
Abstract
The influence of the histone deacetylase (HDAC)-inhibitor, valproic acid (VPA), on bladder cancer cell adhesion in vitro was investigated in this paper. TCCSUP and RT-112 bladder cancer cells were treated with VPA (0.5 or 1 mM) twice or thrice weekly for 14 days. Controls remained untreated. Tumour cell interaction with immobilized collagen was evaluated by a flow-based adhesion assay using a shear force of 2 or 4 dyne/cm2. The effects of VPA on the integrin adhesion receptors α3, α5, β1, β3 and β4 were assessed by flow cytometry to determine integrin surface expression and by western blotting to determine the cytoplasmic integrin level. VPA of 0.5 mM and 1 mM significantly prevented binding of both RT-112 and TCCSUP cells to collagen, compared with the untreated controls. Adhesion was reduced to a higher extent when RT-112 (subjected to 2 dyne/cm2) or TCCSUP (subjected to 2 or 4 dyne/cm2) tumour cells were treated with VPA three times a week, compared to the two times a week protocol. VPA caused a significant up-regulation of the integrin α3, α5, β1, β3 and β4 subtypes on the TCCSUP cell surface membrane. In RT-112 cells, only integrin α5 was elevated on the cell surface following VPA exposure. Western blotting revealed an up-regulation of α3, α5, β3 and β4 integrins and down-regulation of the integrin β1 protein by VPA in TCCSUP. VPA also up-regulated α5 and down-regulated β1 integrin in RT-112 cells, but also reduced α3 and β3 in TCCSUP. VPA exerted adhesion-blocking properties on bladder cancer cells under physiologic flow conditions. The effects were accompanied by distinct modifications of the integrin expression profile, which differ depending on the cell lines used. Application of VPA might be an innovative option to prevent bladder cancer dissemination.
Introduction
Bladder cancer is the fourth most common cancer in men and the eighth most common cancer in women. Despite surgical and medical advances, the mortality of bladder cancer has not substantially improved over the last 20 years. Systemic chemotherapy based on cisplatin and gemcitabine or on methotrexate, vinblastine, adriamycin and cisplatin (MVAC) has induced significant response rates in patients with locally advanced or metastasized urothelial carcinoma. 1 However, over time disease progression inevitably occurs in most patients, presumably caused by development of resistance, and overall survival has only been extended by four months. 1
Epigenetic alterations are closely associated with the development and progression of bladder cancer. 2 Particularly, an increased expression of histone deacetylases (HDAC), leading to an imbalance in the equilibrium between histone acetylation and histone deacetylation, has been proposed as a driving force which causes normal cells to become malignant. 2 Immunohistochemical methods have revealed a strong correlation between the over-expression of HDAC and an increased occurrence of bladder cancer, as well as significant HDAC being positivity associated with the severity of bladder cancer. 3 Hence, down-regulating HDAC by HDAC-inhibitors might be an option in thwarting bladder cancer.
Indeed, the HDAC-inhibitors entinostat (MS-275), belinostat (PXD101) or vorinostat (suberoylanilide hydroxamic acid) have all been shown to block bladder cancer cell growth in vitro and in vivo.4–6 We recently demonstrated that the HDAC-inhibitor valproic acid (VPA) exerts growth-blocking properties on a panel of bladder cancer cell lines, commensurate with dose and exposure time. 7 In a mouse model, VPA administration has also delayed the time to develop urinary bladder tumour. 2 These promising preclinical experiments suggest that HDAC-inhibitors may be an innovative tool to fight bladder cancer. Still, information about the effect of HDAC-inhibitors on bladder cancer cell adhesion, which is a necessary prerequisite to metastatic spread, is lacking. Therefore, the impact of VPA on the adhesive activity of bladder cancer cell lines was investigated.
Materials and methods
Cells
RT-112, RT-4, 5637, UMUC-3 (ATCC/LGC Promochem GmbH, Wesel, Germany) and TCCSUP (DSMZ, Braunschweig, Germany) bladder carcinoma cells were grown and subcultured in RPMI 1640, 10% fetal calf serum (FCS), 20 mM HEPES-buffer, 1% glutamax and 1% penicillin/streptomycin (all: Gibco/Invitrogen, Karlsruhe, Germany). RT-112 is an invasive (pathological stage T2) moderately differentiated (grade 2/3) model of human bladder cancer, whereas TCCSUP is a transitional cell carcinoma, grade 4. RT-4 represents a grade 1 papillary tumour, 5637 a grade 2 and UMUC-3 a high grade 3, invasive bladder cancer. Subcultures from passages 7–24 were selected for experimental use.
Valproic acid
VPA (gift from G. L. Pharma GmbH, Lannach, Austria), at final concentrations of 0.5 or 1 mM, was added to bladder carcinoma cells two or three times a week for 14 days. Controls remained untreated. To exclude toxic effects, cell viability was determined by the trypan blue exclusion test (Gibco/Invitrogen).
Flow-based adhesion assay
Fibrillar collagen from bovine tendon (Sigma, Deisenhofen, Germany) was prepared as described earlier. 8 Adhesion of TCCSUP and RT-112 cells to fibrillar collagen under physiologic flow conditions was performed as reported previously with minor modifications. 9 Briefly, µ-slides (Ibidi, Martinsried, Germany) were coated overnight at 4℃with fibrillar collagen in a humidified environment according to the manufacturer’s instructions. The µ-slides were blocked with 2% BSA (Millipore, Schwalbach, Germany), washed and then placed under an epifluorescent microscope (Nikon Eclipse 600, Amstelveen, The Netherlands). RT-112 or TCCSUP cells were detached from the culture flask, adjusted to 1 × 106 cells/mL, and then perfused at physiologic wall shear stress (2 or 4 dyne/cm2) with a syringe pump (KDS Scientific, Holliston, MA, USA).
Tumour cell interaction with collagen was visualized in real time using a charge coupled device (CCD) camera (Lumenera, Ottawa, Canada) with a temporal resolution of 26 frames per second. The entire period of perfusion with at least five randomly selected fields of view was recorded in real time and the images were stored directly on a hard disk drive. The number of firmly adherent cells was computed by semi-automatic image processing as has been described. 9 Cells were defined as firmly adherent, if no movement greater than the cell diameter occurred within the 10-s observation period.
Integrin surface expression
Bladder cancer cell lines were washed in blocking solution (PBS, 0.5% BSA) and then incubated for 60 min at 4℃with phycoerythrin (PE)-conjugated monoclonal antibodies directed against the following integrin subtypes: Anti-α3 (IgG1; clone C3II.1), anti-α5 (IgG1; clone IIA1), anti-β1 (IgG1; clone MAR4), anti-β3 (IgG1; clone VI-PL2) or anti-β4 (IgG2a; clone 439–9B; all: BD Biosciences, Heidelberg, Germany). Integrin expression of the tumour cells was then measured using a FACscan (Becton Dickinson; FL-2H (log) channel histogram analysis; 1 × 104 cells/scan) and expressed as mean fluorescence units. A mouse IgG1-PE (MOPC-21) or IgG2a-PE (G155–178; all: BD Biosciences) was used as an isotype control.
Western blot analysis
To explore the cytoplasmic integrin level, tumour cell lysates were applied to a 7% polyacrylamide gel and electrophoresed for 90 min at 100 V. The protein was then transferred to nitrocellulose membranes. After blocking with non-fat dry milk for 1 h, the membranes were incubated overnight using the monoclonal antibodies listed above. HRP-conjugated goat-anti-mouse IgG (Upstate Biotechnology, Lake Placid, NY, USA; dilution 1:5.000) served as the secondary antibody. The membranes were briefly incubated with ECL detection reagent (ECL™, Amersham/GE Healthcare, Munich, Germany) to visualize the proteins and then analysed by the Fusion FX7 system (Peqlab, Erlangen, Germany). β-actin (1:1000, Sigma) served as the internal control.
Statistics
All experiments were performed three to six times. Statistical significance was investigated by the Wilcoxon–Mann–Whitney-U-test. Differences were considered statistically significant at a P value less than 0.05.
Results
VPA diminishes bladder cancer cell adhesion under flow
VPA significantly blocked the adhesion of both RT-112 and TCCSUP cells, compared to the untreated controls. The blocking effect was investigated at 2 and 4 dyne/cm2 (Figure 1(a) and (b)), and the tumour cell response was similar with 0.5 mM and 1 mM VPA. At 2 dyne/cm2, adhesion was reduced to a higher extent when tumour cells were treated with VPA three times a week, compared to the two times/week protocol. This was also true when TCCSUP (but not RT-112) cells were subjected to a shear force of 4 dyne/cm2. The trypan dye assay showed that the applied drug dosage did not cause toxic effects.
Adhesion kinetics of TCCSUP and RT-112 cells under a shear flow of 2 dyne/cm2 (1a) or 4 dyne/cm2 (1b). Cells were treated trice weekly with 0.5 or 1 mM VPA over a period of two weeks. Controls were untreated. Cells were counted for 10 s. Each mark on the x-axis represents a 10-s interval. One representative experiment of six is shown. *indicates significant difference to controls. SDintra-assay < 15%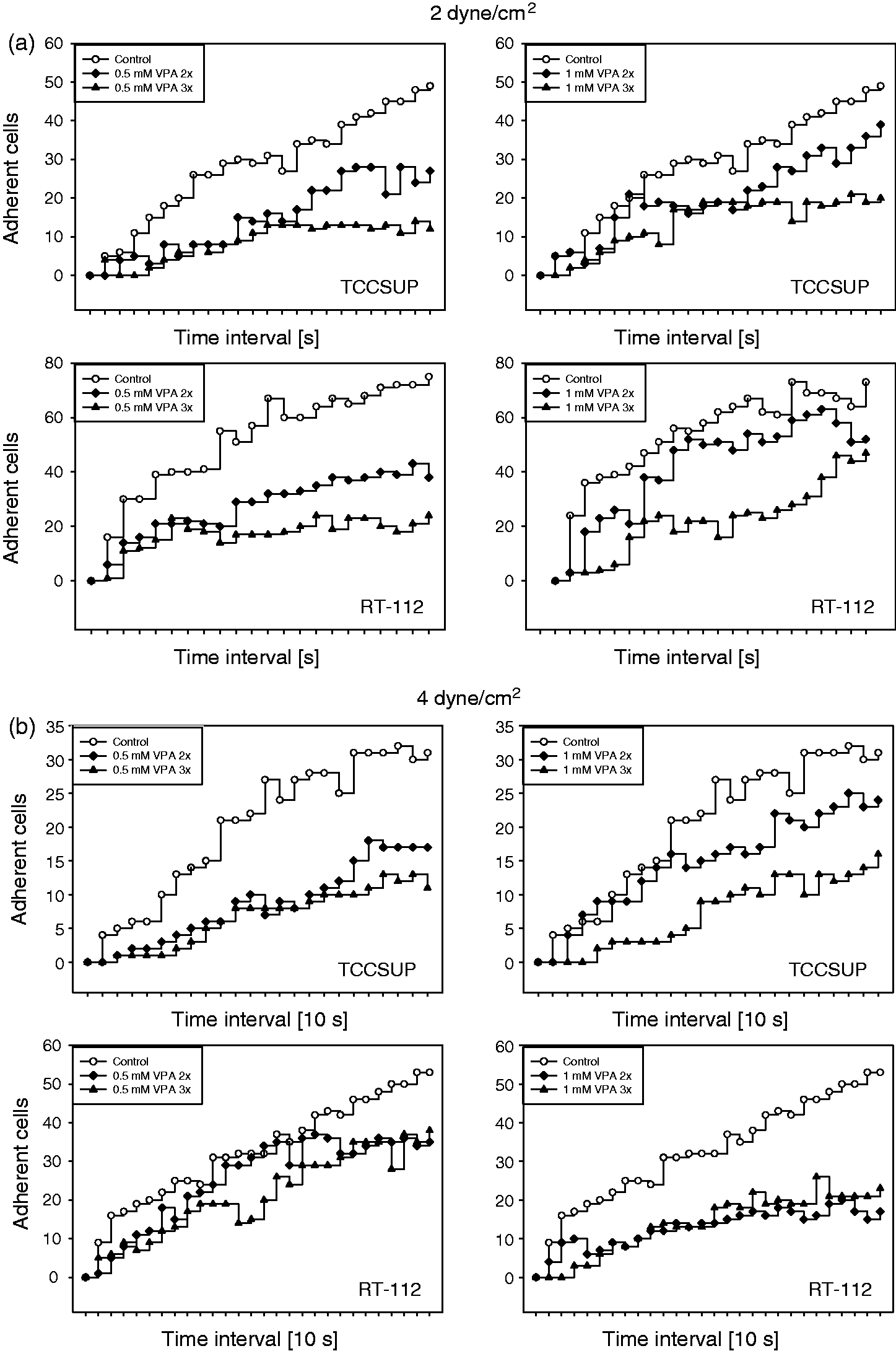
VPA modifies integrin surface expression
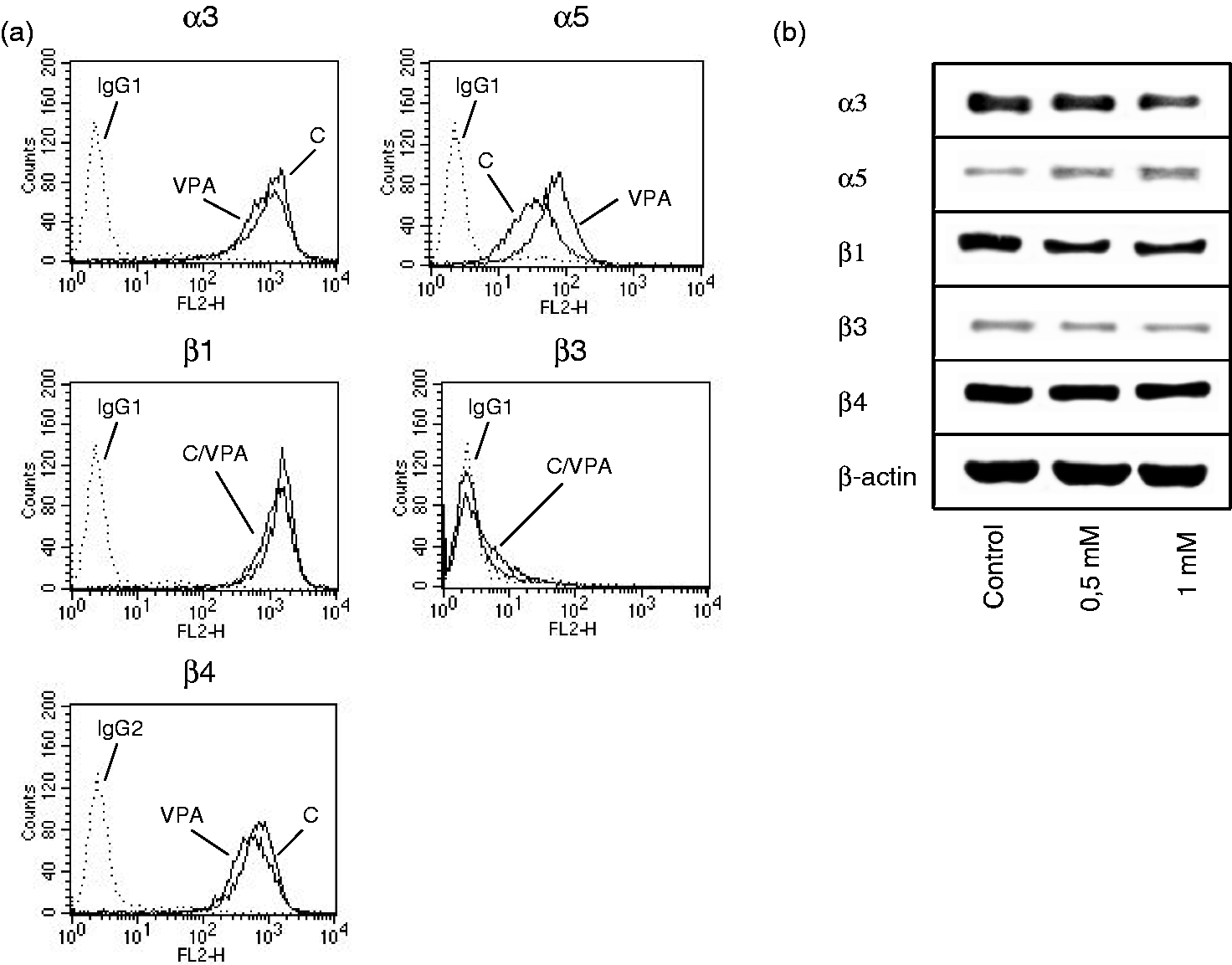
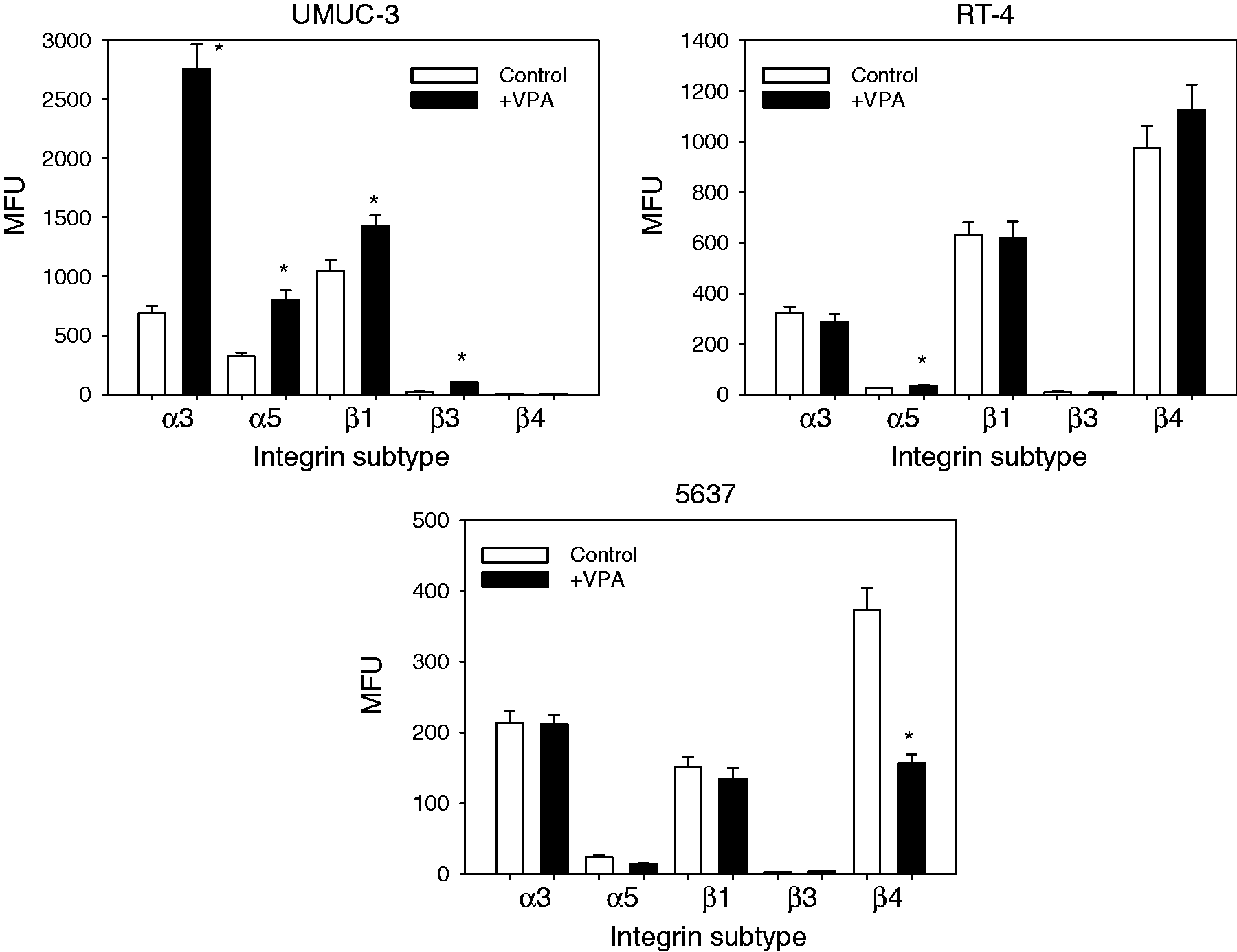
The integrin subtypes α3 and β1 were highly expressed on TCCSUP cells, with integrin α5 and β4 expressed at a minor level. The β3 type was only moderately present on the TCCSUP cell surface (Figure 2a). Both 0.5 and 1 mM VPA, applied three times a week, caused significant up-regulation of all integrin members, α3, α5, β1, β3 and β4, on TCCSUP cells (Figure 2a, representative for 1 mM VPA). The RT-112 cell line expressed the integrin subtypes in the order β1 > α3 > β4 > α5. Integrin β3 was not detected on the cellular membrane by flow cytometry (Figure 3a). VPA of 0.5 or 1 mM for the three times/week protocol significantly elevated α5 on RT-112 cells, whereas α3, β1 and β4 were slightly diminished compared to the controls (Figure 3a, representative for 1 mM VPA). To gain a more comprehensive view of the role of VPA influence on integrins in bladder cancer, additional experiments were carried out on three tumour cell lines derived from increasing grades of bladder cancer. All cell lines strongly expressed α3 and β1 integrins. The α5 and β3 levels in UMUC-3 were similar to those in TCCSUP cells. A minimal α5 level was detected in RT-4 and 5637 cells. Integrin β3 was also moderately expressed in the RT-4 cell line, but undetectable in 5637 cells. Inversely, the β4 level was high in 5637 and RT-4 but not in UMUC-3 cells. Analogous to the behaviour of TCCSUP cells, VPA caused a distinct elevation of α3 (+298.6 ± 66.8%), α5 (+146.9 ± 37.3%), β1 (+36.1 ± 9.3%) and β3 (+302 ± 87.5%) in UMUC-3 cells. VPA also elevated α5 in the RT-4 cell line (+34.9 ± 8.2%). In 5637 cells, VPA potently diminished β4 by −58.1 ± 11.3% (Figure 4).
(a) FACS analysis of integrin α and β subtype expression on treated versus non-treated TCCSUP cells. Cells were washed in blocking solution and then stained with specific monoclonal antibodies as listed in the Materials and methods section. To evaluate background staining of PE conjugated antibodies, goat anti-mouse IgG1-PE or IgG2a-PE was used (dotted lines). Fluorescence was analysed using a FACScan flow cytometer. One of three independent experiments. (b) Modification of intracellular integrin protein level. Lysates of treated versus non-treated TCCSUP cells were subjected to SDS-PAGE and blotted on the membrane incubated with respective monoclonal antibodies. β-actin served as the internal control. The figure shows one representative from three separate experiments. (c) Integrin β1-distribution in non-treated TCCSUP (control) versus TCCSUP cells treated for two weeks with 1 mM VPA. Tumour cells were stained with an FITC-marked anti-integrin β1 antibody and the integrin localization evaluated microscopically (×100 magnification) (a) FACS analysis of integrin α and β subtype expression on treated versus non-treated RT-112 cells. Cells were washed in blocking solution and then stained with specific monoclonal antibodies as listed in the Materials and methods section. To evaluate background staining of PE conjugated antibodies, goat anti-mouse IgG1-PE or IgG2a-PE was used (dotted lines). Fluorescence was analysed using a FACScan flow cytometer. One of three independent experiments. (b) Modification of intracellular integrin protein level. Lysates of treated versus non-treated RT-112 cells were subjected to SDS-PAGE and blotted on the membrane incubated with respective monoclonal antibodies. β-actin served as the internal control. The figure shows one representative from three separate experiments FACS analysis of integrin α and β subtype expression on treated versus non-treated UMUC-3, RT-4 and 5637 cells. Cells were washed in blocking solution and then stained with specific monoclonal antibodies as listed in the Materials and methods section. To evaluate background staining of PE conjugated antibodies, goat anti-mouse IgG1-PE or IgG2a-PE was used. Fluorescence was analysed using a FACScan flow cytometer. Data are depicted as mean fluorescence units (MFU). One of three independent experiments. *indicates significant difference to controls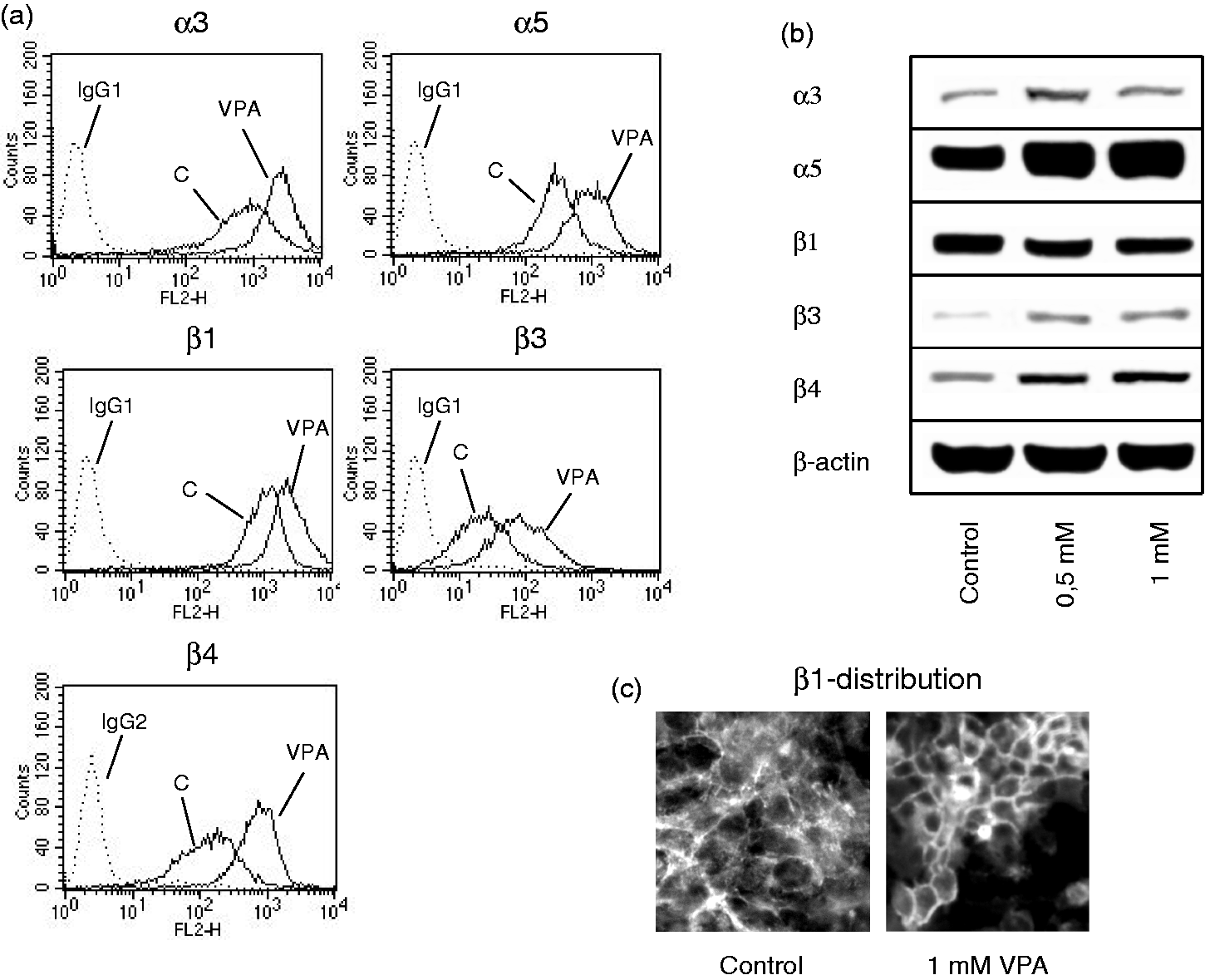
VPA acts on the integrin protein level
The integrin α3 protein was enhanced by VPA (0.5 mM > 1 mM) in TCCSUP cells. The same action was induced by 0.5 or 1 mM VPA on the α5, β3 and β4 proteins. In contrast, the integrin β1 content was reduced following VPA exposure (Figure 2b). Fluorescence analysis revealed a VPA-triggered translocation of β1 from the cytoplasm to the cell surface (Figure 2c). The α5 integrin was also found to be up-regulated, and the β1 integrin to be reduced in RT-112 cells under VPA (Figure 3b). However, in contrast to TCCSUP, VPA triggered the down-regulation of cytoplasmic α3 and β3 in RT-112 cells, and β4 was not modified at all by VPA.
Discussion
Cell adhesion is a fundamental process during the formation of metastases. High risk superficial and, more often, muscle-invasive bladder cancers bear significant metastatic potential, despite radical surgical treatment. 10 Therefore, patients with advanced bladder cancer may particularly benefit from a therapeutic strategy that aims to stop tumour dissemination. No data are available dealing with the relevance of HDAC-suppression in bladder cancer metastasis. However, growth-blocking properties of several HDAC-inhibitors have been documented, pointing to the potential clinical importance of this substance class. Evidence is provided here that the HDAC-inhibitor VPA blocks bladder cancer attachment.
Receptors of the integrin family contribute directly to adhesive interaction of tumour cells with vessel walls and extracellular matrix proteins. Therefore, the integrin expression pattern in VPA-treated versus non-treated bladder cancer cells was investigated. A congeneric integrin entity in the different cell lines could not be detected, with β3 integrin being present in TCCSUP, UMUC-3 and RT-4 but not in the RT-112 and 5637 cell lines. β4 integrin was present in all cell lines, excepting UMUC-3. The lack of common integrin entities has also been reported in UMUC, T24, J82, RT-4, 253J and Hu456 cells.11,12 Integrin expression could then vary among bladder cancer patients. Furthermore, individual dynamic alteration in the integrin expression profile may come about during the process of tumour dedifferentiation. 13 Knowledge of this process is crucial since it has recently been demonstrated that the tumour cell’s response to drug treatment depends on its integrin composition. 14 Therefore, the efficacy of therapeutic intervention may, at least partially, depend on the tumour integrin status.
The integrin members α3, α5, β1, β3 and β4 were all up-regulated in TCCSUP by VPA. The role of α3 integrin in bladder cancer has been discussed controversially. Treatment of T24 cells with a blocking antibody against α3 has resulted in the inhibition of adhesion to an endothelial cell monolayer, 15 whereas the same protocol elevated T24 binding to a collagen matrix. 16 Immunohistochemical analysis of cancerous tissue has revealed a loss of α3 during the progression from low (non-invasive) to high stage (invasive) bladder carcinoma. 11 The adhesion of TCCSUP to endothelium has not been evaluated and, therefore, no assessment can be made about how α3 contributes to endothelium-TCCSUP interaction. Speculatively, α3 strengthens adhesive forces between bladder cancer cells and vascular endothelium, preventing further movement towards matrix proteins. Whatever the precise mechanism is, evidence is provided here that the VPA-induced enhancement of α3 prevents tumour–matrix interaction, which may suppress the invasive activity of the tumour cells.
Elevation of the α5 integrin on the TCCSUP, RT-112, UMUC-3 and RT-4 surface, induced by VPA, is difficult to interpret. Saito and coworkers assumed that the presence of α5 contributes to a more malignant phenotype in transitional cell carcinoma, 17 whereas others have not detected α5 integrins at lower cancer stages, and less than 10% of high stage invasive tumours were found to be α5 positive. 11 In vitro blocking α5 forced the adhesion of bladder cancer cells to immobilized matrix proteins. 16 Based on our in vitro results, α5 integrin expression seems to be inversely correlated with bladder cancer adhesion. It may, therefore, be concluded that a therapeutic increase of α5 may slow metastatic tumour progression. In fact, four out of five bladder cancer cell lines responded well to VPA in terms of integrin α5 up-regulation, supporting this supposition.
The β1 integrin surface expression positively correlated with adhesion of 5637 1 8 and T24 bladder cancer cells 15 to an endothelial monolayer. Recently, β1 integrin blockade has been shown to diminish 5637 1 8 and T24 attachment to fibronectin. 19 A decrease of β1 was found in 5637 cells treated with VPA, which corroborates these publications. Nevertheless, this may contradict findings on TCCSUP cells, where a high β1 integrin surface level induced by VPA was associated with a low attachment rate. When discussing the relevance of integrins in tumour recurrence and malignancy, it should be kept in mind that integrin receptors serve as mechanistic binding as well as differentiation triggering elements. Therefore, up-regulation of the same integrin type might either lead to enhanced cell adhesion by coupling the receptor to its ligand or to reduced cell adhesion by activating integrin-driven differentiation signals. Reduced cell adhesion by activated integrin has recently been postulated to be induced in prostate cancer cells where membranous integrin β1 up-regulation was coupled to adhesion loss.20,21 Therefore, VPA’s action on the β1 integrin in TCCSUP cells could cause receptor elevation, triggering tumour cell re-differentiation, which is associated with a low invasive phenotype.
However, β1 integrin alterations caused by VPA were not consistent. The molecule was up-regulated along the surface of TCCSUP cells, but down-regulated intracellularly. Novel findings suggest that β1 internalization contributes to tumour metastasis and, conversely, β1 translocation from the cytoplasm to the cell membrane may prevent tumour cell migration. 22 Since fluorescence analysis demonstrated β1 integrin translocation from the cytoplasm to the TCCSUP cell surface under VPA, it may be assumed that VPA accelerates dynamic integrin β1 turnover in this cell type, which may be coupled to diminished adhesion.
The β3 molecule has been shown to be a proactive mediator of apoptosis in a bladder cancer in in vitro models. 23 β3 integrin has also been shown to suppress some cancers, and patients with β3-integrin-overexpressing tumours are significantly more disease-free and their overall survival 24 time is increased. A similar observation has been made with respect to integrin β4, as a decrease in β4 was significantly related to an increase in tumour grade and deep invasion.11,25 In vitro, β4 appears to inhibit growth and migration in bladder carcinoma cell lines.26,27 The present results suggest, therefore, that increased expression of β3 and β4 evoked by chronic treatment with VPA might slow the adhesion dynamics of TCCSUP cells. In a recent publication, long-term use of VPA has been reported to stop TCCSUP cell growth. 7 Whether β3 is involved in this process, however, cannot yet be answered.
Since VPA differentially modified the integrin expression pattern on the five bladder cancer cell lines analysed here, its molecular mode of action may vary with the tumour phenotype. Since the strongest intervention was seen in the high grade UMUC-3 and TCCSUP cells, it may be argued that VPA’s effects correlate with tumour staging and grading. However, there is also evidence that VPA’s action correlates with the integrins present on the tumour cell membrane, complicating the concept, since UMUC-3 cells were found to be integrin β4 negative, whereas TCCSUP cells were integrin β4 positive. In this context, exposing VPA to androgen-sensible LNCaP versus androgen-resistant PC-3 prostate cancer cells resulted in a dissimilar alteration of integrin subtype expression. The subtype expression depended, at least partially, on the initial integrin constellation of the particular cell line, 14 whereby PC-3 was β3 integrin positive and LNCaP β3 integrin negative. Since TCCSUP and RT-112 have been characterized by a different integrin profile as well, with β3 being positive in TCCSUP but negative in RT-112, a similar scenario as that observed with PC-3 and LNCaP cells may have become apparent in the present study, as well.
VPA has been shown here to block bladder cancer cell interaction with collagen under flow conditions. Together with the evidence of growth blocking properties of VPA, HDAC-inhibition might become an option in treating advanced bladder cancer. Nevertheless, due to the different molecular influence of VPA on different cell lines, a subgroup of patients may particularly benefit from an HDAC-inhibitor based therapy. In this context, the heterogeneous effect of VPA on integrin expression will need further studies.
Footnotes
Author contributions
All authors participated in the design, interpretation of the studies and analysis of the data and review of the manuscript; EJ, SMS and TS performed all major experimental work of the study. JM and LH carried out western blotting. RAB and CW contributed to the conception, design and coordination of the study and drafted the manuscript. GB and AH participated in the conception of the study and data interpretation and helped to revise the manuscript. All authors have read and approved the final manuscript. The corresponding author has verified that all individuals who made contributions to this study are included either as authors or are acknowledged at the end of the paper.
Acknowledgements
We would like to thank Karen Nelson for critically reading the manuscript. This work was supported by the ‘Alfons und Getrud Kassel-Stiftung’.
*
Contributed equally to this work.
†
Contributed equally as senior authors.