
Abstract
Triple negative breast cancer (TNBC) represents approximately 15% of the newly diagnosed cancers worldwide and is characterized by tissue lacking in estrogen, progesterone and human epidermal growth factor receptors. TNBC disproportionately affects younger women and women of colour, and new treatments are needed. The opioid growth factor (OGF) – opioid growth factor receptor (OGFr) axis is a determinant of cell proliferation in neoplasia, and OGF is an endogenously produced pentapeptide that inhibits cell replication by interacting with OGFr and upregulating cyclin-dependent inhibitory kinase pathways thus reducing DNA synthesis. In these studies we investigated the presence and function of the OGF-OGFr axis in two human TNBC cell lines, as well as in breast cancer cell lines containing hormonal receptors. TNBC cell lines MDA-MD-231 and BT-20, as well as human breast cancer cells SK-BR-3 and MCF-7, were examined for the presence of pentapeptide and receptors, as well as their response to OGF. Specificity of peptide and receptor was confirmed by antibody neutralization and molecular studies to knockdown classical receptor protein. The requirement for protein transcription and translation and RNA transcription were investigated. Growth of TNBC cells in the presence of OGF and standard of care chemotherapeutic agent paclitaxel was evaluated to determine both efficacy and protective effects against toxicity. OGF treatment inhibited TNBC cells in a dosage related, receptor mediated, and reversible manner. OGF was the specific endogenous opioid to inhibit cell proliferation, and this was mediated by p21 cyclin dependent inhibitory kinase pathways, and required protein and RNA synthesis. OGFr was the specific receptor involved; both peptide and receptor were detected in all four cell lines. OGF treatment inhibited growth of all cancer cell lines evaluated, and reduced cell death in cultures exposed to paclitaxel. The OGF-OGFr axis is present and functioning in TNBC cell lines, and provides a novel biological pathway as potential therapy.
Keywords
Introduction
Over 225,000 new cases of invasive breast cancer are reported each year in the United States. 1 Approximately 15% of globally diagnosed breast cancer is designated as triple negative breast cancer (TNBC). TNBC lacks estrogen receptors (ER), progesterone receptors (PR), and the human epidermal growth factor receptor (HER2). The prognosis for women with breast cancer is improving each year, with nearly 98% survival over a five-year period of time if the cancer is localized and diagnosed early. 2 Successful treatment increases with the presence of ER, PR and HER2. TNBC tends to occur in younger women, women of colour, and those from lower socioeconomic countries. To improve the outcome of TNBC we have studied a novel biological pathway present in a wide variety of cancers 3 which may provide knowledge on the progression of TNBC and enable development of novel targets and therapies.
Although endogenous opioid systems are known to regulate cancer proliferation,3–9 little is known about the role of the opioid growth factor (OGF) – opioid growth factor receptor (OGFr) axis and proliferation of TNBC cells. OGF, chemically termed [Met 5 ]-enkephalin, is a constitutively-expressed pentapeptide that inhibits proliferation by stimulating the cyclin-dependent inhibitory kinase pathway and slowing the transition between the G1 and S phases of the cell cycle. Zagon et al. 3 have recently reported that the OGF-OGFr axis is important in MDA-MB-231 and MCF-7 human breast cancer cell lines TNBC, with no changes in the growth of these neoplasias observed with the addition of OGF if OGFr is not available.
Attenuation of the OGF-OGFr axis in human cancer cell lines occurs by disruption of OGF-OGFr interfacing through continuous exposure to an opioid antagonists, neutralization of endogenous opioids, or knockdown of the OGFr stimulated DNA synthesis.3–5 Enhanced growth inhibition through the OGF-OGFr pathway has been documented following exogenous OGF or genetic manipulation to overexpress OGFr in order to repress cell division.4,7 Given that TNBC is an aggressively growing breast cancer, the present study investigated the presence and efficacy of OGF in the inhibition of cell growth. Our research on the OGF-OGFr axis in TNBC was also extended to human breast cancer cell lines expressing one or more of the targeted receptors. These data demonstrate that OGF is a non-toxic, biological therapy for the treatment of TNBC.
Materials and methods
Cell culture
Human breast cancer cell lines MDA-MB-231, BT-20, MCF-7, and SKBR3 were obtained from the American Type Culture Collection (Manassas, VA). The TNBC cell line MDA-MD-231 was grown in Leibovits L-15 media, whereas BT-20 cells were cultured in modified Eagle’s medium. The MCF-7 cell line that demonstrates estrogen and progesterone receptors was maintained in modified Eagle's media, whereas the SK-BR-3 cells that have only the herceptin receptor were grown in RPMI 1640 complete media. All media were supplemented with 10% foetal calf serum and contained antibiotics (5000 units/mL penicillin, 5 µg/mL streptomycin). Cells were grown in a humidified atmosphere of 5% CO2/95% air at 37℃. The MDA-MB-231 cells were obtained from a 44-year-old black female with medullary carcinoma, 10 whereas BT-20 cells were obtained from a 74-year-old Caucasian female. 11 The MCF-7 cell line was acquired from a metastatic site in a 69-year-old Caucasian woman, 12 while the SK-BR-3 cell line was isolated from pleural effusion of breast adenocarcinoma in a 43-year-old Caucasian female. 13
Immunocytochemistry
Log phase human breast cancer cell lines MDA-MB-231, BT-20, MCF-7 and SKBR3 were seeded on 22 × 22 mm cover slips and 72 h later, fixed and stained with anti-OGF (CO-172) and anti-OGFr (BO-344) polyclonal antibodies.14,15 Polyclonal antibodies to OGF and OGFr were generated in the laboratory and have been characterized.14,15 Cells incubated with secondary antibody only were considered controls. Stained cells were examined using an Olympus IX-81 epifluorescent microscope.
OGFr binding assays
Nuclear-enriched fractions of log-phase cultures of MDA-MB-231 cells were evaluated for OGFr binding characteristics using standard binding assays 16 and custom-synthesized [H3]-[Met 5 ]-enkephalin (Perkin Elmer, Waltham, MA; 52.7 Ci/mmol). Three independent binding assays were performed.
Cell growth
For most assays, the triple negative cell line MDA-MB-231 was utilized. Cells were seeded in triplicate wells and treated 24 h later with compounds or untreated (control). Unless otherwise stated, media and compounds were replaced daily. All drugs were prepared in sterile water and dilutions represent final concentrations. Paclitaxel was initially dissolved in DMSO and further diluted in sterile water. At designated times, cells were harvested and counted by a hemacytometer using the trypan blue exclusion methodology. Two counts per aliquot of two aliquots per well, and two or more wells per treatment per time point were counted. Two to five independent experiments were conducted for each assay.
Specificity of endogenous OGF
The specificity of endogenous OGF for cell growth was determined by treating cells with a polyclonal antibody against OGF (1:200; CO172); rabbit serum (1:200) served as a control. Serum and media were changed daily, and cells were counted after 72 h of treatment.
Specificity of OGFr: knockdown with OGFr siRNA
OGFr targeted siRNAs (antisense 5′-uagaaacucagguuuggcg-3′; sense 5′-cgccaaaccugaguuucua-3′) were designed and obtained as a ready-annealed, purified duplex probe from Ambion (Austin, TX); scrambled siRNA (Ambion) served as a control.3,4 Classical mu, delta, and kappa opioid receptor siRNAs were purchased from Santa Cruz. Cells (2.5 × 104 cells/well) were seeded in 24-well plates containing 0.5 mL of media without antibiotics and transfected with 20 nM siRNAs for each opioid receptor, OGFr, or scrambled sequence. Cells were incubated for 4 h at 37℃ prior to adding OGF (10−6 M) or NTX (10−6 M). Cultures then were incubated for an additional 20 h, followed by the replacement of fresh complete media with or without OGF or NTX. Seventy-two hours later, cells were harvested, and either counted or protein isolated for Western blot analysis. 4 Proteins were stained with 1:200 dilutions of anti-OGFr polyclonal antibody (BO344), or mu (sc-15,310, Santa Cruz Biotechnology, Santa Cruz, CA), delta (sc-9111, Santa Cruz) or kappa (sc-9112, Santa Cruz) opioid receptor antibodies.
OGF’s mechanism of action: cell cycle inhibition following siRNA knockdown
OGF is known to act through cyclin-dependent inhibitory kinase pathways by regulation of p16 and/or p21.17,18 To assess the role of p21 as a mediator of the inhibitory effects of OGF, siRNA knockdown of p21 was conducted. Log-phase cells were transfected with 20 nM p21 siRNA (Santa Cruz), or scrambled siRNA (Ambion) using Oligofectamine (Life Technologies, Grand Island, NY) in serum and antibiotic free media for 4 h at 37℃ before the addition of OGF (10−6 M). Cultures were incubated an additional 20 h before the addition of fresh complete media with or without OGF 10−6 M. OGF and media were changed daily. At 72 h after transfection, cells were collected and counted, or processed for Western blot analysis to determine expression levels of p21 protein.
To assess protein expression, cells were harvested with 0.25% trypsin-EDTA (Mediatech, Herndon, VA) and subjected to 15% SDS-PAGE analysis. 4 Blots were probed with a p21 monoclonal antibody (ab-7960, 1:200; Abcam, Cambridge, MA) followed by anti-rabbit horseradish peroxidase conjugated secondary antibody (AP307P,1:3000; Merck Millipore, Billerica, MA), and developed using a chemiluminescence Western blotting detection system. To determine equal loading of total protein, blots were stripped with buffer and reprobed with anti-actin antibody. The optical density of each band was determined by densitometry, and each value was normalized to actin. At least two independent experiments were conducted.
OGF action and protein and RNA synthesis
To examine whether OGF’s inhibitory and NTX’s stimulatory effects are dependent on protein and RNA synthesis, log-phase MDA-MB-231 cells were treated with 5 µg/mL concentrations of cycloheximide or puromycin for protein synthesis or actinomycin D for RNA synthesis, followed by treatment with 10−6 M concentrations of OGF or NTX, and 30 µM BrdU for 3 h. Cell cultures were rinsed and fixed in 10% formalin and stained with antibodies to BrdU. At least 1000 cells on three coverglasses/treatment were counted for BrdU incorporation, and labelling indexes (number of positive cells divided by total cells) were calculated.
Chemicals
The following compounds were obtained from the indicated sources: [Met 5 ]-enkephalin, [Leu 5 ]-enkephalin, [D-Pen2,5]-enkephalin (DPDPE), [D-Ala 2 , MePhe 4 , Glyol 5 ]-enkephalin (DAMGO), β endorphin, naltrexone (NTX), naloxone (NAL), dynorphin A1-8, morphine sulphate, endomorphin 1, endomorphin 2, paclitaxel (Sigma-Aldrich, St. Louis, MO), and U69,583 (Upjohn Diagnostics, Kalamazoo, MI).
Statistical analysis
All data were analysed by GraphPad Prism software using two-way or one-way ANOVA where appropriate, with Newman–Keuls post hoc tests. In some cases, independent Student’s t-tests were applied to the data.
Results
OGF and OGFr are present in human breast cancer cells
Immunohistochemical studies revealed OGF and OGFr in the cytoplasm and nucleus of MDA-MB-231 cells (Figure 1A). Staining was not observed in cells processed with secondary antibody only. Receptor binding assays of nuclear protein from log-phase MDA-MD-231 cells revealed specific and saturable binding with a one-site model of binding. The binding capacity (Bmax) of MDA-MB-231 was 8.5 ± 1.8, with a binding affinity (Kd) of 4.1 ± 1.1 nM.
The presence and function of the OGF-OGFr axis in human triple negative breast cancer cells. (A) The presence of OGF (Co172 1:200) and OGFr (BO344 1:200) in log phase MDA-MB-231 human triple negative breast cancer cells as detected by immunocytochemistry. Cultures were stained with antibodies to OGF or OGFr; cells stained with secondary antibody only served as control (Inset). Bar = 20 µm. (B–E) OGF inhibited growth of MDA-MB-231 cells in a dose-related (B), time-dependent (C), receptor-mediated (D), and reversible manner (E). Cells were treated as described in the Methods. (F) MDA-MB-231 cultures were seeded and treated with 10−6 M naltrexone (HDN) or 10−9 M naltrexone (LDN), 10−6 M OGF, or sterile water. Cells were counted at 24, 48, and 72 h. Data represent means ± SEM for triplicate samples; all experiments were conducted 2–4 times. Significantly different from sterile-water treated controls at the corresponding time point at *P < 0.05, **P < 0.01, or ***P < 0.001as determined by ANOVA with Newman–Keuls post-hoc tests. (A color version of this figure is available in the online journal)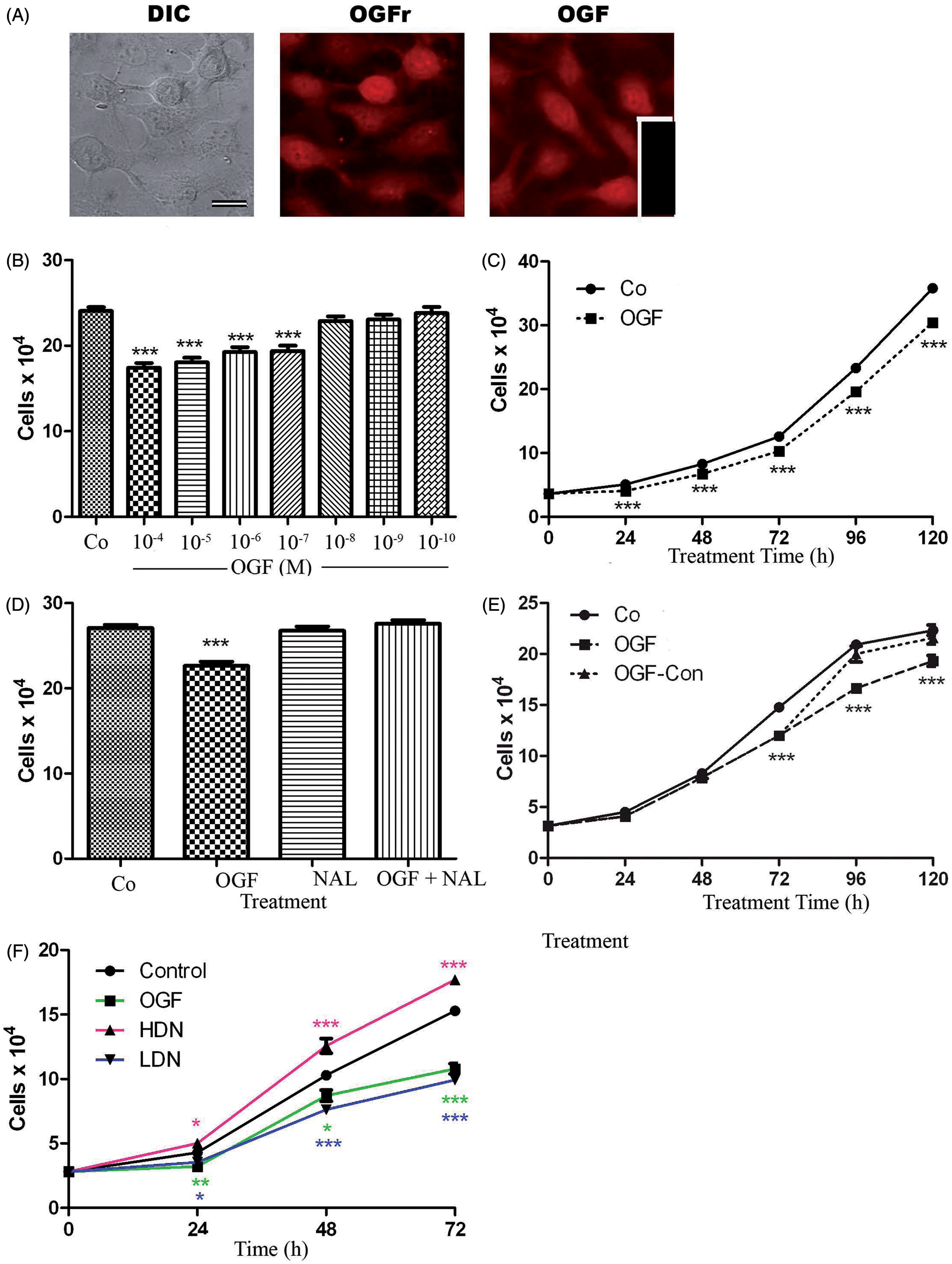
OGF depresses growth of human breast cancer cells
The effects of OGF in concentrations ranging from 10−4 to 10−10 M on growth at 72 h revealed a dose-dependent inhibitory effect on cell proliferation (Figure 1B). The addition of OGF in concentrations from 10−4 M to 10−7 M reduced cell number of MDA-MB-231 cultures by 19–28% relative to controls. Growth curves of cells treated with OGF (10−6 M) revealed that cell number was repressed by 20% relative to untreated cultures within 24 h. Over a 120 h period of time, cell number was reduced by 15–19% relative to controls (Figure 1C). The doubling times for MDA-MD-231 cells were calculated to be ∼34 h, whereas MDA-MD-231 cells grown in the presence of OGF had a doubling time of 37.5 h. To determine whether OGF activity was mediated by an opioid receptor, MDA-MB-231 cells were grown in the presence of 10−6 M concentrations of OGF, OGF and a short-acting, non-selective opioid receptor antagonist, naloxone (NAL), or NAL alone; control cultures were untreated. Cell number was reduced by 16% in cultures treated with OGF compared with controls, whereas cultures receiving both OGF and NAL, or NAL alone, were not significantly different from the controls (Figure 1D). The inhibitory action of OGF was reversible (Figure 1E), with cultures exposed to OGF (10−6 M) for 72 h were decreased in cell number by 18% relative to controls. If media were removed at 72 h and fresh media were added with no OGF (OGF-Co), cell number returned to control levels within 48 h of removal of OGF; OGF-Co cell cultures increased 12–20% at 96 and 120 h from initial seeding. However, cultures maintained on 10−6 M concentration of OGF exhibited 20% and 14% fewer cells than control cultures.
MDA-MB-231 cells are responsive in growth to continuous or intermittent blockade of the OGF-OGFr axis
Log phase cells were subjected to 10−6 M concentrations of the potent long-acting opioid receptor antagonist NTX. An intermittent opioid receptor blockade with naltrexone (10 −6 M) was invoked by adding naltrexone to cultures for only 4 h after which time the media was removed, cells were washed and fresh media replaced on a daily basis. This intermediate blockade was performed daily for three days after which time cells were counted. Intermittent receptor blockade invoked by short-term exposure to NTX resulted in a 17.5% reduction in cell number within 24 h, and a 26% and 35% reduction at 48 h and 72 h, respectively, relative to controls. (Figure 1F). Continuous opioid receptor blockade by 10−6 M NTX (media and compound replaced daily) resulted in an increase in breast cancer cell number within 24 h, with cultures exposed to naltrexone (HDN) having 16% and 22% more cells than control cultures at 24 and 48 h (Figure 1F).
OGF is the specific endogenous opioid agonist related to growth inhibition of human breast cancer cells
To determine whether other endogenous or exogenous opioids are capable of modulating the growth of human breast cancer cells, MDA-MB-231 cultures were treated daily for three days with 10−6 M concentrations of natural (DPDPE, β-endorphin, endomorphin-1, and endomorphin-2) or exogenous (ethylketocyclazocine EKC, U69593, morphine, and DAMGO) opioid-related compounds, many of which are specific for mu, delta, and kappa opioid receptors (Figure 2A). Under growth conditions that demonstrated a 32% reduction in cell number following OGF treatment, all other opioid-related compounds had no effect on cell proliferation. Exposure to daily treatment with naltrexone increased cell number by 28% relative to controls at 72 h.
OGF is the specific opioid agonist that inhibits the growth of log-phase triple negative breast cancer cells. (A) Cultures of MDA-MB-231 cells were treated for 72 h with various endogenous opioids, the opioid antagonist naltrexone (NTX) (10−6 M), or were untreated controls (Co). (B) Endogenous OGF was neutralized by anti-OGF antibody Co172. Cells were treated with a polyclonal antibody specific for OGF, pre-immune serum (IgG), exogenous OGF (10−6 M), or untreated controls (Co) for 72 h. Media and compounds were replaced daily. Data represent means ± SEM for two aliquots/well and at least 2 wells/group. Experiments were performed twice. Significantly different from controls at *P < 0.05 or ***P < 0.001as determined by ANOVA with Newman–Keuls post hoc analyses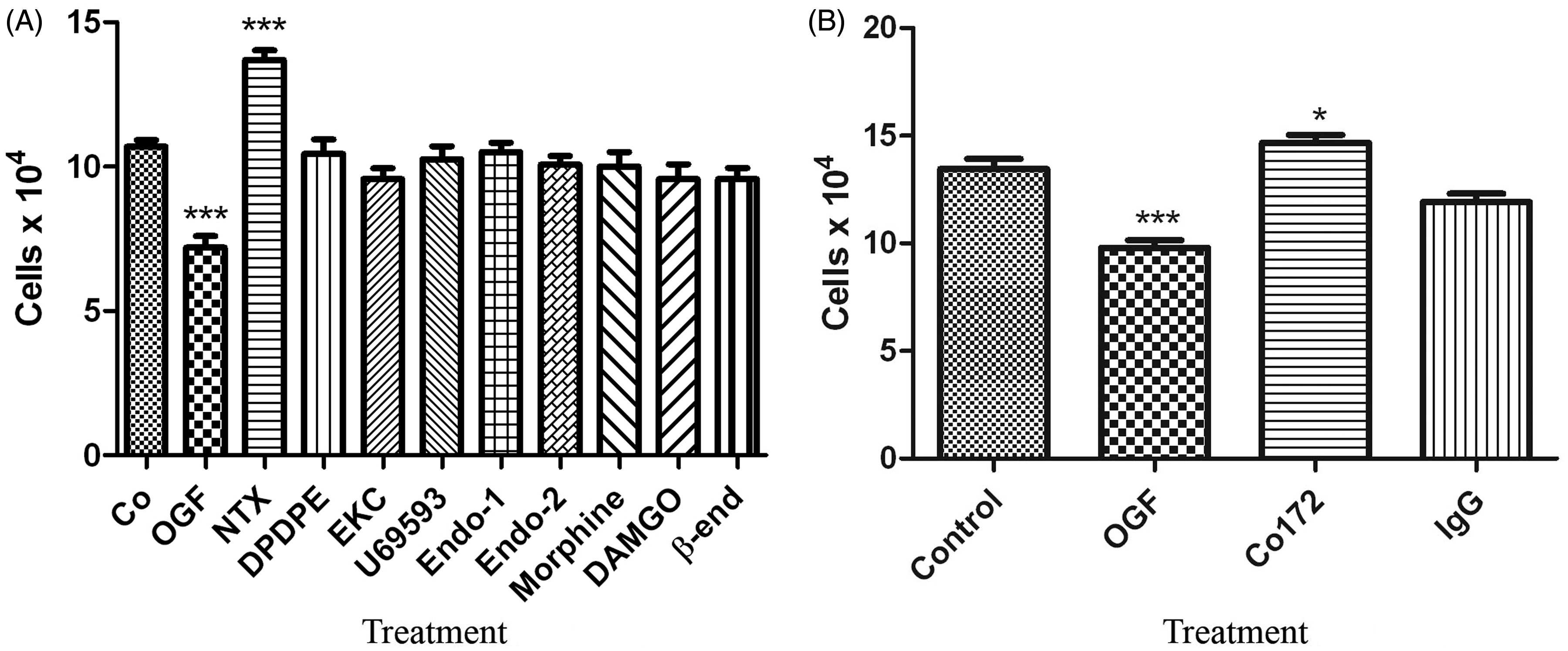
The selectivity of OGF’s inhibitory action was investigated by neutralizing endogenous OGF with the addition of OGF polyclonal antibody. Cultures exposed to the OGF antibody had 10% more cells than untreated controls or cultures treated with control rabbit IgG serum (Figure 2B).
The specific opioid receptor that mediates the inhibitory effects of OGF is OGFr
Opioid receptor specificity was evaluated by knocking down functional opioid receptors using siRNAs specific for classical mu, delta, and kappa opioid receptors and assessing cell proliferation in the presence of OGF or NTX (Figure 3). MDA-MB-231 cells transfected with OGFr siRNA had 40% more cells compared to untreated control cells after 72 h of transfection (Figure 3A). The addition of exogenous OGF or NTX had no inhibitory or additional stimulatory effects, respectively, on cells transfected with OGFr siRNA when compared to untreated cultures transfected with OGFr siRNA. Cells transfected with mu opioid receptor siRNA (Figure 3B) exhibited no significant changes in cell number, and the addition of OGF inhibited growth 15–23% in untransfected, transfected, and scrambled siRNA-transfected cultures in a comparable manner. NTX stimulated growth 56% in cultures transfected with mu opioid receptor siRNAs, relative to controls. Likewise, cultures transfected with delta opioid receptor siRNA (Figure 3C) or kappa opioid receptor siRNA (Figure 3D) were not altered in their growth-related responses to either sterile water, OGF, or NTX in comparison to untransfected cells or those modified by scrambled siRNAs.
The specific opioid receptor in triple negative breast cancer cells that is related to growth regulation by OGF is OGFr. (A-D) MDA-MB-231 cells (50,000) were seeded in 6-well plates and transfected for 24 h with OGFr siRNA (A), mu opioid receptor siRNA (B), delta opioid receptor siRNA (C), or kappa opioid receptor siRNA (D); other cultures were transfected with a commercially available scrambled siRNA and served as controls. In some cultures, OGF (10−6 M) or NTX (10−6 M) were added daily for 48 h and counted at 72 h after transfection. Data represent means ± SEM for at least 2 aliquots per well from 3 wells per group, and were analyzed using ANOVA with subsequent planned comparisons by Newman-Keuls tests. Significantly different from respective controls at **P < 0.01 or ***P < 0.001. (E-H) Cultures were harvested 48 h post-transfection, and protein isolated for western blots to determine the percentage of opioid receptor knockdown. Values represent densitometric analyses from at least 3 blots. Significantly different from wildtype values at **P < 0.01 and ***P < 0.001, and from scrambled siRNA cultures at +P < 0.05 as determined by the independent student t-test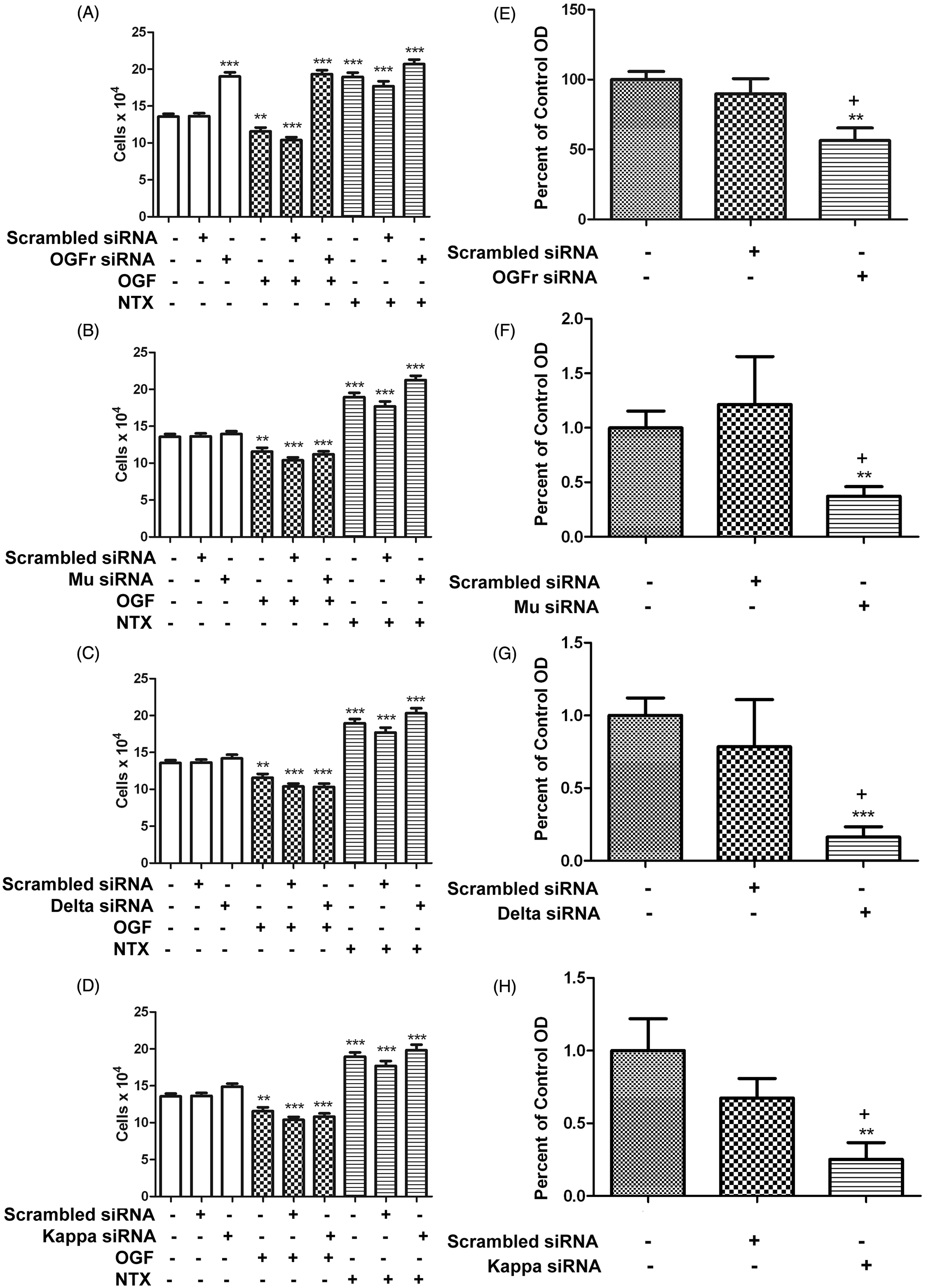
The percentage of knockdown for each siRNA transfection experiment is presented in Figure 3E–H. Based on Western blot analysis, protein expression for the OGFr, mu, delta, and kappa opioid receptors were reduced by 50%, 60%, 75%, and 80%, respectively.
OGF inhibits cell number by decreasing DNA synthesis in MDA-MB-231 cells
To evaluate the mechanism by which OGF inhibits human breast cancer cell growth, DNA synthesis of MDA-MB-231 cultures exposed to OGF or NTX was measured (Figure 4). Relative to untreated controls, the percentage of BrdU labelled cells in cultures treated with OGF (10−6 M) for 3 h was decreased by 23%, while cells treated with NTX (10−6 M) increased by 25%. Moreover, the inhibitory effects of OGF on cell proliferation were abolished in cultures treated with puromycin, cyclohexamide, or actinomycin D which block RNA transcription, RNA translation, as well as protein synthesis (Figure 4A–C). In contrast, the opioid antagonist NTX accelerated DNA synthesis and exerted this action in the presence of cyclohexamide, puromycin, or actinomycin D (Figure 4A–C), suggesting that NTX action is independent of RNA and protein translation.
The inhibitory action of OGF on DNA synthesis is dependent on RNA and protein synthesis. The stimulatory action of NTX on DNA synthesis is independent of RNA and protein synthesis. MBA-MD-231 cells were seeded on 22 mm coverslips and treated for 3 h with OGF (10−6M) or NTX (10−6), and 30 µM BrdU, along with 5 µg/mL concentrations of puromycin (A), cyclohexamide (B) or actinomycin D (C) to measure RNA transcription, protein translation, and RNA translation, respectively. Data represent the percent of BrdU-positive cells (mean ± SEM) and were analyzed using ANOVA and Newman-Keuls post-hoc analyses. Significantly different from control treated cells at **P < 0.01 or ***P < 0.001; significantly different from OGF treated cultures at ^^P < 0.01or ^^^P < 0.001, and from cells treated with water and puromycin, cyclohexamide, or actinomycin D at ++P<0.01 or +++P < 0.001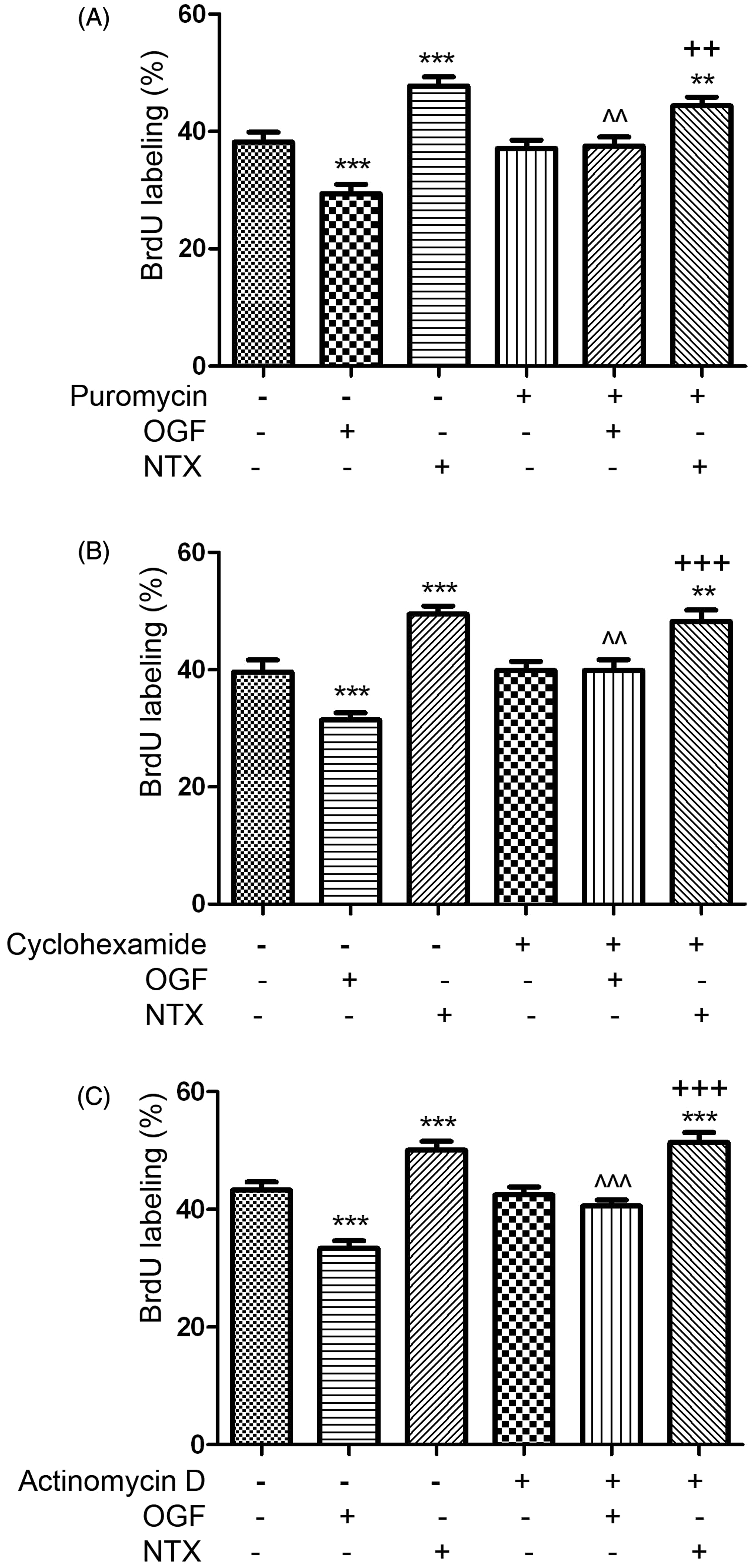
OGF action is mediated by the cyclin-dependent kinase pathway
The OGF-OGFr axis mediates its effects through regulation of the cell cycle. Specifically, OGF upregulates cyclin-dependent inhibitory kinase pathways which regulate downstream retinoblastoma regulation of transition between G1 and S phases of the cell cycle. Both p16 and p21 CKIs are present in human cancer cell lines and either one is effective at regulation of the cell cycle by OGF. Human breast cancer cells MDA-MB-231 have only p21. Cells transfected with p21siRNAs demonstrated approximately 75% knockdown of the protein (Figure 5B, C) and treatment of the transfected cells demonstrated that this reduction in p21 nullified the inhibitory effects of OGF (Figure 5A).
The cell cycle pathway utilized by OGF to repress cell division of human MDA-MB-231 cells. (A) Number of cells in MDA-MB-231 cultures transfected with p21 siRNA or scrambled control siRNA and treated with OGF (10−6 M); cell number was quantitated 72 h later using 2 samples/well and 3 wells per condition. Values represent means ± SEM as determined by ANOVA with Newman–Keuls post-hoc analyses. Significantly different from untreated controls (or scrambled siRNAs) at ***P < 0.001. (B, C) Western blot validation of p21 siRNA knockdown of p21 protein expression. Histograms represent means ± SEM for densitometric values from 3 blots; significantly different from untreated control protein samples at ** P < 0.01. (A color version of this figure is available in the online journal)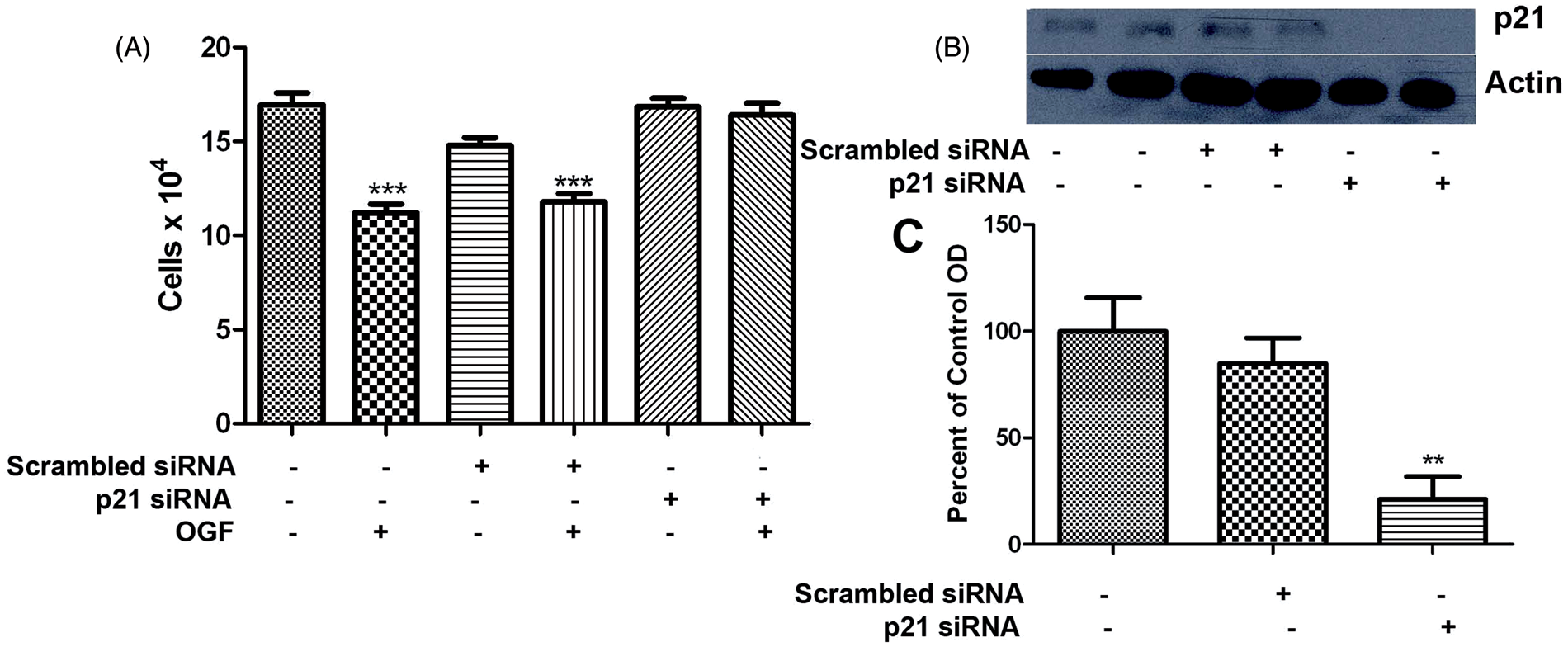
OGF combined with taxol reduces the toxicity of taxol treatment
One of the standard chemotherapies for breast cancer is paclitaxel (taxol). However, paclitaxel’s efficacy is based on the fact that it kills cells and is very toxic to the patient. Addition of OGF to taxol treatment did not further reduce cell number of MDA-MB-231 cells (Figure 6A,B), but did in fact, diminish cell death following taxol treatment at 10−7 M or 10−8 M (Figure 6C). Whereas treatment with 10−8 M taxol alone resulted in greater than 30% dead cells, the cell death in cultures treated with OGF and 10−8 M taxol combined was reduced to 14%, a significant reduction of 60%.
Combination therapy of OGF and/or the standard of care, paclitaxel (Taxol) in MDA-MB-231. (A) Cells were treated with OGF (10−6 M) and/or Taxol (10−7 M) over a period of 96 h. Cell number was significantly repressed by addition of Taxol. To determine whether OGF conferred any protection against the toxicity of taxol, or whether OGF in combination with a lower dosage of taxol was effective, cell number (B) and percent dead cells (C) were reported for cultures treated with taxol at concentrations of 10−7 M and 10−8 M, and combination taxol and OGF (10−6 M). Values represent means ± SEM for 2 aliquots from 2 wells/treatment group. Significantly different from control at P < 0.01 (**) and P < 0.001 (***), and from the respective taxol treatment alone at p < 0.01 (++) as determined by ANOVA with subsequent comparisons made using Newman–Keuls tests. (A color version of this figure is available in the online journal)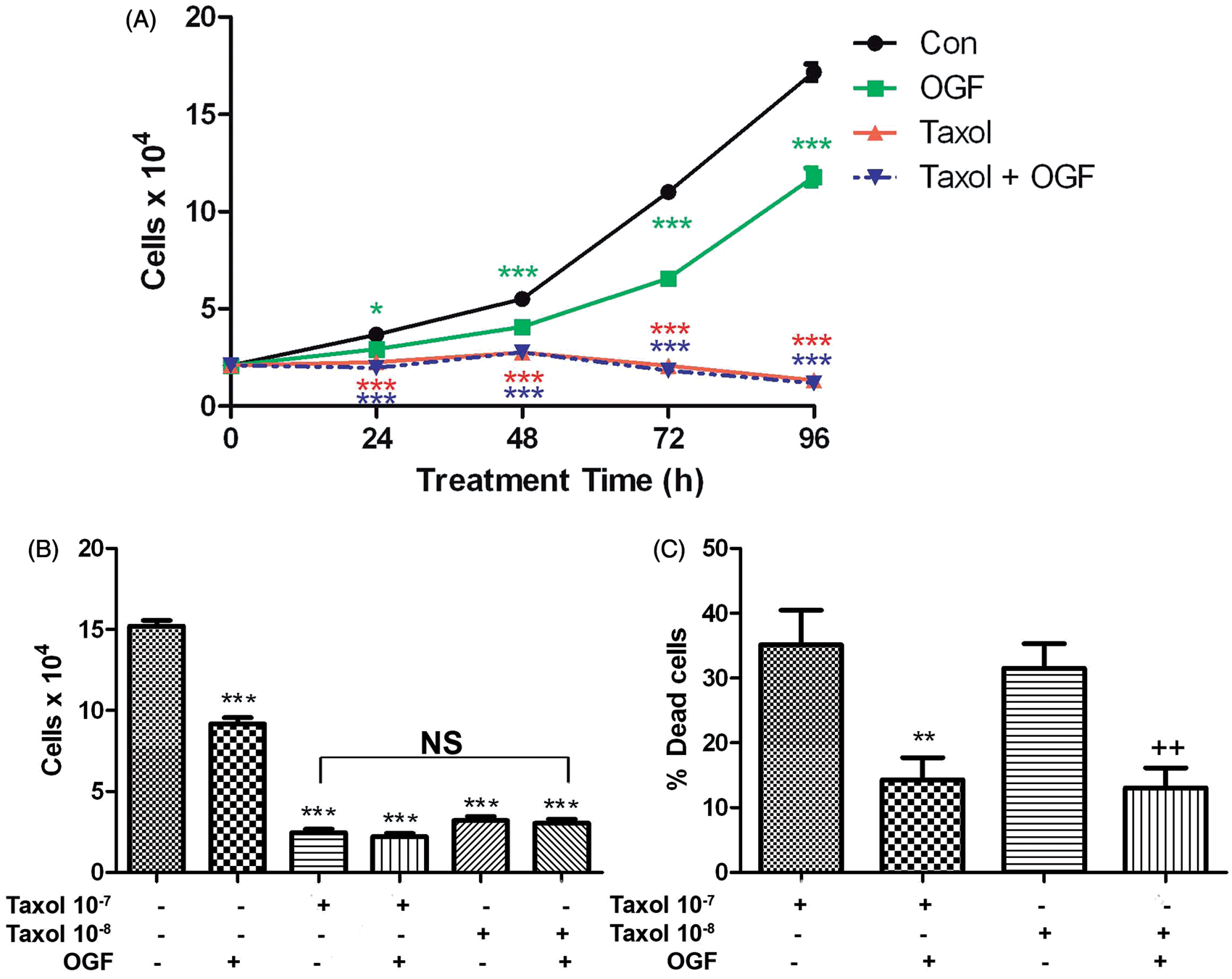
OGF and OGFr are present and attenuate growth of human breast cancer cell lines BT20, SK-BR-3, and MCF7
To examine the ubiquity of the OGF-OGFr axis in human breast cancer cell lines, we demonstrated the presence and activity in three additional cell lines. Epifluorescent microscopy revealed immunoreactive OGF and OGFr in the cytoplasm and nucleus of BT20, SK-BR-3, and MCF7 human breast cancer cells (Figure 7A). Addition of OGF in concentrations ranging from 10−4 to 10−10 M for 72 h revealed a dose-related inhibitory effect on cell proliferation (Figure 7B). Specifically, OGF (10−4–10−6 M) reduced cell number in BT20 cultures by 28–40% from untreated controls. Across a 120-h period, cell number in cultures treated with 10−6 M OGF was significantly reduced by 14% to 23% compared with control cultures (Figure 7B). OGF at concentrations of 10−4–10−8 M depressed cell number in SK-BR-3 cultures by 20–61% from control levels, and across a 120 h period, OGF (10−6 M) reduced cell number by to 41% compared with controls (Figure 7B). MCF7 cells were inhibited by OGF (10−4–10−7 M), with reductions of 20–26% being recorded relative to controls. Over a 120 h period, the cell number in OGF treated MCF7 cultures was significantly reduced by 13% to 22% compared with control cultures (Figure 7B).
OGF and OGFr are present and capable of attenuating growth in human breast cancer cell lines BT-20, SKBR3, and MCF7 that represent triple negative breast cancer (BT-20), and breast cancer cell lines containing HER2 receptors (SKBR3), or estrogen and progesterone receptors (MCF7). (A) Logphase cells were stained with DAPI, or polyclonal antibodies against OGF (Co172) or OGFr (Bo344). (B) Cells were grown in the presence of OGF (10−6 M) for the indicated period of time or with different concentrations of OGF for 72 h; controls were untreated. Data represent means ± SEM for two or more aliquots per well from at least two wells per group at each time point and were analyzed with ANOVA and Newman-Keuls post-hoc analyses. Significantly different from respective controls at P < 0.05 (*) or P < 0.001 (***). (A color version of this figure is available in the online journal)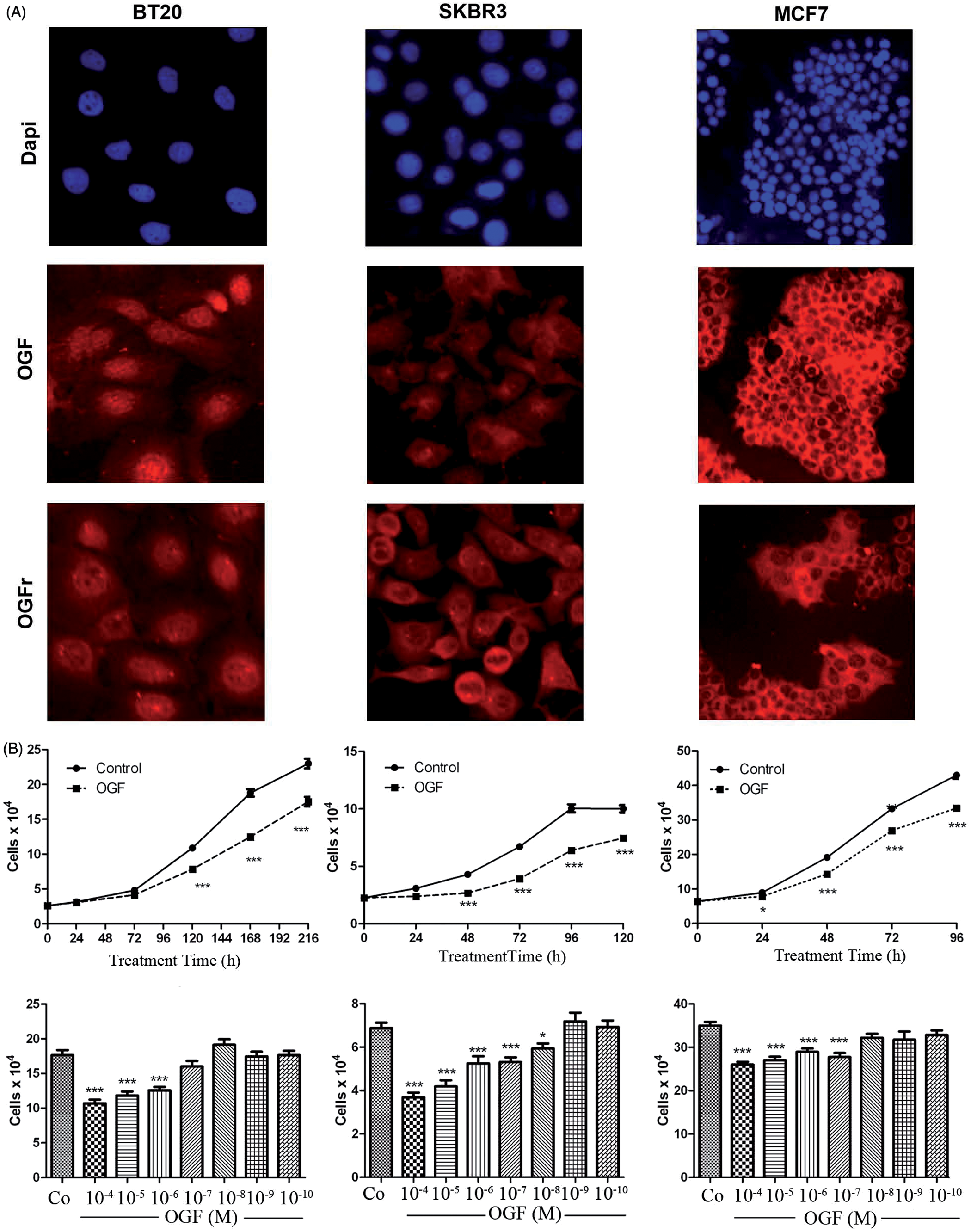
Discussion
This study shows for the first time that a biological pathway in human breast cancer involves the neuropeptide, OGF, and its nuclear-associated receptor, OGFr, is present and functions to inhibit cell proliferation of human breast cancer cells in vitro. Importantly, the OGF-OGFr axis is detectable and functioning in breast cancer cell lines that lack other receptors associated with targeted therapies. The present data showing that OGF inhibits cell replication in a dose-dependent, receptor-mediate, reversal manner provides new information on a potential biotherapy for TNBC. The specificity of both OGF and OGFr demonstrated that other classical opioid receptors and their natural and synthetic ligands are not involved in the mediation of inhibitory cell proliferative events. OGF inhibited the proliferation of MDA-MB-231 cells with a dosage that was of physiological relevance to the binding affinity for OGFr (nanomolar range). OGF repressed TNBC cell number in a rapid and persistent manner that did not exhibit tolerance, with significant reductions in cells recorded within 24 h of treatment.
Mechanistic analyses of the action of OGF revealed that RNA transcription and protein transcription and translation are required for the repression in DNA synthesis as measured by BrdU incorporation. The OGF-OGFr axis activated p21 cyclin-dependent inhibitory kinase, as knockdown of this protein removed much of the inhibitory effect of OGF. Moreover, the present study revealed that the OGF-OGFr axis is present and functioning in other human breast cancer cells, some of which possess estrogen and progesterone receptors (MCF-7) or the human epidermal growth factor receptor (SK-BR-2). These latter two cell lines provide evidence that OGF may be effective for all breast cell types, not only those lacking in the estrogen, progesterone, and HER-2 receptors. Thus, these results reveal for the first time the significance of a novel, biological pathway that regulates cell proliferation in human breast cancer.
OGF was the only endogenous opioid peptide altering growth of TNBC cells culture. Cells treated with the long-acting general opioid antagonist naltrexone were accelerated in their growth suggesting that the OGF-OGFr axis was an integral player in regulating human breast cancer cell proliferation. Moreover, these observations along with evidence that depletion of the peptide by antibody neutralization studies stimulated growth in vitro suggest that the endogenous opioid OGF is constitutively expressed by human breast cancer cells. Whether production or secretion of this peptide is deficient in breast cancer cells is unknown at this time.
OGF has been documented in a number of studies to interact with OGFr and to be the specific ligand for this opioid receptor.4,8,9 Evidence suggests that the growth inhibitory effects observed following both endogenous and exogenous OGF administration are mediated through the OGFr.3–5 The specificity of the OGFr receptor for mediation of cell replication was confirmed at the molecular level by siRNA studies. Transfection studies on other human cancer cell lines establishing stable under- and over-expression of OGFr reveal that the receptor is a physiological determinant of cell proliferation. 3 Although enkephalins bind to other opioid receptors, data from many studies support that OGF is interacting through OGFr to mediate its repressive action on human cancer cell lines.4,8,9 The MDA-MB-231 cell lines that were transiently transfected with OGFr siRNA responded to a partial knockdown of the OGFr by acceleration in growth, and within 72 h more cells were reported than in control cultures. Addition of exogenous OGF which normally reduced cell number by ∼20% had no inhibitory effect. The magnitude of regulation by these transfection studies needs to be considered because some OGFr protein expression remained following transfection and enabled OGF to bind. The outcome of patient tumour cells that may have little or no OGFr expression has not been determined.
Therapeutic regimens for the treatment of cancer often involve cocktails of chemotherapy or combinatorial approaches. Paclitaxel is one of the widespread agents used in the treatment of breast cancer, but this agent has been associated with many side-effects including prolonged nausea, abnormal bruising, joint pain and exhaustion. These effects often result in reduced compliance by patients for continuing therapy. Our studies assessing the combinatorial effects of OGF and paclitaxel revealed that OGF does not provide any additive effect in reduction of cell number to that which follows paclitaxel treatment. However, addition of OGF did reduce by more than 50% the overall number of dead cells. Thus, it appears that when OGF is combined with paclitaxel, OGF inhibits cells and provides some form of protection against apoptosis or necrosis induced by paclitaxel alone. These data extend information reported on squamous cell carcinoma of the head and neck 19 and ovarian adenocarcinoma 20 where the addition of OGF to paclitaxel demonstrated additive effects in tissue culture studies. Moreover, the addition of OGF was both protective against the toxicity of paclitaxel, and provided additional inhibitory influence in nude mouse studies, supporting the use of OGF in combination with other standard chemotherapies. 19
Research in the field of breast cancer is very active, and large-scale clinical trials are ongoing.21–27 However, treatment of TNBC which lacks hormonal receptors 27 that can be targeted by specific drugs such as tamoxifen,22,25 continues to require further research. Tate et al. 23 recently reported that panobinostat, a histone deacetylase inhibitor, showed efficacy by killing TNBC cells in vitro and decreasing tumorigenesis in vivo. The toxic effects of panobinostat were induced by increasing histone acetylation and blocking cell cycle progression at the G2/M phase. Given that OGF blocks cell cycle progression at the G1/S phase, the combination of these novel therapies may be very effective against TNBC. It could be envisioned that OGF would be effective as a combinatorial therapy with activation of TRAIL to accelerate apoptosis, 24 and inhibit DNA synthesis, in TNBC. The broader implications for OGF in regards to its ability to be protective against toxic chemotherapies, as well as its efficacy and potential use in cocktail therapy to treat hormone-receptor containing breast cancers, requires further investigation but provides an exciting new biological pathway to explore.
In conclusion, the data demonstrate for the first time that a native biological pathway is present and plays a role in regulating the proliferation of human breast cancer cell lines including TNBC. OGF is a receptor-mediated, non-toxic, constitutively-expressed, inhibitory peptide that effectively represses cell replication through the p21 cyclin-dependent inhibitory kinase mechanism. The OGF-OGFr axis provides a novel biological pathway for the treatment of human breast cancer.
Footnotes
Author contributions
All authors participated in the design, data analyses and review of the manuscript. NKP performed much of the research. ISZ and PJM interpreted the data and wrote the manuscript.
Acknowledgements
SK-BR-3 cells were provided by Dr. James Connor, Department of Neurosurgery, Penn State University College of Medicine. Funding to support this research included non-discretionary funds (ISZ). There are no conflicts of interest. Penn State University holds a patent (ISZ and PJM co-inventors) on the use of OGF for treatment of pancreatic and gastrointestinal cancer.