
Abstract
Galangin is a member of flavonols and found in Alpinia officinarum, galangal root, and propolis. Previous studies have demonstrated that galangin has anti-cancer effects on several cancers, including melanoma, hepatoma, and leukaemia cells. However, anti-cancer activity of galangin on human colon cancer has not been established yet. In this study, we investigated the anti-cancer effects of galangin on two types of human colon cancer cells (HCT-15 and HT-29). We found that galangin induced apoptosis and DNA condensation of human colon cancer cells in a dose-dependent manner. We also determined that galangin increased the activation of caspase-3 and -9, and release of apoptosis inducing factor from the mitochondria into the cytoplasm by Western blot analysis. In addition, galangin induced human colon cancer cell death through the alteration of mitochondria membrane potential and dysfunction. These results suggest that galangin induces apoptosis of HCT-15 and HT-29 human colon cancer cells and may prove useful in the development of therapeutic agents for human colon cancer.
Introduction
Colorectal cancer resulting from uncontrolled cell growth in the colon or rectum is one of the leading tumours in the world and is considered a ‘big killer’ together with lung, prostate, and breast cancer. Most colorectal cancers occur due to lifestyle and increasing age, with only a minority of cases associated with an underlying disorder. 1 Moreover, colorectal cancer is the second most common cause of cancer deaths in men and women. This corresponds to approximately 10% of all cancer deaths.
Flavonoids are a group of more than 4000 polyphenol compounds that occur naturally in foods of plant origin. These compounds possess a common phenylbenzopyrone structure and are primarily categorized into flavonols, flavones, isoflavones, flavanols, and flavanonols according to the saturation level and opening of the central pyran ring.2,3 These polyphenol compounds display a remarkable spectrum of biological activities including antioxidant, anti-inflammatory, anti-carcinogenic, and modulation of enzymatic activities that might be able to influence processes that are dysregulated during cancer development.4,5
Galangin (3,5,7-trihydroxyflavone), a member of the flavonol class of flavonoids, is present at high concentration in propolis, a natural material produced by honeybees, and in India root, Alpinis officinarum, which is a common spice and herbal medicine in Asia. 6 Recently, many studies have suggested that galangin has anti-cancer effects against several cancers.7–11 However, the effect of galangin on human colon cancer has not been established.
In the present study, we investigated the anti-cancer effect of galangin on human colon cancer cells, including HCT-15 male human colon cancer cell and HT-29 female human colon cancer cell.
Materials and methods
Reagents
Galangin and 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) were purchased from Sigma Chemical Co. (St. Louis, MO, USA). ProLong® Gold Antifade reagent with 4’,6-diamidino-2-phenylindole (DAPI) and tetramethyl rhodamine methyl ester (TMRM) were purchased from Invitrogen (Carlsbad, CA, USA).
Cell culture and galangin treatment
HT-29 and HCT-15 cells were obtained from American Type Culture Collection (Rockville, MD, USA). Cells were grown as described by Fujimoto et al.
12
Briefly, cells were cultured at 37℃ in the presence of 5% CO2 in RPMI 1640 supplemented with 10% fetal bovine serum, 200 IU/mL penicillin, 200 µg/mL streptomycin, 2 mmol/L
Cell viability assay
Cell viability was measured by colorimetric MTT assay. The cells (2 × 104 cells/well) were seeded in 96-well culture plates with RPMI 1640. Cells were treated with various concentrations of galangin (5–200 µmol/L) and incubated in 37℃ for 24 h. After treatment, medium containing galangin was removed and MTT (0.5 mg/mL) in phosphate-buffered saline (PBS) was added to each well. After incubation a 37℃ for 4 h, MTT solution was removed and formazan product was dissolved in solubilization solution (1:1 = DMSO:ethanol) into a coloured solution. Absorbance of the formazan solution was quantified by an enzyme-linked immunosorbent assay microplate reader at 570 nm.
DNA condensation assay
HT-29 and HCT-15 cells were first seeded on a round coverslip in a 12-well cell culture plate. After exposure to various concentrations of galangin for 24 h, the cells were fixed by incubation in 4% paraformaldehyde (PFA) for 10 min. After washing with PBS three times, the cells on coverslip were mounted on glass slides with permanent mounting solution with DAPI. The cells were then observed with a fluorescence microscope (Nikon, Tokyo, Japan).
Annexin-V/PI binding assay
Cells were seeded onto a six-well cell culture plate and treated with various concentrations of galangin. After 24 h, both floating cells and attached cells were collected and pooled into round-bottom tubes. The collected cells were washed with PBS and stained with fluorescein isothiocyanate (FITC)-labelled annexin-V (Annexin-V–FITC) and PE-labelled propidium iodide (PE–PI) (Annexin V Apoptosis Detection Kit II; BD Pharmingen™, CA, USA). The stained cells were analysed using a FACSCalibur flow cytometer for 10,000 event for each experimental setup (BD Biosciences, CA, USA) and FlowJo software (ver 7.6.5; Tree Star Inc., OR, USA).
Western blot analysis
Galangin-treated (5–200 µmol/L) HCT-15 and HT-29 cells were lysed in ice-cold whole cell lysate buffer (RIPA) containing 150 mmol/L sodium chloride, 1% Triton X-100, 0.5% sodium deoxycholate, 0.1% sodium dodecyl sulphate (SDS), 50 mmol/L Tris (pH 8.0), 1 mmol/L phenylmethylsulphonyl fluoride, 2 µg/mL leupeptin, 1 µg/mL pepstatin, 1 mmol/L sodium orthovanadate, and 100 mmol/L sodium fluoride. The mixture was incubated at 4℃ for 30 min. Cell debris was removed by microcentrifugation, followed by quick freezing of the supernatant. The protein concentration of each sample was determined using a micro bicinchoninic acid assay kit (Pierce, Rockford, IL, USA). Equivalent protein samples (20 µg) were separated by 10% SDS-polyacrylamide gel electrophoresis (SDS-PAGE) and electrophoretically transferred to polyvinylidene difluoride (PVDF) transfer membrane. The membrane was placed into a blocking solution (5% nonfat milk) at room temperature for 1 h. After blocking, anti-β-actin, anti-caspase-9, anti-caspase-3, anti-apoptosis inducing factor (AIF), and anti-cytochrome c antibodies (Santa Cruz Biotechnology, CA, USA) were used as primary antibodies. Horseradish peroxidase-conjugated anti-rabbit and anti-mouse antibodies (Santa Cruz Biotechnology) were used as secondary antibodies. Band detection was performed using the enhanced chemiluminescence (ECL) detection system and exposed to radiographic film. Pre-stained blue markers were used for molecular weight determination.
Mitochondrial membrane potential (Δψm) assay
The cells were seeded into six-well cell culture plate and then treated with 200 µmol/L galangin. Both floating cells and attached cells were collected at 0, 15, 30, 60, 120, and 180 min, and pooled into a round-bottom tube. Collected cells were washed with PBS and stained with 20 nmol/L fluorescent dye TMRM ester. TMRM to detect mitochondrial permeability and membrane potential in whole and living cells. The stained cells were analysed using a FACSCalibur flow cytometer and FlowJo software for 10,000 events for each experimental condition.
Mitochondrial fragmentation
For mitochondrial fragmentation, we used pDsRed2-Mito plasmid DNA, which encodes a fusion of Discosoma sp. red fluorescent protein and the mitochondrial targeting sequence. Cells were seeded onto a six-well cell culture plate at 5 × 105 cells/well, and after 24 h the cells were transiently transfected using Lipofectamin 2000 (4 µg of DNA vector per well; Invitrogen). The transfected cells were seeded on polyethyleneimine-coated cover glass in a 12-well cell culture plate at a density of 2 × 105 cells/well in complemented RPMI-1640 medium. After 24 h, the cells were treated with or without 200 µmol/L of galangin. The cells were fixed by 4% PFA at 0 and 4 h and mounted on slide glass with Prolong® Gold Antifade reagent with DAPI. The cells were then observed with a confocal microscope (Carl Zeiss, Stuttgart, Germany).
Statistical analysis
The results are presented as the mean ± standard deviation. The data were analysed by one-way analysis of variance (ANOVA) followed by Duncan’s post hoc test using SPSS (IBM, Armonk, New York, NY, USA). The differences were considered statistically significant at P < 0.01.
Results
Galangin induced the cell death of HCT-15 and HT-29 cells
In preliminary studies, we investigated whether galangin has anti-cancer activity in human colon cancer cells. The cytotoxic effect of galangin on HCT-15 and HT-29 cells was measured by MTT assay. Galangin treatment reduced cell viability of HCT-15 and HT-29 cells in a dose-dependent manner (with increasing concentration, 5–200 µmol/L). In comparison to the control (treated with only 0.4% DMSO), treatment of 100 and 200 µmol/L of galangin induced approximately 75% and 85% reductions in cell viability on HCT-15 and HT-29 cells (Figure 1). This suggested that treatment of galangin induces the cell death in HCT-15 and HT-29 cells.
Galangin treatment induced decreased cell viability and/or increased cell death in HCT-15 and HT-29 cells. (a) Structures of galangin. (b) HCT-15 and HT-29 cells were plated in 96-well microplates. Experimental medium (RPMI 1640, 0.4% DMSO) containing various concentrations of galangin was added, and viable cell numbers were assessed by MTT assay after 24 h incubation, as described in ‘Materials and methods’ section. Data are reported as viable cell numbers of galangin-treated cells, expressed as a percentage of control cells that were exposed to 0.4% DMSO. The values of galangin concentration tested represent the average (±SD) from four replicate wells and are representative of three separate experiments. The differences were considered statistically significant at **P < 0.01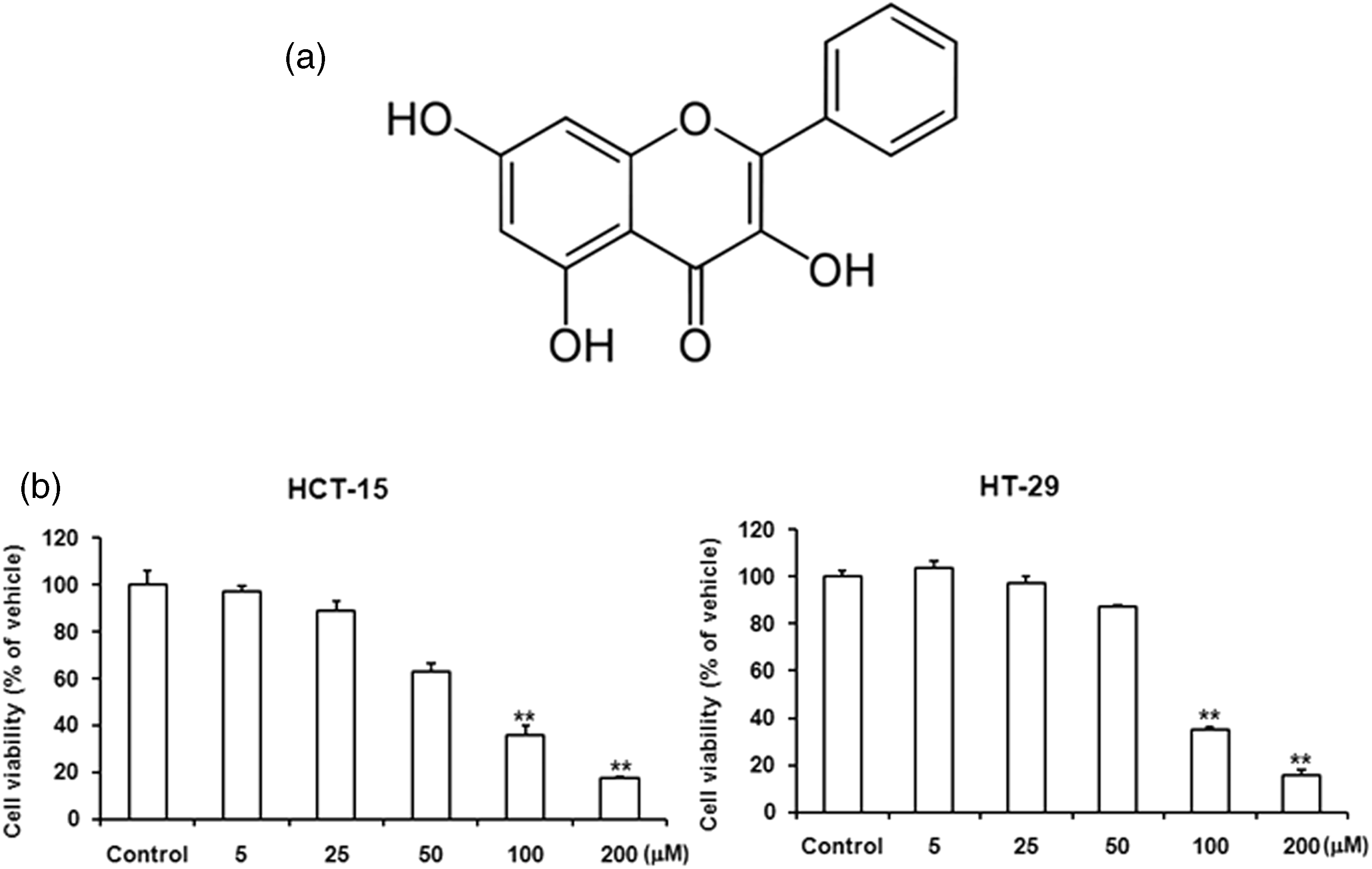
Galangin induced nuclear condensation and apoptotic cell death in HCT-15 and HT-29 cells
Given our results discussed above, galangin at concentrations of 50, 100, and 200 µmol/L were used to study the major mechanisms underlying galangin-induced cell death in HCT-15 and HT-29 cells. A nuclear condensation assay was used to assess the morphological changes of apoptosis in HCT-15 and HT-29 cells. As shown in Figure 2a, DAPI staining revealed that treatment of HCT-15 and HT-29 cells with various concentrations of galangin induced significant nuclear rounding and shrinkage in HCT-15 and HT-29 cells when compared to the control (Figure 2a). In addition, we determined galangin-induced apoptotic cell death by an Annexin/PI staining assay. Galangin treatment dose-dependently led to 50.65% and 61.1% apoptotic cells (Annexin-V+/PI− and Annexin-V+/PI+) on the HCT-15 and HT-29 cells populations, respectively (Figure 2b). Galangin treatment induced some PI-positive cells simultaneously with apoptotic cell death, although we considered this secondary necrotic cell death followed by apoptosis. These results indicated that the treatment of galangin induced cell death of HCT-15 and HT-29 cells via apoptotic events.
Treatment of galangin induced the nuclear condensation and apoptotic cell death in HCT-15 and HT-29 cells. (a) HCT-15 and HT-29 cells were treated with various concentrations of galangin (5–200 µmol/L for 24 h and then DAPI staining was performed as described in ‘Materials and methods’. The lower panels show the corresponding fields of DAPI fluorescence. Original magnification was 200 × and 400 × . (b) Human colon cancer cells were treated with various concentrations of galangin for 24 h and stained with Annexin-V–FITC and PI–PE. Fluorescence intensity was measured by flow cytometry and presented as a percentage of each quadrant. (A color version of this figure is available in the online journal)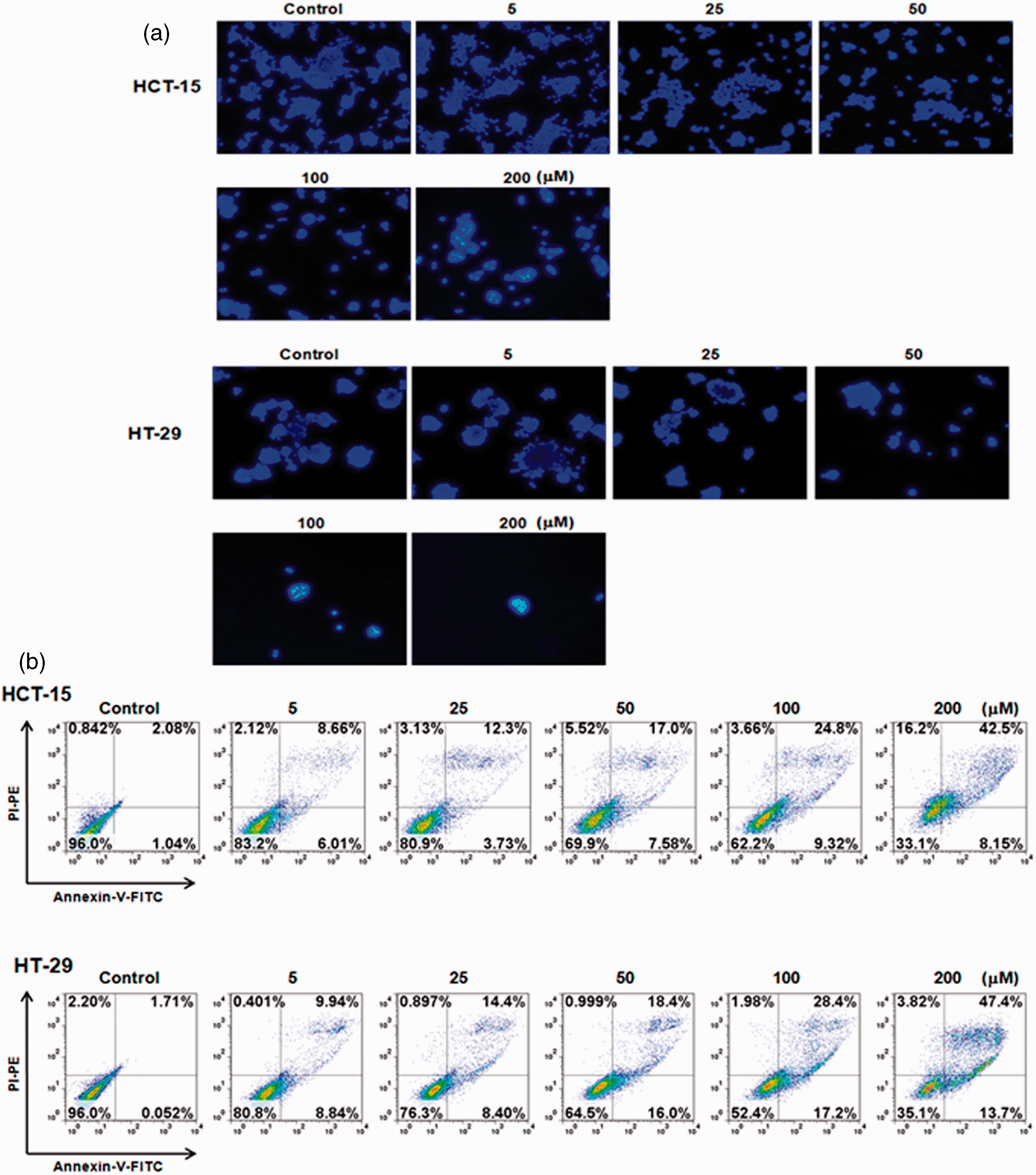
Galangin induced the activation of caspase-3 and caspase-9 in HCT-15 and HT-29 cells
Activation of the caspase cascade is important for the induction and processing of apoptosis. For this reason, the down-stream events of galangin-induced apoptosis in HCT-15 and HT-29 cells were characterized by activation of caspase-3 and caspase-9 using Western blot analysis. As shown in Figure 3a and b, the pro-caspase forms of caspase-3 and caspase-9 were markedly decreased in HCT-15 and HT-29 cells by various concentrations of galangin treatment (Figure 3a and b). These data indicated galangin-induced apoptosis of human colon cancer cells may occur via the caspase-dependent pathway.
Galangin induced cleavage of caspase-3 and caspase-9. Human colon cancer cells were incubated with different galangin concentrations for 24 h and subsequently lysed. Equal amounts of proteins were then separated by SDS-PAGE and transferred to PVDF membranes. Membranes were probed with the indicated antibodies (anti-caspase-3, anti-caspase-9, and anti-β-actin) and detected by ECL solution. The β-actin was used as the internal control. The results are from one experiment representative of four performed that showed similar patterns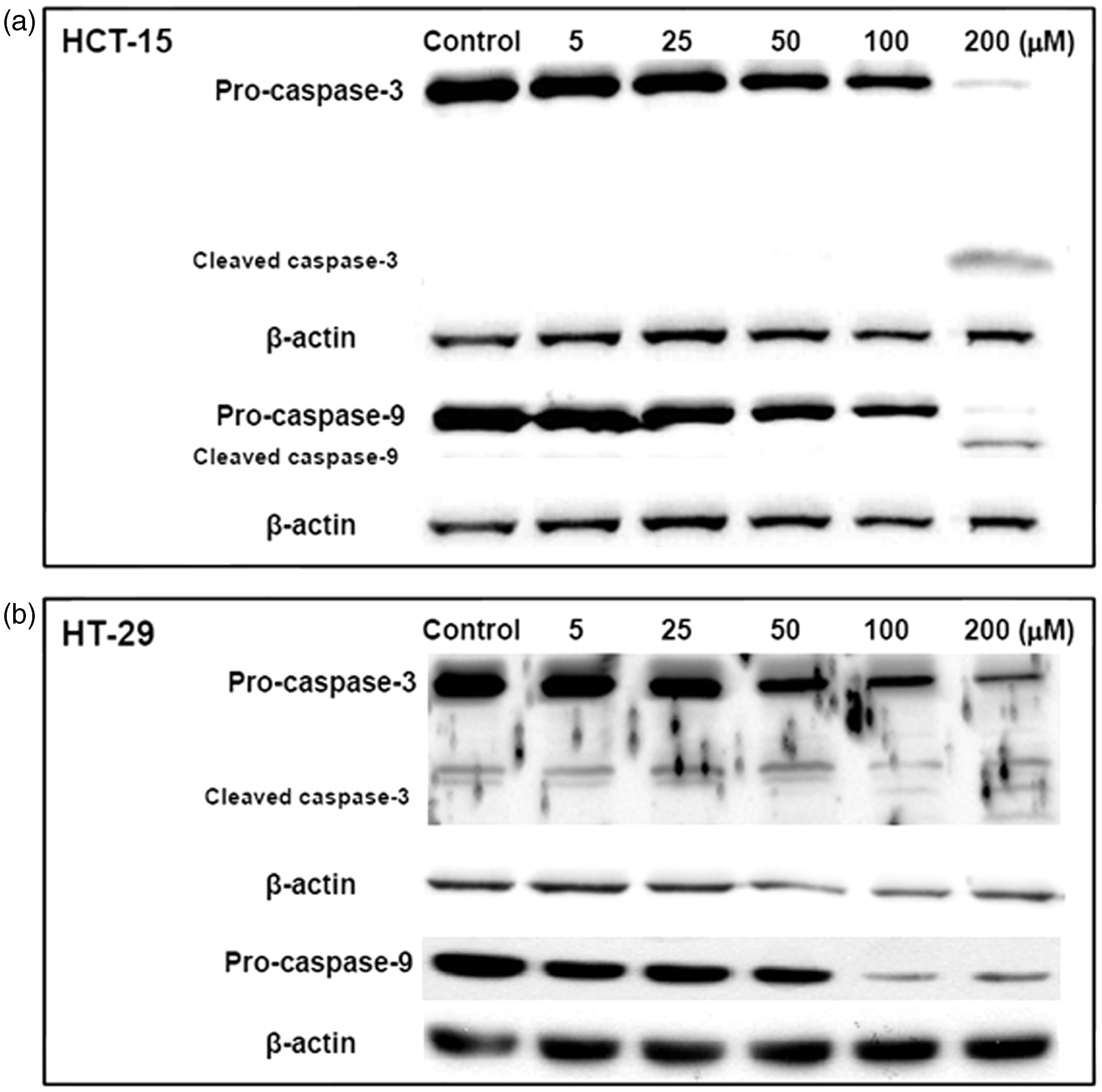
Galangin induced disruption of mitochondrial membrane potential in human colon cancer cells
In the previous experiments, galangin induced human colon cancer cell death through the induction of DNA condensation and activation of caspase-3 and caspase-9. To investigate whether galangin induces a change of mitochondria membrane potential (MMP), we measured the MMP. Galangin rapidly induced the depolarization of mitochondria membrane and subsequently led to hypopolarization. Moreover, the present study sought to establish whether galangin damages the mitochondria by using a mitochondrion-selective fluorescent packaged protein (pDsRed2-Mito), the uptake of which depends on intact mitochondrial fragmentation. In the untreated human colon cancer cells, mitochondria were normal and abundant throughout the cytoplasm (Figure 4b). Mitochondria fragmentation is recognized to be an important event during the onset of apoptosis, involved in apoptosis-associated activation of outer mitochondrial membrane scission during cell death. When the cells were exposure with 200 µmol/L galangin, the reticular mitochondria of human colon cancer cells became fragmented, swollen, and aggregated in circular forms. Therefore, we concluded that the mitochondria fragmentation was induced by galangin treatment at 4 h (Figure 4b).
Galangin induced cell death of human colon cancer cells by mitochondrial alterations. (a) Human colon cancer cells were treated with 200 µmol/L galangin for various times (0–180 min) and stained with 20 nmol/L TMRM. Fluorescence intensity was measured by flow cytometry and presented as mean fluorescence intensity (MFI). (b) HCT-15 cells were transfected with N-terminal mitochondrial signal sequence of DsRed (pDsRed2-Mito) and then incubated 200 µmol/L of galangin for 4 h. After incubation, cells were fixed and then analysed by confocal microscopy. (A color version of this figure is available in the online journal)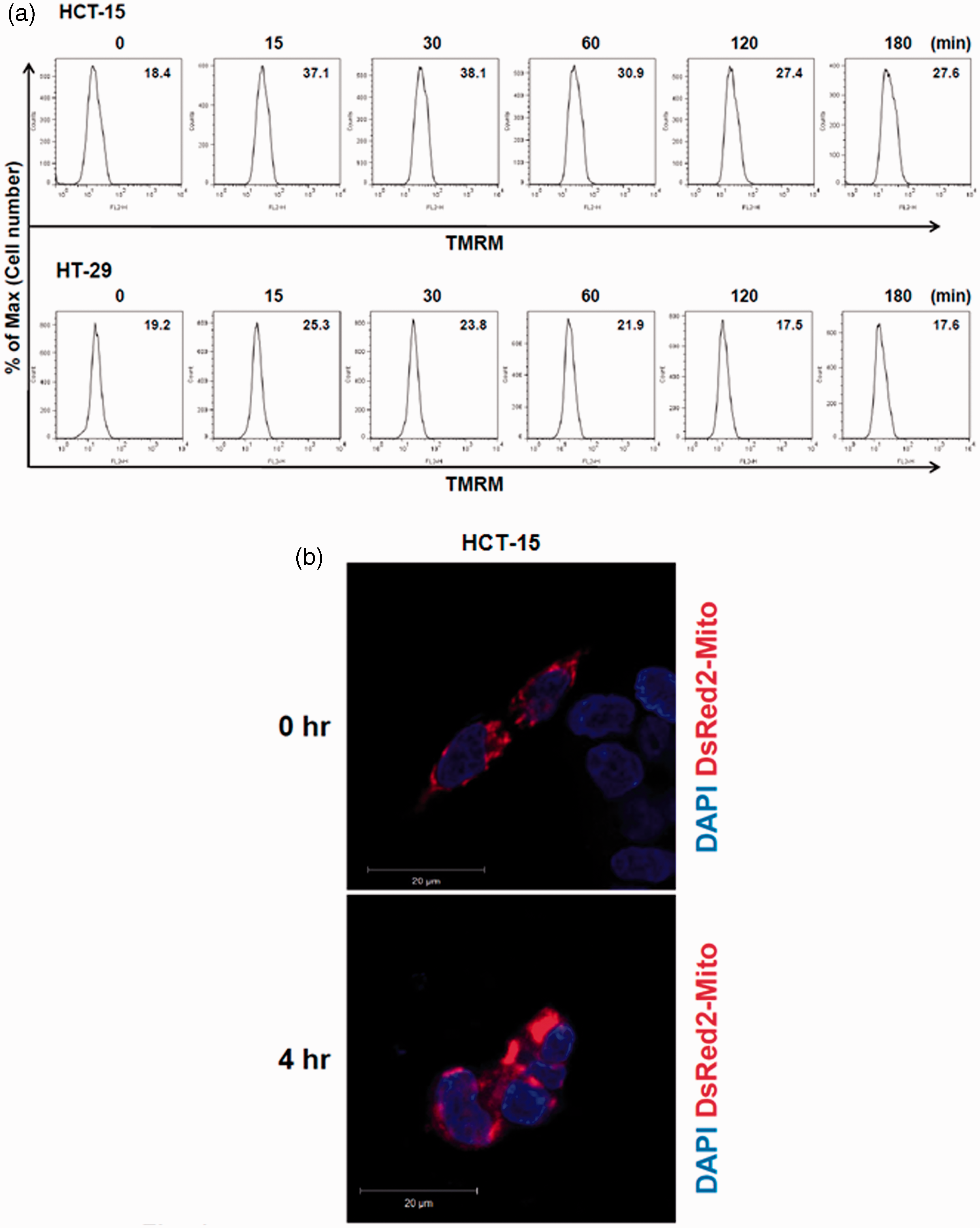
Galangin induced AIF release
Induction of mitochondrial fragmentation proceeded all morphological and biochemical signs of apoptosis. Dysfunction of mitochondria can result in the release of pro-apoptotic molecules such as AIF into the cytosol. In addition, apoptosis can be initiated by mitochondrial damage with significant caspase activation.
13
Galangin treatment induced apoptosis via the intrinsic pathway with release of AIF from mitochondria in HCT-15 and HT-29 cells. These results indicated that galangin induced apoptosis of human colon cancer cells via mitochondria dysfunction and AIF release (Figure 5).
Galangin induced AIF release from mitochondria into cytosol in human colon cancer cells. Human colon cancer cells were incubated with different galangin concentrations for 24 h, subsequently lysed, and equal amounts of cytosolic fraction proteins were separated by SDS-PAGE and transferred to PVDF membranes. Membranes were probed with the indicated antibodies (anti-AIF, and anti-β-actin) and detected by ECL solution. β-actin was used as the internal control. The results are from one experiment representative of four performed that showed similar patterns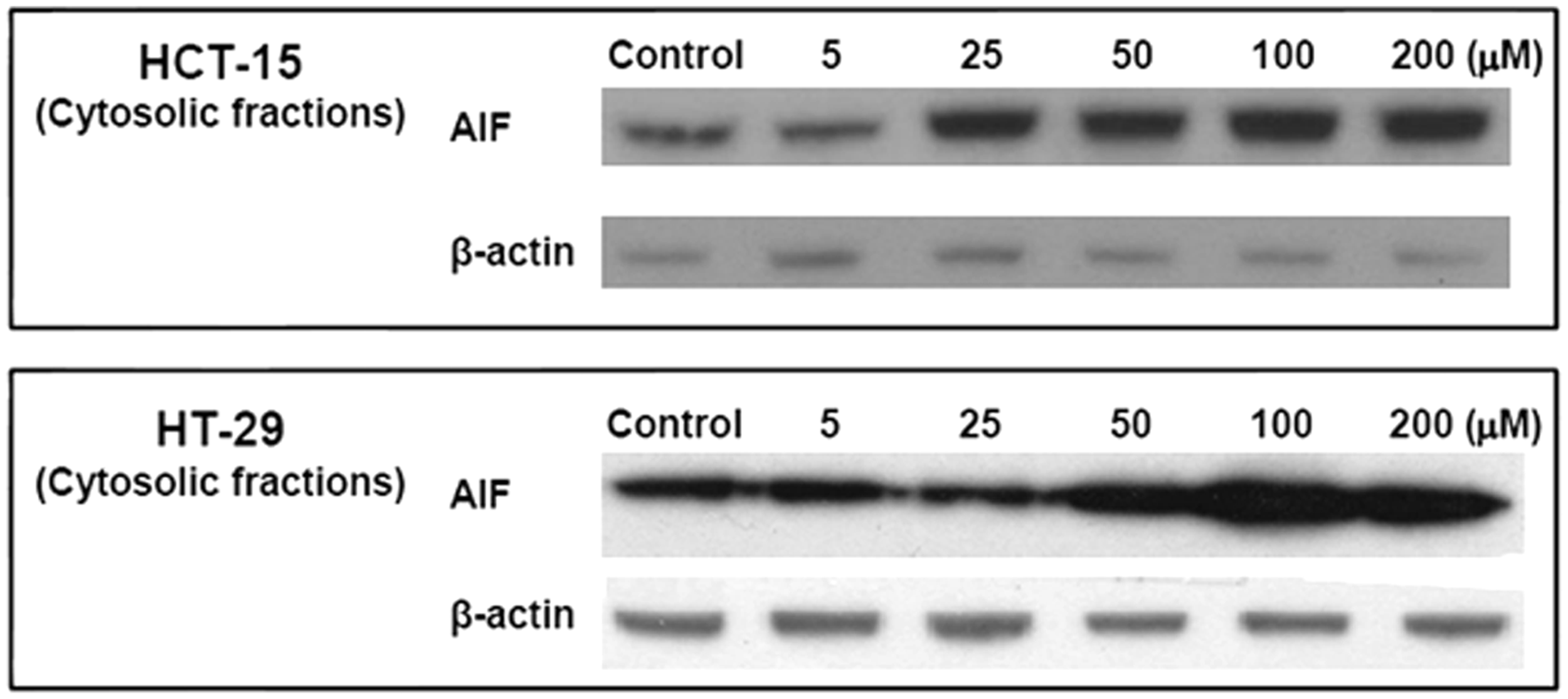
Discussion
Galangin has been used as a spice and an herbal remedy for a variety of ailments in Asia for centuries. Recently, galangin has been reported to induce apoptotic cell death and autophagy on HepG2 hepatocellular carcinoma cells via p53-dependent pathway and induce changes in cells in G0/G1 of the cell cycle, modulate expression of cyclin/cdk, and induce apoptosis in imatinib-resistant Bcr-Abl-expressing leukaemia cells.7,8 Moreover, galangin prevents oxidative damage and has a down-regulatory effect on the inflammatory pathway in livers of fructose-fed rats. 14 However, the effects of galangin on HCT-15 and HT-29 cells were previously unknown. This study focused on the anti-colon cancer activity of galangin on human colon cancer cells in vitro to identify a new therapeutic agent.
As shown in Figure 1, galangin induced human colon cancer cell death in a dose-dependent manner. After 24 h of incubation, at the concentrations of 50, 100, and 200 µmol/L galangin treatment significantly reduced the viability of human colon cancer cells in comparison to control. As expected, human colon cancer cells exhibited an increase in DAPI-positive cells with increased concentrations of Galangin treatment (Figure 2). DAPI-positive cells significantly increased with 100 and 200 µmol/L galangin treatment, although it was not significant with 50 µmol/L of galangin.
The two primary modes of cell death are apoptosis and necrosis. To investigate which type of cell death galangin induced in human colon cancer cells, 15 we determined apoptotic cell death and necrotic cell death by Annexin-V/PI staining. The flipping of phosphatidyl serine from the inside to the outside of the cell membrane, a major event in the apoptotic process, was detected by Annexin-V binding assay after treatment of galangin on HCT-15 and HT-29 cells. Stained cells were classified into apoptotic (Annexin-V+/PI−) and necrotic/late apoptotic cells (Annexin-V+/PI+). As shown in Figure 2b, we found galangin induced apoptotic cell death of human colon cancer cells in a dose-dependent manner.
The mitochondrial pathway is relatively more important than the death-receptor pathway for induction of apoptosis by chemotherapeutic agents. 16 Mitochondrial dysfunction leads to a change in mitochondrial membrane potential and caspase activation. Caspase activation is essential in cells for apoptosis. Previous studies have shown that galangin induces cell death via mitochondria pathway. Galangin increased expression level of cleaved caspase-3 and -9 on SNU-484 gastric cancer cells and HepG2 hepatocellular carcinoma cells.8,9 To investigate whether galangin induced human colon cancer cell death through the activation of caspase cascades, human colon cancer cells treated with different concentrations of galangin (5, 25, 50, 100, and 200 µmol/L) for 24 h. As shown in Figure 3, 200 µmol/L galangin-treatment significantly induced cleavage of caspase-3 and caspase-9. Consistent with previous studies, these results indicated that galangin treatment induced human colon cancer cell death activity through the activation of the caspase-dependent pathway (Figure 3a and b). Alteration of the MMP is involved in mitochondrial dysfunctions. When human colon cancer cells were treated with galangin, the MMP was altered at 30 min (Figure 4a). This result concurs with the finding that galangin decreased mitochondrial membrane potential on HepG2 hepatocellular carcinoma cells. 9 Moreover, to visualize changes in mitochondrial morphology pre and post galangin treatment, mitochondria were labelled with DsRed. As shown in Figure 4b, untreated mitochondria of human colon cancer cells appeared tubular and interconnected in filamentous networks. However, when cells were incubated with galangin, the reticular mitochondria of human colon cancer cells became fragmented. For example, 4 h after Galangin treatment, almost all the mitochondria in human colon cancer cells were fragmented. Moreover, this study demonstrated human colon cancer apoptosis by galangin treatment via enhancement of AIF release from mitochondria into the cytosol (Figure 5).
In summary, galangin treatment reduced cell proliferation and induced apoptotic death in HCT-15 and HT-29 cells. Galangin induced apoptosis through activation of caspase-3/-9 and AIF release from mitochondria into the cytosol. Moreover, galangin-treated human colon cancer cells showed alteration of the MMP and mitochondria fragmentation. These results suggested that galangin-induced apoptosis was mediated by mitochondrial dysfunction and the caspase-dependent pathway, which may prove useful in the development of therapeutic agents for the treatment of human colon cancer.
Footnotes
Author contributions
All authors participated in the design, interpretation of the studies and analysis of the data and review of the manuscript; TKH, MEK, JHY, SJB, and JY conducted the experiments, TKH and JSL wrote the manuscript, and MEK contributed the Statistical analysis.
Acknowledgements
This work was supported by the 2013 Inje University research grant.