
Abstract
This 3-year surveillance study investigated the contamination patterns and genomic characteristics of Staphylococcus aureus in 2133 imported food samples, including frozen beef, frozen pork, frozen fish, frozen chicken, and cheese. An overall S. aureus contamination rate of 4.4% (93/2133) was observed, with frozen beef showing the highest prevalence (6.0%, 68/1139). The 93 isolates showed high resistance rates to ampicillin at 61.3% and penicillin at 58.1%. Methicillin-resistant S. aureus (MRSA) strains were identified in 14.0% (13/93) of them. Substantial genetic diversity was observed, with 24 sequence types (STs) and 41 spa types identified. The major prevalent clones included the globally prevalent ST5 (19.4%), of which 44.4% (8/18) carried at least one enterotoxin gene, as well as the livestock-associated ST398 (11.8%), which was significantly lower than its prevalence rate in China. The clone ST1460-t156 (10 strains) derived from equine-associated lineages was detected in strains isolated from beef samples. Virulence gene profiling revealed diverse enterotoxin carriage, with some strains carrying four or five enterotoxin genes simultaneously. Although a relatively low contamination rate of S. aureus was observed in the imported food products, the high prevalence of MRSA and carriage of several virulent strains necessitate close monitoring of S. aureus within global food supply chains to provide essential data for developing risk-based food safety interventions.
Introduction
Staphylococcus aureus is a resilient Gram-positive bacterium that can be found in a diverse array of food matrices, including egg products (Pondit et al., 2018), dairy (Morar et al., 2021), meats (Thwala et al., 2021), seafood (Sivaraman et al., 2022), and vegetables (Wu et al., 2018a). This ecological versatility intersects with expanding international food trade networks, which grew by 350% in volume between 2000 and 2021 (UNCTAD, 2024), creating unprecedented pathways for pathogen dissemination. Of particular concern is the dual public health threat posed by this bacterium: its capacity to cause rapid-onset foodborne intoxication through heat-stable enterotoxins, coupled with rising antibiotic resistance exacerbated by decades of antibiotic misuse.
Globally, S. aureus ranks as the third most prevalent cause of foodborne illness (European Food Safety Authority; European Centre for Disease Prevention and Control, 2022), responsible for approximately 241,000 annual cases in the United States and 12.5% of bacterial foodborne outbreaks in China (Li et al., 2022; Scallan et al., 2011). The known secreted virulence factors mainly include staphylococcal enterotoxins (SEs) (sea, seb, sec, sed, seh, sel, seq, and sek), exfoliative toxins (ETs A and B), and toxic shock syndrome toxin-1 (de Jong et al., 2018; Yu et al., 2023). SEs are regarded as the primary contributors to staphylococcal food poisoning (SFP). The combined effects of these toxins not only exacerbate clinical symptoms, such as vomiting, diarrhea, and fever, but also increase the risk of systemic complications, including toxic shock syndrome (Zeaki et al., 2019). Furthermore, their resilience to environmental stressors, such as heat and proteolytic enzymes, enhances their persistence in contaminated food matrices, underscoring the challenges in preventing SFP (Kadariya et al., 2014).
In recent decades, the antimicrobial resistance problem has reached alarming levels, with over 25% of foodborne isolates showing at least one class of antibiotics resistant (FAO W, 2018). Among them, the increasing antibiotic resistance in S. aureus strains is of particular concern. Specifically, methicillin-resistant S. aureus (MRSA) has emerged as a dominant pathogen, constituting a substantial proportion of infectious diseases caused by S. aureus (Xing et al., 2025). A comprehensive meta-analysis of meat and meat products shows that MRSA has an overall resistance prevalence of 3.7% (95% CI: 2.8∼5.0%) (Xing et al., 2025). Moreover, many MRSA strains have developed resistance to multiple antibiotics, including ciprofloxacin (CIP), clindamycin (CLI), tetracycline (TET), and erythromycin (ERY), thereby rendering prevention and treatment arduous (Wang et al., 2022).
China is one of the largest global importers of frozen meat products, which constitute a critical component of the national food supply. Consequently, surveillance of S. aureus contamination in these products holds substantial public health significance. Despite the existence of a limited number of studies, there remains a lack of comprehensive research on the prevalence, antibiotic resistance, and molecular epidemiological characteristics of S. aureus in food imported to China. Hence, the objective of this study was to compare the phenotypic and genotypic attributes of S. aureus isolated from diverse food types and geographical locations, while furnishing information essential for risk assessment, early warning systems, and source tracing of this bacterium. These findings can facilitate the anticipation of transmission pathways of specific S. aureus strains across various food items, thereby aiding regulatory bodies in adopting prompt and efficient preventive and control strategies to address contamination in the global food supply chain.
Methods
Food samples
A total of 2133 food samples obtained from various countries were collected during 2018–2020 to monitor the contamination of S. aureus in different imported foods. The food samples included frozen beef (n = 1139), frozen pork (n = 447), frozen fish (n = 272), frozen chicken (n = 74) and cheese (n = 201). The samples were placed in cold sterile containers at a temperature of lower than 4°C and transported immediately to the laboratory and subjected to microbiological analysis within 24 h.
Bacteria isolation
S. aureus was isolated from food samples according to the China national standard method GB 4789.10-2016. Briefly, 25 g sample was homogenized with 225 mL of 7.5% sodium chloride enrichment broth and incubated aerobically at 36°C ± 1°C for 18 h∼24 h. The enriched culture was then streaked onto Baird-Parker agar (BPA) and sheep blood agar, and the plates were incubated at 36°C ± 1°C for 24 h∼48 h (BPA plates) and 18 h∼24 h (sheep blood agar plates), respectively. Presumptive S. aureus colonies on the above plates were selected for Gram staining microscopy and plasma coagulation enzyme test. Isolates were considered coagulase-positive if plasma coagulation occurred within 4 h at 37°C. For definitive molecular confirmation, multiplex real-time PCR was performed according to the AQSIQ Industry Standard (AQSIQ, 2016) (Supplementary Table S1).
Antibiotic susceptibility test
The antibiotic susceptibility test was performed by the microdilution method. The following 13 antibiotics were tested: ampicillin, penicillin, oxacillin, ERY, CLI, CIP, daptomycin, sulfamethoxazole, vancomycin, TET, gentamicin, and chloramphenicol. The minimum inhibitory concentration values for breakpoints were referred to Clinical and Laboratory Standards Institute (CLSI) guidelines (CLSI, 2025). S. aureus ATCC29213, Escherichia coli ATCC25922 and Enterococcus faecalis ATCC 29212 were used as quality control strains. The criterion for defining MRSA is that the strain is resistant to either cefoxitin or oxacillin, or carries the mecA/mecC gene, which were identified using WGS analysis.
Whole genome sequencing
The purpose of conducting WGS was to systematically analyze the virulence genes, antibiotic resistance genes, and evolutionary relationships of S. aureus isolates. Genomic DNA extraction was performed using the DNeasy® Blood & Tissue Kit (Qiagen, Valencia, CA, USA). Sequencing libraries were constructed with the NEBNext® Ultra™ II DNA Library Prep Kit (New England Biolabs, Ipswich, MA, USA) and sequenced on an Illumina HiSeq X Ten platform with paired-end 2 × 150 bp. Raw reads underwent quality control processing, including adapter trimming and removal of low-quality sequences (Phred score ≤ 20) using Trimmomatic v0.39 (Bolger et al., 2014). De novo genome assembly was conducted via SOAPdenovo2 (version r240) with a minimum contig length threshold of 500 bp for downstream analysis (Luo et al., 2012).
Bioinformatic analysis
The presence of virulence factors was determined by BLAST alignment against the Virulence Factor Database (VFDB; http://www.mgc.ac.cn/VFs/). Antimicrobial resistance genes were identified through the Comprehensive Antibiotic Resistance Database (http://card.mcmaster.ca) using default parameters. Staphylococcal Protein A (spa) typing was performed through comparative genomic analysis using the Spa Server platform (https://cge.food.dtu.dk/services/spaTyper/). Multi-locus sequence typing (MLST) was conducted using the S. aureus MLST database (https://pubmlst.org/saureus/). Genome annotation was inferred by Prokka v1.14.6 (Seemann, 2014), core genome analysis was performed by Roary v3.13.0 with default parameters (Page et al., 2015). Gubbins v2.4.1 was used to filter recombinant regions. Finally, based on the core genome alignment, a maximum likelihood tree was constructed using RAxML v8.2.12 with the GTR + GAMMA substitution model and 100 bootstrap replicates. The tree was rooted using Staphylococcus argenteus as the outgroup (Stamatakis, 2014) and visualized using ChiPlot (Xie et al., 2023).
Result
Prevalence of S. aureus in imported foods
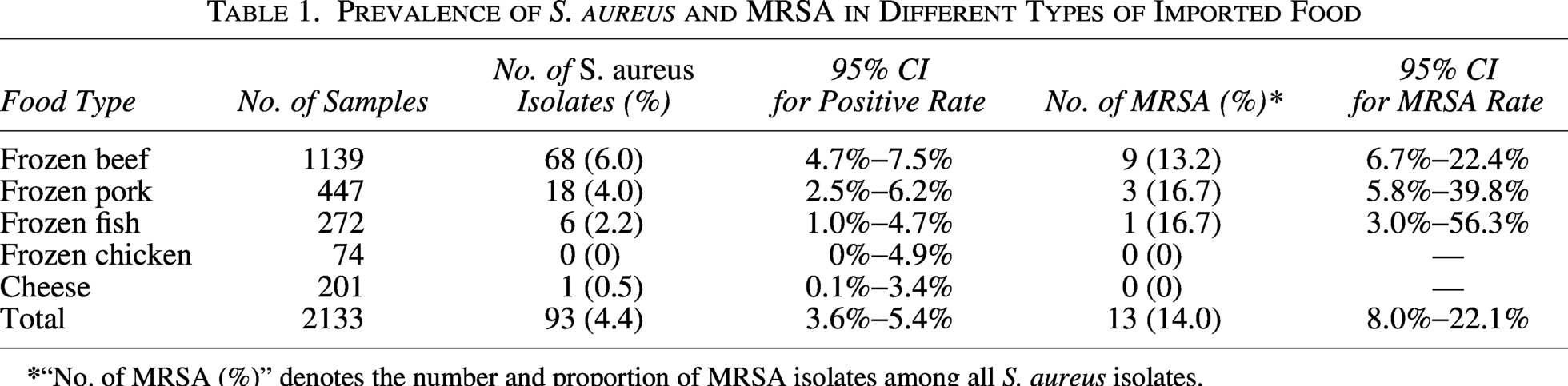
The prevalence of S. aureus varied significantly among food types (χ2 = 13.85, df = 4, p < 0.05). The overall prevalence of S. aureus in 2133 food samples analyzed was 4.4% (Table 1). The highest contamination was found in frozen beef at 6.0%, followed by frozen pork (4.0%), frozen fish (2.2%), and cheese (0.5%).
Prevalence of S. aureus and MRSA in Different Types of Imported Food
Antibiotic susceptibility
A total of 93 isolates of S. aureus were tested for their resistance to 13 different antibiotics (Table 2). Resistance to ampicillin was the highest, reaching 61.3% (57/93), followed by penicillin (58.1%) and ERY (24.7%). Resistance to the remaining antibiotics was as follows: oxacillin (14.0%), TET (12.9%), cefoxitin (11.8%), CLI (12.9%), chloramphenicol (8.6%), daptomycin, and CIP (6.5%). In this study, the overall detection rate of MRSA was 14.0% (13/93). Among these 13 MRSA isolates, 11 harbored the mecA gene and one carried the mecC gene.
Antibiotic Resistance of S. aureus Isolates Obtained from Imported Foods
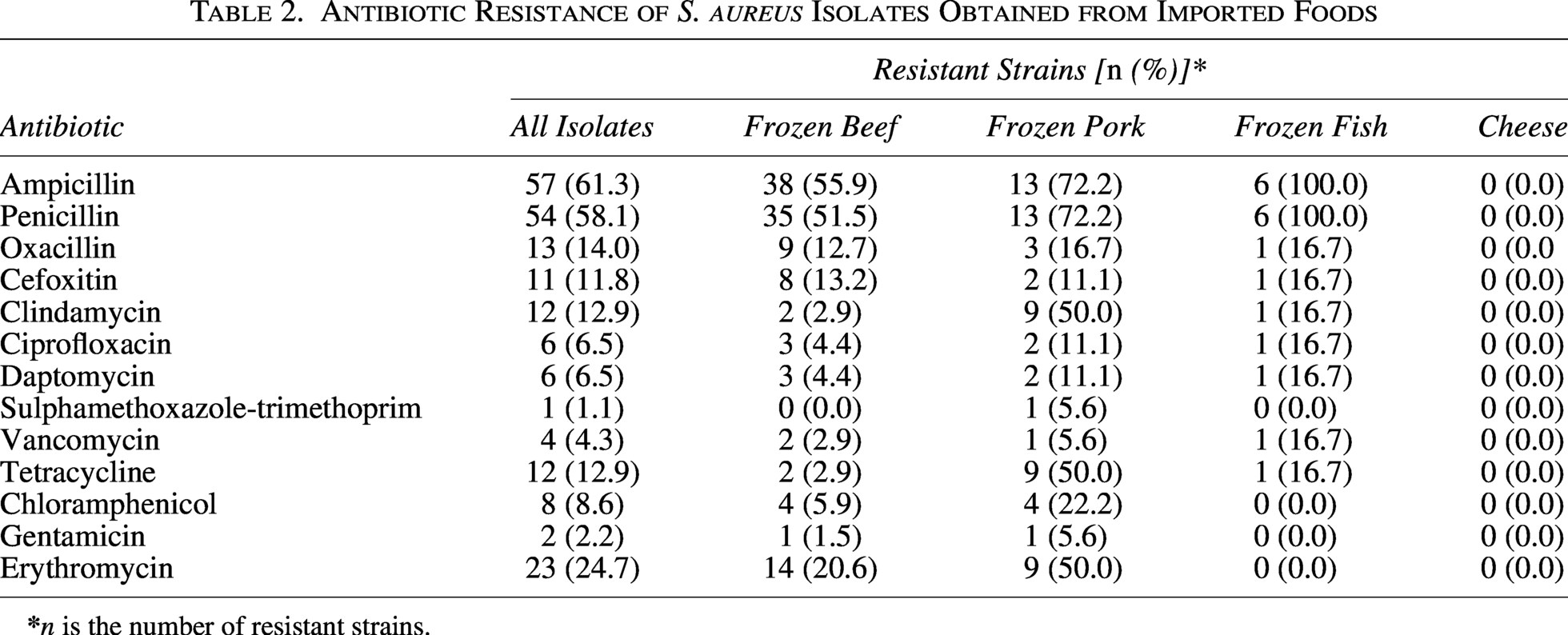
In terms of multidrug resistance (MDR), defined as resistance to three or more antimicrobial classes, 21 strains (22.6%, 21/93) were identified as MDR (Supplementary Table S2). Among these 21 MDR strains, 4 (4.3%, 4/93) and 2 (2.2%, 2/93) exhibited resistance to 5 and 6 antimicrobial classes, respectively. Notably, one strain showed resistance to nine classes.
Multi-locus sequence typing analysis
A total of 24 sequence types (STs) were identified (Table 3), with ST5 (18/93, 19.4%) and ST398 (11/93, 11.8%) representing the most predominant lineages, followed by ST1460 (10/93, 10.8%) and other minor STs including ST9, ST188, ST6, ST7, ST15, and ST45. Among the 13 MRSA isolates, ST398 was the most prevalent ST, accounting for 23.1% (3/13). This was followed by ST5, ST59, and ST45, each representing 15.4% (2/13). The remaining four isolates were distributed among ST88, ST9, ST72, and ST6, each comprising 7.7% (1/13).
The STs and Spa Types of S. aureus Identified in Different Foods

*“No.” indicates the number of isolates for each spa type. “Food types” denotes the source food category: B = frozen beef, P = frozen pork, F = frozen fish, C = cheese.
Notably, 25% of STs (6/24) were detected in multiple food types, including ST5, ST7, ST15, ST88, ST188, and ST398, each found in two distinct food categories. (Fig. 1). In contrast, 13 STs were only isolated from frozen beef samples. Among them, the most frequent was ST1460, followed by ST6, ST45, ST25, ST30, ST59 and ST72, and ST78, ST81, ST152, ST352, and ST3510.
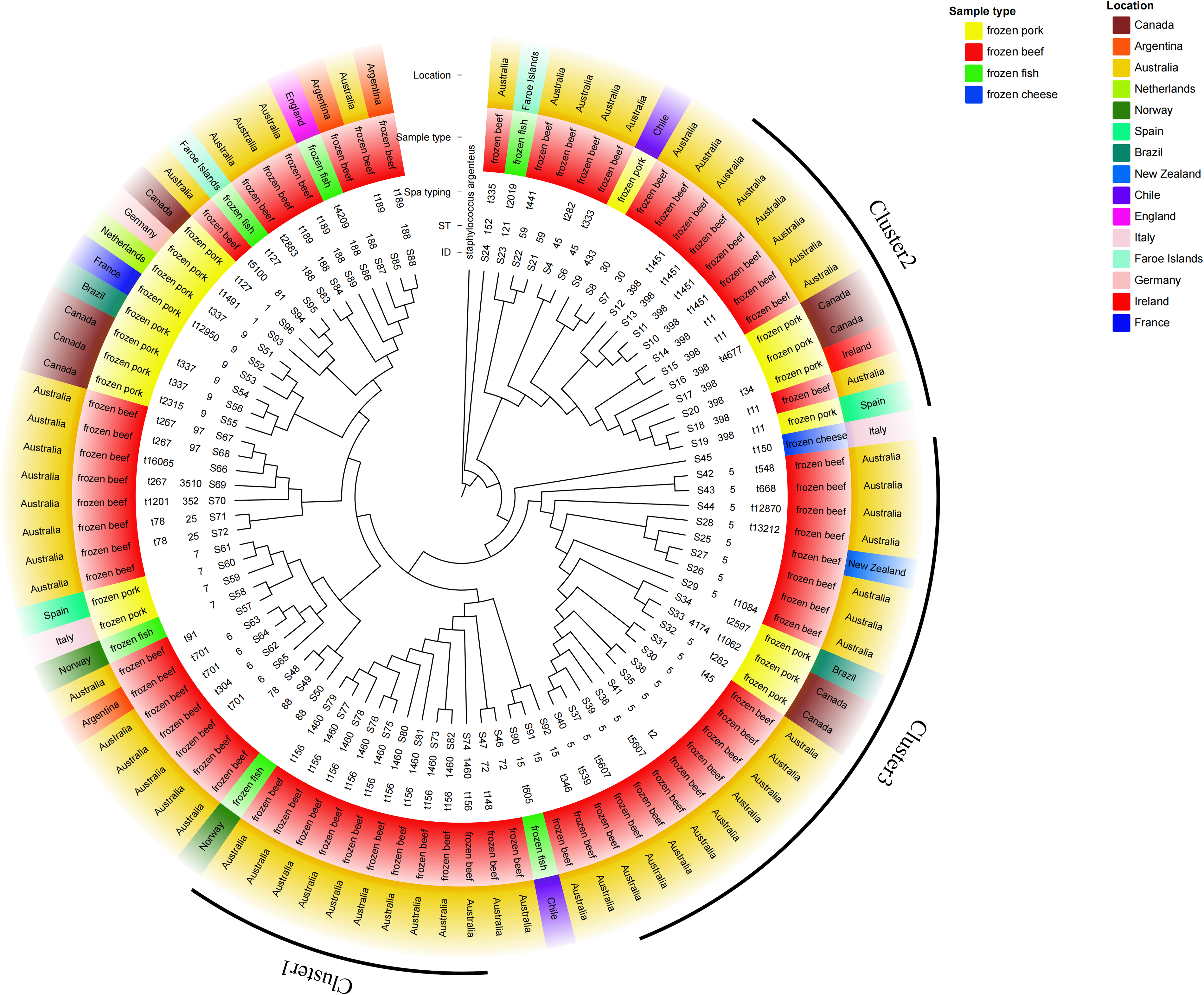
WGS based phylogenetic tree of S. aureus isolates from imported foods. A Staphylococcus argenteus strain was used as outgroup. Phylogenetic tree was annotated from inside to outside by ST, spa type, sample type, and location.
Spa typing analysis of S. aureus
A total of 41 different spa types were identified (Table 3). The t156 type was the most frequent spa type, identified in 10.8% (10/93) of isolates and exclusively associated with ST1460 lineages. The secondary prevalent types included t011, t189, and t1451, each accounting for 4.3% (4/93) of the isolates. Other spa types included t189, t701, t337, t333, and etc. Of the 13 MRSA isolates, spa types were successfully detected in 9 strains. Among these detected spa types, t011 and t333 had the highest detection rates, both at 15.4% (2/13). The remaining detected spa types were t1451, t282, t441, t668, and t304, with each type showing a detection rate of 7.7% (1/13). Out of the 41 distinct spa types identified in this study (Supplementary Table S3), 28 spa types were detected in the 68 beef strains, 11 spa types were detected in 18 pork strains, 4 spa types were detected in 6 fish strains, and one spa type was identified in the only cheese strain. Specifically, in beef strains, t156 emerged as the most prevalent type, with a frequency of 10 out of 68. In pork strains, t337 was found in 3 out of 18 isolates, which stood out among the detected spa types.
Carriage of enterotoxins genes
A variety of enterotoxin genes were detected among the 93 S. aureus strains (Supplementary Table S4). Among SE genes, sec was the most prevalent (20.4%, 19/93), followed by seq, sea and sek (16.1%, 15/93), sel (15.1%, 14/93), seb (11.8%, 11/93), sed (5.4%, 5/93), and seh (4.3%, 4/93). Notably, 49.5% (46/93) of strains harbored at least one SE gene, with seven strains carrying three distinct SE types, four strains harboring four, and two strains encoding five SE genes.
Phylogenetic analysis based on WGS
Phylogenetic analysis of all 93 S. aureus isolates identified multiple evolutionary clades in S. aureus strains, among which Cluster 1 exhibited a high degree of concordance with MLST and spa typing (Fig. 1). The Cluster 1 clade, comprising the representative ST1460-t156 genotype, consisted exclusively of 10 frozen beef isolates originating from Australia. The Cluster 2 clade also presented a unique phylogenetic clade, as all 11 strains within it belonged to a single ST type, namely ST398, but there were more than four different spa types within it. Among them, t1451 and t11 accounted for 4/11 each, t034 and t4677 accounted for 1/11 each, and spa types were not identified for the remaining one isolate. For the remaining clades, although they exhibit a certain degree of correlation with ST types, some STs are interspersed among them. For instance, ST4174 was found interspersed among the ST5 dominant Cluster 3.
Discussion
Monitoring the prevalence, antibiotic resistance, and molecular epidemiology of S. aureus in globally imported foods is essential for microbial risk assessment, contamination source identification, and targeted surveillance. This study showed that the S. aureus contamination levels in imported food samples collected at Shanghai port were significantly lower than the global average values. For instance, frozen beef showed a 6.0% prevalence rate, significantly lower than the 29.7% (18.4%∼42.3%) global average for raw beef; frozen pork exhibited a 4.0% detection rate, well below the 23.5% (15.6%∼32.5%) contamination rate reported for raw pork worldwide (Ou et al., 2017). In addition, the positive detection rate of frozen fish was only 2.2%, which is significantly lower than the levels of 30%∼48.7% that have been reported in multiple countries (Costa et al., 2015; Okoli et al., 2018; Vázquez-Sánchez et al., 2012). In addition, the overall isolation rate of S. aureus in the retail meat products in China ranged from 19.7% to 35% (Wu et al., 2018b; Zhao et al., 2025), which is significantly higher than our observed rate of 4.4%. The relatively low S. aureus contamination rate observed in this study might be attributed to cold-chain transportation and storage conditions, which inhibited bacterial growth and therefore its spread (Arriaga-Lorenzo et al., 2023; Wang et al., 2024).
In this study, the MDR rate of S. aureus was 22.6%. While in China, the prevalence of MDR in S. aureus isolated from foods ranges from 26.1% to 82.6% (Gan et al., 2021; Wang et al., 2017a; Zhou et al., 2025). The relatively low MDR levels from imported foods are likely attributable to the stringent regulatory controls on antimicrobial use in the exporting countries (Australian Government, 2024; Australian Pesticides and Veterinary Medicines Authority, 2023). Similar to that had been documented in other research findings (Ghabbour et al., 2022; Mahros et al., 2021; Torki Baghbaderani et al., 2020), our S. aureus isolates showed high resistance rates of 61.3% and 58.1% to ampicillin and penicillin, respectively. The findings indicate that penicillin and ampicillin may have relatively limited efficacy in the treatment of foodborne S. aureus infections, and thus, priority should be given to other classes of antibiotics in clinical treatment or environmental intervention. ERY also exhibited considerable resistance at 24.7%, which falls within the median range of the global surveillance interval (8.7%∼87.3%) (Mesbah et al., 2021; Tsehayneh et al., 2021; Zhou et al., 2025; Zwe et al., 2025), and is also lower than the 31.5%∼87.2% resistance rates reported in Chinese studies (Li et al., 2022; Liao et al., 2018; Wu et al., 2018b; Zhou et al., 2025). Therefore, it remains critical to strengthen supervision in this field and raise public awareness of antibiotic resistance.
MRSA is a significant public health concern in humans and animals. In this study, the overall positive rate of MRSA was 14.0%. Similarly, several studies have reported that the percentage of positive isolates of MRSA in food at 11.5% in Turkey (Şanlıbaba, 2022), 6.7% in the United States (Ge et al., 2017), 16.7% in Iran (Safarpoor Dehkordi et al., 2017), 21.5% in Algeria (Chaalal et al., 2018), 4.9% in Japan (Hiroi et al., 2012), and 6.8% in China (Wu et al., 2019). These findings indicate that the overall MRSA positive rate in this study is consistent with international reports on food contamination and exceeds that observed in domestic Chinese meats. Therefore, targeted prevention and control measures are critical to reduce MRSA transmission risks through imported meats.
In the present study, ST5, ST398, ST1460, and ST9 collectively accounted for 48.4% of all STs. Notably, ST5 also exhibits a high clinical prevalence (Gu et al., 2020; Piper, 2025; Zhan et al., 2024) and accounted for 21.4% of the S. aureus strains causing bloodstream infections (Li et al., 2018). Additionally, ST5 strains frequently harbor a diverse array of toxin genes (Lee et al., 2014; Zheng et al., 2024). Indeed, among the 18 ST5 strains isolated in this study, nearly half (about 44.4%) carried at least one enterotoxin gene. These toxins not only serve as the primary pathogenic factors in SFP but also exacerbate the severity of clinical manifestations, including vomiting, diarrhea, and fever (Zeaki et al., 2019). ST398 was detected at 11.8%, consistent with its global prevalence as a livestock-associated MRSA (Voss et al., 2005). Of note, 36.4% (4/11) of ST398 isolates originated from pork, confirming its porcine association. While ST1460 is found associated with equine hosts (Mama, 2019), all 10 strains in this study were recovered from beef, indicating a possible risk of cross-species transmission. ST9 strains were exclusively pork-derived (Wang et al., 2017b), further confirming the specific carriage of ST9 strains in pork-derived foods.
The most prevalent spa type in this study was t156 (10.8%, 10/93), yet it is not a globally predominant spa type, warranting attention to its potential for localized outbreaks. Ranking second in detection rate, all t189 isolates were from beef samples. Notably, a Chinese study on dairy farms reported a high proportion of t189 (Zhao et al., 2021), and it was the most prevalent spa type in community-associated S. aureus from pediatric bloodstream infections (Wang et al., 2018), underscoring the need for enhanced surveillance of t189. Regarding the spa types with the third-highest prevalence, spa type t701 is commonly detected in food isolates and is often associated with ST6. Importantly, ST6-t701 clones are frequently implicated in foodborne intoxication outbreaks and have become the predominant clones responsible for SFP in China (Li et al., 2015; Liao et al., 2020; Yan et al., 2012). Several spa types identified in this study require heightened attention despite their relatively low detection rates. Specifically, t437 is a predominant community-associated MRSA clone in Asia (Song et al., 2011; Zhao et al., 2019), while t127 ranks sixth among the most common clones from human infections in Europe (Grundmann et al., 2010; Karampatakis et al., 2021).
SEs are pivotal virulence factors in S. aureus-mediated foodborne poisoning, with epidemiological studies indicating that 95% of outbreaks are associated with classical SE genes (De Buyser et al., 2001). In our study, classical SE genes (e.g., sec, sea) ranked among the most frequently detected enterotoxin genes, further supporting the potential of these strains to cause foodborne diseases (Aydin et al., 2011; Buyser et al., 2001). Notably, 49.5% of the strains harbored three or more SE genes. This is particularly concerning given that strains carrying multiple enterotoxin genes have been implicated in prior S. aureus-related food poisoning incidents (Roussel et al., 2015). Therefore, greater attention should be paid to monitoring S. aureus strains with multiple enterotoxin genes in food safety surveillance.
Conclusion
This study revealed a diverse array of S. aureus strains were found in the imported food items in China, with a notable prevalence of antimicrobial-resistant and MDR isolates. These findings highlight the need for heightened awareness of the potential risks posed by these strains and emphasize the importance of strengthening surveillance efforts focused on detecting S. aureus with high virulence potential and antimicrobial resistance in imported food products.
Authors’ Contributions
Conceptualization: J.S. and S.Z.; Methodology: J.Y., Y.J., Q.L., J.S. and S.Z. Software: H.M.; Data curation: H.M. Formal analysis: H.M. Investigation: L.Z., Y.Z. and X.W. Resources: J.S. and S.Z. Validation: L.Z., Y.Z., X.W., Y.F., M.X., Q.L. and J.S. Visualization: H.M. Writing—original draft: H.M., L.Z., Y.Z., X.W., Y.F. and M.X. Writing—review and editing: H.M., J.S. and S.Z. Supervision: J.Y., Y.J., J.S. and S.Z. Funding acquisition: J.S. and S.Z.
Supplemental Material
sj-docx-1-fpd-10.1177_15353141261466472 — Supplemental material for Prevalence, Antibiotic Resistance, and Molecular Epidemiology of Staphylococcus aureus Isolated from Imported Foods in China
Supplemental material, sj-docx-1-fpd-10.1177_15353141261466472 for Prevalence, Antibiotic Resistance, and Molecular Epidemiology of Staphylococcus aureus Isolated from Imported Foods in China by Hangtian Ma, Lina Zhao, Yuqiu Zhang, Xin Wang, Jilin Yang, Yuan Jiang, Yu Feng, Min Xu, Qiong Liu, Jinling Shen, and Shuai Zhi
Footnotes
Acknowledgment
This work was supported by the National Key R&D Program for the 14th Five Year Plan “National Quality Infrastructure System” (2024YFF0619404), the Ningbo Natural Science Foundation (2023S071), the One health Interdisciplinary Research Project, Institute of One Health Science, Ningbo University (NBUOH202503), and State Key Laboratory for Quality and Safety of Agro-products (SKLQSAP-2026XD01).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.