
Abstract
The mouse is a quickly reproducing, inexpensive animal and often used for transgenic approaches. Due to its small size, only the aorta is frequently taken to assess vascular function. However, atherosclerosis is a generalized disease and becomes symptomatic when the perfusion of specific organs is impaired. We have therefore compared the thoracic and abdominal aorta with carotid, femoral, mesenteric, renal and coronary arteries to see whether aortic vasomotion can indeed serve as a surrogate for other, organ-specific vascular territories. Arterial segments of male C57BL/6J mice were dissected and mounted on a myograph for isometric force measurement. Vasoconstriction was determined in response to depolarization by potassium chloride (KCl), which was not different with or without an α-adrenoceptor antagonist. Vascular responses were determined in response to receptor activation by the neurotransmitter norepinephrine (±inhibition of nitric oxide synthase; ±α- and β-adrenoceptor antagonists) and the platelet-derived mediator serotonin (±inhibition of nitric oxide synthesis; ±5-hydroxytryptamine receptor antagonist). Endothelium-dependent and -independent vasodilation was determined in response to carbachol and nitroprusside after norepinephrine-induced pre-constriction (±β-adrenoceptor antagonist). Vasoconstriction in response to KCl, norepinephrine and serotonin differed in magnitude between thoracic and abdominal aorta and between aorta and the other arterial segments. Endothelium-dependent and -independent vasodilation differed also in magnitude between the arterial segments. Thus, the murine aorta is not a general surrogate to assess vascular function of organ-specific vascular territories.
Introduction
Mice are the most commonly used mammals in cardiovascular research because of their availability, small size, low cost, ease of handling and fast reproduction rate. Mice with cell-, tissue- and pathway-specific transgenic modifications are available. C57BL/6J is the most widely used mice strain. 1
Atherosclerosis is a generalized disease. However, atherosclerosis becomes only symptomatic when perfusion of a specific organ is impaired: in the coronary arteries to cause myocardial ischemia or myocardial infarction, in the femoral arteries to cause peripheral arterial disease, in the renal arteries to cause hypertension and renal failure, in the aorta to cause aneurysms or increased aortic stiffness, in the carotid arteries to cause stroke and in the mesenteric arteries to cause angina abdominalis. For studies of vascular function in mice, however, only the aorta is frequently used, 2 –5 assuming that the murine aorta is a reasonable surrogate for all vascular territories. In fact, the vasomotion of different vascular territories is heterogeneous. 6 –11 Therefore, we here compared the thoracic aorta, abdominal aorta, carotid, femoral, mesenteric, renal and coronary arteries to see whether and possibly under which conditions aortic vasomotion can indeed serve as a surrogate for other, organ-specific vascular territories. We focus on the responses to the humoral vasoconstrictors norepinephrine and serotonin, and on carbachol as endothelium-dependent and nitroprusside as endothelium-independent vasodilators.
Materials and methods
Mice and dissected arterial segments
Male C57BL/6J mice (5.4 ± 0.1 months; 28.3 ± 0.8 g) were sacrificed under anaesthesia with enflurane, and their hearts were rapidly removed. Thoracic aorta, abdominal aorta, carotid arteries, femoral arteries, mesenteric arteries, renal arteries and coronary arteries were carefully dissected and immediately placed into carbogenated (5% CO2; 95% O2) Krebs–Henseleit buffer (mmol/L: 119 NaCl, 4.7 KCl, 2.5 CaCl2 × 2 H2O, 1.17 MgSO4 × 7 H2O, 25 NaHCO3, 1.18 KH2PO4, 0.027 EDTA, 5.5 glucose). After removal of perivascular tissue, vessels were perfused with buffer or fixed in formalin. For analysis of vascular morphology, the dissected vessel length was retained, and for vasomotor measurements, vessels were cut into segments of 2 mm length and suspended in an isometric small vessel myograph (Danish Myo Technology, Aarhus, Denmark).
Vascular morphology
Inner and outer circumference of arterial segments
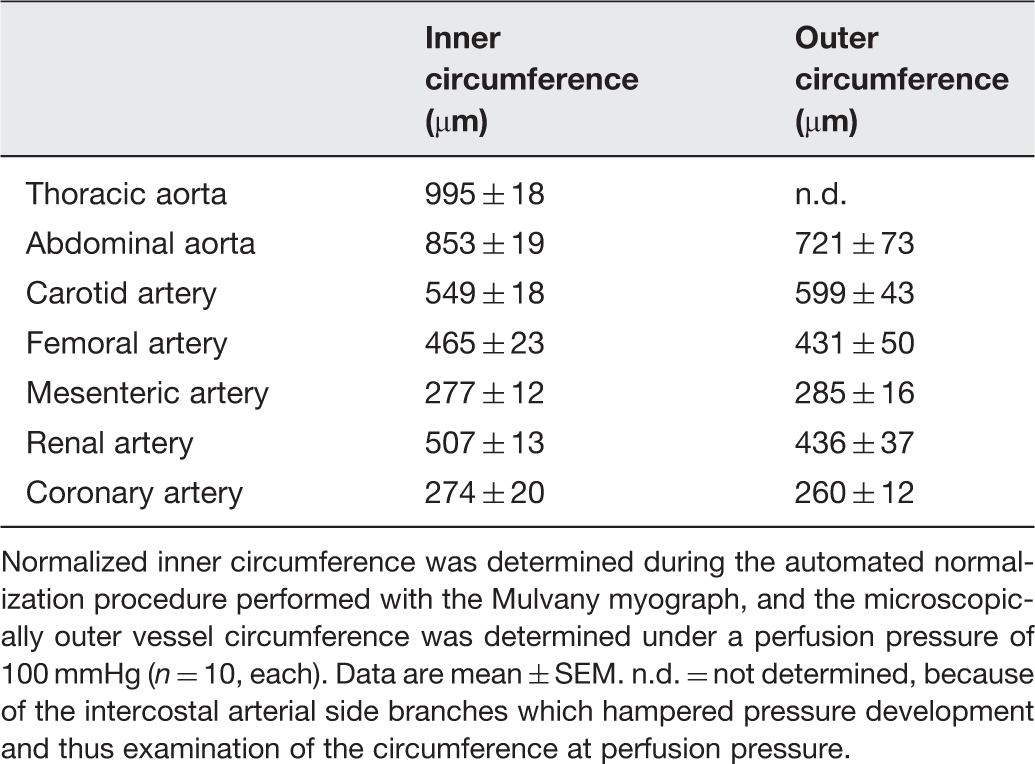
Normalized inner circumference was determined during the automated normalization procedure performed with the Mulvany myograph, and the microscopically outer vessel circumference was determined under a perfusion pressure of 100 mmHg (n = 10, each). Data are mean ± SEM. n.d. = not determined, because of the intercostal arterial side branches which hampered pressure development and thus examination of the circumference at perfusion pressure.
For histological examination, vessels were stabilized by introduction of a stainless steel wire (40 µm) and fixed in 4.5% formalin for 24 h. Vessels were embedded in paraffin ((1) seven incubation steps in ethanol from 70% to 100% for 1 h each, (2) xylene incubation for 1 h twice and (3) two subsequent steps in paraffin at 65℃for 2 h each) with an automatic tissue processor (TP 2010 Leica, Wetzlar, Germany). Vessels were embedded in paraffin blocks (Embedding Center EG 1140, Leica, Wetzlar, Germany), cut into 4 µm slices with a microtome (RM2155, Leica, Germany) and mounted onto polylysine-coated slides (R. Langenbrinck, Emmendingen, Germany). Slices were malted (at 65℃for 1 h) and dewaxed in xylene. The xylene incubation (twice for 5 min) was followed by rehydration in ethanol (five incubation steps from 100% to 70% for 2 min each). The staining procedure was started after rinsing with water. Elastica van Gieson staining kit (Merck KGaA, Darmstadt, Germany) was used according to the manufacturer’s protocol. After rinsing with water, elastin-solution according to Weigert was incubated for 10 min. Weigert’s A- and B-solutions (1:1) were added for 5 min. After rinsing with water, picrofuchsin solution was incubated for 2 min. After a standard dehydration procedure ((1) seven incubation steps in ethanol from 70% to 100% for 1 h each, (2) xylene incubation for 1 h twice) slides were covered with a mounting medium (CV mount, Leica, Nussloch, Germany) and a glass plate.
To estimate the proportion of elastic fibres per arterial segment, the Elastica van Gieson-stained colour picture slices were converted into black/white pictures (CorelDRAW® Graphics Suite 12, Fremont, USA), and their relative area (%) per tunica-intima-media area was calculated.
Vasomotor assay
Arterial segments were mounted onto two stainless steel wires (40 µm in diameter, for coronary and mesenteric arteries 25 µm in diameter) which were connected to a force transducer and a micrometer, respectively. Vasoconstriction was measured as active wall tension (mN). Arteries were equilibrated in carbogenated Krebs–Henseleit buffer at 37℃before an automated normalization procedure was performed. This normalization is controlled from the interface using a standardized procedure according to the manufacturer’s protocol. The normalization uses an approximation of the lumen diameter (d100) which the artery would have had in vivo, when relaxed and subject to a transmural pressure of 100 mmHg, and the Laplace law for vessels with infinitely thin walls: P = 2 T/d, where P is transmural pressure, T is wall tension and d is lumen diameter. The arteries were then set to a lumen diameter d = 0.9 × d100, where active force development is maximal. This normalized inner circumference was read and compared to the outer circumference determined microscopically (Table 1). The active force was divided by twice the arterial segment length (AD converter: PowerLab8/30, software: LabChart6, ADInstruments GmbH, Spechbach, Germany). The vessels were equilibrated for a further 30 min with frequent buffer changes. Maximal vasoconstriction was induced repetitively via depolarization with potassium chloride (KCl initially; 0.6 × 10−1 mol/L and then 1.2 × 10−1 mol/L over 15 min each). Between KCl-exposures, vessels were washed with frequent buffer changes until baseline force was reached. The maximal vasoconstrictor response to KCl of all arterial segments, which were subsequently used for the different experimental protocols to determine vasoconstrictor and vasodilator response, was comparable.
To determine the potential influence of sympathetic innervation of the arteries on their vasoconstrictor response to KCl, in a subset of experiments, maximal vasoconstriction induced by KCl 1.2 × 10−1 mol/L was determined after pre-incubation (30 min) of the vessels with the α-adrenoceptor antagonist phentolamine (10−4 mol/L) or placebo, respectively.
Maximal force development (ΔmN) at automatically determined and manually chosen vessel diameter
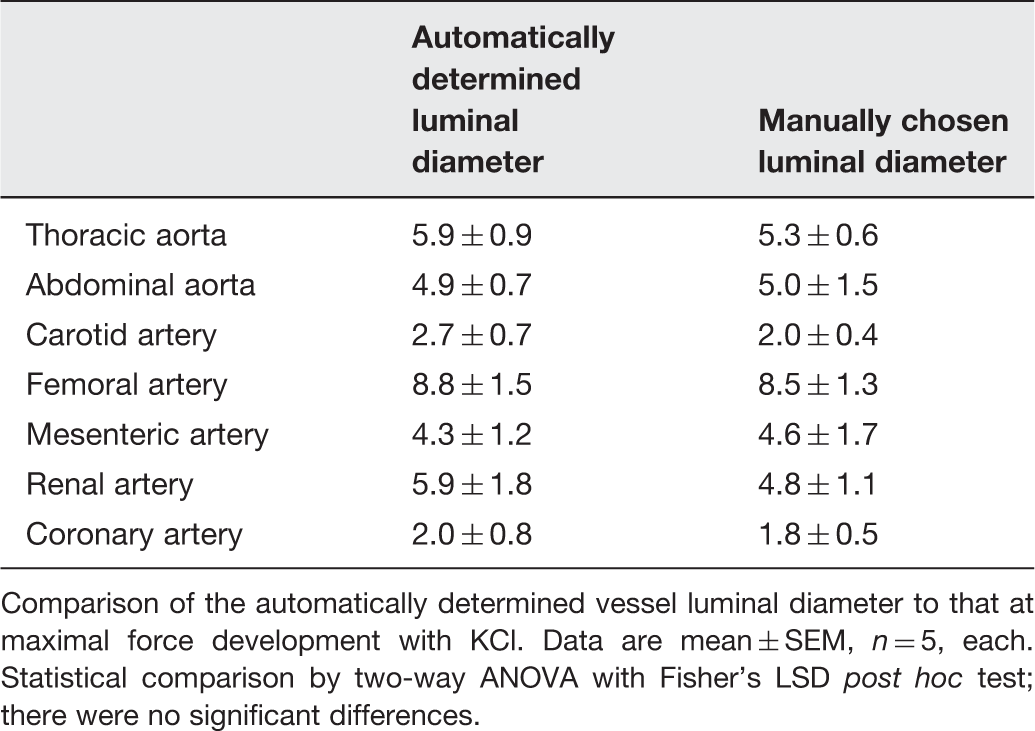
Comparison of the automatically determined vessel luminal diameter to that at maximal force development with KCl. Data are mean ± SEM, n = 5, each. Statistical comparison by two-way ANOVA with Fisher’s LSD post hoc test; there were no significant differences.
Experimental protocols
Cumulative concentration–response curves were determined in response to 1 × 10−9 mol/L–1 × 10−4 mol/L for norepinephrine and serotonin. The effect of nitric oxide synthase (NOS) activity and the role of α- and β-adrenoceptors in the vasoconstriction by norepinephrine was determined by pre-incubation (30 min each) of the vessels with the NOS-inhibitor L-NG-nitroarginine methyl ester (L-NAME, 10−4 mol/L), the α-adrenoceptor antagonist phentolamine (10−4 mol/L), the β-adrenoceptor antagonist propranolol (2 × 10−5 mol/L) and their combination (propranolol with L-NAME or phentolamine, respectively). The effect of NOS activity and the role of serotonin receptors in the vasoconstriction by serotonin were determined by pre-incubation (30 min, each) of the vessels with the NOS-inhibitor L-NAME (10−4 mol/L) or the 5-hydroxytryptamine (5HT)2 receptor antagonist ketanserin (10−6 mol/L).
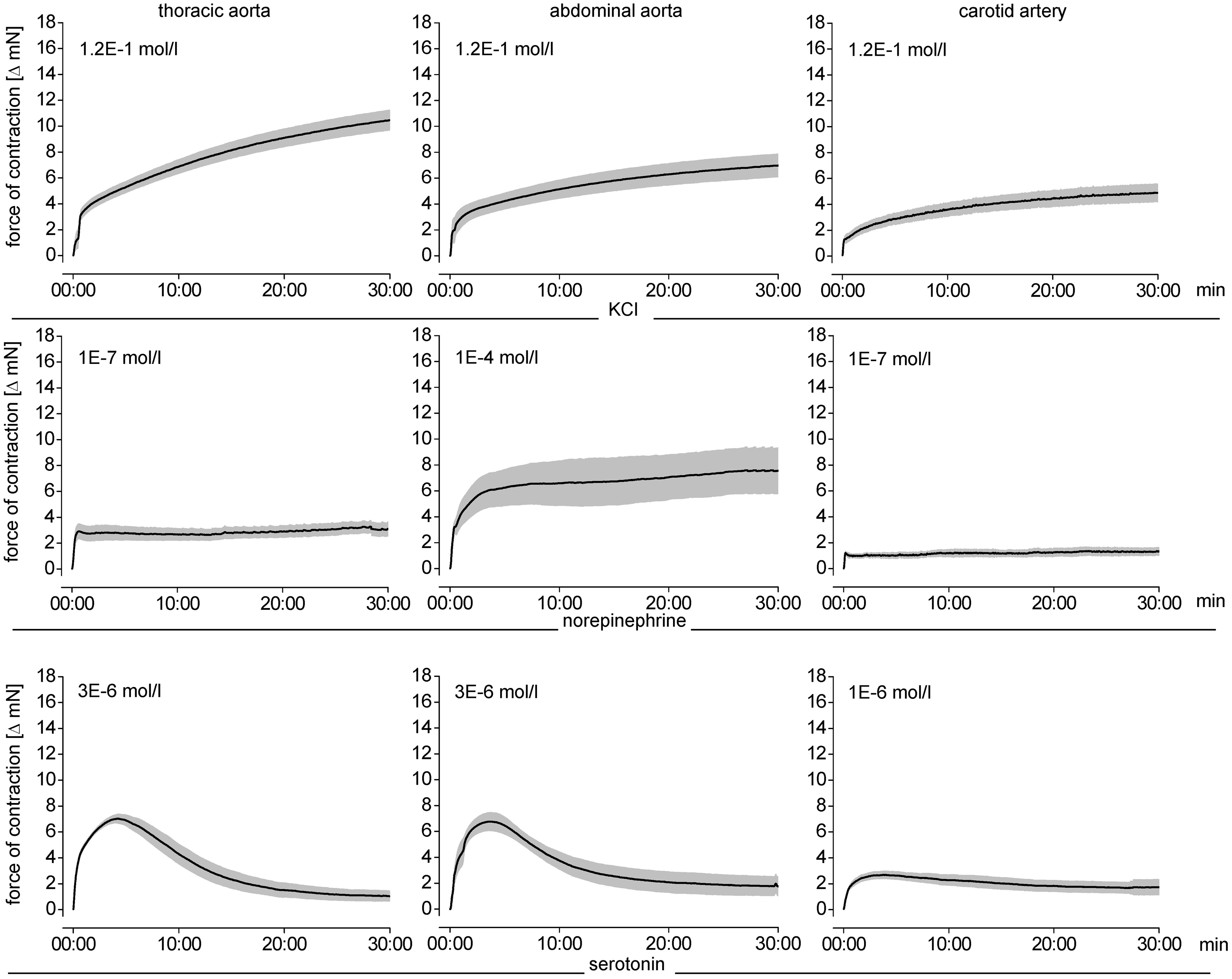
Endothelium-dependent and -independent vasodilation was measured in response to carbachol and nitroprusside (1 × 10−9 mol/L–1 × 10−4 mol/L each) after pre-constriction by norepinephrine; for each arterial segment the norepinephrine concentration which caused a maximal vasoconstriction was used. Norepinephrine was used for pre-constriction because neither KCl nor serotonin induced a temporally stable vasoconstriction in all investigated vessels over the required time period, that is, 30 min (Figure 3).
For the coronary arteries, the combination of propranolol, L-NAME and norepinephrine was used, because only this combination caused a stable pre-constriction (Figure 3). Consequently, for the coronary arteries only the endothelium-independent vasodilation in response nitroprusside was determined.
Chemicals and drugs
Carbachol, nitroprusside, [-]-norepinephrine bitartrate, phentolamine hydrochloride, propranolol hydrochloride and serotonin were purchased from Sigma, Deisenhofen, Germany. Ketanserin tartrate was purchased from Tocris, R& D Systems Inc. Minneapolis, USA. L-NAME was purchased from MP biomedicals, Ohio, USA. All chemicals for the Krebs–Henseleit buffer were purchased from AppliChem, Darmstadt, Germany. Formalin (Roti Histofix) was purchased from Carl Roth, Karlsruhe, Germany. Xylene (REF 6764506) was purchased from Thermo Scientific, Kalamazoo, USA. All chemicals were of the purest grade commercially available.
Statistical analysis
Data are means ± standard error of mean (SEM). Maximal vasoconstrictor responses to KCl were compared by one-way analysis of variance (ANOVA) followed by Dunn’s multiple comparison procedure. Maximal vasoconstrictor responses to KCl between the respective arterial segments, which were subsequently used for the different experimental protocols (vasoconstriction by norepinephrine and serotonin and vasodilation by carbachol and nitroprusside) were compared by two-way ANOVA followed by Fisher’s least significant difference (LSD) post hoc tests (SigmaStat 2.03, SPSS Inc., Chicago, IL, USA).Concentration–response curves were analysed in terms of half-maximal effective concentration (EC50) after least-square sigmoidal curve fitting of individual curves using Origin 7 G SR2 (Northampton, USA); P-values less than 0.05 were considered significant.
Results and discussion
Vascular size versus strength of vasoconstriction
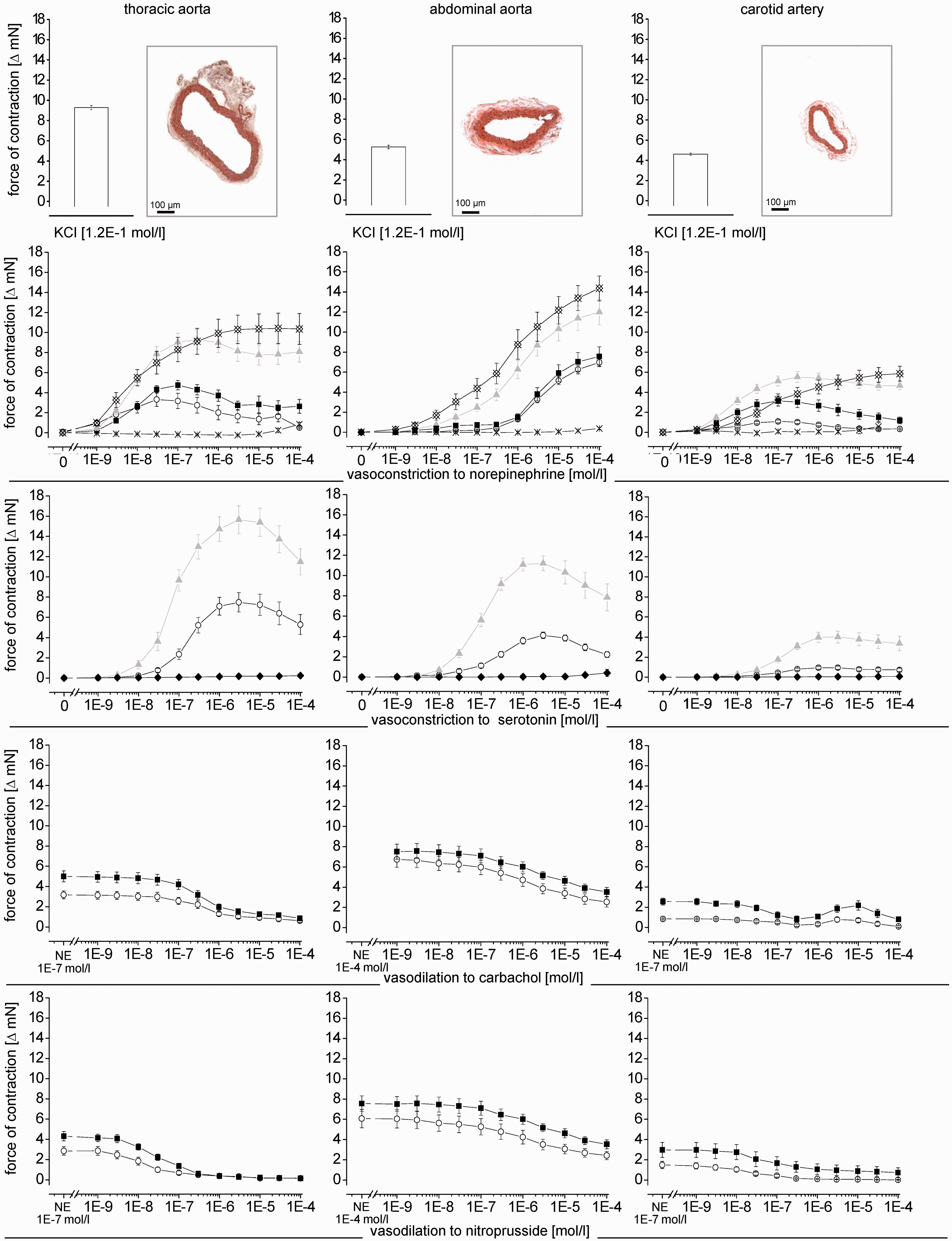
The preparation of particularly the smaller vessels required manual skills and expertise. As expected, the arterial segments differed in their size (Table 1). The number of elastic fibres in the tunica media of the elastic vessels (thoracic and abdominal aorta, carotid arteries) was higher than that in the muscular vessels (femoral, mesenteric, renal and coronary arteries) (Figure 1).
Proportion of elastic fibres in the tunica-intima-media in all arterial segments with examples of Elastica van Gieson staining (dark areas = elastic fibres) converted into black and white pictures (white areas = elastic fibres), respectively. Data are mean ± SEM of more elastic (grey bars) and more muscular arterial segments (white bars), n = 6, each. One-way repeated measures ANOVA is followed by Dunn’s multiple comparison; #
P < 0.01 vs. muscular arterial segments; *P < 0.01 vs. thoracic and abdominal aorta. (A color version of this figure is available in the online journal)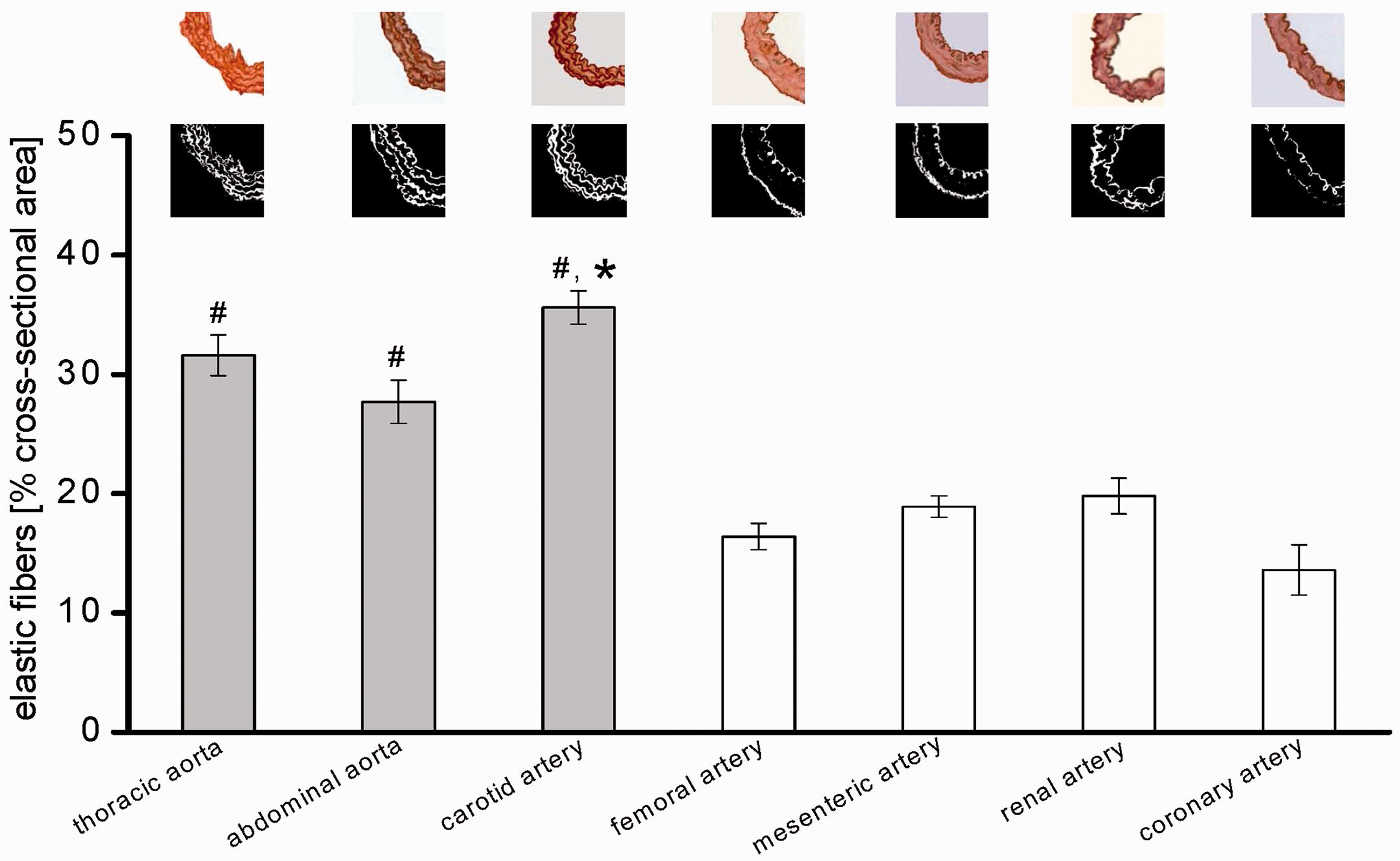
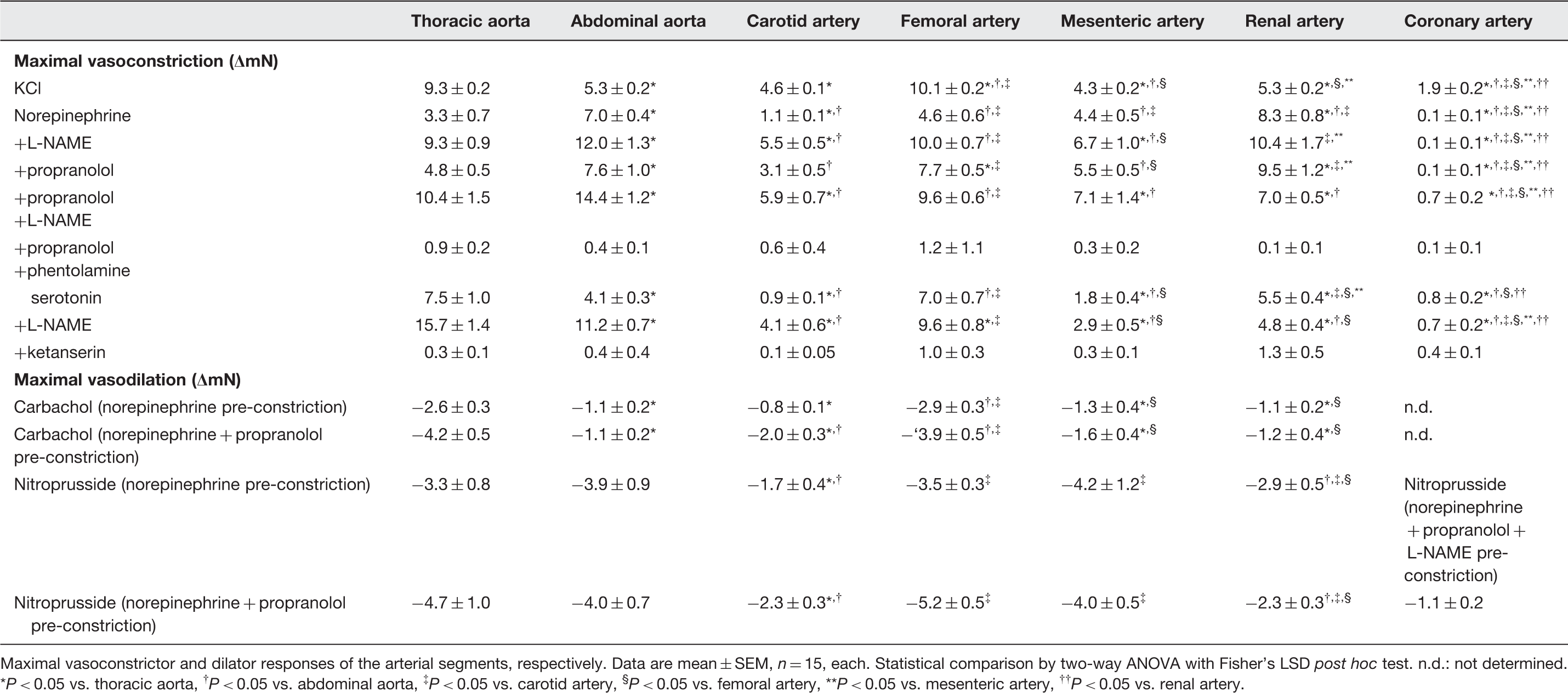
KCl, norepinephrine (with the exception of coronary arteries) and serotonin induced vasoconstriction (Table 3 and Figures 2 to 4). Different from prior studies in the pulmonary vascular territory of sheep,
12
in our hands the vasoconstriction of the different vascular territories did not depend on vessel size (Figure 4); although smaller, the femoral artery developed a force comparable to that of larger arterial segments (e.g. thoracic or abdominal aorta) in response to different stimuli (Tables 1 and 3). Our data are in line and extend those from a prior report on a stronger vasoconstriction of femoral than carotid arteries in response to phenylephrine.
6
The two elastic aortic vascular segments differed in their responses to all used vasoconstrictor stimuli (Table 3). Apparently, vascular size does not determine the magnitude of vasoconstrictor responses.
Vasoconstriction of different arterial segments in response to KCl- and KCl+ phentolamine. Data are mean ± SEM, n = 6, each Temporal stability of vasoconstriction induced by KCl, norepinephrine (for coronary arteries: norepinephrine with propranolol and L-NAME) and serotonin for each arterial segment. Data are mean ± SEM, n = 8, each Vasoconstriction and vasodilation of different arterial segments. Micrographs of the arterial segments (Elastica van Gieson staining) at 100-fold magnification. Vasoconstriction in response to KCl, norepinephrine and norepinephrine +L-NAME, or +propranolol, or +propranolol and L-NAME, or +propranolol and phentolamine and to serotonin and serotonin +L-NAME, or +ketanserin. Vasodilation in arterial segments pre-constricted with norepinephrine (abbreviated on the x-axis as: NE) ±propranolol induced by carbachol and nitroprusside. For coronary vessel segments, the vasodilation was studied with norepinephrine +propranolol +L-NAME pre-constriction. Data are mean ± SEM, n = 15, each. (A color version of this figure is available in the online journal) Maximal vasomotor responses to norepinephrine, serotonin, carbachol and nitroprusside Maximal vasoconstrictor and dilator responses of the arterial segments, respectively. Data are mean ± SEM, n = 15, each. Statistical comparison by two-way ANOVA with Fisher’s LSD post hoc test. n.d.: not determined.
P < 0.05 vs. thoracic aorta, †
P < 0.05 vs. abdominal aorta, ‡
P < 0.05 vs. carotid artery, §
P < 0.05 vs. femoral artery, **P < 0.05 vs. mesenteric artery, ††
P < 0.05 vs. renal artery.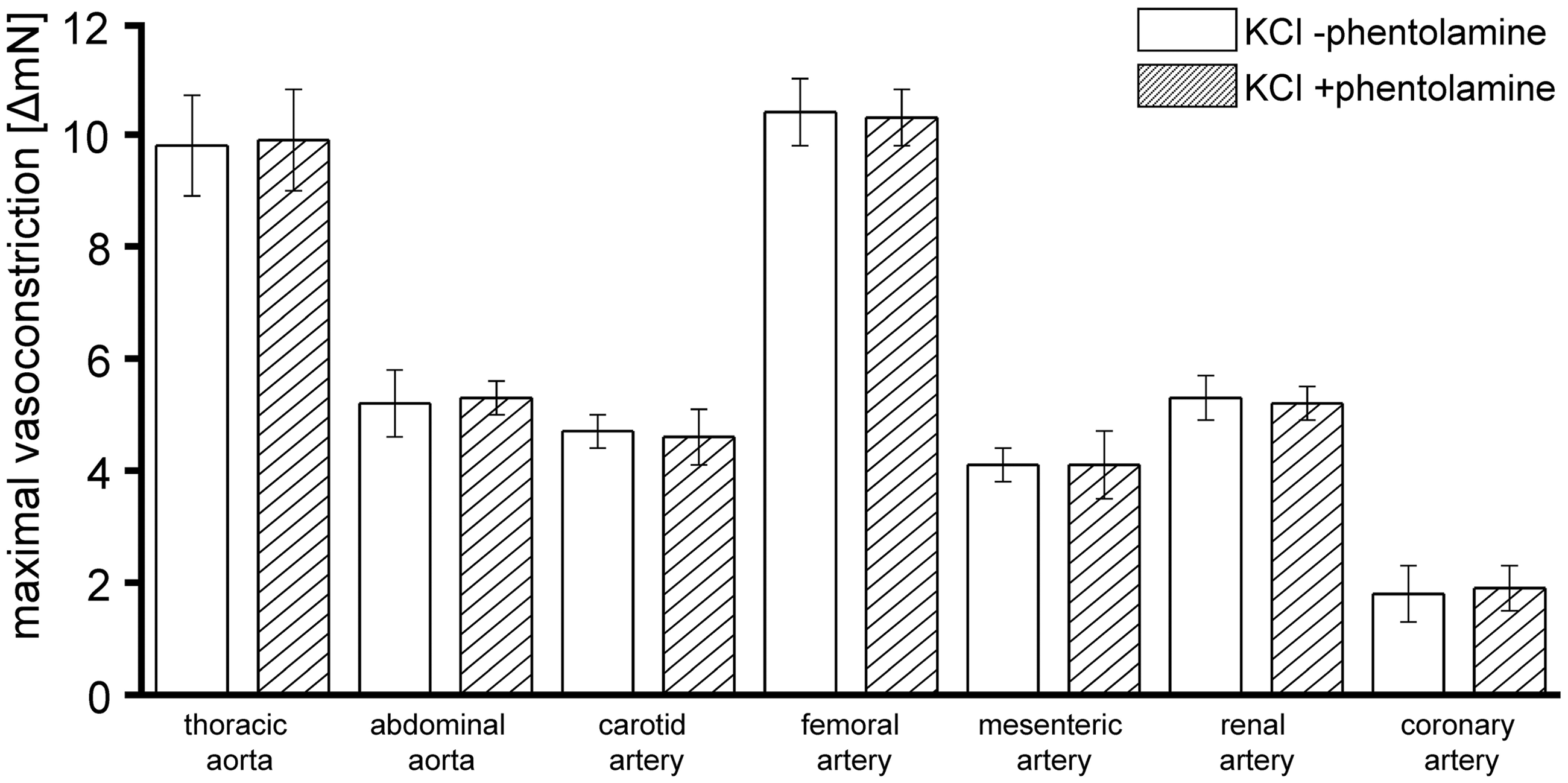
Vasoconstrictor responses in different arterial segments
Half-maximal effective concentration for vasoconstriction and vasodilation
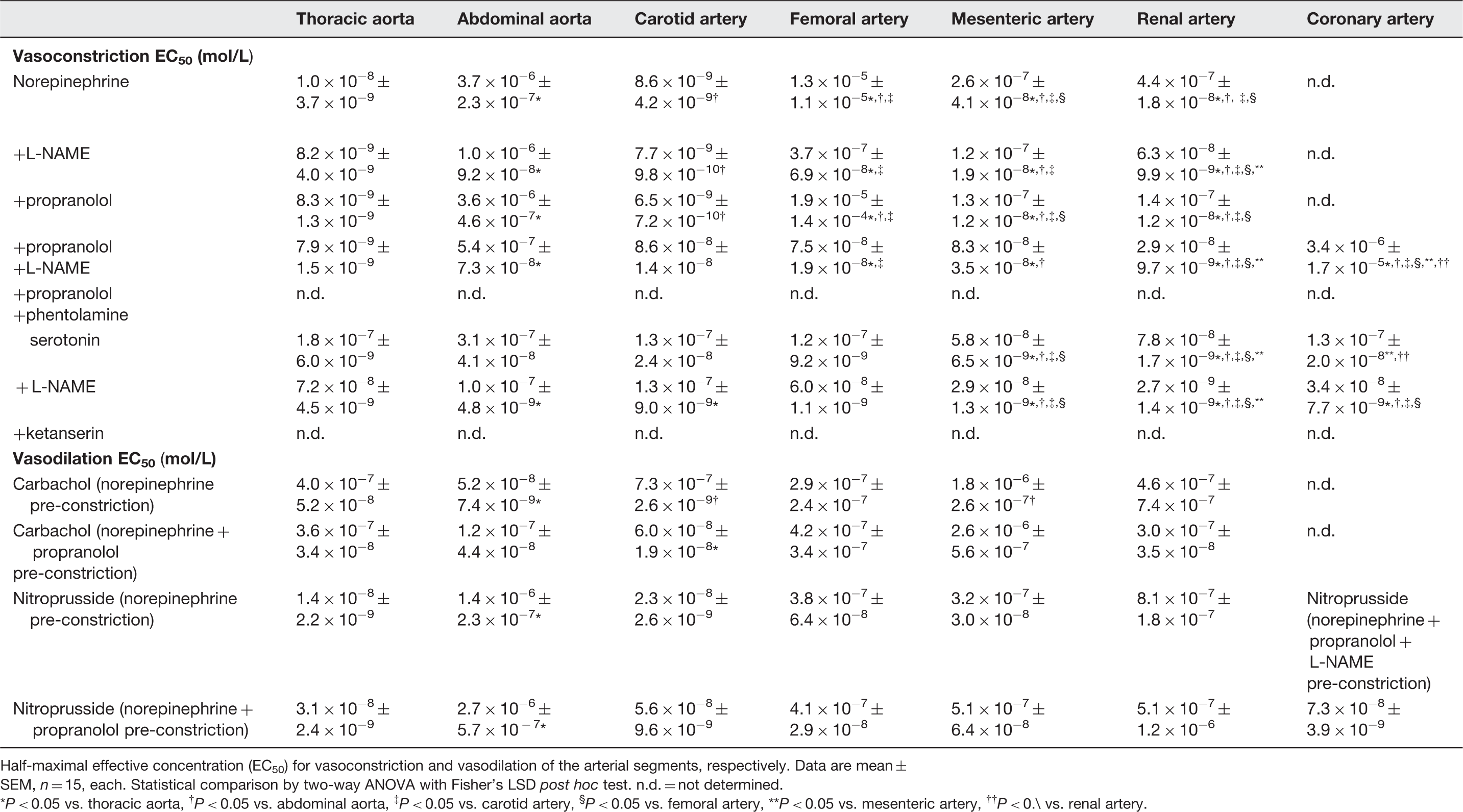
Half-maximal effective concentration (EC50) for vasoconstriction and vasodilation of the arterial segments, respectively. Data are mean ± SEM, n = 15, each. Statistical comparison by two-way ANOVA with Fisher’s LSD post hoc test. n.d. = not determined.
P < 0.05 vs. thoracic aorta, † P < 0.05 vs. abdominal aorta, ‡ P < 0.05 vs. carotid artery, § P < 0.05 vs. femoral artery, **P < 0.05 vs. mesenteric artery, †† P < 0.\ vs. renal artery.
5HT2 receptors are primarily responsible for the vasoconstrictor response to serotonin 15 and 5HT1 receptors for vasodilation. 16 However, the vasomotor response to serotonin is very heterogeneous; serotonin induces vasoconstriction in larger arteries, whereas it relaxes more downstream resistance vessels. 16 The net blood flow response to serotonin is therefore balanced by expression, localization and consequently activation of the different receptor subtypes. In our setup, serotonin caused vasoconstriction in all arterial segments which was characterized by a bell-shaped concentration–response curve, pointing indeed to an interaction of different vasomotor mechanisms (Figure 4). The vasoconstriction by serotonin was abrogated by the 5HT2 receptor antagonist ketanserin in all arterial segments, as expected (Table 3 and Figure 4).
The extent of vasoconstriction (Table 3 and Figure 4) and sensitivity to the vasoconstriction (Table 4 and Figure 4) by norepinephrine and serotonin were potentiated by NOS inhibition with L-NAME, reflecting the well-established endothelial NOS-derived nitric oxide-induced dilation in all types of vessels. 17 As expected, the NOS activity was higher in larger aortic vascular segments, 18 as reflected by the stronger counteraction of vasoconstriction than in the smaller arterial segments. The impact of NOS activity, however, also differed among smaller arterial segments (Tables 3 and 4 and Figure 4).
Apart from vascular caliber, receptor distribution and their respective downstream pathways, the different vasoconstrictor responses could be related to the different developmental origin of the smooth muscle cells from different embryonic lineages. 19 However, an obvious similarity in responses of segments from the same developmental origin, for example, abdominal aorta versus femoral artery, in comparison to a difference in responses of segments from different developmental origin, for example, mesenteric artery versus femoral artery, was not apparent.
Vasodilator responses
For comparison of the vasodilator responses of the different arterial segments, we used pre-constriction by norepinephrine because it was stable over the investigational period of 30 min (Figure 3). In the coronary arteries, norepinephrine induced a stable pre-constriction only in combination with propranolol and L-NAME (Figure 3).
The endothelial-dependent vasodilation to carbachol differed among the arterial segments. The extent and responsiveness were most intense in the thoracic aorta and weaker in mesenteric and renal arteries, in line with the literature 18 and the results during vasoconstriction with and without NOS inhibition (Tables 3 and 4 and Figure 4).
Endothelium-independent vasodilation by nitroprusside was more intense than the endothelium-dependent vasodilation by carbachol; however, even nitroprusside did not abrogate vasoconstriction completely. Vasoconstriction was almost non-reversible in the renal artery (Table 3 and Figure 4). Thus, to achieve a vasodilator response in a sensitive manner for the different arterial segments, pre-constriction cannot be used under all circumstances in all arterial segments.
Limitations and conclusions
The vasomotor response of different vascular territories is a complex and integrated process. This descriptive study on vasomotor responses in different arterial macrocirculatory vascular territories of C57BL/6J mice demonstrates that vasoconstriction and vasodilation differ between vascular territories. Here, we provide a methodological reference framework for studies on vascular reactivity in C57BL/6J mice. Each vascular bed is unique and one cannot be substituted by another one in experiments assessing vascular function and their relevance to atherosclerosis.
Our results relate only to the macrocirculation, but not the microcirculation and may be different in other mouse strains and other species. Furthermore, we have used just a few paradigmatic vasoactive substances to characterize vascular diversity. For a more detailed characterization of pharmacological vascular reactivity, a more comprehensive approach should be used. 5 Nevertheless, our data may provide a methodological reference for further use of isolated mouse vascular segments.
Footnotes
Author contributions
PK designed the study, performed data analysis and statistics and drafted the paper; AS performed vasomotor bioassays and GH supervised the study and finalized the paper. The data of the present paper are part of the MD thesis of AS.
Acknowledgements
The technical assistance of Sandra Krueger, Jelena Loeblein and Astrid Buechert is gratefully acknowledged.