
Abstract
In the present study, the protective effects of luteolin were investigated against psychological stress-induced cognitive impairment. To emulate the psychological stress, mice received restraint stress for six hours daily, between 9:00 and 15:00 hours, for 21 consecutive days. The results of step-through test, open-field test and Morris Water Maze test demonstrated that psychological stress treatment could result in cognitive impairments in mice. This cognition dysfunction was improved by treatment with low- and medium-dose luteolin. In addition, psychological stress induced an increased serum corticosterone concentration with a decreased serum norepinephrine and dopamine concentration. These alterations were attenuated by treatment with luteolin. Also, psychological stress significantly decreased the glutathione (GSH) concentrations and superoxide dismutase (SOD) activities in prefrontal cortex and hippocampus, while the malondialdehyde (MDA) concentrations were enhanced. However, these oxidative alterations in prefrontal cortex and hippocampus induced by psychological stress were significantly reversed by treatment of luteolin. Further, the current study indicated a decline of catalase (CAT) activities in the hippocampus of the ST group, which was significantly prevented by low, medium and high dose of luteolin. On the other hand, there was no significance in CAT activities of the prefrontal cortex among the six groups. Collectively, the present results suggest that luteolin treatment serves as a key role in improving the psychological stress-induced cognitive impairments.
Introduction
Stress is defined as a condition in an organism that results from the action of one or more stressors that may be of either external or internal origin. 1 Brain is the target for different stressors because of its high sensitivity to stress-induced degenerative conditions. It is well known that psychological stress can induce a series of dysfunctions of central nervous system, such as cognitive impairment, anxiety, amnesia and insomnia. The development of many psychosomatic diseases was also correlated with psychological stress, for example, tumor, hypertension and diabetes. 2–5 These observations indicate a need to develop therapeutics to alleviate the psychological stress responses in individuals. Therefore, close attentions were focused on studies of the protective effects of natural potential ingredients on psychological stress-induced injury.
Flavonoids, plant polyphenolic compounds abundant in fruits and vegetables, exhibit a wide array of biological effects, including antioxidant, free-radical scavenging and antinflammatory properties. Recently, interests in luteolin, a flavonoid isolated from Cirsium japonicum, have been intensified because of their physiological function, such as antioxidant, anti-inflammatory and neuroprotective activities. 6,7 Several studies also demonstrate that luteolin could improve learning and memory in aged mice. 8,9 Moreover, previous research has indicated that psychological stress could impair central nervous system and result in brain tissue injury. 10–12 However, little information is available on the protective effects of luteolin against cognitive dysfunction induced by psychological stress. Therefore, the primary aim of this research was to evaluate whether luteolin could improve cognitive impairments induced by psychological stress in mice and to explore the possible mechanisms underlying this action.
Materials and methods
Agents and chemicals
Luteolin (purity >98%) was purchased from Shanxi Sciphar Biotechnology Co. Ltd (Shanxi, China). Corticosterone, norepinephrine and dopamine enzyme-linked immunosorbent assay (ELISA) kits were purchased from Adlitteram Diagnostic Laboratories (San Diego, CA, USA). Malondialdehyde (MDA), superoxide dismutase (SOD), glutathione (GSH) and catalase (CAT) kits were purchased from Nanjing Jiancheng Biotechnology Co. Ltd (Nanjing, China).
Animals
Adult male Kunming mice (6 weeks old, weighing 18–23 g) were housed with free access to food and water. The animals were kept at regulated temperature (22 ± 2°C) and humidity (50 ± 10%) on a 12-hour light/dark cycle with ad libitum access to standard commercial food and pure water. After acclimating to laboratory conditions, the mice were divided at random into six groups of 10 animals each: control group (CT), stress group (ST), control + medium dose of luteolin group (CM), and three stress groups with low, medium and high doses of luteolin modulation, respectively (LD, MD and HD). The mice received humane care throughout the experiment according to the Chinese Council on Animal Care and Institutional Care Committee of Xuzhou Medical College. All efforts were made to minimize both the number of animals used and their suffering.
Drugs and treatment schedule
CT and ST groups of mice received 0.5% sodium carboxymethyl cellulose as vehicle treatment. Luteolin was suspended in 0.5% sodium carboxymethyl cellulose and daily administered intragastrically at doses of 25, 50 and 100 mg/kg 30 min before the animals were subjected to psychological stress for three consecutive weeks. The CM group received the medium dose of luteolin at the same time to that of LD, MD and HD groups, while was not received psychological stress treatments. All compounds were administered systemically in a volume of 0.01 mL/g body weight.
Psychological stress model
The method used in this study was a modified method for psychological stress as described previously, 13,14 of which the animal model of psychological stress was developed by restraint for six hours daily in well-ventilated polypropylene tubes, with no physical suppression or pain experience in mice. The tubes were 10.5 cm in length long and 3.2 cm in diameter. Each tube was covered on one end with a Plexiglas covering, which had a small air hole (0.2 cm in diameter) in it. This protocol was scheduled for three consecutive weeks. During the fourth week, animals were subjected to step-through test, open-field test and subsequently followed by a Morris Water Maze (MWM) test. Thereafter, the animals were anesthetized with ether and killed by decapitation after aortic exsanguinations. Blood was collected and serum isolated from the whole blood. Brains were rapidly removed, and the prefrontal cortex and hippocampus tissues were isolated. The samples were stored at −80°C until processed for biochemical estimations.
Step-through test
One day after the end of the treatment with luteolin or vehicle, the step-through passive avoidance response was immediately performed according to previous study with minor modifications. 15 The step-through apparatus consists of six compartments of equal size (15 × 10 × 11 cm). Each compartment was divided into a light and a dark chamber by a guillotine door (3 cm in diameter). The light chamber was equipped with an illuminator. As described previously, 15 a single acquisition (training) trial was conducted prior to test. The trial consisted of a 30-s adaptation in one chamber which was dark at first, then followed by the onset of a bright light. To avoid the light, the mice crossed over into the darkened adjacent chamber and received a mild 0.8 mA footshock for one second. The time to crossover from the light chamber to the darkened chamber (latency) was recorded with a maximum trial length of 180 s. The mice were tested for recall 24 h after the acquisition period to determine whether they had been trained to criterion by remaining in the light chamber for 180 s to avoid the footshock. Any mice that did not meet the criterion were restrained. The latency and number of mistakes were used to measure the ability of the rats to remember adverse stimuli.
Open-field test
One day after the step-through test, the ambulatory behavior was assessed in an open-field test apparatus consisting of a square wooden arena (30 × 30 cm) marked with black lines. 16 The floor of the wooden arena was evenly divided into 6 × 6 cm squares marked with black lines. In the test, mice were individually placed in the center of the arena and permitted for free explorations. After 20-min adaptation, the number of squares crossed by the mice and the number of rearing on the hind paws were recorded during a test of three minutes. Between trials, the mouse was returned to its home cage in the same room, and open field was cleansed with a detergent and dried before occupancy by another mouse.
MWM test
Following the open-field test, the MWM test was carried out based on the method described previously 17 with minor modifications. The experimental apparatus consisted of a circular water tank (120 cm in diameter, 60 cm in height) located in a test room with several cues on white walls. With its inner surface painted black, the pool was filled with water to a height of 40 cm (maintained at 25 ± 1°C), covering an invisible (black) 10 cm square platform which was located approximately 1.5 cm below the surface of the water. The pool was virtually divided into four quadrants and the platform was placed at a fixed position in the center of a quadrant. Four different start points (NE, SE, SW and NW) were equally spaced around the circumference of the pool.
The MWM test lasted for five consecutive days following the 21 d of stress treatment, in which mice were trained in the MWM and their latency to platform was recorded with four trials per day over four days. At the beginning of each trial, the mice were released at one of the four possible starting points facing the wall, and allowed to swim freely until they reached the platform. If mouse did not find the escape platform within 90 s, it was given a latency score of 90 s. The inter-trail interval was 10 min. At the end of each session, all animals were dried and returned to their home cages. Twenty-four hours following the four consecutive days, the platform was removed. Mice were scored during a 90-s probe trial for latency to reach the platform and the time spent in the target quadrant to assess consolidation and retrieval of memory.
Brain homogenate
Following the behavior assessments, the animals were sacrificed with the brain removed, and the prefrontal cortex as well as hippocampus was isolated. To examine biochemical parameters, the prefrontal cortex and hippocampus were weighed, homogenized in nine volumes (1:9 w/v) of cold saline to prepare a 10% tissue homogenate in an ice bath and centrifuged at 1699
Estimation of oxidative alterations in prefrontal cortex and hippocampus
The changes of MDA, SOD, GSH and CAT in prefrontal cortex and hippocampus were measured by their corresponding kits (Jiancheng Bio-tech, Nanjing, China) according to the manufacturer's instructions. Measurement in all assays described above was carried out in triplicates. The protein amount in all samples was measured by the bicinchoninic assay.
Blood samples collection
Blood samples were collected from the orbital sinus in 2 mL Eppendorf tubes and centrifuged at 1699
Measurement of serum corticosterone, dopamine and norepinephrine concentrations
Corticosterone concentrations in the serum were measured using the Corticosterone ELISA kit (Adlitteram Diagnostic Laboratories) for mice according to the manufacturer's instructions. The kit uses a double-antibody sandwich ELISA to assay the level of mouse corticosterone in samples and the detection limit of the assay was 5 ng/mL. Similarly, the concentrations of dopamine and norepinephrine in serum samples were also respectively analyzed by their corresponding ELISA kits.
Statistical analysis
All statistical analyses were performed using the SPSS software, version 13.0 (SPSS Inc., Chicago, IL, USA). Values are expressed as mean ± SEM (standard error of the mean). For the MWM test, data in escape latency were analyzed by two-way analysis of variance (ANOVA), the factors being treatment and training day. Statistical significance was set at P < 0.05. For the open-field, step-through, biochemical and neuroendocrine assays, differences between groups were analyzed with one-way ANOVA followed by the Dunnett's post hoc test. Results were considered significant at P < 0.05.
Results
Performances in step-through test of mice
In the acquisition trial, the step-through latencies did not differ among the five groups (P > 0.05). The step-through latency in the 24-h retention trail significantly decreased in the ST group in contrast to the CT group (P < 0.001). Treatment with luteolin (25, 50 and 100 mg/kg) significantly increased the step-through latency as compared with the ST group (P < 0.01 or P < 0.001, Figure 1). Treatment with luteolin did not affect the step-through latency during either the training or retension sessions for the mice without restraint stress treatments.
Behavioral performances of mice in step-through test. The CT group and the ST group were given daily with 0.5% sodium carboxymethyl cellulose for three consecutive weeks. Three stress and luteolin modulation groups were administered daily with low, medium and high doses of luteolin, respectively. The CM group received medium dose of luteolin without restraint stress treatments. All stress groups received restraint for three consecutive weeks. At the fourth week, the performances of mice in step-through test were examined. Data are presented as mean ± SEM (n = 10). ***P < 0.001 versus CT group.
##P < 0.01, ###
P < 0.001 versus ST group. CT, control group; ST, stress group; LD, low dose; MD, medium dose; HD, high dose; CM, control + medium dose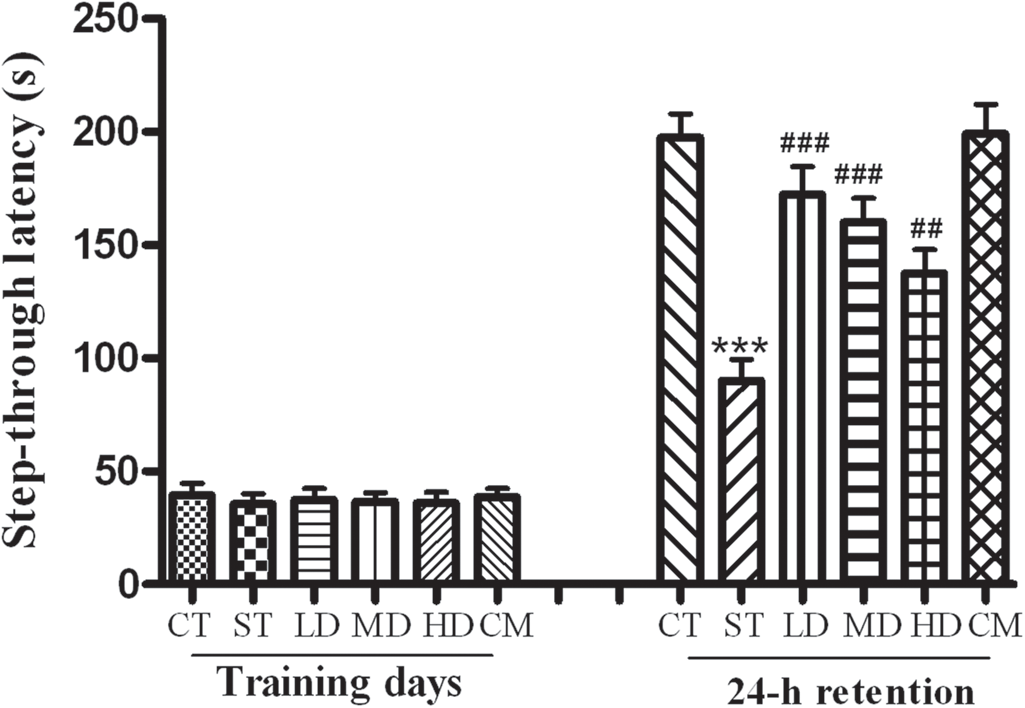
Performances in open-field test of mice
Compared with the CT group, the crossing and rearing frequencies of the ST group in the open-field test were significant decreased, and these changes were improved in the LD group (P < 0.05). Furthermore, there were no remarkable difference in crossing and rearing frequencies of the MD and HD groups in contrast to that of the ST group (Figures 2a and b, P > 0.05). However, no significance in the crossings and rearings was found between mice treated with luteolin alone and the mice of the CT group.
Number of line crossings, rearings of mice in the open-field test. Data are presented as mean ± SEM (n = 10). *P < 0.05 versus CT group,
#P < 0.05 versus ST group. CT, control group; ST, stress group; LD, low dose; MD, medium dose; HD, high dose; CM, control + medium dose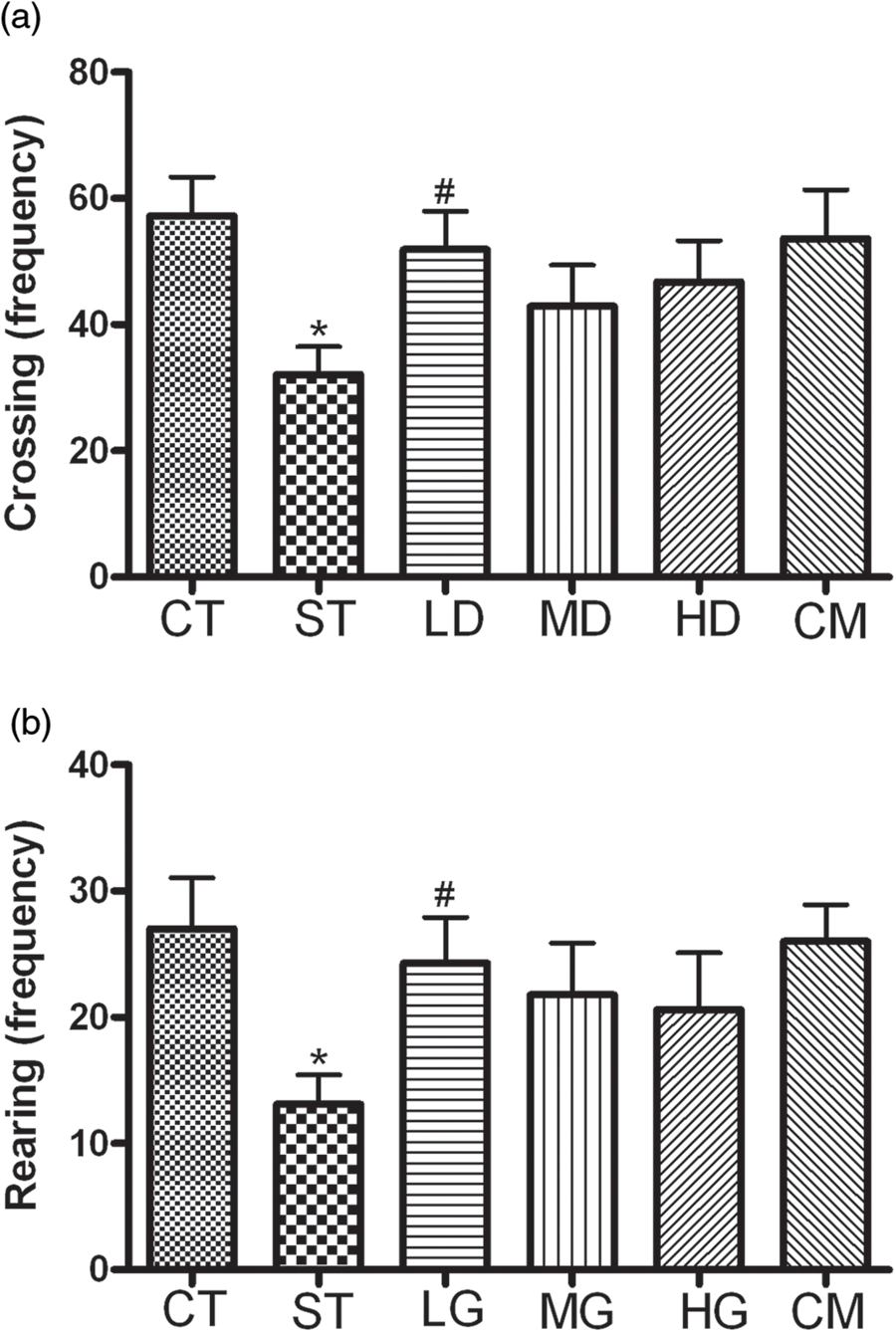
Performances in MWM test of mice
To examine the influences of luteolin on cognitive impairment induced by psychological stress, we tested the learning and memory using the MWM test and the results are shown in Figure 3a. The mean escape latency did not differ between any of the groups on the first day of testing in MWM test. While from the second day onwards, there was a significance difference in escape latency between CT and ST animals (P < 0.05). Chronic luteolin treatment significantly decreased the escape latency in stressed mice (Figure 3a). However, the mice in the HD group showed no significance in latency to reach the platform compared with the ST group (P > 0.05).
Behavioral performances of mice in Morris Water Maze test. Escape latency appeared during the training and the probe sessions (a). Time spent in target quadrant during the probe trial (b). Data are presented as mean ± SEM (n = 10). *P < 0.05, ***P < 0.001 versus CT group.
#P < 0.05,
##P < 0.01, ###
P < 0.001 versus ST group. CT, control group; ST, stress group; LD, low dose; MD, medium dose; HD, high dose; CM, control + medium dose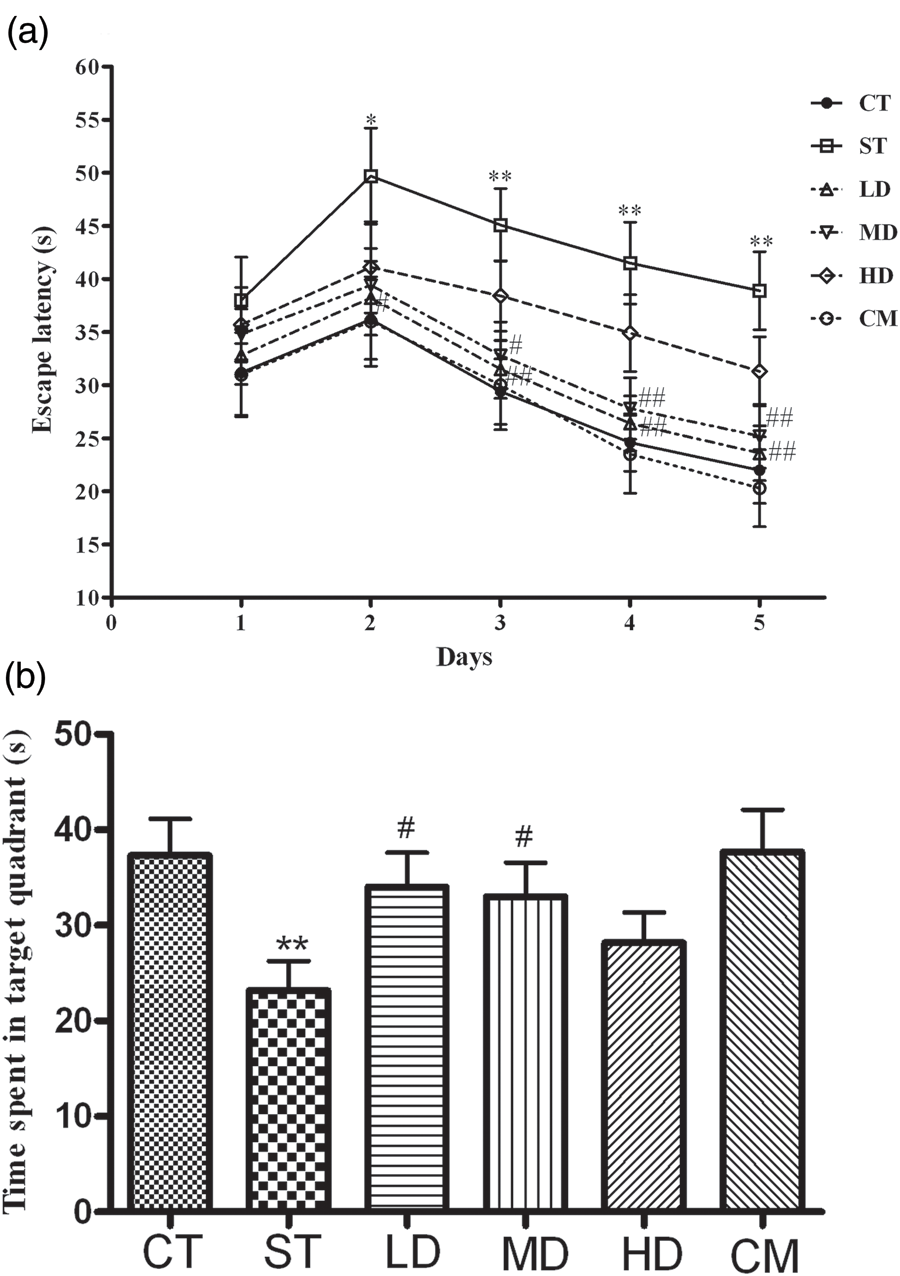
To assess the consolidation of memory, the mice were subjected to a probe trial on day 5. During the probe trial, the time of mice staying in the platform quadrant was determined (Figure 3b). A significantly shorter stay was observed in the ST group, which was significantly reversed by LD and MD treatment (P < 0.05). In contrast, no significant difference was found in the time staying in the platform quadrant of HD treatment mice when compared with the ST group (P > 0.05).
Oxidative alterations in prefrontal cortex and hippocampus
Changes of MDA, SOD, GSH and CAT in prefrontal cortex of mice
MDA, malondialdehyde; SOD, superoxide dismutase; GSH, glutathione; CAT, catalase; CT, control group; ST, stress group; LD, low dose; MD, medium dose; HD, high dose; CM, control + medium dose
Mice were divided into six groups (n = 10). CT group and ST group were given daily with 0.5% sodium carboxymethyl cellulose for three consecutive weeks. Three stress and luteolin modulation groups were administered daily with low, medium and high doses of luteolin, respectively. CM group received medium dose of luteolin without restraint stress treatments. All stress groups received restraint stress for three consecutive weeks. At the end of test, mice were sacrificed and the oxidative markers were measured. Data are expressed as mean ± SEM determined by one-way analysis of variance (ANOVA)
*P < 0.05, **P < 0.001 versus CT group
# P < 0.05, ## P < 0.01 versus ST group
Changes of MDA, SOD, GSH and CAT in hippocampus of mice
MDA, malondialdehyde; SOD, superoxide dismutase; GSH, glutathione; CAT, catalase; CT, control group; ST, stress group; LD, low dose; MD, medium dose; HD, high dose; CM, control + medium dose
Data are expressed as mean ± SEM determined by one-way analysis of variance (ANOVA)
*P < 0.05, **P < 0.001 versus CT group
# P < 0.05, ## P < 0.01 versus ST group
Concentrations of serum corticosterone
Levels of corticosteronel, norepinephrine and dopamine in serum of mice
CT, control group; ST, stress group; LD, low dose; MD, medium dose; HD, high dose; CM, control + medium dose
Data are expressed as mean ± SEM determined by one-way analysis of variance (ANOVA)
*P < 0.01, **P < 0.001 versus CT group
# P < 0.05, ## P < 0.01 versus ST group
Concentrations of serum norepinephrine and dopamine
Compared with the CT group, mice exposed to the psychological stress exhibited significant decline in both the serum norepinephrine and dopamine concentrations (P < 0.01 or P < 0.001). Additionally, the concentration of norepinephrine was increased in the LD group in contrast to that of the SD group (P < 0.05). Furthermore, the serum dopamine concentrations were observed to be significantly higher in LD and HD groups when compared with the CT group (P < 0.05). However, luteolin did not alter the levels of norepinephrine and dopamine in the serum of mice without restraint stress treatments (P > 0.05, Table 3).
Discussion
Nowadays, there are growing evidences that psychological stress is related to many diseases and increasing attentions were paid to the prevention of stress-induced injury. Flavonoids are a class of polyphenolic compounds which display a variety of biologic activities. The neuroprotective activity of flavonoids is well reported and dietary supplementation studies, in human and animals, using flavonoid-rich plant or food extracts have highlighted their potentials to influence cognition and learning. 18–20 In the present study, mice were subjected to chronic restraint stress to emulate the psychological stress in daily life. Our research revealed that luteolin treatment can reverse impaired cognition induced by psychological stress in mice. In addition to the maintenance of cognitive function, a normalizing of serum corticosterone, norepinephrine and dopamine concentrations, as well as an attenuation of oxidative damage in prefrontal cortex and hippocampus were also seen in mice that were treated with luteolin.
Corticosterone concentrations could reflect the body's stress intensity. 21,22 A significant increase of corticosterone concentrations was observed after acute stress. 23 Otherwise, previous research has indicated that corticosterone concentrations were elevated during the first seven days in the stressful environment, but returned to basal levels by day 14. 24 However, growing evidence showed that animals chronically exposed to chronic stress showed a depression-like dysfunction of hypothalamic–pituitary–adrenal (HPA) axis feedback, associated with adrenal hypertrophy and elevated corticosterone secretion. 25,26 We speculated that the different responses of corticosterone concentrations to chronic stress might be associated with the intense intensities and the animal species. Additionally, many studies inferred that elevated corticosterone concentrations contributed to the pathogenic impairments induced by stress. 2,5 In the present study, a significant increase of corticosterone concentration in serum was observed, which was consistent with the hypothesis that serum corticosterone was up regulated by psychological stress treatment. 5,26 Furthermore, our data showed that the performances of psychological stress mice in the open-field test, step-through test and MWM test were all changed abnormally. Also, appropriate doses of luteolin modulations could improve these behavioral disturbances induced by stress. Therefore, our data suggested that luteolin was able to attenuate the psychological stress-induced learning and memory deficits which might be associated with preventing the increase of corticosterone concentrations in mice.
In addition, psychological stress also activated SNS, and then catecholamines were released from the sympathetic nerve terminal and the adrenal medulla. 27,28 Catecholamines consist of epinephrine, norepinephrine and dopamine, which are involved in the modulation of body's cognition, awareness, attention, emotional state and other psychoactivities. In case of the intensity and duration of stress overtopping the body's copping capacity, dysfunction of these catecholamines results due to the prolonged stressful conditions associated with a wide range of behavioral disturbances. 5 In this study, a significant reduction was observed in the levels of catecholamines, including norepinephrine and dopamine in animals subjected to psychological stress. Therefore, our study indicated that the cognitive impairments induced by psychological stress might be related to the declines of norepinephrine and dopamine, which were consistent with previous research. 29,30 Moreover, evidence reveals that luteolin has an inhibitory activity against monoamine oxidase (MAO) enzyme. 31 In the present study, luteolin exhibited considerable reversal effects on the psychological stress-induced variations in the values of norepinephrine, dopamine as shown by their quantification. Consequently, luteolin treatment could attenuate the cognitive impairments, which might be related with its normalizing effect on the serum catecholamines concentrations.
Luteolin had been shown to prevent various neuronal damages induced by oxidative stress and toxic substances. Previous research suggested that luteolin could obviously attenuate scopolamine hydrobromide (SCOP)-induced amnesia. 14 In other studies, luteolin consumption enhanced spatial working memory by mitigating microglial-associated inflammation in the hippocampus of aged mice. 8 Also, luteolin showed protective effect on learning and memory impairments induced by cerebral hypoperfusion. 32 Moreover, in vitro studies also revealed that luteolin has neuroprotective effects on neuronal damage induced by toxic substances. 23,33 N-methyl-4-phenyl-pyridinium (MPTP) was commonly used as a neurotoxic drug to produce experimental animals or an in vitro model of Parkinson disease. In MPTP-injured PC12 and C6 cells, luteolin can enhance the oxidative stress defense and maintain the cellular redox homeostasis, in an ERK-dependent way. 23 In addition, luteolin showed a dose-dependent protective effect on primary cultured rat cortical neurons injury induced by Amyloid β-25–35. 33 Psychological stress could induce a series of dysfunctions of central nervous system, such as cognitive impairment, aberrations in the HPA system, brain tissue damage and neurotransmitter levels changes. 34 In the present study, our results demonstrated that the cognitive impairments induced by psychological stress were improved by luteolin treatment in mice.
Furthermore, psychological stress caused conspicuous oxidative stress which damaged lipid membranes, proteins and nucleic acids, accordingly disturbed the related biological activities. Otherwise, prefrontal cortex, as well as hippocampus, is structure within the brain that plays key roles in memory, attention, thought, language and consciousness. But they were more susceptible to free radical damage, compared with other tissues. 35,36 In the present study, the oxidative modifications in prefrotnal cortex and hippocampus were investigated. Our results showed a significant elevation of lipid peroxidation markers (MDA) in the prefrontal cortex and hippocampus of the ST group induced by psychological stress, which was in agreement with previous reports, 5,34 indicating a lipid peroxidation in brain tissues caused by psychological stress. Additionally, psychological stress significantly decreased the SOD and CAT activities, as well as GSH concentration in prefrontal cortex and hippocampus of stressed mice. These results indicated that the delicate balance between oxidative species and antioxidative defenses was disturbed in prefrontal cortex and hippocampus of mice exposure to psychological stress treatment. However, these oxidative alterations were normalized in mice by repeated treatment with luteolin. Indeed, luteolin has been demonstrated to be a free radical scavenger and may effectively minimize lipid peroxidation in brain tissues. 37,38 In the present study, repeated administration of luteolin for three weeks significantly prevented the decrease of antioxidant capacity and the increase of lipid peroxidation in the prefrontal cortex and hippocampus induced by psychological stress. Collectively, these results are supportive of the hypothesis that luteolin could inhibit the psychological stress-induced prooxidant–antioxidant disequilibrium.
In conclusion, the current study suggested that chronic administration of luteolin results in an improvement on cognitive performances in mice exposure to psychological stress. The related mechanisms under these actions might be attributed to the prevention of oxidative damage in the prefrontal cortex and hippocampus, as well as the attenuating effect on the hypersecretion of corticosterone and hyposecretion of catecholamines induced by psychological stress.
Footnotes
ACKNOWLEDGEMENT
This work was supported by the ‘Qing-Lan’ Project of Jiangsu Province, ‘Liu Da Ren Cai Gao Feng’ Project of Jiangsu Province (2011-YY-13), the Industrialization of Scientific Research Promotion Projects of Universities and Colleges in Jiangsu Province (2011–2016), the Natural Science Fund for Universities and Colleges in Jiangsu Province (11KJB350005), The foundation of School of Pharmacy in Xuzhou Medical College (2011YKJ004), The Foundation of Xuzhou Medical College Key Laboratory of Tumor Biology Therapy (C0904, JSBL0803, C0903), the Science and Technology Plan Projects of Xuzhou (XF11C037; XF11C062, XF11C065; XZZD1227; XZZD1219), Superiority Academic Discipline Construction Project of Jiangsu Higher Education Institutions.