
Abstract
Disruption of peripheral blood B-cell homeostasis and variation of surface receptors occur with certain infections and autoimmune diseases. However, the impact of antiviral therapy on B-cell alteration during chronic hepatitis B (CHB) infection remains unclear. Our study aims to document the effects of B-cell alteration in CHB patients treated with tenofovir or adefovir. A total of 21 CHB patients and 10 healthy donors were recruited into the study. We identified B-cell subsets by flow cytometry and observed changes in the B-cell repertoire of CHB patients upon tenofovir or adefovir antiviral treatment. The total and percent of B cells and CD5 + B-cell subsets were significantly increased in CHB patients compared to healthy donors. Total and percent of CD5 + B cells gradually decreased following the diminution of the HBV DNA load after tenofovir and adefovir treatment. Upon tenofovir treatment, the percent of memory CD27 + B cells was increased but the absolute number declined, whereas naïve CD27− B cells declined in both percent and absolute number. In the adefovir treatment group, neither naïve nor memory B cells were altered by the treatment. Furthermore, CHB patients displayed higher levels of activation markers (CD69 and CD24) and trended towards restored B-cell homeostasis after antiviral treatment. In conclusion, disrupted B-cell homeostasis is an important feature of CHB patients and is partially restored after control of viral replication by antiviral treatment. B-cell antiviral immunity is improved by restoring B-cell homeostasis and activation.
Keywords
Introduction
Chronic hepatitis B (CHB) is a global health problem affecting more than 350 million people worldwide. 1 Hepatitis B virus (HBV) infection persists in 1–5% of infected adults, 20–30% of young children, and up to 90% of perinatally infected subjects. 2 The goal of therapy for CHB is to prevent the progression of the disease. The two main strategies currently used are antiviral therapy with nucleoside/nucleotide analogues (NAs) or immunomodulators, such as pegylated interferon (PEG-IFN).3–5 A finite duration of treatment with PEG-IFN has been shown to have a better chance of achieving a sustained antiviral response. 6 However, side effects, the need for subcutaneous injections, and the contraindication profile, including individuals with cirrhosis, remain major limitations. NAs suppress HBV DNA replication by inhibiting the viral polymerase. 7 Tenofovir (TDF) was the latest NA approved by the FDA in 2008. It is structurally related to, and more potent than, the NA adefovir (ADV). Clinical trials indicate that TDF has superior antiviral efficacy with a similar safety profile as compared with ADV through 48 weeks.8,9 However, treatments with some NAs are associated with mutations and subsequent selection of resistant strains resulting in therapeutic failure. 10 A better understanding of the interaction between host and HBV will aid in developing more effective therapies and better biomarkers for disease monitoring of the large number of infected individuals.
The immune system has an inherent capacity to control HBV infection. 11 Thus, a synergistic approach comprising boosting the host immune response, specific immunotherapeutic interventions, and effective suppression of viral load can promote sustained viral clearance in chronic HBV infection. There is a large amount of data on the functionality of HBV-specific T cells, such as CD4 + T cells, CD8 + T cells, and regulatory T cells, that suggests their functionality is improved in NA-treated CHB patients12–16; however, this effect is often only transient and partial.12,16 HBV-specific B cells are also important effector cells during antiviral immune responses.17,18 A study using human PBLs in a NOD/SCID mouse model demonstrated that HBV core antigen binds and activates naïve human B cells in vivo. 19 In addition, sCD40L-activated B cells may function as antibody-producing cells and induce HBV-specific cytolytic T lymphocytes. 20 B lymphocytes are the most critical immune cells involved in humoral immune responses. They play important roles in defense against viral infections and are responsible primarily for the basic function of producing antibodies, whether antigen-specific or not. In addition, B cells are also important antigen-presenting cells. 21 In humans, aberrant circulating B-cell subpopulations are an increasingly recognized consequence of infectious 22 and inflammatory disease23–25; these alterations are reversed by anti-retroviral or immunoregulatory therapy.26,27 Moreover, protracted antigenic stimulation may result in B-cell activation and polyclonal proliferation, potentially leading to autoimmunity and lymphoproliferative disorders. 25 The differences observed between chronic HBV-infected patients and healthy controls has provided compelling evidence that B cells play an important role in the antiviral immune response,28,29 but altered B-cell homeostasis and B-cell surface receptors during antiviral therapy remain poorly understood. Investigation of the effects of NAs on B cells and the B-cell compartment would better define the significance of B-cell homeostasis in CHB pathophysiology and antiviral immunity.
To this aim, we performed a longitudinal study to examine the dynamic changes in the compartments and surface receptors of B cells in CHB-infected patients after the initiation of TDF or ADV therapy. We found that CHB patients displayed perturbation of B-cell homeostasis and higher levels of activation markers on B cells. This effect was partially improved after antiviral therapy with TDF or ADV.
Materials and methods
Patients
Characteristics of the study population
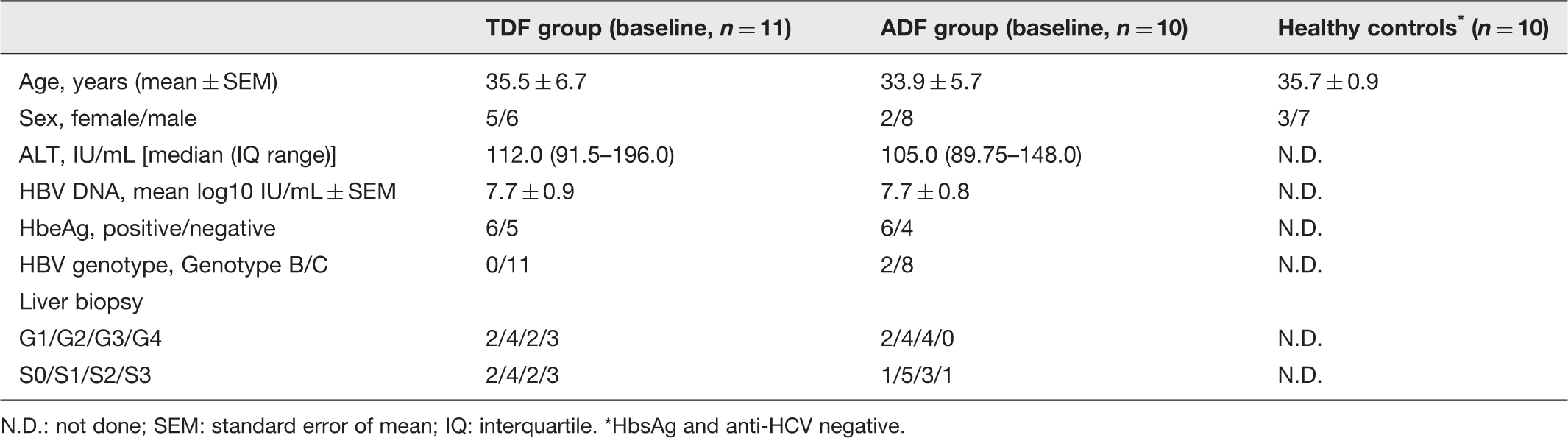
N.D.: not done; SEM: standard error of mean; IQ: interquartile. *HbsAg and anti-HCV negative.
Treatment
CHB patients were randomized into TDF and ADV treatment groups and treated with 300 mg once daily TDF (GSK Pharma, UK) or 10 mg once daily ADV (GSK Pharma, UK). All patients met the most recent European guideline criteria for treatment of CHB 30 and were followed up for 24 weeks. Six milliliters of EDTA-treated peripheral blood were obtained longitudinally before and after the treatment.
Virological and other biochemical measurements
HBV serology (HbsAg/Anti-HBs, HbeAg/Anti-Hbe, quantitative HbsAg, quantitative HbeAg) was determined by microparticle enzyme immunoassay. Serum HBV DNA load was measured by quantitative PCR using the Roche COBAS Taqman HBV test (Roche, UK); the limit of detection for HBV was 20 IU/mL. Serum levels of aspartate aminotransferase (AST) and alanine transaminase (ALT) were determined with a Biochemistry Automatic Analyzer (Roche Diagnostics, Branchburg, USA).
Flow cytometry
A six-parameter flow cytometric analysis of fresh whole blood cells was performed using a two-laser FACSCalibur instrument equipped with CellQuest Pro software (BD Biosciences, Erembodegem, Belgium). Absolute lymphocyte numbers and lymphocyte differentiation were measured by an accredited clinical lab. Fifty microliters of whole blood were stained with 1 µg/106 cells of different combinations of the following monoclonal antibodies: fluorescein isothiocyanate (FITC)-conjugated CD19 (clone HIB19) and allophycocyanin (APC)-conjugated CD27 (clone MT271), or CD5 (clone UCHT2) and Peridinin-Chlorophyll-Protein Complex (PerCP)-conjugated CD3 (clone SK7), and stained with Phyco-Erythrin (PE)-conjugated antibodies against CD69 (clone L78), CD80 (clone L307.4), CD86 (clone FUN-1), CD24 (clone ML5), CD22 (clone S-HCL-1), or with isotype-matched monoclonal antibodies of irrelevant specificity FITC-, PE-, APC-, and Tricolor-conjugated as negative controls.
After a 30-min incubation at 4℃, whole blood was lysed with lysing solution (BD Pharm Lyse), and the cells were washed twice with PBS 2% fetal calf serum, resuspended in FACS buffer, and immediately analyzed.
Statistical analysis
Descriptive statistics were used to summarize patient characteristics. The difference between the TDF and ADV groups was analyzed by the Mann-Whitney U test and the difference between paired variables was determined by the Wilcoxon matched pairs test using the Prism 5.0 (GraphPad software, USA). Spearman’s rank correlation coefficient was used for correlation analysis. All tests were two-sided and P-value of less than 0.05 was considered statistically significant.
Results
Disrupted B-cell homeostasis in CHB-infected patients is associated with liver damage
To determine B-cell homeostasis, 21 CHB patients and 10 healthy subjects were recruited into this study. CHB patients displayed elevated ALT and HBV loads. Liver biopsy showed all patients without severe fibrosis (Table 1).
Characterization of peripheral blood B cells by flow cytometry analysis indicated that both the total and percent of CD19 + CD3 − B cells in CHB patients were significantly higher than in HD (P < 0.001; Figure 1(a1) and (a2)). Furthermore, the total and percent of CD5 + B cells were also significantly higher than HD (P < 0.001; Figure 1(b1) and (b2)). The distribution of CD27 + /CD27 − B-cell subsets was similar between CHB-infected patients and HD (data not shown).
The percent and absolute number of total (CD19 + CD3−) B cells, CD5 + B cells, and the expression of activating receptors on total B cells at baseline. The percent and absolute number of total (n = 21) and CD5 + (n = 17) B lymphocytes from CHB patients and 10 HD were analyzed ex vivo by flow cytometry. Data are shown as the percentage (a1, b1) and absolute numbers (a2, b2) of gated lymphocytes. Activation markers CD69 (c1) and CD24 (c2) on total B cells were determined by mean fluorescence intensity (MFI). Data are expressed as mean ± SEM. ***P < 0.001, **P < 0.01, *P < 0.05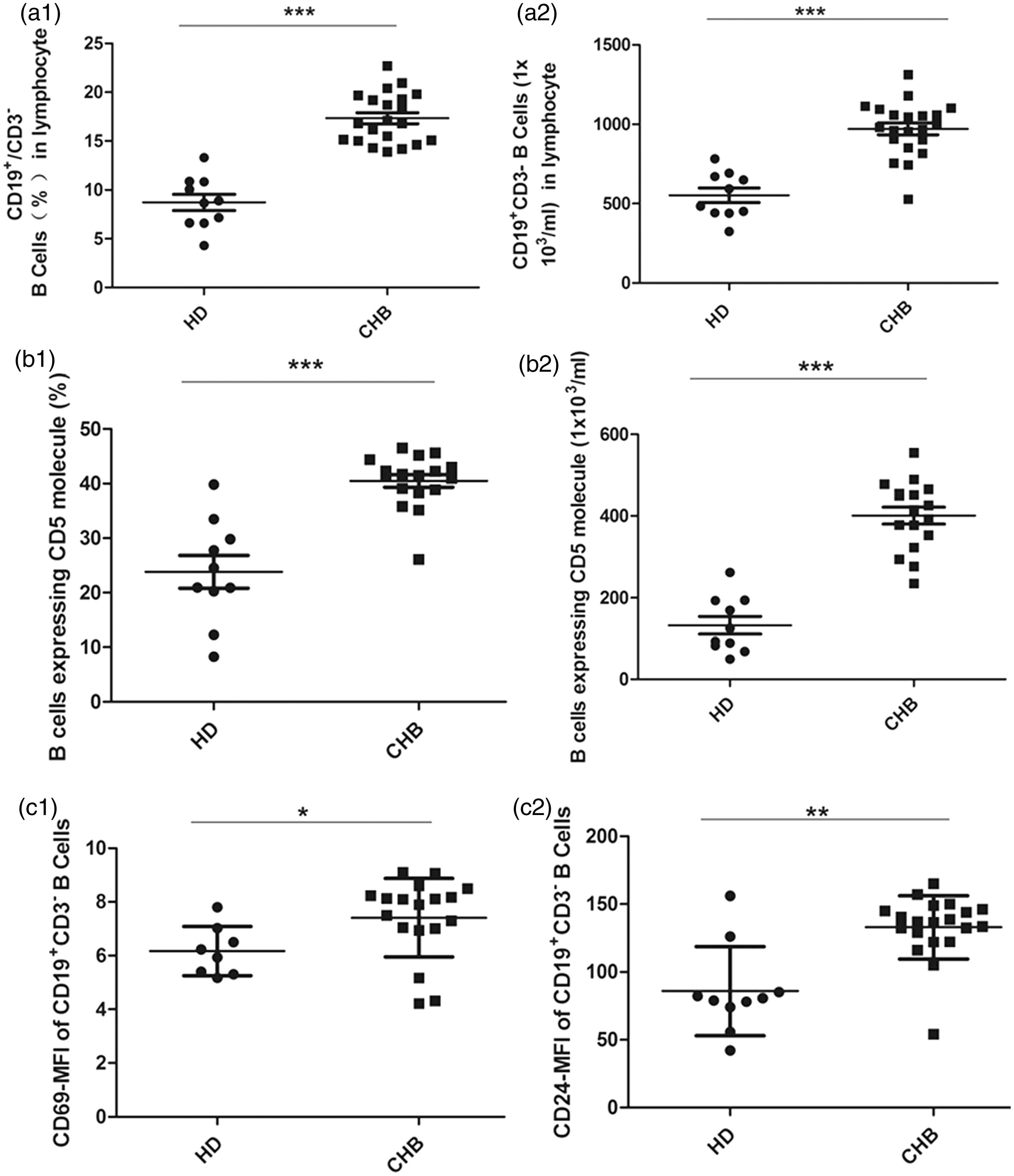
Cell surface marker on B cells observed in the study

CHB patients demonstrate different therapeutic responses to TDF and ADV treatment
Comparison of liver function in CHB patients after treatment with ADV or TDF
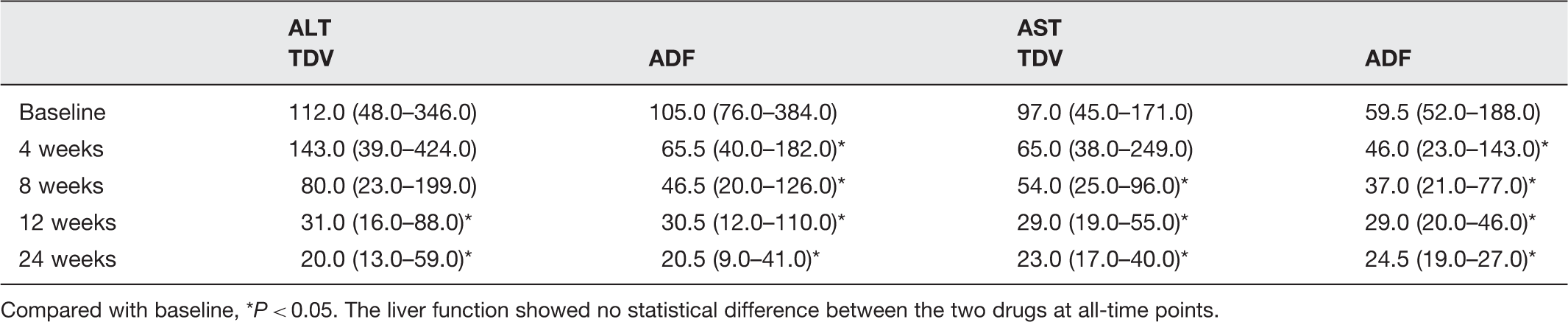
Compared with baseline, *P < 0.05. The liver function showed no statistical difference between the two drugs at all-time points.
The HBV DNA level before and after antiviral treatment
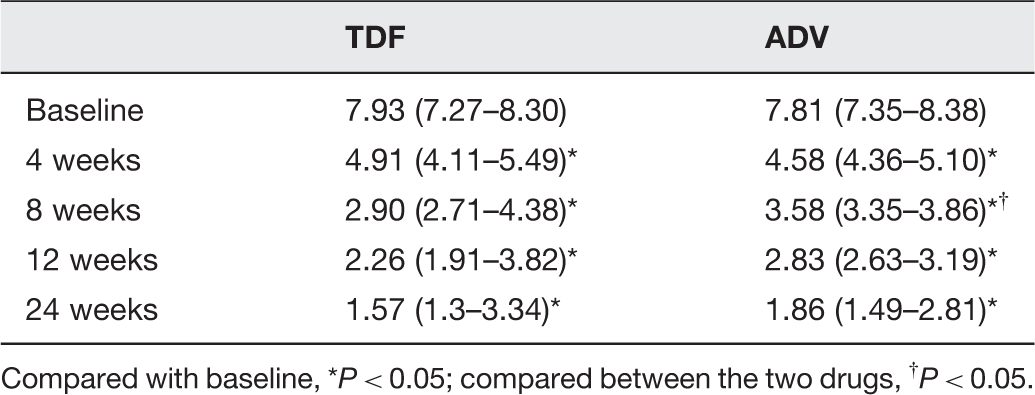
Compared with baseline, *P < 0.05; compared between the two drugs, †P < 0.05.
Treatment with TDF or ADV improves altered peripheral B lymphocyte homeostasis and liver inflammation in CHB-infected patients
We next asked whether altered peripheral blood B lymphocyte homeostasis could be restored following different treatments. We found that the total and percent of CD19 + /CD3 − B cells transiently decreased at week 8,12, and 24 post initial treatment in both the TDF (Figure 2(a1) and (a2)) and ADV groups (Figure 2(b1) and (b2)). These data clearly indicate that treatment with TDF or ADV modulated the homeostasis of B cells, but TDF demonstrated a stronger effect than ADV. This effect is shown in a representative flow cytometry scatter plot in Figure 2(c).
Decreased CD19 + CD3 − B cells after antiviral treatment. CD19 + CD3 − B-cell relative and absolute number in CHB patients who received TDF (a) (n = 11) or ADV (b) (n = 10) treatment were analyzed at the indicated time points. Data are expressed as mean ± SEM for (a) and (b) and percent for (c). (c) Representative FACS plots of CD19 + CD3 − B cells (gating on lymphocytes) from TDF group. ***P < 0.001. (A color version of this figure is available in the online journal)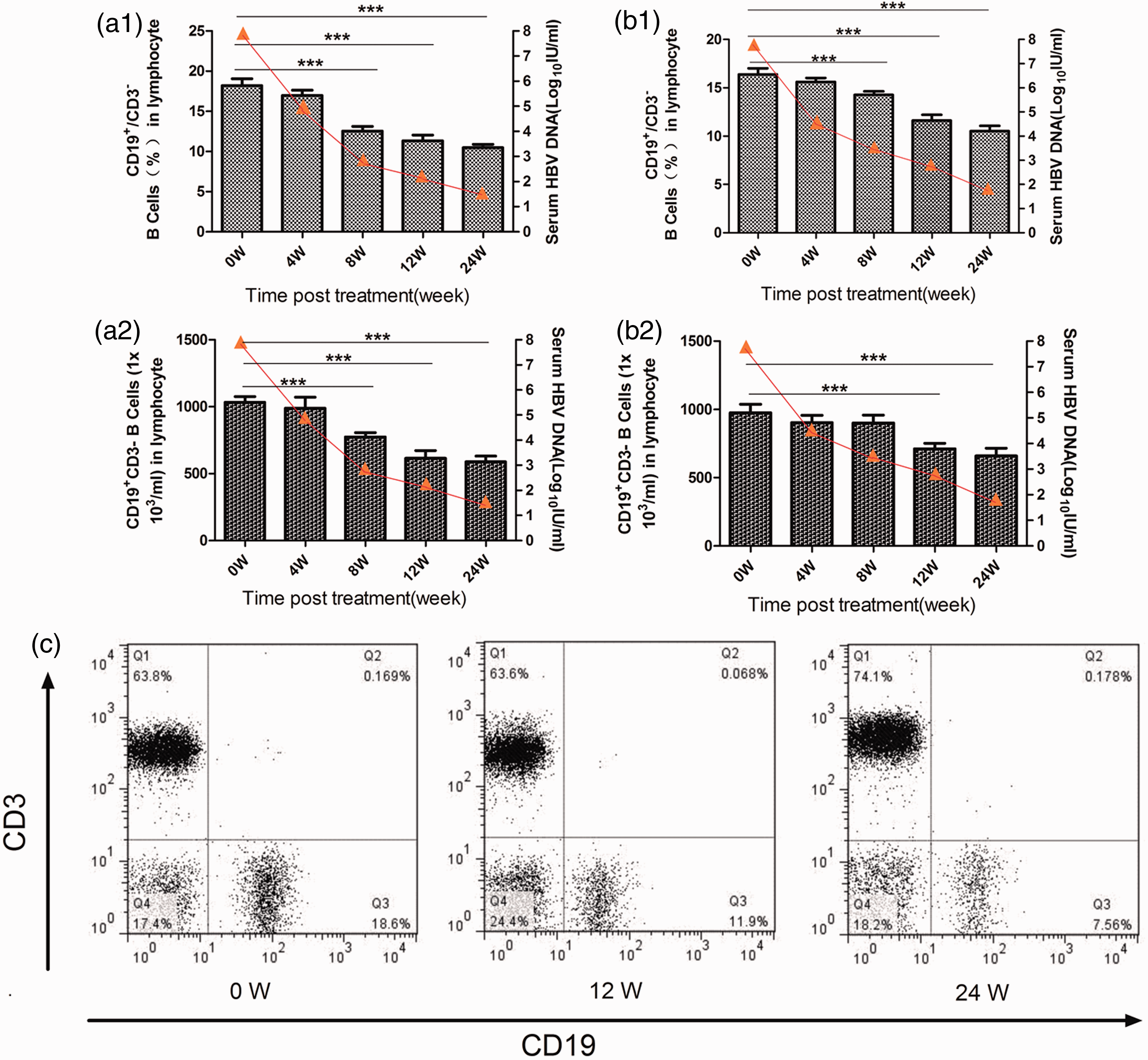
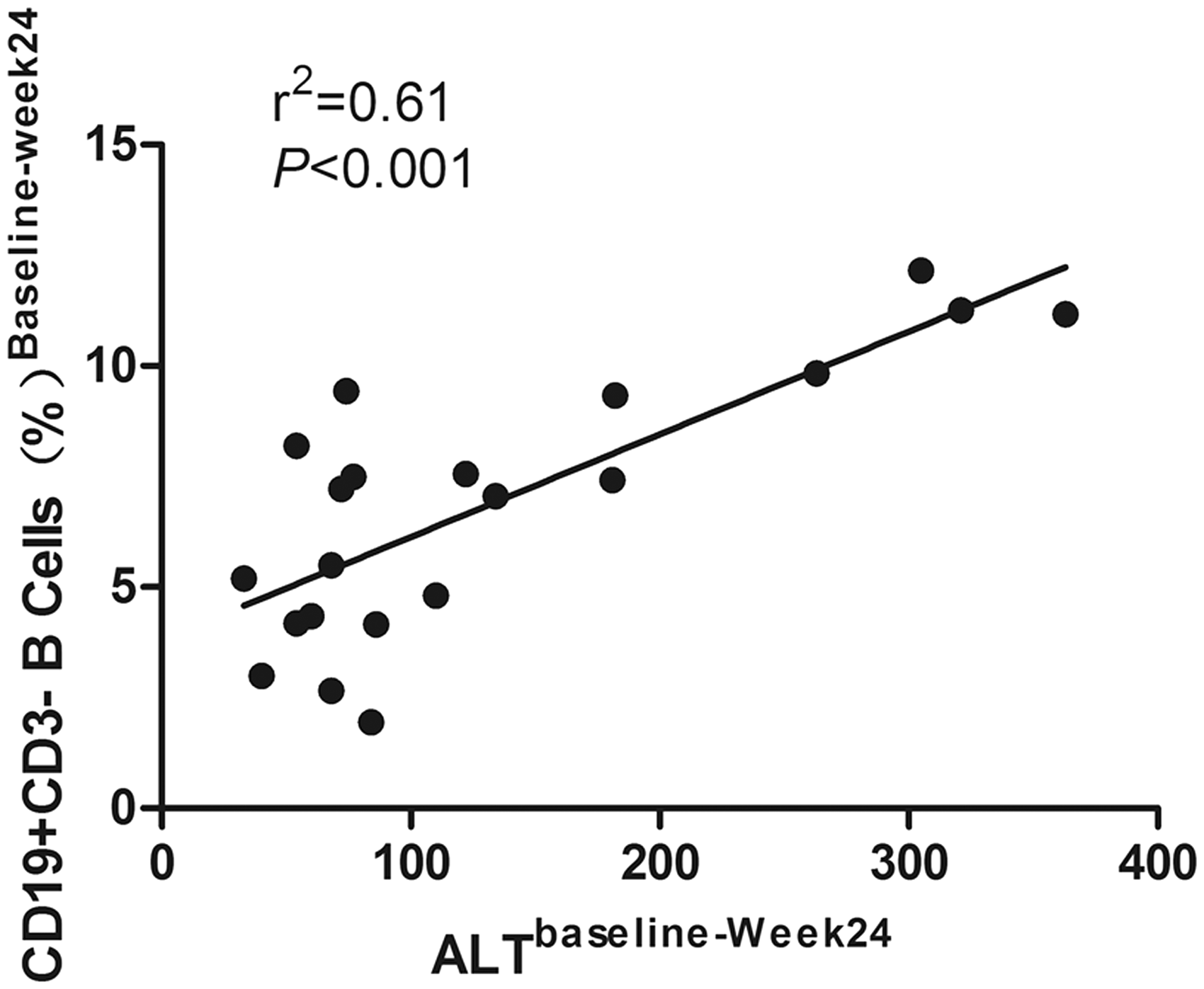
We also examined the CD5 + B-cell subpopulation before and after antiviral treatment. The results revealed that the decrease in both total and percent of CD5 + B cells was similar to that seen for CD19 + CD3 − B cells upon TDF (Figure 3(a1) and (a2)) and ADV (Figure 3(b1) and (b2)) therapy; the decline in absolute number is more obvious than in the percent. The number of CD5 + B cells was significantly different between the two groups at week 8 after antiviral treatment, when the TDF group had a CD5 + B-cell number of 202.2 ± 64.3 × 103 cells/mL (average ± standard deviation) compared with 284.3 ± 74.3 × 103 cells/mL (P = 0.032) in the ADV group. A representative flow cytometry scatter plot is shown in Figure 3(c). Interestingly, a strong correlation was demonstrated between ALT and the percent of CD19 + B cells after antiviral therapy, as shown in Figure 4; data were generated in both groups of CHB-infected patients (r2 = 0.61, P < 0.001).
Decreased number of CD5 + B cells after antiviral treatment. CD5 + B-cell subset relative and absolute number in patients with CHB who received TDF (a) (n = 11) or ADV (b) (n = 10) treatment at the indicated time points. Peripheral blood mononuclear cells were prepared from individuals and stained with a FITC-CD19 and percp-CD3 and APC-CD5 antibodies or isotope controls. The cells were gated on lymphocytes and gated on CD3 − CD19 + B cells for dynamic change in the relative and absolute number of CD5 + B cells. ##P < 0.05 for the TDF group compared ADV group. Data are expressed as mean ± SEM for (a) and (b) and percent for (c). (c) Representative FACS plots of CD5 expression from the TDF group. *P < 0.05, **P < 0.01, ***P < 0.001. (A color version of this figure is available in the online journal) The possible correlation between CD19 + CD3 − B-cell percent and serum levels of ALT in 21 CHB patients at 24 weeks. Spearman’s rank correlation coefficient between the therapy-induced decrease in ALT and a decrease in CD19 + CD3 − B cells was determined at baseline and 24 weeks. r2 indicates Pearson’s correlation coefficient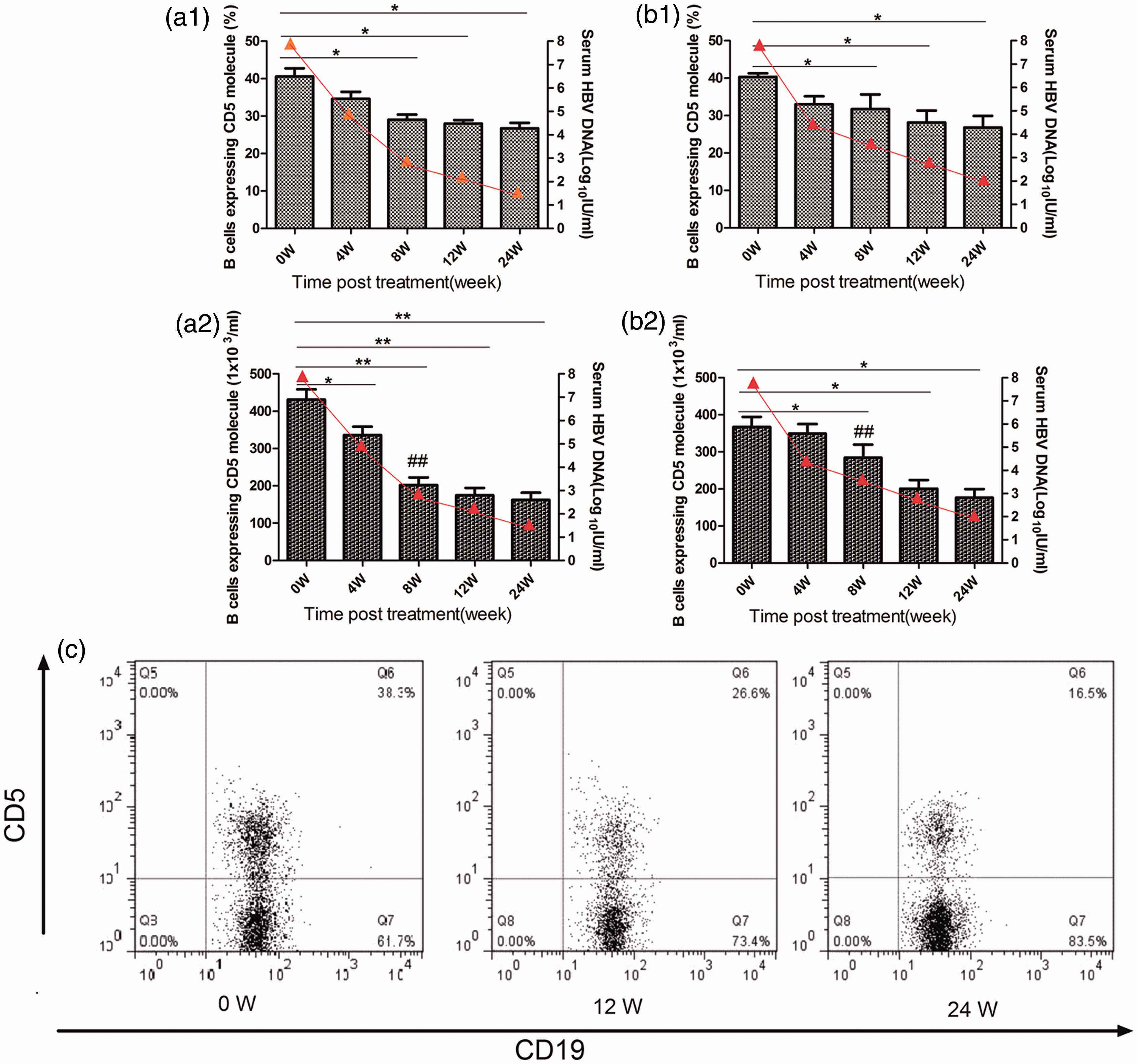
Furthermore, we determined whether CD27 + /CD27 − B-cell subsets were effector cells in antiviral immunity induced by TDF and ADV. Interestingly, we observed that the percentage of CD27 + memory B cells was increased and the absolute number of these cells was reduced in the TDF group (Figure 5(a) and (b)). We next analyzed the change in total and percent of CD27 − naïve B cells in the TDF group. As expected, the total and percent of CD27 − naïve B cells was significantly lower at week 24 compared to baseline (P = 0.003 and P = 0.026, respectively, Figure 5(c and d)). Thus, we conclude that the increased percentage of CD27 + B cells upon treatment with TDF is a result of the reduction in the CD27 − naïve B-cell pool and a reduced number of CD27 + memory B cells. However, no significant differences were noted in the CD27 + /CD27 − B-cell subpopulation before or after initial treatment in the ADV group (data not shown).
The change in CD27 + /CD27 − B cells during antiviral treatment with TDF. The relative and absolute number of CD27 + /CD27 − B cells at baseline (week 0), week12 and week 24 after TDF treatment (n = 11). Data are shown in box-and-whisker plots: (a) percent CD27 + B cells; (b) absolute number of CD27 + B cells; (c) percent CD27 − B cells; and (d) absolute number of CD27 − B cells. ***P < 0.001, **P < 0.01, * P < 0.05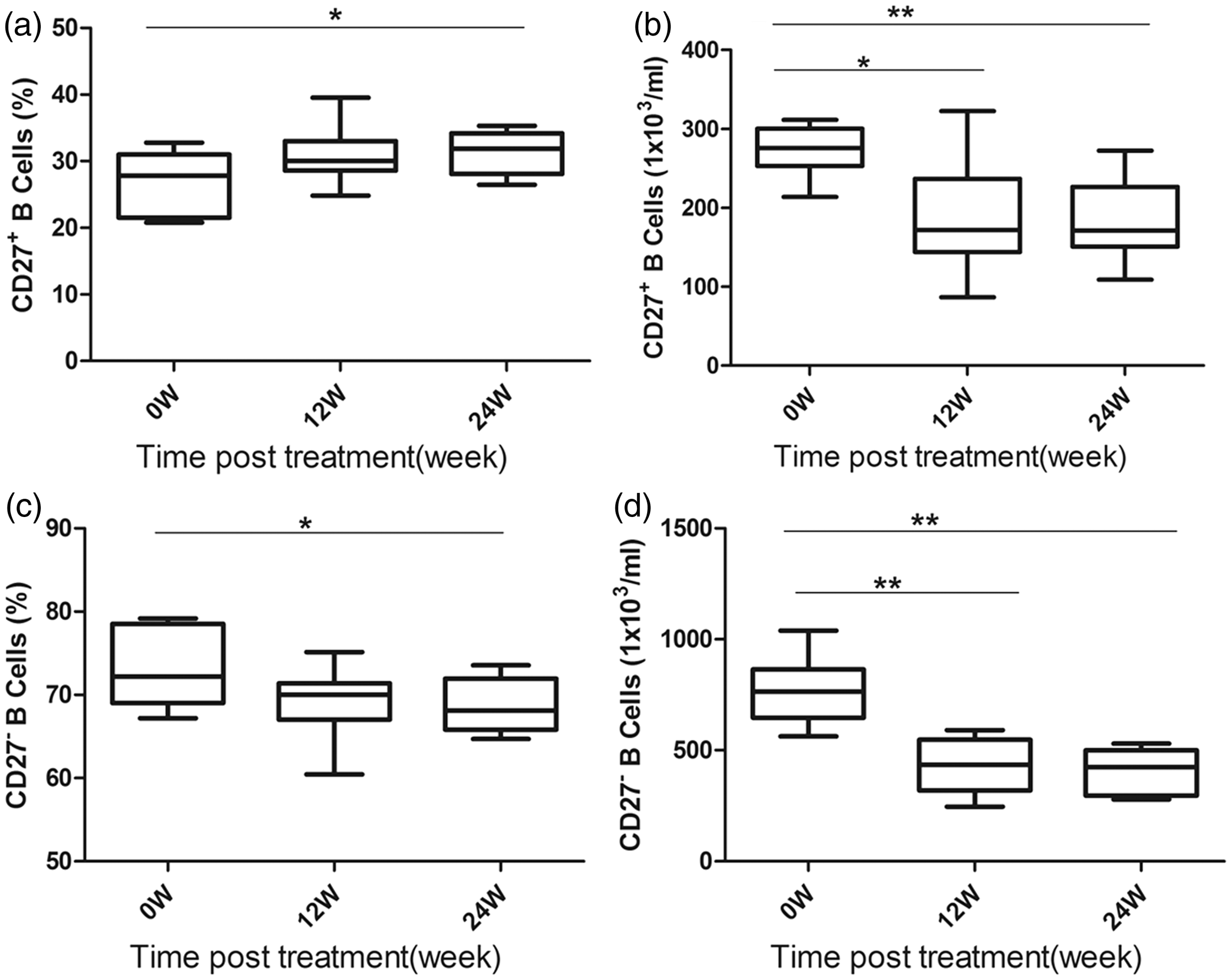
Discussion
To better understand the source of disrupted B-cell homeostasis associated with HBV infection and test the hypothesis that homeostasis will recover following a decline in HBV DNA load induced by antiviral therapy, we characterized circulating B cells in patients chronically infected with HBV. In this study, we examined the percent and absolute number of total and differentiated subsets of circulating B cells in patients with CHB following NA antiviral treatment. We found significant differences in both the total and percent of B cells between CHB patients and HD. The percent of CD19 + CD3 − B cells in CHB patients was significantly higher than those in HD, consistent with recent data on chronic HCV infection. 31 B cells play a decisive role in fibrosis and later stages of liver disease, 32 though we did not find a significant difference in liver fibrosis stages.
Cooper 33 and Good, 34 in the late 1960s, established that B cells comprise a specific subset of lymphocytes, and that they possess unique characteristics and functions consistent with their role as initiators and effectors of the humoral immune system. Of particular interest are recent findings showing disrupted B-cell homeostasis in some autoimmune diseases. We observed the CD5 + /CD5 − and CD27 + /CD27 − B-cell subsets to characterize homeostasis in CHB patients compared to HDs. In the present study, we found that peripheral blood CD5 + B cells significantly increased in patients with CHB infection when compared to HD. By contrast, memory/naïve (CD27 + /CD27 − ) B cells were similar in CHB patients and HD. CD5 is expressed on a subset of B lymphocytes associated with the production of low-affinity “natural antibodies” or autoantibodies. These cells are the source of much of the IgM in the circulation and may act more autonomously than the CD5 − B-cell subset.35,36 There have been studies showing that “natural antibodies” bind to virus-antibody-complement immune complexes and aid in their clearance by the spleen. 37 Furthermore, CD5 + B cells are long lived, which may be due to their autocrine expression of IL-10 following BCR activation. 38 CD5 + B cells have the ability to regulate the immune response. Therefore, our data suggest that clonal proliferation of CD5 + B cells may affect peripheral B-cell homeostasis and the hepatic immune response in CHB patients. The disrupted B-cell homeostasis is involved in liver damage, as the liver is the first line of defense in antiviral humoral immunity. It is well known that a sustained B lymphocyte-mediated immune response against HBV, that is, a memory B lymphocyte response, provides protection against further HBV infection. 39 In addition, long-term protection after hepatitis B vaccination is dependent on the persistence of a strong immunologic memory.40,41 However, we did not find an alteration of CD27 + /CD27 − B cells in CHB patients compared to HD, which would be indicative of a defect in immune capacity to resist chronic HBV infection or produce and secrete antigen-specific antibodies. Further antiviral therapy would be necessary to modulate humoral immune responses for inhibiting viral replication or secreting antibodies.
It has been suggested that chronic antigenic stimulation of B cells by persistent virus is a cause of B-cell activation and transformation. 42 We examined circulating B cells for expression of activation markers in CHB patients compared to HD. We found that CD69 and CD24 expression on B cells was higher in CHB patients, in keeping with data of Rosa and Oliviero,28,43 and CD80 levels mildly decreased. However, in contrast with the previously cited study, others have failed to detect activated B cells in chronic HCV infection. 31 The activation of B cells did not appear to be affected by TDF or ADV treatment in our study. Weakly activated B cells may be accounted for by evidence that HBV replication is not sufficient to induce B-cell activation or accumulation.
Variable recovery of B-cell homeostasis has previously been found in patients with immune dysfunction following effective immunomodulatory therapy, suggesting that the recovery of B-cell homeostasis in this setting contributes to clinical improvement in these patients.26,27,44 We treated CHB patients with the NAs TDF or ADV and characterized the change in total and differentiated subsets of B cells by flow cytometry. We found that treatment with either regimen resulted in a transient decline in the total and percent of peripheral total and CD5 + B cells at weeks 8, 12, and 24 post-treatment. The implications are that the B-cell pool size and repertoire can be manipulated by diminishing HBV DNA load following antiviral treatment with TDF or ADV. Interestingly, decreased ALT levels and an altered CD19 + B-cell profile induced by treatment showed a strong positive correlation. These data suggest that treatment with TDF or ADV induce total B-cell depletion, in contrast to the accumulation of B cells after rituximab treatment for chronic graft-versus-host disease and systemic lupus erythematosus,27,44 where low levels of B cells exist in the active phase before treatment. Moreover, the reduced percent CD19 + B cells and the restoration of ALT levels would appear to alleviate liver damage and the inflammatory response. In addition, the decrease in CD5 + B cells following treatment with TDF or ADV indicates that disrupted B-cell homeostasis induced by CD5 was gradually restored. Interestingly, at week 8 after treatment, the decrease in CD5 + B-cell numbers is more significant in the TDF group than in the ADV group, which may contribute to the difference in HBV DNA load between the two groups at week 8. Taken together, our data suggest an important role of B-cell homeostasis in recovery of both liver damage and the antiviral effect.
When the overall percent and total number of peripheral B cells expressing CD27 were examined post initiation of treatment, a general reduction of B cells was observed following reduced viral replication and HBV Ag release, particularly CD19 + /CD27 − naive B cells during antiviral treatment with TDF. Remarkably, the disrupted naïve/memory B-cell homeostasis was not observed in the ADV group. Thus, the naïve/memory B-cell subpopulation reconstitution may be associated with the stronger antiviral response after TDF in CHB.
In summary, the current study provides clear evidence that there are persistent disruptance in the peripheral B-cell compartments of CHB patients, and that partial recovery is likely associated with decreased HBV DNA load. After treatment with one of two different NAs, TDF or ADV, the accumulation of peripheral B cells underwent either incomplete or transient B-cell depletion; activation markers on B cells did not change significantly during antiviral therapy and no toxicity was observed. Our results, together with recent evidence, suggest ongoing, antigen-driven immune responses may lead to profound B-cell depletion, which may accelerate B-cell differentiation into effector cells. Reducing the HBV DNA load by NA antiviral treatment, rather than through innate immune cells such as NK cells, causes changes in proliferation and activity to help regulate the immune response. Despite the small number of patients in this study, our findings provide data that will inform future translational studies. We provide a unique insight into the disrupted B-cell in CHB patients during TDF- and ADV-induced reduction of viral load. Based on our data, further studies with a larger population, along with liver tissue analysis and studies of B-cell function, can be performed to validate these findings.
Footnotes
Author contributions
HS and JL contributed equally to this work. All authors participated in the design, interpretation of the studies, and review of the manuscript. HS, JL, XH, HY, and XC performed the experiments. JN, ZT, and YP analyzed the data. HS, DX, and JN wrote the paper.
Acknowledgements
This study was supported by grants from the Natural Science Foundation of China (81172727; 30972611) and Natural Science and Technology Major Project (2012ZX1 0002003), which are the funding sources for this study. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the paper.