
Abstract
Cronobacter sakazakii, an opportunistic foodborne pathogen associated with powdered infant formula, causes severe neonatal infections, yet its interaction with gastric epithelial cells remains poorly characterized. Here, we conducted a preliminary in vitro study to evaluate early host–pathogen responses of C. sakazakii in AGS gastric epithelial cells. AGS cells were exposed to C. sakazakii ATCC 29544 at a multiplicity of infection of 100 for 2 h, and cytotoxicity and transcriptional responses were assessed by viability assay, gentamicin protection assay, and reverse transcription-quantitative polymerase chain reaction. Infection significantly reduced AGS cell viability and increased expression of bacterial virulence-associated genes (ompA and hfq) and host inflammatory genes (IL-8 and NF-ƙB) compared with mock-infected controls (p < 0.001). Gentamicin-protected recovery was consistent with bacterial entry into AGS cells, although confirmatory imaging-based analyses were not performed. These findings indicate that C. sakazakii (ATCC 29544) can elicit early cytotoxic and pro-inflammatory responses in AGS gastric epithelial cells under in vitro conditions, supporting the need for confirmatory studies in more physiologically relevant gastric study models.
Introduction
Cronobacter spp., belonging to the Enterobacteriaceae family, have been recognized as an emerging opportunistic foodborne bacterial pathogen causing acute intestinal and extraintestinal diseases such as meningitis, necrotizing enterocolitis and septicemia in neonates and infants (Lang et al., 2024). More than 40% of the infants with meningitis infection caused by C. sakazakii have died around the world (Rounds et al., 2026). This foodborne bacterial pathogen has been isolated from a wide range of dried food products, especially the powdered infant formula (PIF) (Ekundayo and Ijabadeniyi, 2024). Systemic and intestinal infections caused by Cronobacter species follow the colonization and direct infection of the intestinal epithelial cells and subsequently reach the bloodstream (Lehner et al., 2018). Multiple in vitro and in vivo studies have demonstrated that outer membrane protein A (ompA) and hfq protein play a critical role in the adhesion to, invasion of eukaryotic intestinal epithelial cells by, and stress adaptation in Cronobacter species, especially C. sakazakii (Phair et al., 2022). Cronobacter strains lacking ompA and/or hfq genes exhibit a markedly reduced ability to invade host cells, underscoring the essential roles of these factors in Cronobacter infection (Cechin et al., 2023).
Previous work has shown that C. sakazakii can survive and express virulence-associated genes under simulated neonatal gastric conditions. However, direct evaluation of early bacterial interaction with gastric epithelial cells remains limited. The present study did not model gastric acidity or simulated gastric fluid, mucus, microbiota, immune components, or neonatal feeding matrices. Instead, it was designed to evaluate early epithelial responses to bacterial exposure under standard cell culture conditions. Direct investigation of these early host–pathogen interactions in gastric epithelial cells remains limited. (Phair et al., 2022). We hypothesized that C. sakazakii can induce gastric epithelial injury accompanied by increased transcription of key bacterial virulence determinants and host inflammatory markers. Therefore, we assessed AGS cell viability, bacterial invasion, and expression of ompA, hfq, IL-8 and NF-ƙB following short-term infection with C. sakazakii ATCC 29544.
Materials and Methods
In this study, C. sakazakii strain ATCC 29544 was purchased from ATCC (Manassas, VA, USA), revived, and cultured according to the supplier’s instructions. The human gastric cancer cell line (AGS cell line) was used for in vitro assays. AGS cells were purchased from the ATCC (Manassas, VA, USA) activated based on the institute’s instructions. AGS cells were seeded at 104 cells per well. The cells were infected at a multiplicity of infection (MOI) of 100 for 2 h at 37°C with 5% CO2, treated and evaluated according to the method previously described by Allahyari et al. (2026). Infection was performed in 2 mL serum-free and antibiotic-free medium, and the cells were not serum-starved before infection. An MOI of 100 was selected to ensure detectable host-cell and bacterial transcriptional responses during the short exposure period and to remain consistent with previously described epithelial infection approaches. The cells were harvested and evaluated for viability and expression of cellular pro-inflammatory and bacterial virulence factor genes. Mock-infected controls received the same volume of sterile PBS without bacteria. Cytotoxicity was quantified using the 3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide assay following the manufacturer’s protocol. After infection, bacterial invasion was assessed by a gentamicin protection assay as previously described by (Almajed and Forsythe 2016; Almajed and Forsythe, 2016).
Expression of IL-8, NF-ƙB, ompA, and hfq genes was measured and evaluated in AGS cells by using the reverse transcription-quantitative polymerase chain reaction (RT-qPCR) method and specific primers as previously described by Allahyari et al. (2026). Infected cells and mock-infected (PBS-treated) were subjected to total RNA extraction, cDNA synthesis, and RT-qPCR assay by using Qiagen’s RNeasy, QuantiTect Reverse Transcription kit, and RT2 SYBR Green qPCR Master-Mix kits (QIAGEN, Valencia, CA, USA), respectively, according to the manufacturers’ instructions (Allahyari et al., 2026). Relative gene expression was calculated using the 2-ΔΔCt method. Bacterial gene expression was normalized to 16S rRNA, whereas AGS-cell gene expression was normalized to Glyceraldehyde 3-Phosphate Dehydrogenase. One-way analysis of variance (ANOVA) was used to compare and determine significant (p < 0.05) differences using SPSS software version 29.0.2 (SPSS Inc., Chicago, USA). All measurements were performed in three biological replicates with technical triplicates.
Results and Discussion
After 2 h of exposed to C. sakazakii, AGS cells were evaluated for viability and expression of pro-inflammatory and bacterial virulence factor gene expression. Bacterial internalization was further supported by gentamicin protection, yielding 2.4 ± 1.8 × 104 colony-forming unit (CFU)/mL at 2 h post-infection compared with mock controls. We observed a significant reduction (p < 0.0001) in the viability of AGS cells exposure to C. sakazakii (Fig. 1A). Relative gene expression analysis of virulence factor-encoding genes indicated that (Fig. 1B and C) ompA and hfq genes were significantly (p < 0.0001) upregulated during the infection of the AGS cell line with C. sakazakii (Fig. 1B and C). Similarly, the host inflammatory genes IL-8 and NF-ƙB were significantly (p < 0.001) upregulated in infected cells NF-ƙB (Fig. 1D and E). Relative expression analysis revealed significant upregulation of ompA, hfq, IL-8, and NF-ƙB in infected cultures. These findings indicate that, within this simplified AGS-cell system, C. sakazakii ATCC 29544 elicited early cytotoxic and transcriptional responses.
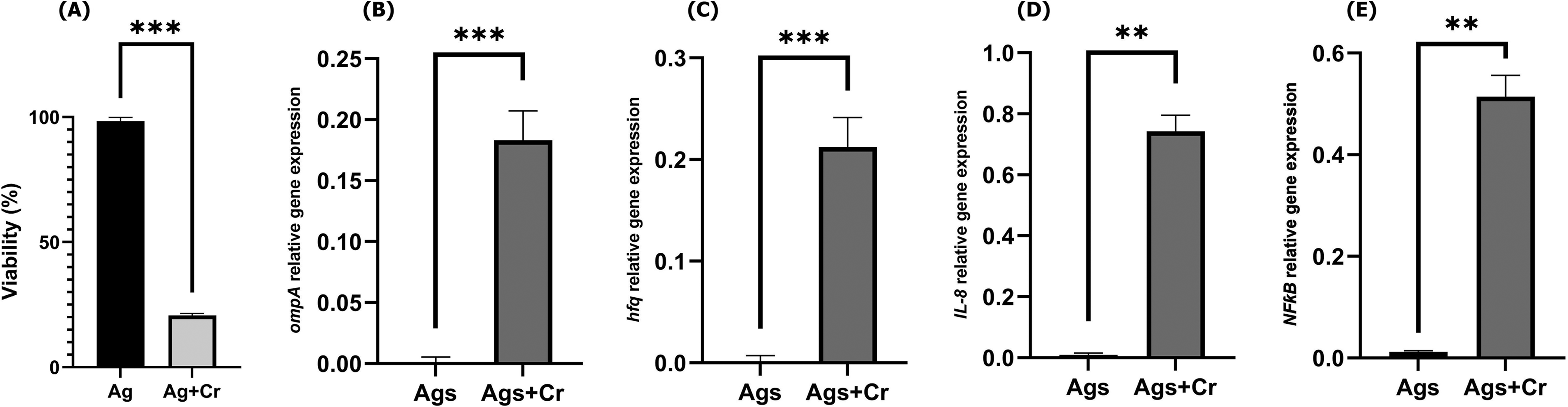
Early cytotoxic and transcriptional responses of AGS cells following infection with C. sakazakii.
The observed expression patterns are consistent with previous reports implicating ompA and hfq in C. sakazakii virulence and epithelial interaction. Hfq protein, as an RNA chaperone, interacts with the small bacterial RNAs and post-transcriptionally induces the expression of bacterial outer membrane proteins. Lack or suppression of the hfq gene in C. sakazakii contributes to attenuated pathogenicity (Kim et al., 2015). OmpA in C. sakazakii plays the most important role in the invasion of intestinal epithelial cells and expression of inflammation-associated proteins (Allahyari et al., 2026; Cechin et al., 2023; Fan et al., 2023).
Gentamicin-protected recovery of bacteria was consistent with intracellular persistence/entry in AGS cells; however, because microscopy-based confirmation and separate adhesion controls were not included, this result should be interpreted as supportive rather than definitive evidence of invasion. In addition, the present study used a single reference strain, a short exposure period, and a carcinoma-derived AGS cell line under standard culture conditions without gastric acidity, mucus, microbiota, immune components, or neonatal feeding matrices. Accordingly, the biological relevance of the model is limited, and strain-dependent variability cannot be excluded. Kireina et al. (2024) recently carried out a study to investigate the expression of C. sakazakii virulence factors in PIF under simulated neonatal gastric conditions. They exposed C. sakazakii to a simulated neonatal gastric fluid for a 4-h duration and found that C. sakazakii (ATCC 29544) can survive and express rpoS and grxB virulence factor genes during the exposure to these harsh environmental conditions (Kireina et al., 2024). Therefore, this study should be viewed as a preliminary exploratory in vitro observation rather than a physiological model of gastric infection or pathogenesis. Its contribution lies in showing that C. sakazakii ATCC 29544 can trigger early host–pathogen responses in a gastric epithelial cell context that remains less studied than intestinal models. Confirmation in more physiologically relevant gastric systems and with multiple strains will be needed to determine the broader significance of these findings. We also acknowledge that the use of a relatively high inoculum may have amplified early cytotoxic and inflammatory responses and should be considered when interpreting effect size.
Authors’ Contributions
B.P.: Conceptualization, methodology, investigation, formal analysis, data curation, visualization, and writing—original draft. S.U.M.-T.: Conceptualization, supervision, resources, writing—review and editing, and project administration.
Footnotes
Ethical Considerations
This study involved in vitro experiments using a commercially available human cell line (AGS) and did not involve human participants or animals; therefore, institutional ethics approval was not required.
Data Availability
The data supporting the findings of this study are available from the corresponding author upon reasonable request.
Author Disclosure Statement
The authors declare no competing interests.
Funding Information
This research received no external funding.