
Abstract
IgA nephropathy (IgAN) is an immune complex glomerulonephritis that is characterized by recurrent hematuria as the main clinical manifestation. In this study, we used the IgAN mouse model which was previously established to investigate the possible mechanism by which IgAN fibrosis correlates with decreased plasma gelsolin (pGSN) levels. We investigated the levels of pGSN, transforming growth factor β1 (TGFβ1), and oxidative stress markers including superoxide dismutase (SOD), catalase (CAT), glutathione (GSH), and malondialdehyde (MDA) in the serum and renal tissues of different groups. The between-group differences and correlations in the results for the IgAN group were analyzed with statistical methods. The pathological and urinalysis results obtained from the IgAN mouse model showed that this model conforms to the basic lesion characteristics observed in human IgAN. The serum pGSN levels and SOD, CAT, GSH levels in renal tissues were decreased in the IgAN group (P < 0.01), and pGSN, TGFβ1, MDA levels in renal tissues of the IgAN group were increased which compared with those in the other groups (P < 0.01). The correlation analysis for serum pGSN levels in the IgAN group showed a significant correlation with different test results (P < 0.01). The possible mechanism by which IgAN fibrosis correlates with decreased pGSN levels involves the regulation of TGFβ1 and oxidative stress.
Introduction
IgA nephropathy (IgAN) is the most common primary glomerulonephritis worldwide and accounts for 20–50% of primary glomerulonephritis cases. Approximately 20–40% of patients develop end-stage renal failure within 10–15 years of disease onset. 1 The majority of IgAN patients exhibit recurrent hematuria as the main clinical manifestation, however, it is difficult to diagnose this immune complex glomerulonephritis early due to the lack of effective clinical detection markers.
Gelsolin, which was first identified in rabbit pulmonary macrophages by Yin and Stossel in 1979, 2 was shown to regulate actin reorganization in a calcium-dependent manner. Gelsolin was also found in platelet cells 3 and serum. 4 It was subsequently shown that gelsolin is widely distributed from lower eukaryotes to higher mammals and is an important component of cytoskeletal proteins. Gelsolin plays an important role in cancer, apoptosis, infection and inflammation, aging, and different diseases. 5 Plasma gelsolin (pGSN) is the extracellular isoform of gelsolin that circulates in the plasma of healthy humans and other mammals at average levels of 200 ± 50 mg/L. 6 It has been suggested that pGSN acts as an actin-scavenging protein in plasma. It is putatively designed to remove actin released from damaged cells in order to prevent increases in blood viscosity and possible toxicity of insoluble F-actin filaments. 7 Many researchers have reported that pGSN levels are subnormal in different diseases, including acute liver injury, idiopathic lung injury, allogeneic stem cell transplantation, trauma, burns, and bacterial and protozoal sepsis.8–13 Osborn et al. found that pGSN is decreased in the serum of patients with rheumatoid arthritis compared with healthy patients 6 and concluded that the decreased levels of circulating pGSN may be linked to autoimmune disease. These preliminary findings prompted us to hypothesize that decreased serum pGSN levels may be involved in the development and progression of IgAN, which is an immune complex glomerulonephritis and has similar autoimmunity pathogenesis with rheumatoid arthritis.
It is widely recognized that IgAN development and glomerular fibrosis are related closely. A variety of cytokines are involved in glomerular fibrosis, with the TGFβ superfamily being considered one of the most important. Some studies have indicated a central role for TGFβ and its downstream signaling cascades in the activation of the cellular pathomechanisms that underlie the progression of glomerular fibrosis. 14 Sharma et al. found that TGFβ1 can provoke and accelerate the effects of oxidative stress in isolated rat glomeruli. 15 Oxidative stress can be further activated by inflammatory cytokine networks, causing glomerular injury and the aggravation of fibrosis in IgAN. Oikonomou et al. found that pGSN exhibits an anti-inflammatory effect in a pulmonary inflammation and fibrosis model. 16 Chauhan et al. reported that pGSN counteracts oxidative stress in Alzheimer’s disease. 17 However, no study has investigated the interactions between pGSN, TGFβ1, and oxidative stress in IgAN. We used the IgAN mouse model previously reported by our group 18 to clarify the role of decreased pGSN levels in IgAN fibrosis. Interestingly, our study indicated that pGSN levels are decreased in the serum of IgAN mice, which, together with the interaction between pGSN, TGFβ1, and oxidative stress, makes pGSN an attractive candidate for further studies aimed at better understanding IgAN progression.
Materials and methods
Establishment of murine IgA nephropathy model
The IgAN animal model used in this study was previously established. 18 Briefly, 120 four-week-old Balb/c mice (body weight 20–22 g) were used, which could ensure that sufficient quantities were available for statistical analysis. All animals were divided into four groups. The 30 mice in the control group received no treatment. The mice in the remaining groups were injected subcutaneously every 2 weeks. Of these animals, the 30 mice in the IgAN group received 3 mg/kg of the MBP-20-peptide fusion comprising the maltose-binding protein (MBP) and a 20-amino-acid peptide which was found to be the antigenic determinant derived from Staphylococcus aureus responsible for the immune manifestations of IgAN. 19 The 30 mice in the elution buffer group received elution buffer, which have the composition of Maltose, Tris–Hcl, NaCl, EDTA, and double-distilled water. Because the MBP-20-peptide fusion in the IgAN group was diluted by elution buffer, the aim of this group was to exclude the possibility that our animal model was induced by elution buffer. The 30 mice in the irrelevant 20 peptide group received 3 mg/kg of the MBP-irrelevant-20-peptide fusion protein, which has a different amino acid sequence (YVEFGSSRVDLQSFPRKA SE) with the 20-peptide in the IgAN group (NVGGDNVDIHSIVPVGQD PH). This group was used to confirm that the 20-peptide in the IgAN group was the antigenic determinant responsible for the immune manifestations of the IgAN group mice. The MBP-20-peptide/irrelevant-20-peptide fusion protein were synthesized separately according to our previous report. 18 The urine of the mice was collected weekly and analyzed for the hematuria and proteinuria levels. The mice in each group were sacrificed by transection of spinal cord at the end of 21 weeks, and fresh kidney tissues and serum were preserved for pathological, biochemical, immunofluorescence, western blotting, real-time polymerase chain reaction (PCR), oxidative stress (spectrophotometry), and enzyme-linked immunosorbent assay (ELISA) tests. The use of animals has been approved by the Animal Care and Use Committee of Harbin Medical University.
Indirect immunofluorescence
Frozen slices from the renal tissue of mice were fixed in acetone for 5 min. Non-specific protein binding sites were blocked with 5% normal goat serum in Phosphate Buffered Saline (PBS) (pH 7.4). A mouse monoclonal anti-pGSN (1:50 dilution) antibody (Abbiotec, CA, USA) and a mouse monoclonal anti-TGFβ1 (1:100 dilution) antibody (Abcam, Hong Kong) were incubated with the sections (4 µm) overnight at 4℃. After rinsing, the sections were incubated with fluorescein isothiocyanate-labeled antimouse IgG (Vector Labs, Burlingame, CA, USA). After rinsing, the sections were observed and images were acquired with a fluorescence microscope (Nikon E800, Japan) equipped with a digital camera (1200F; Nikon) and image acquisition software (ACT-1; Nikon). To evaluate the level of glomerular staining with the pGSN and TGFβ1 antibodies, the gray scale values of the glomeruli were measured by software (ACT-1; Nikon) in all of the groups.
Western blotting
Equal amounts (50 ug) of nephridial tissue extracts were resolved by sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred to a nitrocellulose membrane before being incubated with a primary mouse monoclonal antibody against pGSN (1:500 dilution; Abbiotec, CA, USA) and a mouse monoclonal antibody against TGFβ1 (1:800 dilution; Abcam, Hong Kong). A horseradish peroxidase-conjugated secondary antibody (1:2000; Santa Cruz, CA, USA) was used for detection. The proteins were visualized by enhanced chemiluminescence detection reagents (Pierce, Rockford, IL), and the protein bands were quantified with Quantity One Software (Bio-Rad, Hercules, CA).
Quantitative real-time PCR
The whole nephridial tissue total RNA was extracted with TRIzol reagent (Invitrogen), and complementary DNA was synthesized by using a Reverse Transcription Kit (Toyobo, Osaka, Japan) according to the manufacturer’s protocol. The sequences of the mouse pGSN sense and antisense primers were 5′-AGGCGACAGCTA-CATCATTC-3′ and 5′-TCAGAGCACCAGACTTAGGC-3′, respectively. The sequences of the TGFβ1 sense and antisense primers were 5′-TCGACATGGAGCTG-GTGAAA-3′ and 5′-GAGCCTTAGTTTGGACAGGATCTG-3′, respectively. The sequences of the sense and antisense primers for β-actin were 5′-GCCA-ACACAGTGCTGTCTGG-3′ and 5′-GCTCAGGAGGAGCAATGA- TCTTG-3′, respectively. The PCR was performed as recommended by the manufacturer using SYBR Green PCR Master Mix (Toyobo, Osaka, Japan). The level of each specific transcript was normalized to the level of β-actin and subsequently calculated using the comparative threshold cycle (Ct) method (2_ΔΔCt). 20 Each sample was run in duplicate.
ELISA
Mice blood samples were obtained by enucleation of eyeball from live anesthetized animals, and sera were collected after centrifugation. The concentrations of pGSN and TGFβ1 in the mice serum were detected by separate ELISA assays. Each well of a polystyrene microtiter plate (Corning, NY, USA) was coated with mouse anti-pGSN (1:1000 dilution) antibody (Abbiotec, CA, USA) in carbonate buffer (0.05 M, pH 9.6) overnight at 4℃. After washing with PBS containing 0.05% Tween 20 (PBS-T), the plates were then incubated with pGSN protein (Cytoskeleton, Inc.) and mouse serum samples from the IgAN group and the other control groups (1:50 dilution) at 37℃ for 1.5 h. After the plates were washed with PBS-T, peroxidase-conjugated goat antimouse IgG (Vector Labs, Burlingame, CA, USA) was added (1:2000 dilution) to the plates, which were then incubated at 37℃ for 1 h. The plates were washed with PBS-T, TMB peroxidase substrate was added, and the reaction was stopped with 2 M H2SO4. The absorbance was measured with a BIO-RAD 550 microplate reader (Bio-Rad, Tokyo, Japan) at a wavelength of 450 nm. The concentration of TGFβ1 was determined by following the instructions of the TGFβ1 ELISA kit (Santa Cruz, CA, USA).
Spectrophotometric oxidative stress test
The superoxide dismutase (SOD) and catalase (CAT) activities as well as the GSH and malondialdehyde (MDA) content in the samples were determined by spectrophotometry to evaluate oxidative stress. Briefly, homogenates were prepared from mouse kidney tissues. The SOD, CAT, GSH, and MDA activities and protein levels were determined by following the instructions provided with the spectrophotometry kits (Jiancheng, Nanjing, China). The optical density values (OD value) were recorded and translated into activity or content with the corresponding formulas.
Statistical analysis
The results were analyzed with SAS version 9.1 software (SAS Institute Inc., Cary, NC, USA). The between-group differences were calculated with a one-way ANOVA, and the results are presented as means ± standard deviation (SD). Pearson’s correlation was used to evaluate the correlations between the results. All P values quoted are two-tailed, and the significance is defined as P < 0.05.
Results
Pathological characteristics of the murine IgA nephropathy model
Figure 1 shows the pathological characteristics of the mouse IgAN model, which were determined with the immunofluorescent detection of IgA deposits, light microscopy, and electron microscopy. Immunofluorescence testing with an anti-IgA antibody showed IgA deposits in the glomeruli of the IgAN group (arrow), while in the other three groups, no significant IgA deposition was found. Hematoxylin and Eosin (H&E) staining showed a mild to moderate increase in the mesangial matrix and proliferation of the mesangial cells in the IgAN group (arrow), while other three groups show no obvious pathological changes in renal tissues. The electron microscopic analysis showed numerous large, electron-dense deposits in the mesangium of the IgAN group (arrow), but no electron-dense deposits were found in the other groups.
Pathological detection of the mouse IgA nephropathy model. Immunofluorescence showed that IgA deposits in the glomeruli of the IgAN group (arrow, IF × 200), while no significant IgA deposition was observed in the other three groups (control, elution buffer, and irrelevant 20 peptide group, IF × 200). Hematoxylin and eosin staining showed a mild to moderate mesangial cells proliferation in the IgAN group (arrow, HE × 400), and no significant pathological changes in renal tissues of the other three groups were found (HE × 400). The electron microscopic detecting showed numerous large, electron-dense deposits in the mesangium of the IgAN group (arrow, EM × 6000), but no electron-dense deposits were found in the other groups (EM × 6000). (A color version of this figure is available in the online journal)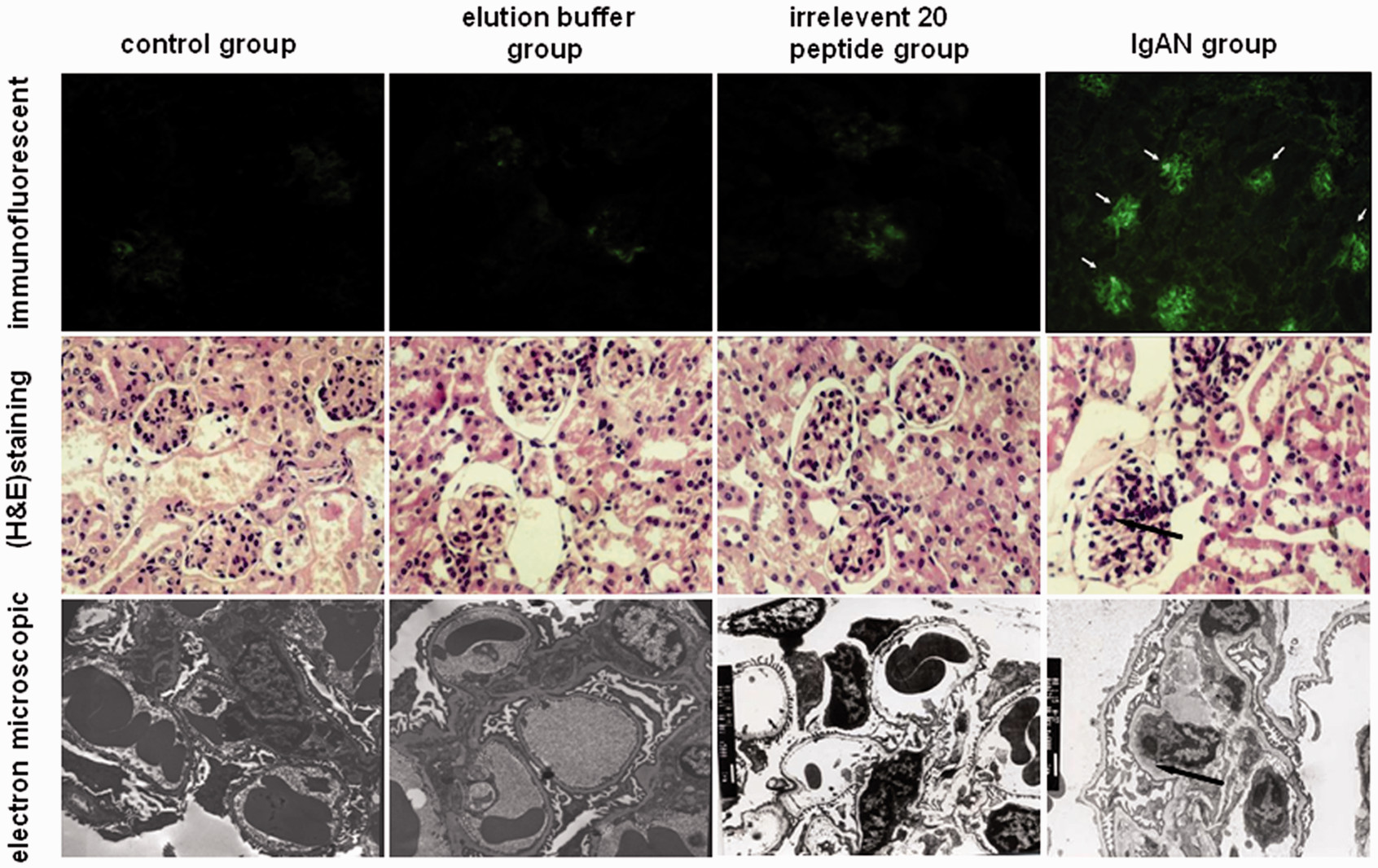
Immunofluorescence detection
As shown in Figure 2(a), the intensity of the pGSN and TGFβ1 staining in the glomeruli of the IgAN group was significantly higher than that in the other groups. A quantified analysis of the immunofluorescence intensity in the glomeruli shows that the pGSN and TGFβ1 gray scale values in the IgAN group were significantly higher than those in the other groups (P < 0.01) (Figure 2(b) and (c)).
The immunofluorescence detection of pGSN and TGFβ1 in IgAN group and other groups. (a) pGSN and TGFβ1 intensity in glomeruli of IgAN group was significantly higher than that in other groups. Quantified analysis of the immunofluorescence intensity in glomeruli shows that the (b) pGSN and (c) TGFβ1 gray scale values in the IgAN group (n = 30) were significantly higher than those in the other groups (n = 90, each group 30), comparisons between groups were tested by one-way ANOVA analysis and least significance difference (LSD) test (*P < 0.01). (A color version of this figure is available in the online journal).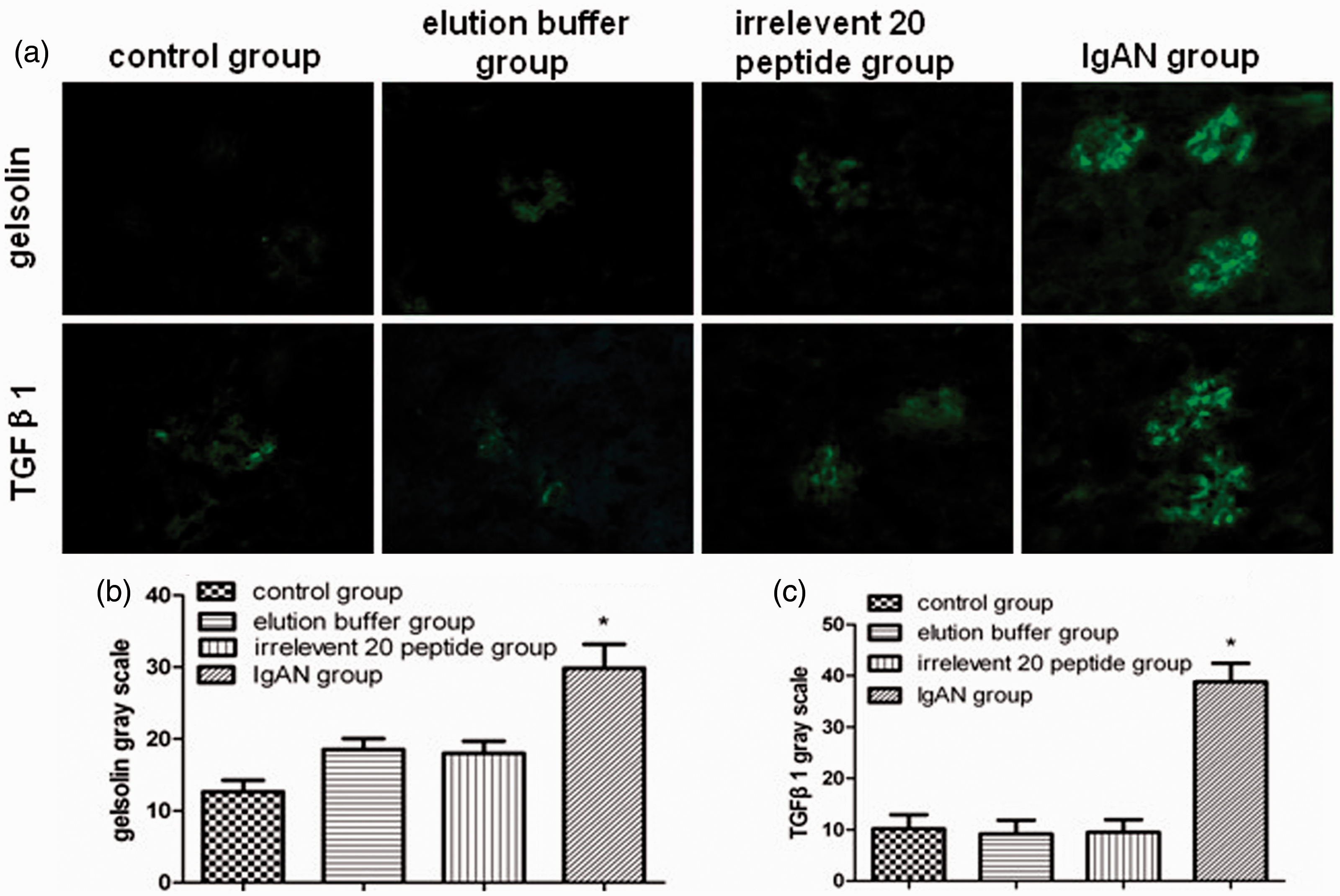
Western blot and real-time PCR for pGSN and TGFβ1
The relative levels of pGSN and TGFβ1 protein in the whole nephridial tissue in four groups are shown in Figure 3(a) and (b). The pGSN and TGFβ1 content was significantly higher in the IgAN group than in the other groups (P < 0.01); the quantified results are shown below the blot (Figure 3(c) and (d)). The relative levels of pGSN and TGFβ1 mRNA in the IgAN group were significantly higher than those in the other groups (P < 0.01), which were shown in Figure 4(a) and (b).
Western blot detection of pGSN and TGFβ1. The protein levels of (a) pGSN and (b) TGFβ1 in four groups were showed. The (c) pGSN and (d) TGFβ1 content in IgAN group (n = 10) were significantly higher than those in other groups (n = 30, each group 10); results were quantified analysis and showed below the blot. Comparisons between groups were tested by one-way ANOVA analysis and LSD test (*P < 0.01) Real-time PCR detection of pGSN and TGFβ1. The (a) pGSN and (b) TGFβ1 mRNA levels relative to control in IgAN group (n = 10) were significantly higher than those in other groups (n = 30, each group 10), which were tested by one-way ANOVA analysis and LSD test (*P < 0.01)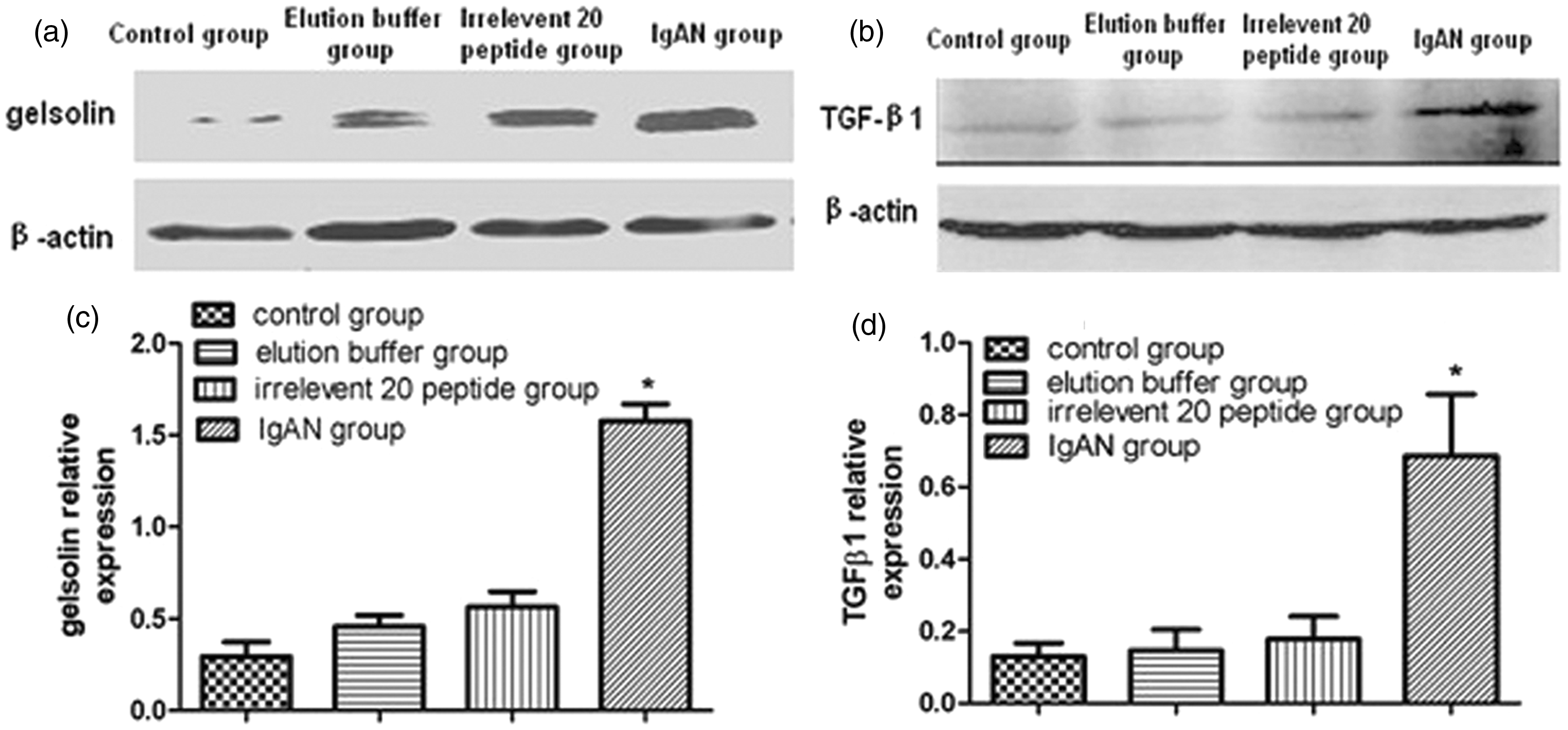
Urinalysis and serological ELISA findings
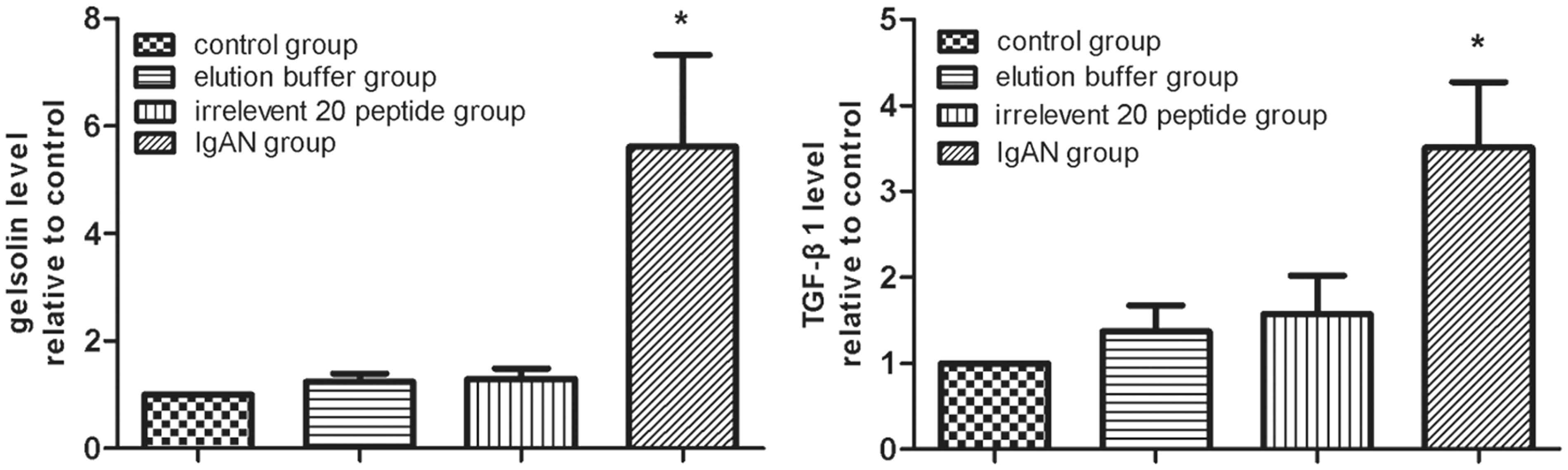
Figure 5(a) and (b) shows the differences in hematuria and proteinuria levels between the IgAN group and the other control groups. The hematuria and proteinuria levels were significantly higher in the IgAN group than other three groups (P < 0.01), but there were no significant differences between the three control groups (P > 0.05). The pGSN and TGFβ1 levels in the serum were detected with the double-antibody sandwich method, which indicated that the pGSN levels in the IgAN group were significantly decreased (P < 0.01) (Figure 5(c)) and TGFβ1 levels in the IgAN group were significantly increased than those in the other control groups (P < 0.01) (Figure 5(d)).
Urinalysis and serological ELISA test. The difference between IgAN group and other control groups in hematuria (a) and proteinuria (b) levels was showed. The hematuria and proteinuria levels were significantly higher in the IgAN group than other three groups (P < 0.01), which were tested by one-way ANOVA analysis and LSD test. The pGSN and TGFβ1 levels in the serum were detected by double-antibody sandwich method ELISA separately, which indicated that pGSN levels were significantly decreased (P < 0.01) (c) and TGFβ1 levels were significantly increased (P < 0.01) (d) in the IgAN group than those in the other control groups tested by one-way ANOVA analysis and LSD test (*P < 0.01). Total animals (n = 120) were used for statistics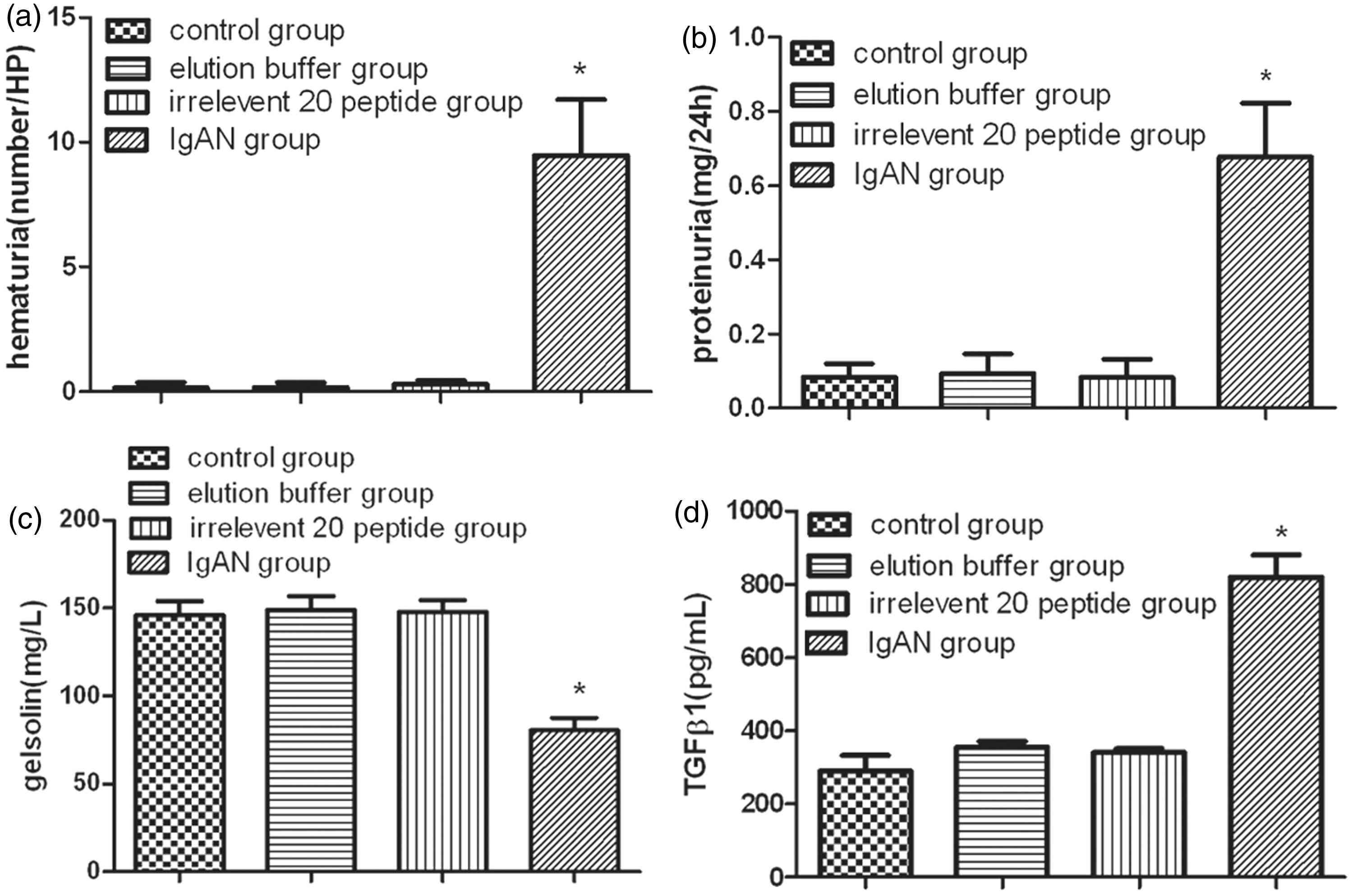
Oxidative stress findings
The SOD and CAT activities as well as the GSH and MDA content were detected by spectrophotometry. The SOD and CAT activities and GSH content in the IgAN group were significantly lower than the same measures in the other control groups (P < 0.01) (Figure 6(a) to (c)), while the MDA content was significantly higher in IgAN group than in the other groups (P < 0.01) (Figure 6(d)).
Detection of oxidative stress in four groups. The activities of SOD, CAT, and the contents of GSH and MDA were detected by spectrophotometry. The activities of (a) SOD, (b) CAT, and the content of (c) GSH show a significant decrease in IgAN group than other control groups (P < 0.01), while the content of (d) MDA was significantly increased in IgAN group (P < 0.01) tested by one-way ANOVA analysis and LSD test (*P < 0.01). Total animals (n = 120) were used for statistics.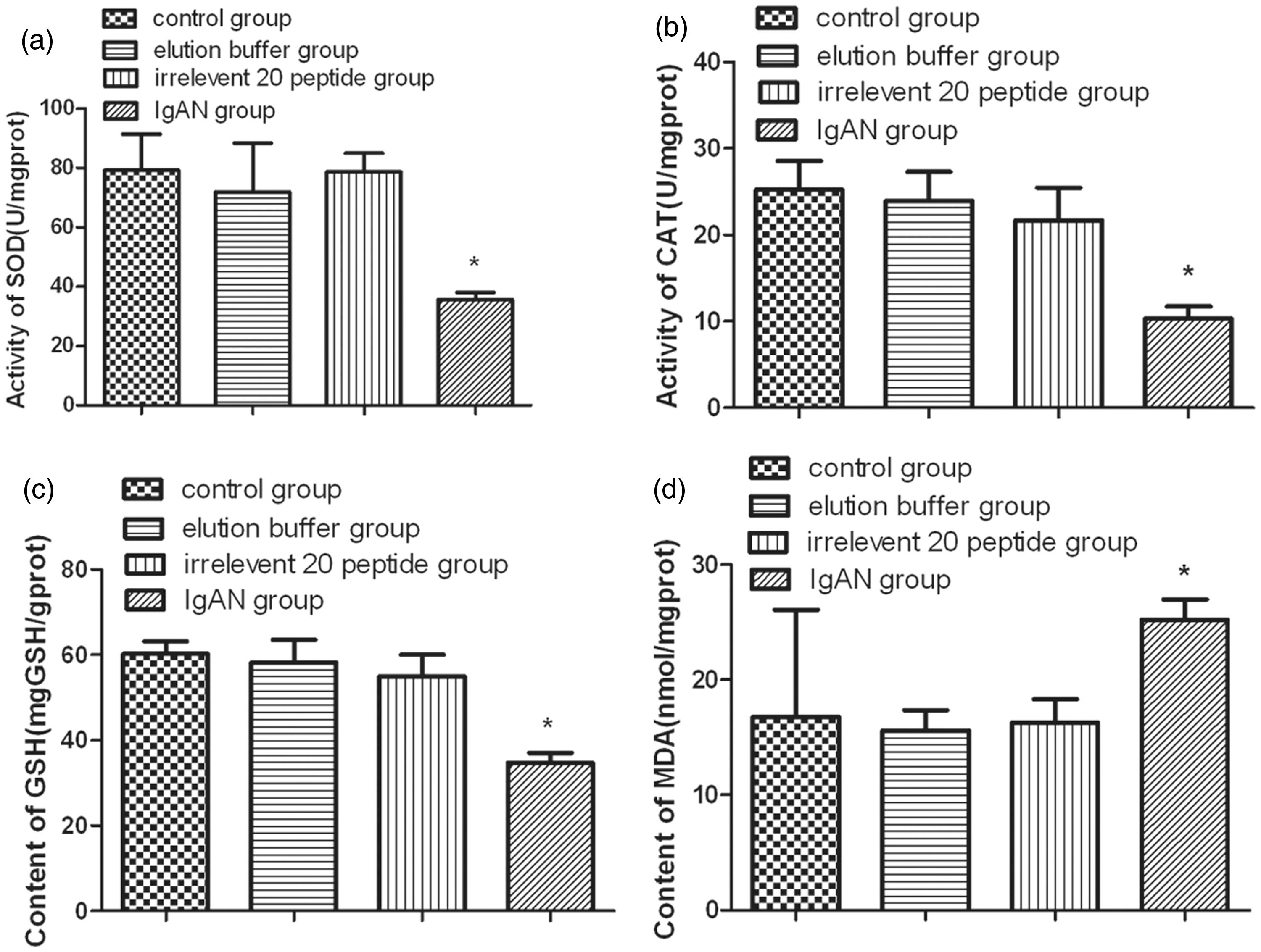
Correlation between serum pGSN levels and urinalysis results, pGSN and TGFβ1 immunofluorescence gray values, real time-PCR results, and oxidative stress test results in the IgAN group
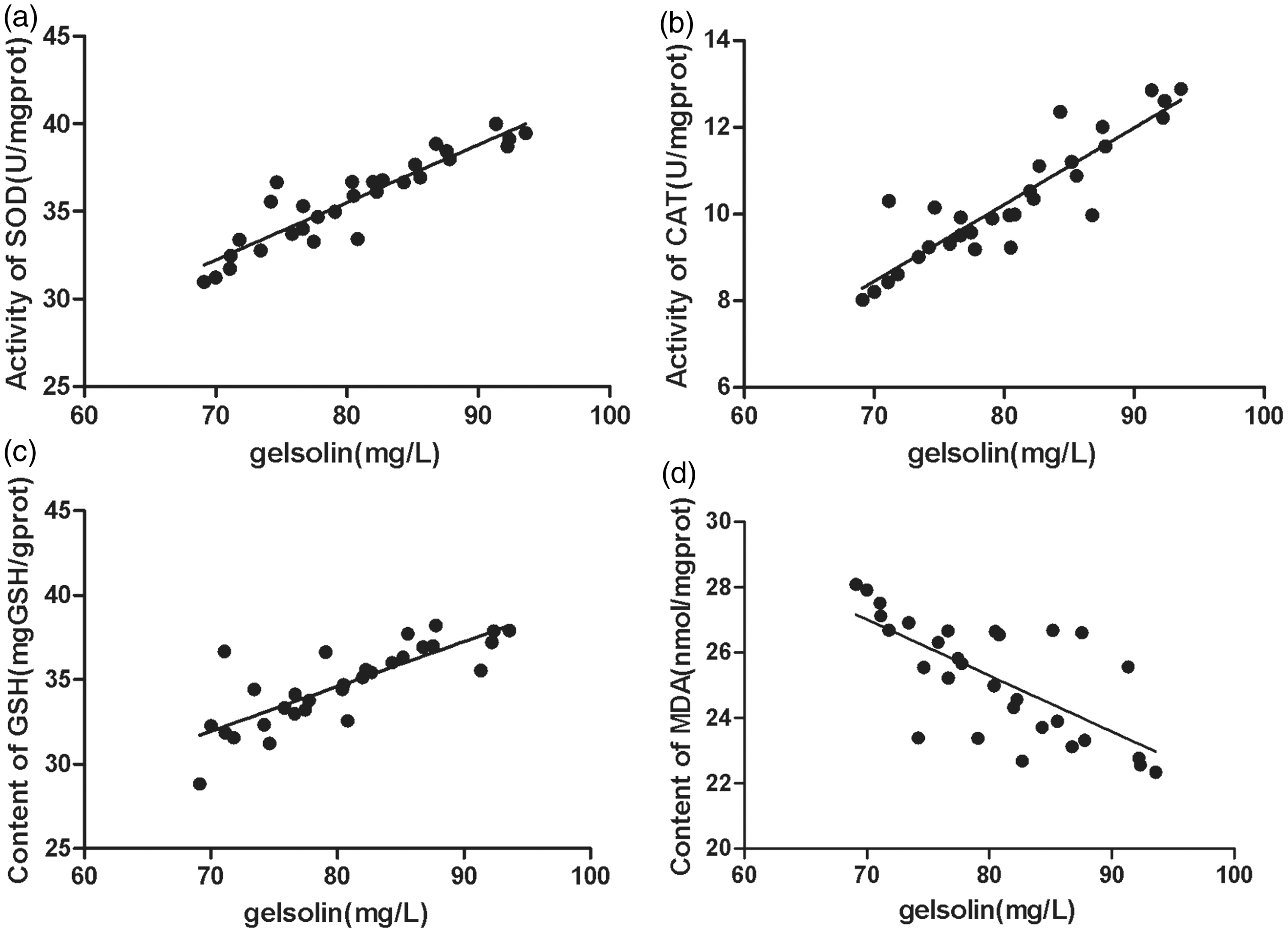
We examined the possible relationships between serum pGSN level and the other test results by analyzing correlations between them. The pGSN level was negatively correlated with hematuria (r = −0.911, P < 0.01) (Figure 7(a)), proteinuria (r = −0.745, P < 0.01) (Figure 7(b)), pGSN deposited in glomerulus (r = −0.803, P < 0.01) (Figure 7(c)), the pGSN mRNA level relative to the control (r = −0.701, P < 0.01) (Figure 7(d)), the TGFβ1 deposited in glomerulus (r = −0.922, P < 0.01) (Figure 7(e)), and the TGFβ1 mRNA level relative to the control (r = −0.817, P < 0.01) (Figure 7(f)). The pGSN level was positively correlated with SOD activity (r =+0.844, P < 0.01) (Figure 8(a)), the CAT activity (r = +0.811, P < 0.01) (Figure 8(b)), and GSH content (r = +0.641, P < 0.01) (Figure 8(c)), but negatively correlated with MDA content (r = −0.483, P < 0.05) (Figure 8(d)) in kidney tissues.
Correlation analysis between serum pGSN levels and urinalysis results, pGSN and TGFβ1 immunofluorescence gray values, real-time PCR results in the IgAN group. Correlation analysis between serum pGSN level and other testing results was investigated to examine the correlations between them. The pGSN level was negatively correlated with (a) hematuria (r = –0.911, P < 0.01), (b) proteinuria (r = –0.745, P < 0.01), (c) pGSN deposited in glomerulus (r = −0.803, P < 0.01), (d) the pGSN mRNA level relative to control (r = −0.701, P < 0.01), (e) the TGFβ1 deposited in glomerulus (r = −0.922, P < 0.01), and (f) the TGFβ1 mRNA level relative to control (r = −0.817, P < 0.01). The evaluation of correlations between results was performed using Pearson’s correlation. A P value < 0.05 was considered statistically significant. Total animals in IgAN group (n = 30) were used for statistics. Correlation of serum pGSN level with oxidative stress testing results in the IgAN group. The pGSN level was positively correlated with the activity of (a) SOD (r = +0.844, P < 0.01), the activity of (b) CAT (r = +0.811, P < 0.01), the content of (c) GSH (r = +0.641, P < 0.01), but was negatively correlated with the content of (d) MDA (r = −0.483, P < 0.05), which was performed by Pearson’s correlation. A P value < 0.05 was considered statistically significant. Total animals in IgAN group (n = 30) were used for statistics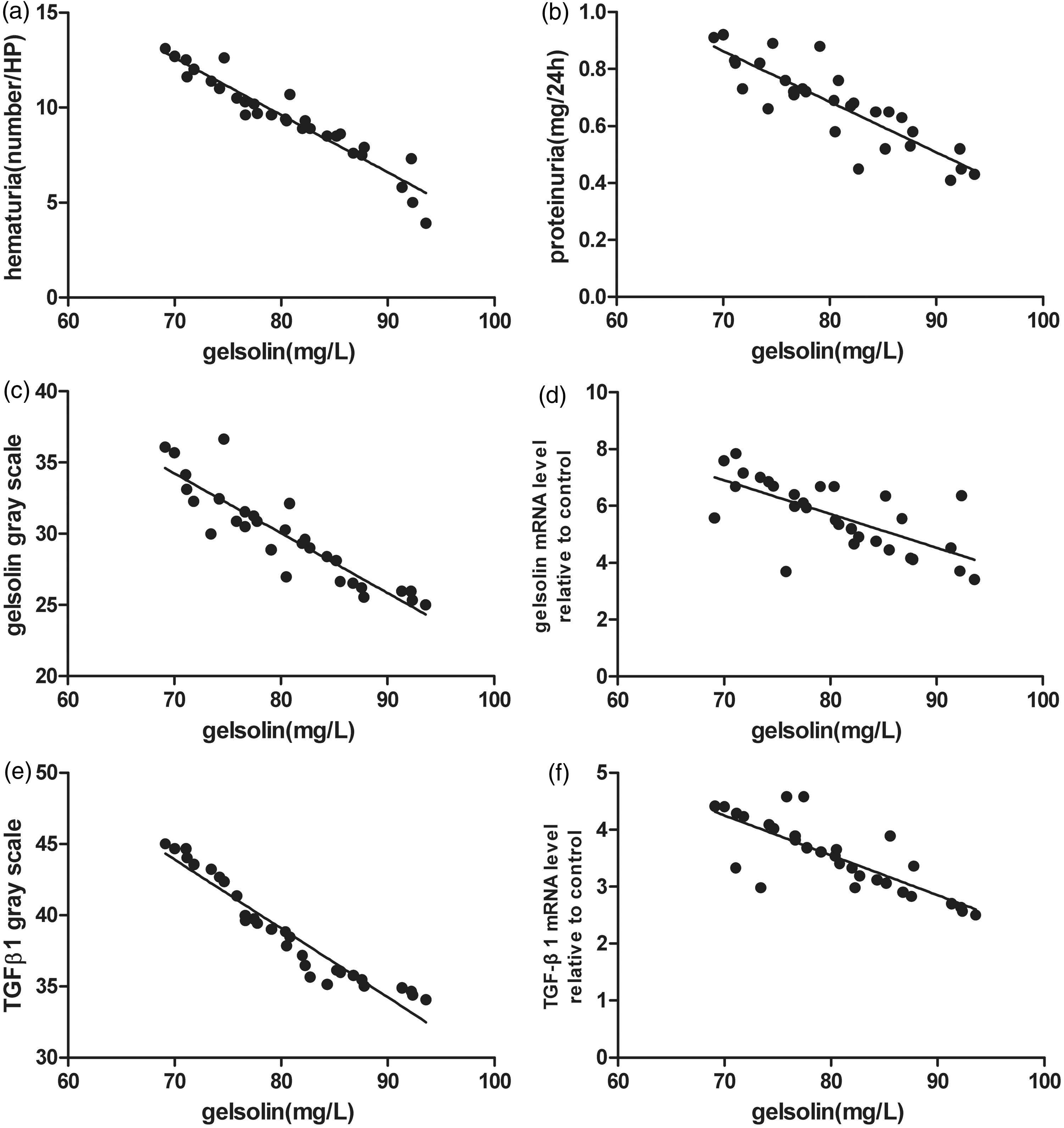
Discussion
IgAN is the most common primary glomerulonephritis worldwide and exhibits the pathological characteristics of extensive immunoglobulin-A deposition in the mesangial area, mesangial cell proliferation, and increased mesangial matrix. 21 The pathogenesis of IgAN is currently unclear, and because of the limitations in obtaining renal biopsy tissue, researchers have designed many different IgAN animal models to provide a research platform for IgAN. Miyawaki et al. used ddY mice, which have high levels of serum IgA, to establish an inbred mouse line that stably expresses high levels of serum IgA. High IgA (Higa) mice have abnormally high concentrations of IgA at 10–60 weeks of age, but do not exhibit a high urinary protein content and never develop hematuria. 22 In a previous study, we established a mouse IgAN model with the MBP-20-peptide fusion protein. 18 Pathological and clinical biochemical evaluations of this animal model revealed that the pathological characteristics of these mice are similar to the characteristics of human IgAN, and the mice in the experimental group have increased proteinuria and hematuria.
Gelsolin was first identified as a cytoplasmic actin-regulatory protein essential for cell locomotion and phagocytosis. pGSN is the extracellular isoform of gelsolin, the physiological significance of pGSN was far from apparent. The prior studies in some acute disease showed that the lower the levels of pGSN, the less favorable the prognosis of acute illness becomes.9,10,23 Rheumatoid arthritis represents a chronic inflammatory and autoimmune disease; Osborn found that pGSN is lower in the serum of patients with rheumatoid arthritis compared with healthy control patients and that gelsolin–actin complexes exist in the synovial fluid, 6 this finding that circulating pGSN levels decrease during chronic joint inflammation is consistent with observations of acute inflammatory diseases. Until now, no any research about the link between pGSN and glomerular nephritis was reported. As an immune complex glomerulonephritis, IgAN have similarity in the pathogenesy of autoimmunity with rheumatoid arthritis.
In our present study, we found that pGSN levels are lower in the sera of IgAN mice than in other control mice, this observed changes are similar to the observations of rheumatoid arthritis. We also found that pGSN levels are significantly higher in the kidneys of IgAN group mice than in the kidneys of mice in the other groups. Furthermore, we found a notable negative correlation between the serum pGSN levels and tissue pGSN test results in the IgAN group, this result of our animal model was the first to show that the serum pGSN levels correlate with tissue pGSN levels which deposited in glomeruli. Our present findings suggest that various pathogenic factors might stimulate the combination and consumption of serum pGSN in the IgAN group, causing the formation of macromolecular immune complexes that escape removal in the liver and are deposited in the kidney, leading to subsequent renal injury and glomerular fibrosis. In our study, the pGSN content in the serum of the IgAN mice was negatively correlated with hematuria and proteinuria levels, indicating that pGSN may be a potential biological marker of IgAN severity.
Fibrosis can occur in a variety of organs, and the persistent progression of fibrosis can cause organ damage, dysfunction, and failure. 24 Tissue fibrosis is a major cause of disability and death from many diseases. Of the various cytokines and signal transduction pathways involved in the pathogenesis of fibrosis, 25 the signal transduction mechanisms of the TGFβ family and the downstream Smad family are good candidates for research. 26 The TGFβ family consists of three subtypes (TGFβ1, TGFβ2, and TGFβ3), and researchers believe that TGFβ1 plays a critical role in the pathogenesis of glomerular fibrosis. 27 The pathogenesis of TGFβ1/Smad-mediated glomerular fibrosis has been widely recognized. There is increasing evidence that blockade of TGFβ1 action can ameliorate renal fibrosis.28–30 Researchers also found that fibrosis was modulated by inhibition of TGFβ1/Smad signaling pathways in adriamycin-induced nephropathy. 31 In our study, we determined the serum and tissue TGFβ1 levels, both of which were significantly higher in the IgAN group compared with the other groups. Our test result was consistent with this known evidence. Some researchers have reported that pGSN has an anti-inflammatory effect 16 in modeled pulmonary inflammation and fibrosis and that decreased levels of pGSN in the serum can increase the levels of some inflammatory factors such as TGFβ1; this increase can further trigger the inflammatory cytokine network and cause renal tissue damage and fibrosis.
Oxidative stress occurs when there is an imbalance between the generation of reactive oxygen species (ROS) and a biological system’s ability to detoxify the reactive intermediates. 32 Oxidative stress is associated with processes such as infection, inflammation, and ischemia/reperfusion injury. 33 Aerobic organisms have developed antioxidant defense mechanisms to minimize the damaging effects of ROS. A limited number of endogenous molecules that fight against oxidative stress have been investigated to date. Some researchers have reported that pGSN can counteract oxidative stress in vivo. 17 Low pGSN levels can cause an imbalance between oxidants and antioxidants, increasing ROS level. Some inflammatory cytokines such as TGFβ1 can promote oxidative stress, resulting in extensive damage to the nephrons and renal functions, including the histological damage of glomeruli and the tubulointerstitial region. In our study, the severity of oxidative stress in the IgAN model was evaluated by determining the activity or content of SOD, CAT, GSH, and MDA, all of which are important markers of oxidative stress. The test results showed that SOD, CAT, GSH activity or content in renal tissues were decreased in the IgAN group, and MDA content in renal tissues of the IgAN group was increased compared with those in the other groups; these oxidative stress markers dynamics were similar to known data, which have been reported in clinical observation of human IgAN by other researchers.34,35 Our study was the first to investigate oxidative stress markers in IgAN animal model and suggest that oxidative stress may be involved in the pathogenesis of IgAN by interactions with TGFβ1 and pGSN.
In this study, we performed a correlation analysis for pGSN content in serum and the other test results. Pearson’s correlation analysis showed that the pGSN content in serum and the TGFβ1 level in tissue had a significant negative correlation. The correlation analysis for pGSN content and oxidative stress markers indicated a significant negative correlation between pGSN levels and MDA content, but a significant positive correlation between pGSN levels and SOD or CAT activity and GSH content. These data provide a possible link between pGSN and IgAN fibrosis. Under endogenous or exogenous stimulation, pGSN may combine with other macromolecules to form an immune complex. In accordance with this hypothesis, the pGSN level in the serum of the IgAN group decreased sharply, leading to an anti-inflammatory effect and counteracting the oxidative stress that occurs in response to reduced pGSN levels. Therefore, the inflammatory cytokine network will be triggered, and oxidative stress will be activated. The interactions between the inflammatory cytokine network and oxidative stress will accelerate renal injury and fibrosis. On the other hand, the resultant immune complexes are too bulky to escape normal clearance mechanisms and reach the renal circulation overlying the mesangium. These complexes bind to mesangial cells and induce cellular activation. As a result, mesangial cells start to proliferate and overproduce extracellular matrix components, cytokines and chemokines, which may cause glomerular injury and fibrosis.
In summary, the findings in our present study suggest that pGSN levels could potentially be used as a biological marker for pathogenesis in the mouse IgAN model. Decreased pGSN levels correlate with cytokine networks and oxidative stress, and the interactions between pGSN, TGFβ1, and oxidative stress correlate with glomerular injury and fibrosis in the mouse IgAN model. However, the molecular mechanisms underlying the interactions between pGSN, TGFβ1, and oxidative stress were not elucidated in our study, and the process leading to mesangial cell activation and the initiation of glomerular inflammatory processes needs to be elucidated. Future investigation should use pGSN-deficient animals or infusion of recombinant pGSN, which will be more beneficial to clarify the role of pGSN in IgAN development.
Footnotes
Statement of Author Contributions
All authors participated in the design, interpretation of the studies, and analysis of the data and review of the manuscript; CH, LZ, XZ, and JT conducted the experiments, XJ supplied all of the reagents and animals in this study, CH and XJ wrote the manuscript, and LZ contributed to the statistical analysis of all the data.
Acknowledgements
We would like to thank the technician Fulai Chen in our department for his technical support. This work was supported by Harbin special fund for technological innovation (2006RFXXS035, 2007RFLXS017) and The Innovation Foundation of Harbin Medical University.