bedding materials in cleaned and disinfected houses. The basal diet was formulated with maize, wheat, and soybean meal, in line with the official genetic line recommendations (Ross Broiler Management Handbook, 2018). The data are available in Supplementary Table S1. The chickens were fed isoenergetic and iso-protein diets in mash form, including pre-starter (days 0–14), starter (days 15–24), grower (days 25–35), and finisher (days 36–42) phases. Feed and water were provided ad libitum. The lighting program began at 7 d of age, with 4 h of darkness and 20 h of light each day. In accordance with the local vaccination schedule, the birds were vaccinated against infectious bursal disease, infectious bronchitis, and Newcastle disease at the hatchery.
A double-blind design was employed, ensuring that the personnel involved in the clinical evaluation of the trial animals were unaware of the group assignments or treatments, as were those performing laboratory analyses. All experimental animals were monitored daily by an experienced veterinarian for adverse clinical signs (diarrhea, respiratory signs, loss of appetite, morbidity, etc.).
Experimental material
A novel mycotoxin neutralizing agent in liquid form (Biōnte® Quimitōx® Livōx®, developed by Biōnte Nutrition S.L., Reus, Spain) were tested. It is composed of grape pomace extract (skin and seed, V. vinifera), rich in polyphenolic compounds (proanthocyanidins, anthocyanins, procyanidins, catechin, ferulic acid, quercetin, resveratrol, and syringic acid, among other bioactive compounds and derivatives) and olive pomace extract (O. europaea), rich in hydroxytyrosol, tyrosol, and other polyphenolic derivatives. In addition, it contains citric acid, sorbic acid, water, and sorbitol of high purity as chemical components. The selected phytogenic ingredients have demonstrated antioxidant, anti-inflammatory, and antimicrobial properties that contribute to mitigating the negative effects of mycotoxins (Arora et al., 2025).
Experimental design
This study included two groups: (a) group C (control group), including 22,950 broiler chickens, which were placed in the control house, and (b) group T (treatment group), including 23,256 broiler chickens, which were placed in the treatment house of farm, in where Biōnte® Quimitōx® Livōx® was administered in 500 mL/ton of clean drinking water consecutively for 3 days, on days 9th, 10th, 11th, then at 23rd, 24th, 25th day and finally at days 31st, 32nd, 33rd day. A flowchart of the trial is presented in Figure 1.
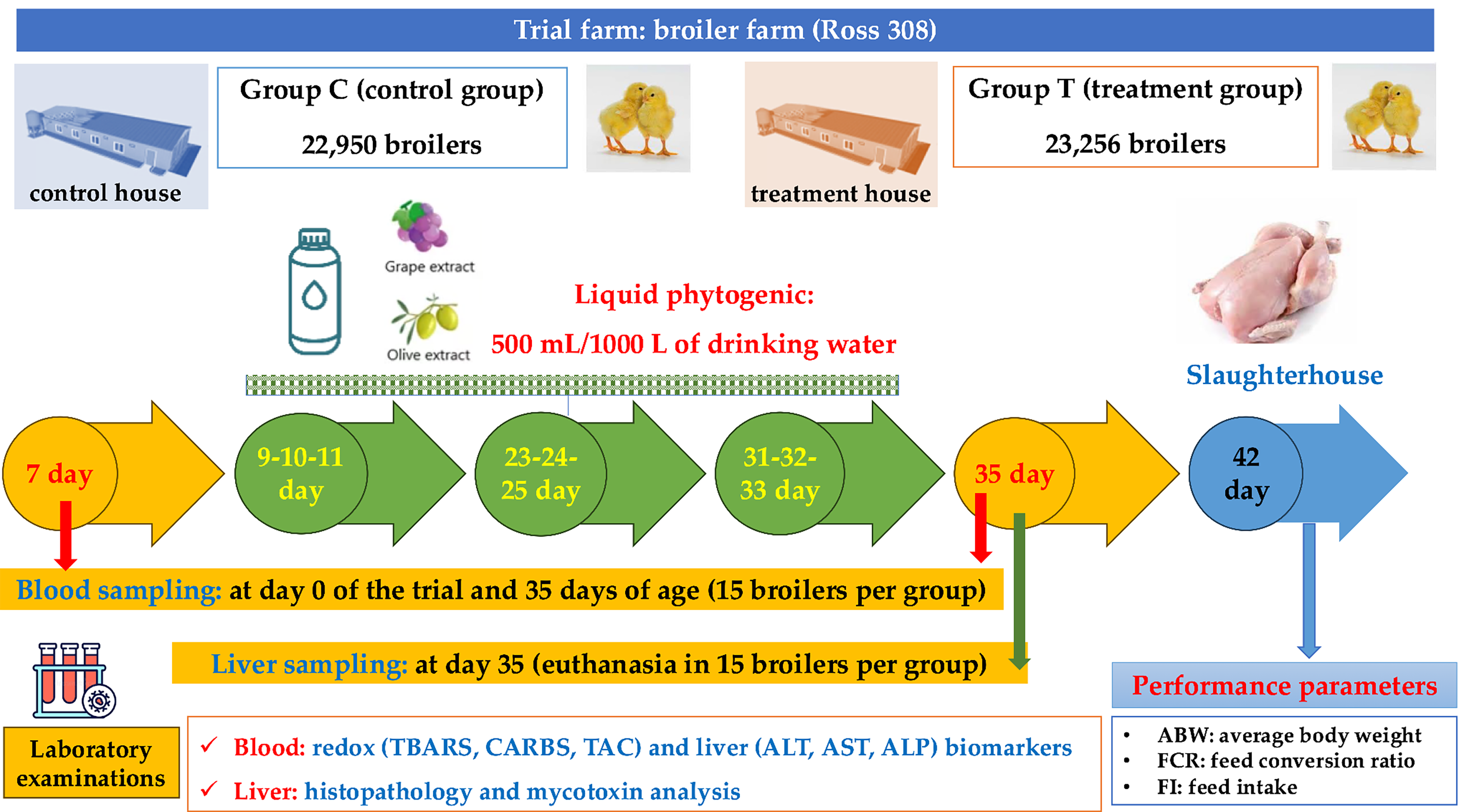
A flowchart of the trial (groups, study design, sampling, laboratory examinations).
Quantification of mycotoxins in feed
During the study, two feed samples weighing 500 g of the final feed were collected every 15 d five times (a total of 10 feed samples). The samples were analyzed and quantified for mycotoxin and their metabolites in an officially accredited laboratory (Biōnte Nutrition S.L, Reus, Spain).
Samples were stored at −20°C upon arrival to preserve their status until the analysis. Feed samples were prepared for analysis. Mycotoxins were present in feed samples using lateral flow kits based on immunochromatography. Thus, feed samples were analyzed to determine DON, FUB, AF-B1, OTA, ZEN and T2/HT2. Feed samples were analyzed using lateral flow strip kits supplied by ProGnosis Biotech (Larissa, Greece). Briefly, previously homogenized feed samples were ground until a fine powder was obtained. Five grams of the obtained powder was weighed into a 50 mL falcon tube, and 15 mL of the extraction solution (included in the kit) was added. The mixture was vortex shaken for 2 min. An aliquot (1 mL) of the supernatant was transferred to a 1.5 mL Eppendorf and centrifuged at 12,500 rpm for 2 min. A volume of 100 µL of the supernatant was added to a dilution tube (included in the kit) and further mixed. Finally, 100 µL of the diluted extract was added to a microwell containing the lyophilized gold particles and homogenized using the micropipette until completely diluted. Once completely diluted, the strip was introduced 3 min after the introduction of the strip, it was retired from the microwell, and the strip’s cotton pad was peeled before its quantification using a 3PR-Mini instrument from ProGnosis. The quantification was performed using the S-Flow software from the same company.
Blood and tissue sampling
On days 7 and 35 of the trial, blood samples were collected from 15 randomly selected birds per group in EDTA tubes. Blood designated for obtaining plasma (EDTA) was kept on ice until centrifugation (3000 × g, 4°C, 15 min) and then stored at −20°C for analyses.
On the 35th day, 15 randomly selected birds from each group after blood sampling were euthanized. From each bird, one entire liver lobe was collected and directly collocated in a plastic bag and stored at −20. In addition, from the second liver lobe, around 3 cm long segments were collected and placed in 10% neutral buffered formalin as described by Bakeer et al. (2013).
Blood examinations for redox and liver biomarkers
The assessment of thiobarbituric acid (TBA) reactive substance (TBARS) and protein carbonyl (CARB) levels in plasma, utilized as redox biomarkers, was conducted based on the methodology of Gerasopoulos et al. (2015). TBARS values were determined using a modified version of the analysis by Keles et al. (2001), where TBARS concentration (μmol/L plasma) was calculated using the molar extinction coefficient of malondialdehyde (MDA). MDA reacts with TBA in a 1:2 ratio to form an adduct measurable spectrophotometrically, with an MDA molar extinction coefficient of 155 × 103 M−1 cm−1. For CARB quantification (nmol/mg protein), the method outlined by Patsoukis et al. (2004) was applied, relying on the molar extinction coefficient of 2,4-dinitrophenylhydrazine (22 × 103 M−1 cm−1). In addition, total antioxidant capacity (TAC) in plasma (mmol DPPH/L plasma) was evaluated using the approach described by Janaszewska and Bartosz (2002).
Plasma total protein, alanine aminotransferase (ALT), aspartate aminotransferase (AST), and alkaline phosphatase (ALP) were determined spectrophotometrically using available commercial kits (Randox B, I234, and Foster City, CA, USA; catalog numbers: TP 3869, AL 3801, AS 3804, AP 3802 for total protein, ALT, AST, and ALP, respectively).
Liver examinations: Histopathological examination and mycotoxin analysis
The collected liver samples were fixed in 10% buffered formalin for 48–72 h for histopathological examination. Slides with 3- to 5-μm-thick sections of formalin fixed paraffin–embedded specimens (Paraplast Plus®, Kendall, UK) were then prepared and routinely stained with hematoxylin and eosin (Feldman and Wolfe, 2014).
The collected liver samples were analyzed using ultra high-performance liquid chromatography tandem mass spectrometry with a quadrupole time of flight analyzer (HPLC-QTOF). Liver samples were extracted. The final extracts were injected into the HPLC-QTOF instrument, and a screening method was conducted with over 400 registered fungi metabolites.
Liver samples were analyzed by Microlab Biotech S.L. (Valencia, Spain). Briefly, samples were homogenized using a Stomacher homogenizer for 1 minute. Once homogenized, 3 g of sample was added to a 50 mL falcon tube with 3 mL of ultrapure water, 3 mL of acetonitrile, and 1 g of sodium chloride. The tubes were mixed using an ultraturrax for 2 min and then centrifuged for 5 min at 4500 rpm. The supernatant was filtrated using a 0.45 µm filter, and 2 mL of the filtrated extract was added to a 15 mL falcon tube to perform a dispersive liquid–liquid microextraction. A total of 6 mL of ultrapure water was added to the tube and 600 µL of chloroform was included as extraction solvent. The mixture was centrifuged for 3 min at 2500 rpm, and the organic phase was further evaporated to dryness using a gentle nitrogen stream. Finally, the extracts were reconstituted with 500 µL of a methanol:water (50:50, v/v) mixture, filtrated with a 0.2 µm syringe filter, and injected in the UHPLC-MS/TOF system. The detection was performed using an untargeted suspect screening approach with a database containing over 400 registered fungi metabolites. Calculation of the relative abundance of the TeA in the samples was performed based on the peak of chromatograms, using the database, and the software of the laboratory.
Mortality and performance parameters
To assess potential growth differences between the two groups, the average body weight (ABW) was recorded when the broiler chickens were loaded at slaughterhouse at 42 d of age. The feed intake (FI) was recorded, and the feed conversion ratio was calculated by dividing the total feed intake by the total body weight gain. Mortality was recorded daily throughout the entire trial period. Mortality data were collected on a flock basis for each group.
Statistical analysis
All the experiments were performed in triplicate to account for experimental variability and ensure the reliability of the results. Obtained results are expressed as means ± standard deviation (±SD). The assumptions of normality (Kolmogorov–Smirnov and Shapiro–Wilk tests) and homogeneity of variance (Levene’s test) were verified prior to parametric testing. The differences between the investigated groups, at days 7 and 35, were assessed by performing an independent two-tailed t-test, at a significance level of 0.05. A G*Power analysis was performed to estimate study power and determine the appropriate sample size. A group size of 15 animals was deemed sufficient, based on a range of effect sizes, to achieve a study power of 95% (Kang, 2021). Specifically, the effect size was evaluated based on the expected mean and SD of the oxidative stress biomarkers between groups. By examining a range of effect sizes (2–3.5) for a two-tailed independent group comparison and assuming type I and type II errors rates of 0.05, we concluded that a group size of 15 animals would yield a study power greater than 95%. All statistical analyses were performed using the R programming language (R Core Team, 2023).
Results
Quantification of mycotoxins in the feed and liver samples
Laboratory testing of the five feed samples showed that the individual risk associated with mycotoxins is low, as the levels were not elevated (Table 1).
Mycotoxins Analysis in Feed During the Trial
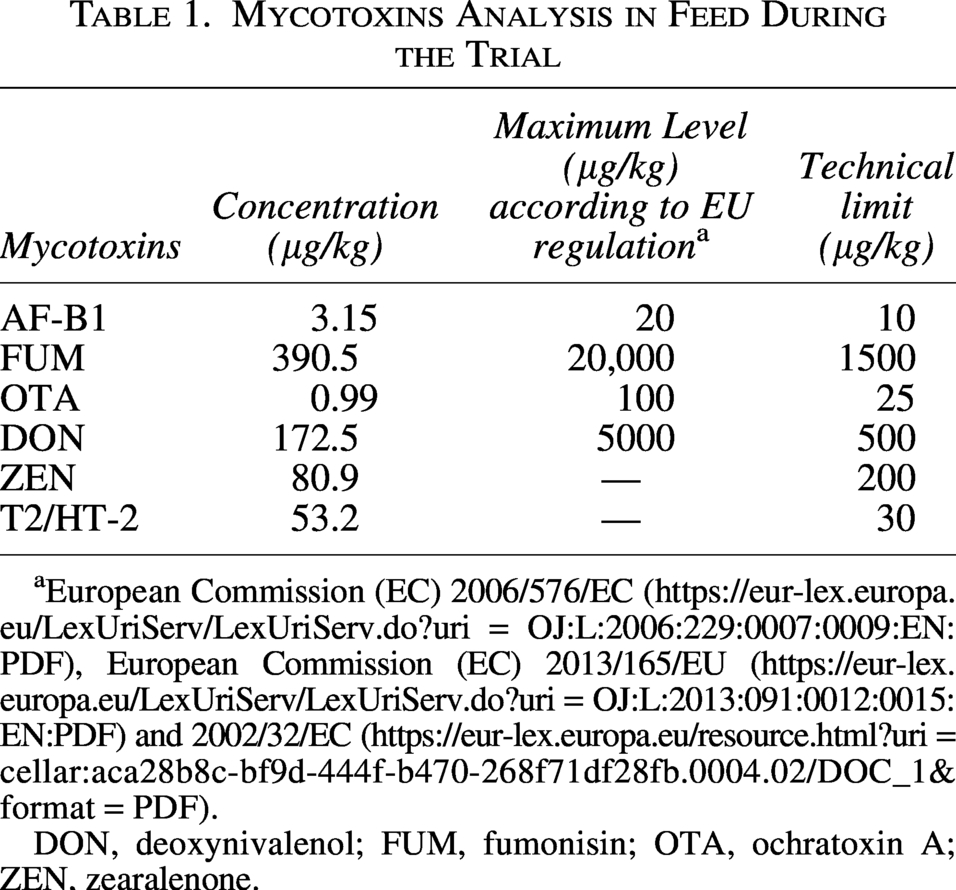
European Commission (EC) 2006/576/EC (https://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri = OJ:L:2006:229:0007:0009:EN:PDF), European Commission (EC) 2013/165/EU (https://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri = OJ:L:2013:091:0012:0015:EN:PDF) and 2002/32/EC (https://eur-lex.europa.eu/resource.html?uri = cellar:aca28b8c-bf9d-444f-b470-268f71df28fb.0004.02/DOC_1&format = PDF).
DON, deoxynivalenol; FUM, fumonisin; OTA, ochratoxin A; ZEN, zearalenone.
Blood examinations for redox biomarkers
Table 2 presents the results concerning the parameters responsible for broilers’ antioxidant status. In both groups, plasma TBARS and CARB declined significantly (p < 0.05) between days 7 and 35, while TAC increased significantly (p < 0.05) over the same period, indicating a general maturation-related improvement in antioxidant capacity (exact values are reported in Table 2). Critically, however, the magnitude of these changes differed substantially between the groups: the phytogenic treatment (group T) produced markedly greater reductions in TBARS and CARBs, and a greater increase in TAC, than the untreated control (group C) by day 35, indicating a treatment-specific enhancement of antioxidant defenses beyond age-related effects. As shown in Table 2, at the beginning of the experiment (day 7), the content of TBARS, CARB, and TAC remained almost intact between groups C and T, confirming that the two groups were comparable at baseline. On the contrary, by day 35, both TBARS and CARBs were significantly lower (p < 0.05) in group T than in group C, while TAC was significantly higher (p < 0.05) in group T. These between-group differences are the most clinically relevant finding of the study, as they demonstrate that the phytogenic anti-mycotoxin agent conferred a measurable antioxidant benefit above and beyond spontaneous age-related changes.
Comparison of the Oxidative Stress Biomarkers and Plasma Biochemical Parameters in Broilers on Day 7 and Day 35 of the Trial
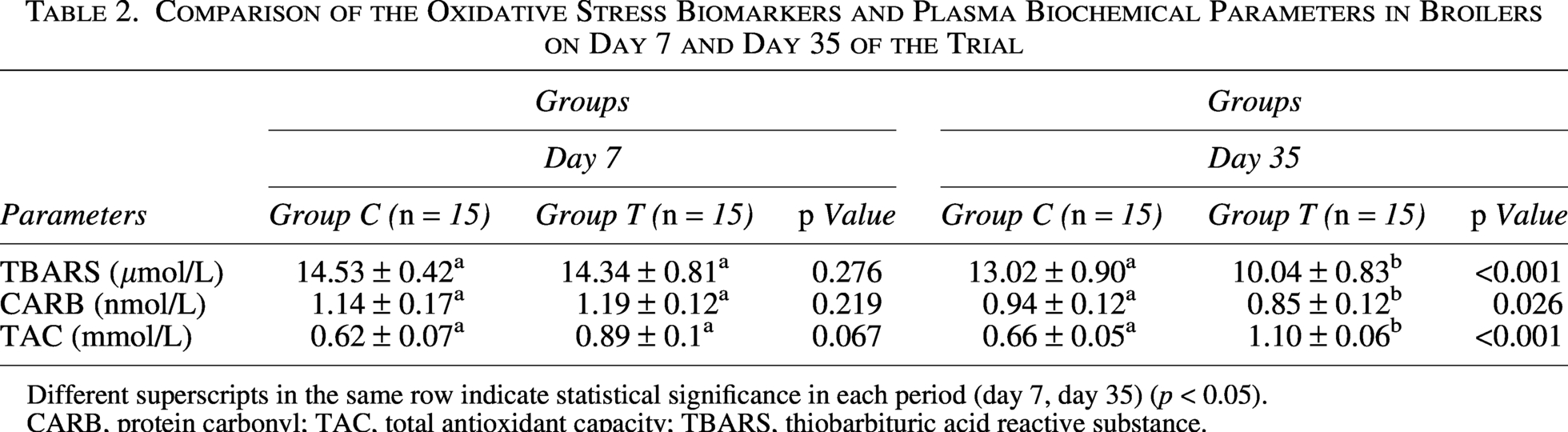
Different superscripts in the same row indicate statistical significance in each period (day 7, day 35) (p < 0.05).
CARB, protein carbonyl; TAC, total antioxidant capacity; TBARS, thiobarbituric acid reactive substance.
The total protein content was almost identical for groups C and T at the beginning of the experiment (Table 3). The total protein content on day 35 was increased for both experimental groups, but only the increase for group T was significantly (p < 0.05) increased. The parameters of ALT, AST and ALP presented a similar pattern Table 3. Specifically, ALT, AST, and ALP activities were comparable between groups C and T at day 7. On day 35, ALT, AST, and ALP were significantly (p < 0.05) reduced in group T, but not group C.
Evaluation of Plasma Biochemical Parameters in Broilers on Day 7 and Day 35 of the Trial
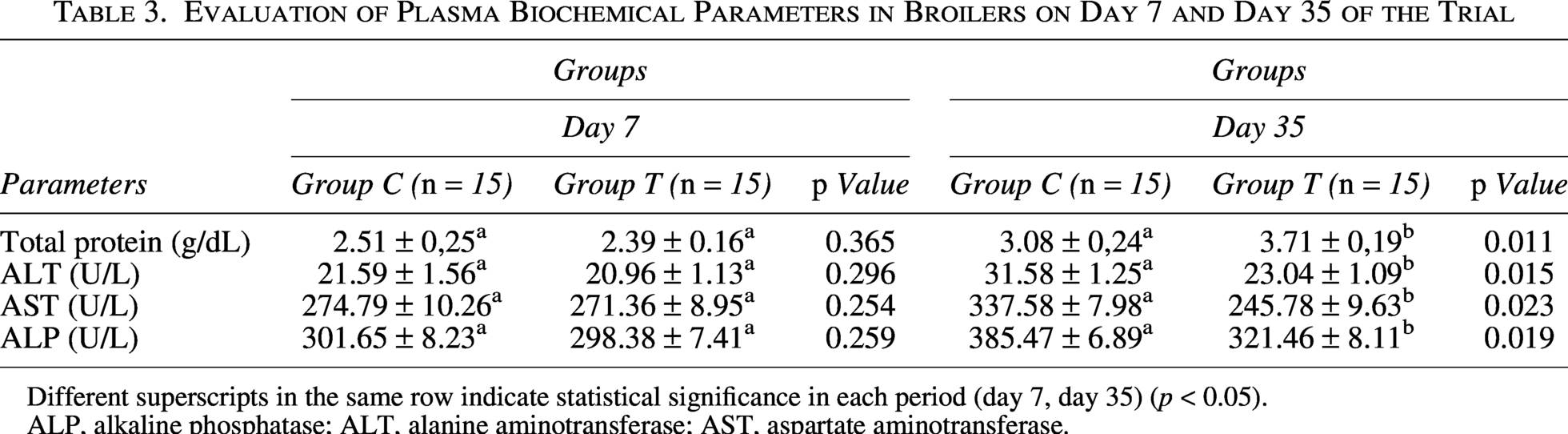
Different superscripts in the same row indicate statistical significance in each period (day 7, day 35) (p < 0.05).
ALP, alkaline phosphatase; ALT, alanine aminotransferase; AST, aspartate aminotransferase.
The temporal changes in both oxidative stress biomarkers and plasma biochemical parameters in broilers on day 7 and day 35 (Fig. 2), comparing control (Group C) and treatment (Group T) groups, are shown in Figure 2. The trajectories illustrate that, while both groups followed broadly similar trends across the study period, group T diverged markedly from group C by day 35 in all measured biomarkers, with convergent evidence of reduced oxidative damage (lower TBARS and CARBs) and improved systemic antioxidant capacity (higher TAC), alongside lower circulating liver enzyme activities (ALT, AST, and ALP), collectively suggesting a hepatoprotective effect of the phytogenic.
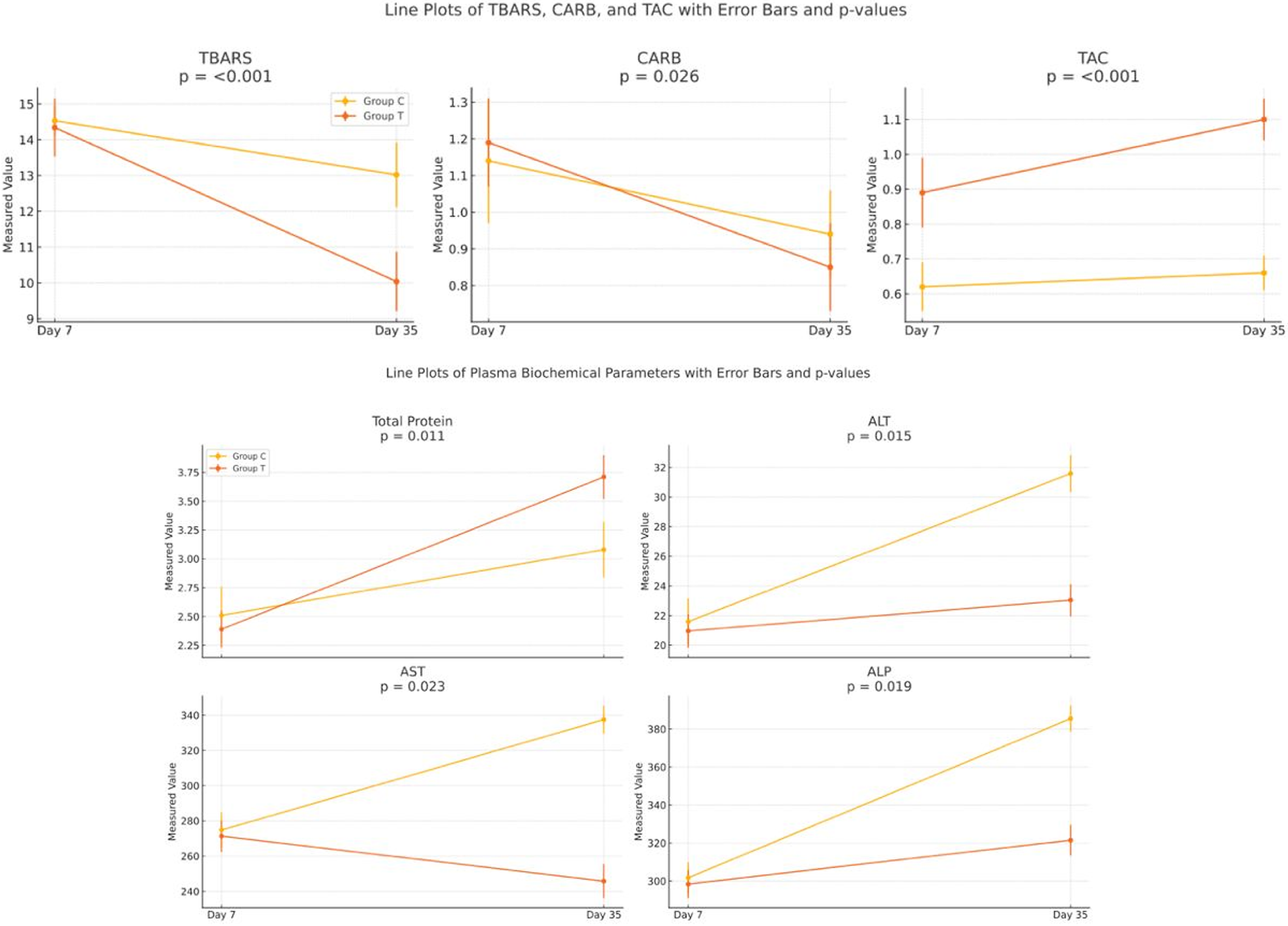
Comparative line plots of oxidative stress biomarkers and plasma biochemical parameters in broilers on day 7 and day 35.
Liver examinations: Histopathological examination and mycotoxin analysis
According to histopathological examination of liver samples, gross examination of the dissected liver lobes showed discoloration in a few cases of both groups (Fig. 3). Histopathologic evaluation of the liver samples revealed sporadic changes such as congestion (3/15 of group C and 2/15 of group T) (Fig. 3a, broiler No6 from control group), vacuolar degeneration of hepatocytes (2/15 of group C and 3/15 of group T) (Fig. 3b, broiler No6 from control group), hydropic degeneration (4/15 of group C and 4/15 of group T), and foci of inflammatory infiltration by heterophils (2/15 of group C and/1/15 of group T) (Fig. 3c, broiler No6 from control group) or lymphocytes and plasma cells (Fig. 3d, broiler No6 from control group). The lesions did not differ between group C and group T.

Liver, hematoxylin–eosin stain
According to the quantification of mycotoxins in the liver samples, the obtained values indicate that the individual risk of mycotoxins was low in our study, as the levels are not elevated. Figure 4 depicts the relative abundance of the TeA in the liver sample A (group C), and sample B (group T). However, the relative abundance is not considered a quantification of the compound. Notably, the most important finding of this study was that broilers receiving the phytogenic anti-mycotoxin agent (group T) showed an 83% reduction in the relative hepatic abundance of TeA compared with untreated controls (group C)—a substantially large effect size that directly demonstrates the capacity of the tested agent to reduce the exposure of this EM at the target organ level (Table 4).
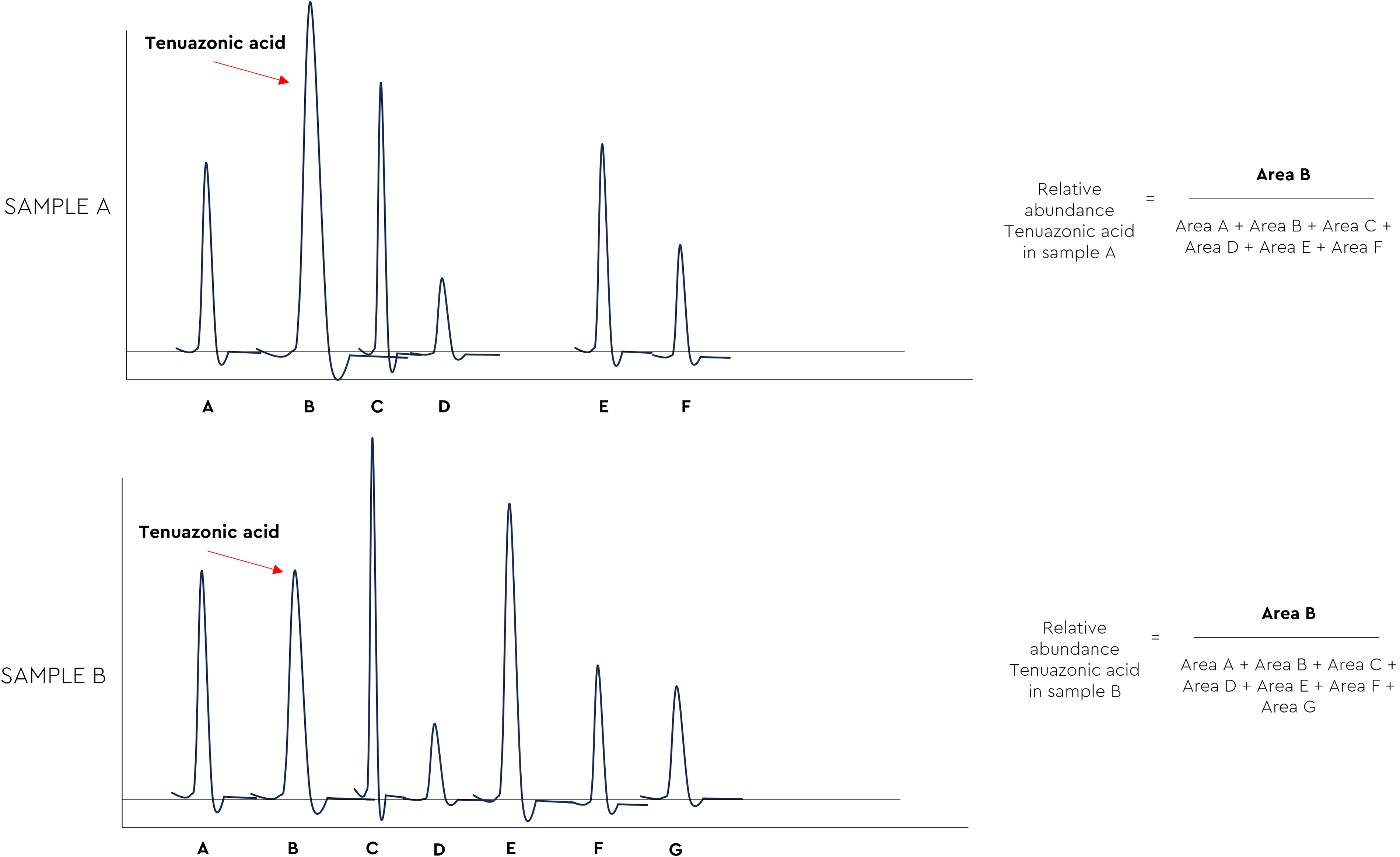
Two chromatograms for the calculation of the relative abundance of the TeA in the samples of both sample A (group C), and sample B (group T). TeA, tenuazonic acid.
Relative Abundance of Fungal Metabolites in Liver Samples

TeA, tenuazonic acid.
Mortality and performance parameters
The mortality rate (%) and performance parameters during the total trial period are presented in Table 5. No significant differences were noticed between groups in mortality rate, while a potential improvement in the growth performance of group T was noticed.
Mortality and Performance Parameters in Control and Treatment Groups During the Total Trial Period
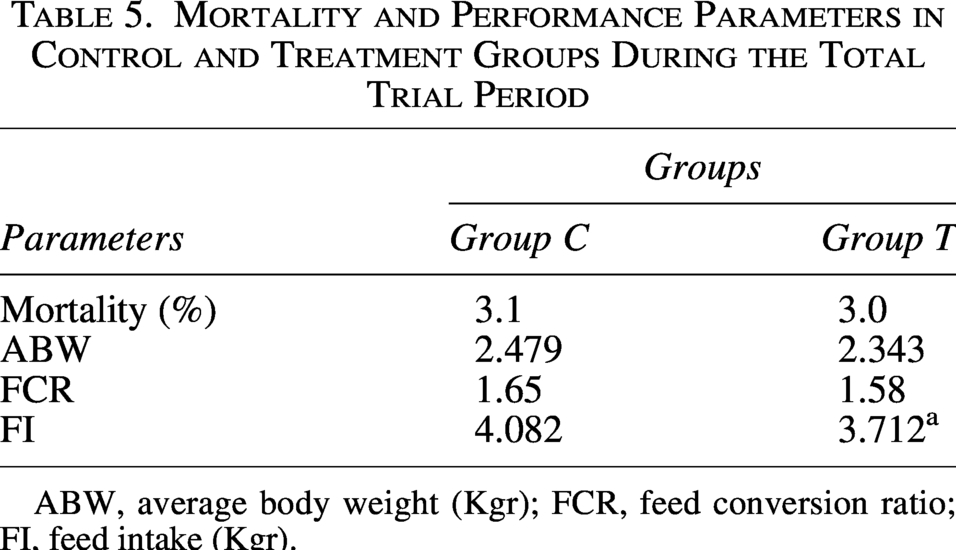
ABW, average body weight (Kgr); FCR, feed conversion ratio; FI, feed intake (Kgr).
Discussion
The contamination of feed ingredients with mycotoxins continues to be a serious global issue, as the Food and Agriculture Organization reported about 25% of worldwide mycotoxin contamination of agricultural products (Eskola et al., 2020). In our study, the levels of mycotoxins detected in the feed during the trial indicated a low individual mycotoxin risk. However, as described in detail in the literature, low levels of mycotoxins can cause subclinical symptoms, such as oxidative stress, metabolic damage, and immunotoxicity (Mavrommatis et al., 2021). When the liver samples were analyzed, the relative abundance of the EM TeA was found in both the control and treatment groups, indicating that the broiler chickens were exposed to this mycotoxin during our experiment. TeA is widely used in food and feed and, for example, 65% of the samples analyzed between 2010 and 2012 were contaminated with TeA (Gross et al., 2011; Asam et al., 2013; Streit et al., 2013; Fraeyman et al., 2015). In our trial, the group receiving the tested mycotoxin neutralizer containing phytogenics (grape seed and olive extracts) showed an 83% decrease in the exposure of TeA in broiler chickens. Although EMs such as TeA are frequently detected, there is no legislation or guidance and data on in vivo toxicity are limited. The complete oral bioavailability of TeA has been reported in both pigs and broiler chickens (EFSA, 2013). The oral bioavailability of TeA is also likely to be high in humans, as human exposure to TeA through the consumption of animal products has been reported (Ostry, 2008). Our results could form the basis for further studies as required for the update of the risk assessment of most EMs by the European Food Safety Authority (Asam and Rychlik, 2013; Fraeyman et al., 2017). The limitations of our study include the administration of a single dose of the tested anti-mycotoxin agent under field conditions with low-level contaminated feed. During the trial period, no health issues related to mortality were observed in broilers. However, further research with varying dosages of the tested agent, under both experimental and field conditions, is necessary to gain a deeper understanding of its impact on exposure of TeA in broiler chickens, as well as its effects on mortality and performance parameters.
Given the problem of mycotoxins in animal production, it is known that the most used method is the addition of adsorbents to reduce the uptake of toxins and their distribution in the blood and target organs (Vázquez-Durán et al., 2022). Globally, phytogenic feed additives rich in polyphenolic compounds have drawn the attention of producers to promote sustainable practices. Their secondary metabolite content, including polyphenols, exerts a wide range of beneficial effects due to their immunomodulatory, anti-inflammatory, antioxidant, antimicrobial, and detoxifying effects (Abdel-Moneim et al., 2020; Papatsiros et al., 2023, 2024a; Papakonstantinou et al., 2023). The mechanisms underlying the antioxidant effects of polyphenols involve several synergistic actions that explain the observed decreases in TBARS and CARB values, as well as the increase in TAC in the present study. First, the hydroxyl groups of polyphenols serve as hydrogen donors, neutralizing reactive oxygen species and interrupting the lipid peroxidation chain reaction and MDA production, which is the main substance measured in the TBARS test (Scalbert et al., 2005). Second, polyphenols, particularly proanthocyanidins that predominate in grape seeds, can chelate metal ions such as Fe2+ and Cu2+, which are responsible for generating Fenton reactions that produce hydroxyl radicals (Heim et al., 2002). Finally, and most importantly for long-term antioxidant effects, polyphenols induce activation of the Nrf2/Keap1 transcription system, which increases the expression of antioxidant enzymes such as SOD, CAT, and GPx, the main contributors to overall TAC levels (Surai, 2014; Sun et al., 2020). The simultaneous decrease in CARB concentration is also explained by the reduced activity of hydroxyl radicals, as CARBs are produced primarily through metal-assisted oxidation of amino acid chains in proteins. Therefore, the biochemical mechanism underlying these changes in redox biomarkers is clear for group T in the present study. According to the literature, olive oil or its derivatives have been used to improve performance parameters, feed efficiency, the heat resistance by combating heat stress and the health status of broilers, and as a growth promoter or protein source (Mujahid et al., 2009; Mahmoud et al., 2013; Berrougui et al., 2015; Gerasopoulos et al., 2015; Seifi et al., 2018; Agah et al., 2019; Saleh et al., 2020; Ibrahim et al., 2021; Vázquez-Durán et al., 2022). Grape seed extracts (V. vinifera) are a byproduct of wine and grape juice processing and contain polyphenols with high antioxidant capacity (Santos-Buelga and Scalbert, 2000; Shi et al., 2003). It is also a known potent antioxidant that contributes to disease prevention and protection against oxidative stress, tissue damage, and inflammation due to its ability to absorb oxygen radicals and its anti-inflammatory effects (Ouédraogo et al., 2011; Sayin et al., 2014). Previous studies with its use as a feed additive for chickens have shown antioxidant, hepatoprotective, and immune-boosting properties as well as positive effects on chickens with coccidiosis (Wang et al., 2008; Farahat et al., 2017; Rajput et al., 2017). The findings of our study are in agreement with these aforementioned studies, as the tested phytogenic agents (grape extract: skin and seed, V. vinifera, and olive extract: pomace, O. europaea) in our trial have shown antioxidant properties based on the significant mitigation of redox biomarkers (TAC, TBARS, and CARBs) in the treatment group, helping to reduce the negative effects of mycotoxins. In addition, grape and olive extracts have been shown to exhibit effective mycotoxin-binding and adsorption in vitro properties against various mycotoxins, such as AF-B1, ZEN, and OTA. These extracts help to reduce the harmful effects of mycotoxins, likely due to the presence of micronized fibers and phenolic compounds (EFSA Panel on Contaminants in the Food Chain [CONTAM] et al., 2011; Avantaggiato et al., 2014; Gambacorta et al., 2016; Abdel-Wahhab et al., 2017; Taranu et al., 2019; Fernández-Poyatos et al., 2019). Based on the findings of the previously published studies, our results about the possible positive antioxidant and antitoxins properties against TeA in poultry could be associated with the administration of the tested phytogenic based anti-mycotoxin agent. Further research is needed to identify which specific phenol or combination of phenols is responsible for the reported significant reduction in the exposure of TeA in broiler chickens, under both experimental and field conditions, considering varying levels of feed contamination. Our previous studies in sows and weaned piglets, using anti-mycotoxin agents containing different phytogenics enriched in polyphenols compared with the present study, showed positive effects not only on oxidative redox status but also on health and growth performance (Papatsiros et al., 2023, 2024a). In contrast, the present study observed a potential improvement in the performance parameters of the treatment group. The biological basis for the performance advantage under mycotoxin exposure involves several factors. Mycotoxin-induced oxidative stress imposes an energetic cost on the organism, as the upregulation of endogenous antioxidant defenses (increased synthesis of SOD, CAT, GPx, and glutathione) diverts amino acid precursors and metabolic energy away from growth and protein accretion (Surai and Fisinin, 2015; Grenier and Applegate, 2013; Wu et al., 2004). By reducing the oxidative load through polyphenol supplementation, the plant-derived product may have partially alleviated this metabolic burden, thereby increasing the availability of nutrients for productive activities (Surai, 2014). In addition, the hepatoprotective effect (evidenced by decreased ALT, AST, and ALP activity) ensures that the liver remains capable of its metabolic, gluconeogenetic, and protein synthesis functions—essential biological mechanisms for maintaining feed utilization efficiency in broilers (Davis et al., 1998; Leeson and Summers, 2009). Although there were no statistically significant differences in performance metrics in the present study, due to the relatively moderate levels of mycotoxin exposure and insufficient statistical power at the flock level, the biological plausibility of the observed trends is supported by the proposed mechanism described above. However, further studies under both experimental and field conditions, using various levels of contaminated feed, larger numbers of broilers and multiple replicates, along with different dosages of the tested agent, are needed to fully access the effects of the tested product.
After crossing the gastrointestinal barrier, mycotoxins typically target the liver as their next site of action. However, Hessel-Pras et al. (2019) highlight the limited understanding of the potentially harmful effects of Alternaria mycotoxins on liver cells. While studies have demonstrated that TeA is almost entirely absorbed in the intestines of pigs and broiler chickens (Fraeyman et al., 2017), no information is currently available on the cellular metabolism of TeA. Certain mycotoxins affect various organs or tissues (e.g., liver, kidneys, gastrointestinal tract, reproductive and urogenital systems), leading in extreme cases to reduced productivity and mortality in livestock and poultry (Devegowda and Murthy, 2005; Murugesan et al., 2015; Papatsiros et al., 2021). However, in our study, no differences in mortality rates were observed between the control and treatment groups. Chronic exposure to TeA reduces animal performance, as a reduction in weight gain was observed in a previous study where broilers were exposed to TeA (Lopez et al., 2016). In the treatment group of our study, the reduction of the exposure of TeA in the liver of broiler chickens by 83% was associated with a potential improvement of performance parameters.
While the exact mechanism underlying the decrease in TeA concentration in the liver of broiler chickens remains unclear, several possible mechanisms may be involved based on the chemical properties of TeA and the composition of the test substance. The TeA molecule contains a tetramic acid group that can form hydrogen and hydrophobic bonds with dietary fiber. Adsorption of TeA molecules onto the surface of micronized lignocellulose fibers in grape and olive pomaces in the gut may reduce the fraction of the substance available for intestinal absorption (Avantaggiato et al., 2014; Gambacorta et al., 2016). In addition, TeA molecules may form complexes with polyphenols through π–stacking interactions between aromatic groups, which limits their solubility and permeability in the intestine (Fernández-Poyatos et al., 2019). Finally, activation of Nrf2 transcription factors by polyphenols, resulting in increased activity of phase II detoxification enzymes, can promote faster conjugation and biliary excretion of TeA from the body (Surai, 2014). Grape seed (V. vinifera) extracts have been shown to provide significant protection against free radicals, lipid peroxidation induced by free radicals, and DNA damage (Bagchi et al., 2000; Xu et al., 2015). It is well-established that these extracts support the functions of vital organs by boosting the antioxidant system and preventing liver damage caused by carbon tetrachloride and ischemia/reperfusion (Dai et al., 2014; Xu et al., 2015).
ALT, AST, and ALP are leaking into the circulation from hepatocellular cells and are the determining enzymes of hepatocellular damage. They are key indicators of liver health, as they play crucial roles in protein and amino acid metabolism within liver cells (Zhang, 2011). Increased ALT and/or AST activities are indicative of hepatocellular injury (Yang et al., 2016). It is also known that mycotoxins (such as AF, FUM, ZEN, and T2HT2) led to aggravated liver dysfunction in broilers, increased liver enzymes (ALT, AST, and ALP) activities, and decreased total protein levels in serum (Fernandez et al., 1994; Tessari et al., 2010; Yang et al., 2016; Rajput et al., 2017; Chen et al., 2019; Ates and Ortatatli, 2021; Zou et al., 2023). The significantly lower plasma ALT, AST, and ALP activities, along with higher total protein levels observed in group T at day 35, are best explained mechanistically as a result of reduced hepatocyte oxidative injury, rather than a primary effect on enzyme synthesis or activity. Hepatocellular membrane integrity depends critically on the lipid bilayer remaining free from peroxidation; when lipid peroxides accumulate—as indicated by elevated TBARS—the resulting aldehydic products (particularly 4-hydroxynonenal and MDA) form adducts with membrane phospholipids and proteins, increasing membrane permeability and allowing cytosolic enzymes to leak into the bloodstream (Comporti, 1998; Esterbauer et al., 1991). The polyphenols in the grape and olive extracts tested in this study, by quenching lipid peroxidation (reduced TBARS) and protein oxidation (reduced CARBs), are proposed to have maintained hepatocyte membrane integrity, thereby reducing the passive efflux of ALT, AST, and ALP into the circulation. The concurrent increase in total plasma protein in group T supports this interpretation, as oxidative damage to hepatocytes also impairs protein synthesis capacity. Importantly, this mechanistic sequence, whereby polyphenol-mediated ROS scavenging leads to reduced lipid peroxidation, which in turn preserves hepatocyte membrane integrity and results in lower circulating transaminase activities, is directly supported by the co-occurrence of these biomarker changes in this study. The beneficial effects of grape and olive extracts on liver function and liver enzyme activities in broilers have also been documented in previous studies (Agah et al., 2019; Almusawi et al., 2019; Kumanda et al., 2019; Tavakolinasab et al., 2020; Erinle et al., 2022; Almuhayawi et al., 2023). In contrast, our findings differ from those of recent studies, which reported no significant effect on liver enzyme activity in broilers fed diets containing olive leaves and olive pulp (Rezar et al., 2025) or grape seed extracts (Rajput et al., 2017). The discrepancies between the positive results and those that did not show anything significant can perhaps be explained by the following factors. The first factor is the polyphenol concentration and bioavailability of the particular botanical source used (olive leaves, grape seeds, and pomace extracts contain different quantities of proanthocyanidins and total phenols). The second variable may be the extent of oxidative stress experienced by the sheep flock (significant improvements would only be observed in flocks experiencing considerable oxidative stress, either due to exposure to mycotoxins or other factors). Third, we must consider the method of supplement administration, such as oral dosing, compared with its inclusion in the diet. However, we have taken under consideration that an increase in serum of ALT and AST, whereas a decline in serum of ALP levels occurs with increasing age of broilers (Meluzzi et al., 1992). Our study’s results suggest that the tested anti-mycotoxin agent based on olive and grape extracts, has notable antioxidant properties. The identified gaps in our study, particularly regarding the use of a single dose of tested agent under low-level contamination field conditions, emphasize the need for further research. Some limitations of the current research should be noted. First, only a single treatment protocol was investigated in the field under conditions where mycotoxins naturally occur at lower concentrations, which restricts its applicability to different or more complex situations involving multimycotoxin contamination. Second, the commercial nature of this experiment does not permit statistically valid conclusions regarding flock performance. Third, it is not possible to standardize individual mycotoxin intake in each flock under field conditions, which may result in differences in oxidative stress biomarker responses among individuals within a group. Lastly, the exact mode of action by which polyphenols from grapes and olives decrease TeA bioavailability requires further investigation. Future studies should investigate the potential of tested anti-mycotoxin agent in preventing liver damage caused by mycotoxins and reducing the exposure of TeA, using various dosages under mycotoxin contamination conditions ranging from low to high.
The lack of identification of EMs during routine analysis leads to an underestimation of mycotoxin risk. It should be highlighted that mycotoxins appear not only in their most known forms but also in their modified forms. Therefore, biomonitoring of mycotoxin risk by analyzing raw materials, feed, and biological matrices is the key to ensuring animal health and product safety. Although EMs have been shown to pose health risks only at very high concentrations, recent studies indicate that they may interact with regulated mycotoxins and produce synergistic or additive effects at low to moderate exposure levels. As a result, there is a need for (a) cumulative risk assessments that address both emerging and regulated mycotoxins at low exposure levels and (b) the development of cost-effective, environmentally friendly products that can bind a wide range of mycotoxins (Papatsiros et al., 2024a).
Conclusions
In a commercial environment with low levels of natural mycotoxins, adding a phytogenic liquid mycotoxin detoxifier to broiler drinking water lowered plasma levels of TBARS and CARBs, increased TAC, and reduced ALT, AST, and ALP activities, demonstrating antioxidant and hepatoprotective effects in the animals. Exposure of the recently discovered TeA toxin in chickens decreased by about 83%. Mortality rates were similar across all groups, although some improvement in growth performance was observed in the treatment group. The results suggest that using a phytogenic feed additive with high levels of polyphenols may be useful as a mycotoxin detoxification strategy in commercial poultry farms. However, further research on this issue is necessary.
Authors’ Contributions
All authors have made considerable contributions to this study. The conception and experimental design of the study were developed by the authors V.G.P., Z.P., and I.R. The preparation of the material and experiment, data collection, and analysis were performed by V.G.P., Z.P., C.E., N.P., G.I.P., D.A., E.M., Ó.C., N.V., I.R., P.K., K.K., and D.P. The article was prepared by the authors V.G.P., Z.P., C.E., G.I.P., E.M., N.V., K.K., and D.P. All authors read and approved the final article.
Ethical Approval
This research was approved by the Institutional Ethical Research Committee of the University of Thessaly (project identification code: 70, date of approval: 11.10.2022).
Footnotes
Acknowledgments
The authors wish to thank the managers of the trial farms for their support.
Disclosure Statement
The authors I.R. and Ó.C. were employed by Biōnte Nutrition S.L. (Reus, Spain). The other authors declare no potential conflicts of interest related to the study.
Funding Information
The authors declare that no funds were received during the preparation of this article. However, they received support by Biōnte Nutrition S.L. for the mycotoxin analysis in feed samples of the trial.
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.