
Abstract
Although transplanting mesenchymal stem cells (MSCs) can improve cardiac function and contribute to endothelial recovery in a damaged artery, natural MSCs may induce neointimal hyperplasia by directly or indirectly acting on vascular smooth muscle cells (VSMCs). Receptor activity-modifying protein 1 (RAMP1) is the component and the determinant of ligand specificity of calcitonin gene-related peptide (CGRP). It is recently reported that CGRP and its receptor involve the proliferation and the apoptosis in vivo and in vitro, and the exogenous RAMP1 enhances the antiproliferation effect of CGRP in VSMCs. Here, we investigated the effects of MSCs overexpressing the human receptor activity-modifying protein 1 (hRAMP1) on heart function and artery repair in rabbit models of myocardial infarction (MI) reperfusion and carotid artery injury. MSCs transfected with a recombinant adenovirus containing the hRAMP1 gene (EGFP-hRAMP1-MSCs) were injected into the rabbit models via the ear vein at 24 h after carotid artery injury and MI 7 days post-EGFP-hRAMP1-MSC transplantation. The cells that expressed both enhance green fluorescent protein (EGFP) and CD31 were detected in the neointima of the damaged artery via immunofluorescence. EGFP-hRAMP1 expression was observed in the injured artery and infarcted myocardium by western blot analysis, confirming that the engineered MSCs targeted the injured artery and infarcted myocardium and expressed hRAMP1 protein. Compared with the EGFP-MSCs group, the EGFP-hRAMP1-MSCs group had a significantly smaller infarcted area and improved cardiac function by 28 days after cell transplantation, as detected by triphenyltetrazolium chloride staining and echocardiography. Additionally, arterial hematoxylin–eosin staining revealed that the area of the neointima and the area ratio of intima/media were significantly decreased in the EGFP-hRAMP1-MSCs group. An immunohistological study showed that the expression of α-smooth muscle antigen and proliferating cell nuclear antigen in the neointima cells of the carotid artery of the EGFP-hRAMP1-MSCs group was approximately 50% lower than that of the EGFP-MSCs group, suggesting that hRAMP1 expression may inhibit VSMCs proliferation within the neointima. Therefore, compared with natural MSCs, EGFP-hRAMP1-engineered MSCs improved infarcted heart function and endothelial recovery from artery injury more efficiently, which will provide valuable information for the development of MSC-based therapy.
Keywords
Introduction
Even though percutaneous coronary intervention (PCI) has been extensively used to recover blood flow after myocardial infarction (MI) and improve heart function, the ischemic complications of PCI, including death, periprocedural MI, restenosis, delayed endothelialization, and late in-stent thrombosis, continue to occur. However, there has been great progress in stem cell-based therapies for cardiac repair in the past decade.1,2 Stem cells are a class of multipotent/pluripotent cells that are capable of self-renewal and clonal expansion, 3 and the seed cells that have been used for cell therapy include embryonic stem cells, endothelial progenitor cells, and mesenchymal stem cells (MSCs).4,5 MSCs, which are found in bone marrow, adipose tissue, umbilical cord blood, and peripheral blood, are an optimal candidate for myocardial repair owing to their inherent ability to both self-renew and differentiate into multiple lineages including osteoblasts, chondrocytes, adipocytes, endothelial cells, and cardiocytes. In addition, they are suitable for transplantation because of their immunoprivilege characteristics (a low expression of human leukocyte antigen major histocompatibility complex (HLA MHC) class I proteins and no expression of CD40, CD80, and CD86). 6 Furthermore, MSCs have the potential to differentiate into vascular smooth muscle cells (VSMCs) and may be the source of proliferating VSMCs during neointima formation after vascular injury.7,8 However, normal MSCs may induce to neointimal hyperplasia and restenosis by directly or indirectly acting on VSMCs. 9 MCSs have demonstrated a significant effect on the repair of the infarcted heart by improving vascular perfusion; reducing fibrosis; and releasing angiogenic, antiapoptotic, and mitogenic factors. 10 Recently, genetically modified MCSs, such as CXCR4-, heme oxygenase-1 (HO-1)- or Bcl-2-overexpressing MCSs, have proven to be more efficient at ameliorating infarcted myocardium than administering MCSs alone.11–13
Calcitonin gene-related peptide (CGRP) is one of the most well-known potent vasodilators and can regulate vascular tone and other aspects of vascular function. The receptors for CGRP include the calcitonin receptor-like receptor (CRLR), the receptor activity-modifying protein 1 (RAMP1), and the receptor component protein. 14 Of the known RAMP proteins, RAMP1 confers ligand specificity for CGRP, whereas an association of CRLR with RAMP2 or RAMP3 results in a receptor with a high affinity for adrenomedullin. 14 The relaxation of the artery in response to CGRP is dependent on RAMP1 expression. The response to CGRP is augmented after the increased expression of RAMP1 in VSMCs in culture. RAMP1 overexpression increased CGRP-induced vasodilation and protected against angiotensin II-induced endothelial dysfunction as well as prevented VSMCs proliferation. 15 In the present study, we tested the effects of human RAMP1-overexpressing MSCs on infarcted heart function and intimal hyperplasia by means of cell transplantation in rabbit models for MI reperfusion and carotid artery injury. Our data showed that hRAMP1 overexpression in MSCs through genetic modification significantly inhibit neointimal proliferation and improve infarcted heart function.
Materials and methods
Plasmid constructs and virus production
The gene that encodes human RAMP1 was excised from the pOTB7-hRAMP1 vector (a kind gift from Dr Andrew F Russo, University of Iowa, USA) via the EcoRI/Xhol restriction sites and cloned into the pShuttle-enhance green fluorescent protein (EGFP) vector and named pShuttle-EGFP-hRAMP1. pAdxsi-EGFP or pAdxsi-EGFP-hRAMP1 was generated by cutting the EGFP or EGFP-hRAMP1 sequence from the pShuttle-EGFP or pShuttle-EGFP-hRAMP1 vector via I-CeuI/I-SceI digestion and subcloned into the pAdxsi vector (SinoGenoMax Co. Ltd, China) at the corresponding sites. Adenovirus particles were produced by transfecting pAdxsi-EGFP-hRAMP1 or pAdxsi-EGFP into the 293 T packaging cell line. The generated adenovirus particles were used to create more transfected 293 T cells and then collected and purified using conventional techniques. 16
MSCs isolation, expansion, and flow cytometry
Bone marrow-derived MSCs were obtained using density gradient centrifugation and adherent culture, as previously described. 17 The isolated MSCs were cultured in deulbecco's modified eagle medium (DMEM) containing 10% fetal bovine serum (FBS). To characterize the MSCs, the third-passage cultured MSCs were analyzed using a fluorescence-activated cell sorter (FACS). Briefly, a total of 2 × 105 cells were incubated for 30 min at 4℃ with monoclonal FITC-conjugated mouse anti-rabbit antibodies against CD45, CD29, and CD90 (US Biological) followed by detecting the FITC-positive cells using a FACS machine (Becton-Dickinson, US).
After 3–5 passages, cultured MSCs were transfected with either the pAdxsi-EGFP-hRAMP1 or pAdxsi-EGFP vector, named as EGFP-hRAMP1-MSCs or EGFP-MSCs; the GFP-positive infected cells were visualized using fluorescence microscopy or analyzed by FACS analysis.
Rabbit models of MI surgery and carotid artery injury
MI and carotid artery injury were induced in male New Zealand rabbits (each 2.0 ± 0.5 kg and from the animal center of Third Military Medical University, China) via provisional ligation of the left ventricular branch (LVB) and balloon dilation, respectively, as previously described.18,19 The investigation conforms to the Guide for the Care and Use of Laboratory Animals by the Ethics Committee of Zunyi Medical College. Briefly, the animals were anesthetized using pentobarbital sodium (3%, 1 mg/kg), and the LVB was permanently ligated for 60 min followed by reperfusion. Penicillin was used to prevent infection by dosing with 800,000 units per day. The standard of success for having induced a MI was that the myocardial tissue became pale at the regions relevant to the ligated artery and that the ST segment was permanently elevated as shown by an animal electrocardiogram. Forty-eight hours after MI, the animals were again anesthetized by the same method, and the left carotid artery was exposed via a midline incision of the ventral side of the neck. Two ligatures were placed proximally and distally to the external carotid artery. After temporary occlusion of the common carotid artery, a small transverse arteriotomy was performed. A 2.0F balloon (0.36 mm in diameter, COOK, Cordis Corporation, USA) was inserted into the common carotid artery and then dilated to 6 atm and dragged back and forth for 1 min 3 times with 1-min intervals. When the intima was peeled off, the ligatures were removed and blood flow was recovered. After 24 h, all successful models were injected with EGFP-hRAMP1-MSCs (EGFP-hRAMP1-MSCs group, n = 18) or EGFP-MSCs (EGFP-MSCs group, n = 18) (2 mL solution of approximately 1 × 108 cells) via the ear vein. Phosphate buffered saline (PBS) was injected for the control group (n = 18). At 7, 14, and 28 days after cell transplantation, six animals in each group were sacrificed and histologically examined.
Morphometric analysis of the injured carotid artery
The injured carotid arteries were carefully harvested 7, 14, and 28 days after transplanting EGFP-hRAMP1-MSCs or EGFP-MSCs and fixed using 4% paraformaldehyde and separated into two parts. One part was paraffin embedded for hematoxylin–eosin (HE) staining and immunohistochemistry analysis of CD31, α-smooth muscle antigen (α-SMA), and proliferating cell nuclear antigen (PCNA) expression. The other part was used for immunofluorescence analysis of the expression of EGFP, EGFP-hRAMP1, and CD31. The neointimal areas and intima/media (I/M) ratios analysis were performed on HE-stained sections and recorded via a Leica QWin Plus analysis system.
For immunohistochemistry assay, the sections (5 µm) were incubated in a 1 : 200 dilution of mouse anti-rabbit monoclonal CD31 (also known as platelet and endothelial cell-adhesion molecule 1, Abcam Bioscience). Adjacent sections were incubated in 1 : 100 monoclonal mouse anti-rabbit α-SMA and PCNA (Abcam Bioscience) overnight at 4℃. All sections were then incubated in 1 : 400 biotin conjugated secondary antibody (goat anti-mouse, Abcam Bioscience) at 37℃ for 30 min and stained with DAB. The section pictures were taken by optical microscopy and analyzed using the Leica QWin Plus analysis system.
For immunofluorescence assay, the sections of the 7-day injured carotid arteries were incubated with goat anti-GFP polyclonal antibody and rat anti-CD31 monoclonal antibody (Abcam Bioscience) overnight at 4℃. After washing 3 times with PBS, the tissues sections were incubated with fluorescein isothiocyanate (FITC)-labeled donkey anti-goat IgG and tetramethyl rhodaine isothiacyanate (TRITC)-labeled goat anti-mouse IgG for 60 min at 37℃. Fluorescence was observed using a Zeiss inverted microscope. Instead of using a primary antibody, the sections were incubated in PBS as a negative control.
Morphometric analysis of the infarcted heart
Twenty-eight days after transplanting the EGFP-hRAMP1-MSCs or EGFP-MSCs, the myocardial tissues of the rabbits were cut into 1–2 µm thick cross-sections. All sections were incubated in tetrazolium chloride (TTC) solution at 37℃ for 15 min. The infarcted myocardium showed no color, while the noninfarcted tissues were brick-red. The sections were fixed using 4% paraformaldehyde for 15 min and photographs of the sections were taken using a digital camera. The infarcted area and the whole ventricle area were measured by the Image tool 3.0 software. Some sections were used for HE staining and Masson’s trichrome dying.
Western blot analysis
The infarcted myocardium and injured vascular tissues were harvested on the 7th day after cell transplantation. Lysis buffer was added to the samples, and the samples were homogenized on ice and then subjected to ultracentrifugation under 4℃. The concentration of the proteins extracted from the supernatants was determined using a bicinchonininc acid (BCA) Protein Quantitative Kit (Abcam Bioscience). Equal amounts of protein (50 µg) were separated by 10% sodium dodecyl sulfate (SDS)-polyacrylamide gel electrophoresis, and the expression of hRAMP1 was detected using a mouse anti-hRAMP1 polyclonal antibody (Abnovo, USA).
Measurement of heart function by echocardiography
Twenty-eight days after cell transplantation, the heart function of the animals was evaluated with 12 MHz Color Doppler Flow Imaging (Philip Corporation) for the mitral valve M-type curve of the left ventricle, the left ventricular diastolic dimension (LVDd), the left ventricular systolic dimension (LVDs), the left ventricular ejection fraction (LVEF), and the left ventricular fractional shortening (LVFS). The results were attained by determining the mean value of three consecutive cardiac cycles.
Statistical analysis
Statistical analysis was performed using the SPSS 16.0 software. The results are presented as the mean ± SEM and were compared among experimental groups using one-way ANOVA followed by the Newman–Keuls multiple comparison test. Values of P < 0.05 were considered to be significant.
Results
Characterization and transfection of MSCs with recombinant adenoviruses
The MSCs showed a polygon or a long spindle shape after 24 h cell culture and then gradually showed a uniform, long spindle shape in the passage cells similar to that of fibroblastic cells (Figure 1A (a) and (b)). The MSCs of the third passage were identified by FACS (Figure 1B (a) to (c)). The result demonstrated that the MSCs strongly expressed CD29 (95.79%) and CD90 (98.59%) while weakly expressing CD45 (12.49%). Seventy-two hours posttransfection with pAdxsi-EGFP-hRAMP1 or pAdxsi-EGFP vector at 200 of MOI, 80% of the MSCs exhibited green fluorescence (Figure 1A (c) and (d); 1B (d)), indicating successful gene transfer. Importantly, EGFP-hRAMP1- or EGFP-MSCs grew well without abnormal morphological changes during a 3-week observation (data not shown).
(A). The MSCs were efficiently transfected with recombinant adenoviruses. The primary MSCs on 7 days grew nonuniformly and colony like (a, b). The third passage of MSCs showed green fluorescence at 72 h posttransfection with pAdxsi-EGFP-hRAMP1(c) or pAdxsi-EGFP vector at 200 of ultiplicity of infection (MOI) (d). Magnification ×100. (B). Flow-cytometric analysis of MSCs. Most MSCs expressed CD29 and CD90 but little CD45 (a, b, c). The EGFP expressions in EGFP-hRAMP1-MSCs were detected via FACS assay (d). (A color version of this figure is available in the online journal)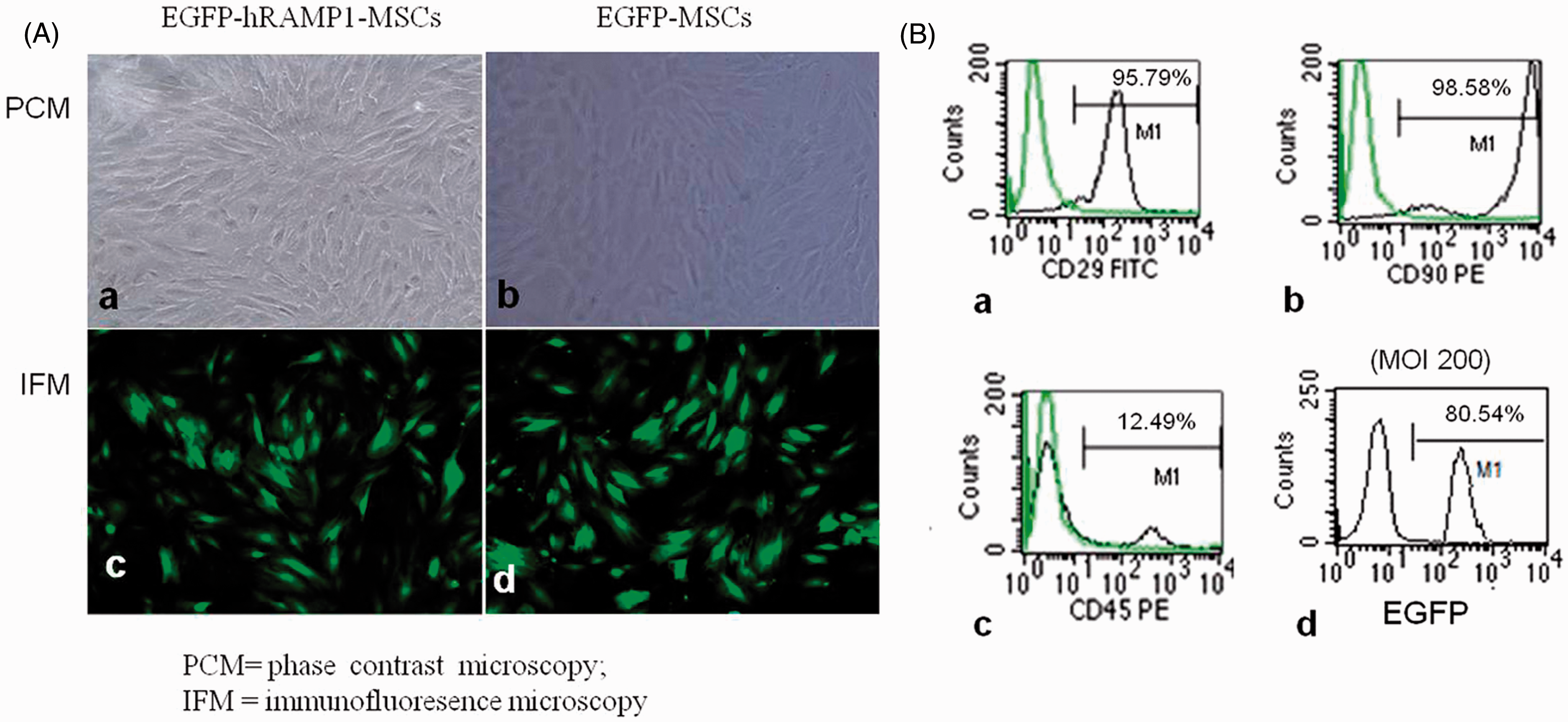
EGFP-hRAMP1-MSCs were targeted to and expressing in the injured carotid artery and infarcted myocardium
The expression of CD31 indirectly indicates the level of re-endothelialization in injured vessels given that it is one of the specific antigens of endothelial cells. With immunohistochemistry and immunofluorescence, we observed that CD31 protein highly accumulated in the endothelial cells of the injured vessels of animals at 7 days posttransplantation with EGFP-hRAMP1-MSCs (Figure 2(a)). Interestingly, the anti-GFP immunofluorescence also showed that most of the endothelial cells were GFP positive and colocalized with CD31 protein, suggesting that MSCs transfected with pAdxsi-EGFP-hRAMP1 vector were targeted to the injured vessels and contributed to the endothelialization of the injured artery (Figure 2(a)). Meanwhile, MSCs that were transfected with pAdxi-EGFP-hRAMP1 vector were recruited to the region and the border of the MI. Therefore, significantly higher levels of hRAMP1 were detected in both the infarcted myocardium and the injured vessel in the EGFP-hRAMP1-MSCs group, as demonstrated by western blot analysis (Figure 2(b)), confirming the expression of the transgene (hRAMP1) after adenoviral gene transfer.
(a) Seven days after cell transplantation, immunofluorescence showed that the neointima of the damaged artery expressed both EGFP and CD31 in the EGFP-hRAMP1-MSCs group (n = 6) but not in the control group (n = 6). Immunohistochemistry also showed an increased expression of CD31 protein in the endothelial cells of the injured vessels of the EGFP-hRAMP1-MSCs animal but not in those of the control group. Magnification is 100 × Scale bar = 50 µm. (b) hRAMP1 protein in both the myocardium (left upper panel) and the carotid artery (right upper panel) of the different groups (n = 6). Red and white arrowheads indicate EGFP and CD31-positive cells, respectively. (A color version of this figure is available in the online journal)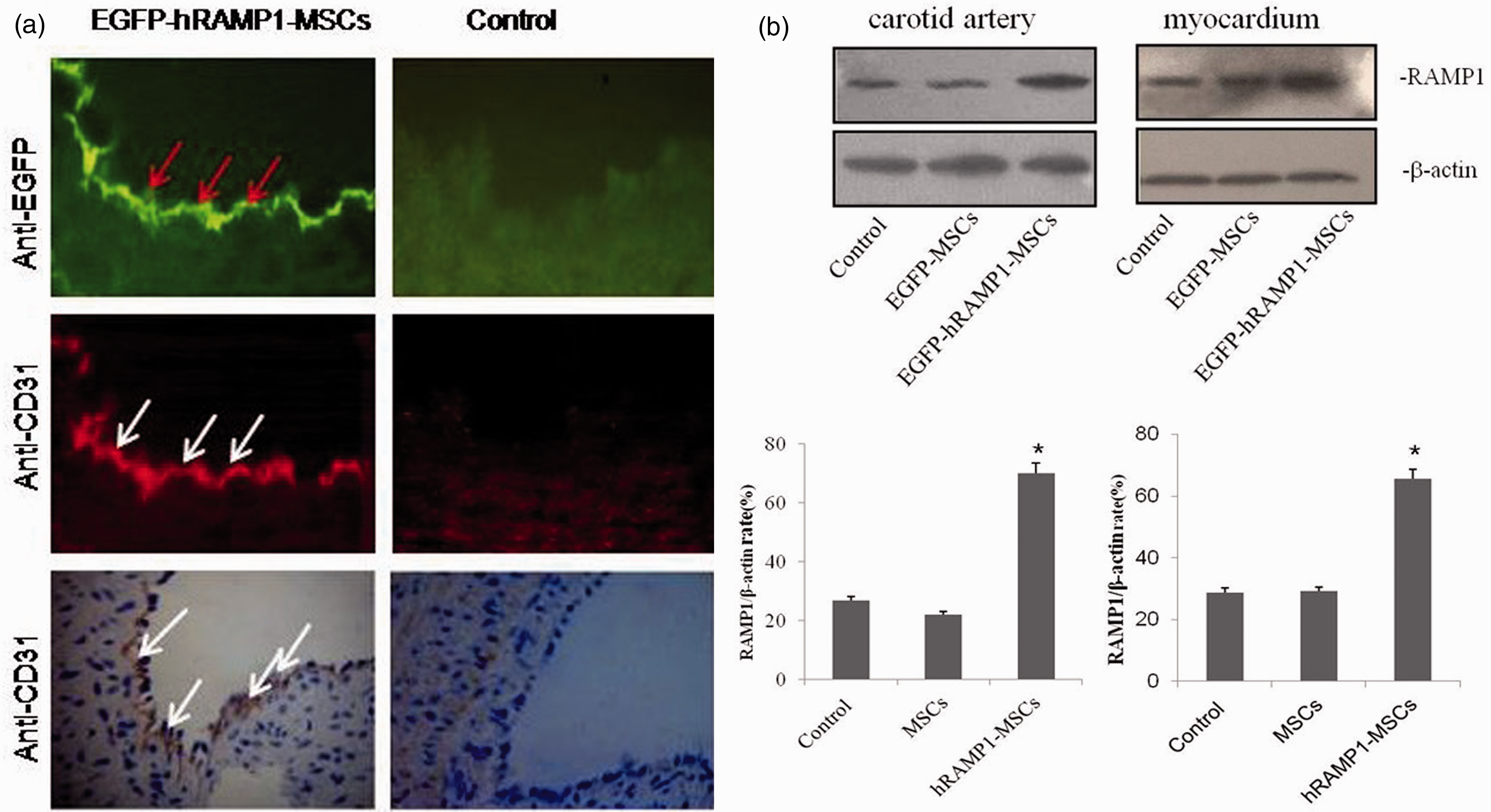
Impact of transplanting EGFP-hRAMP1-MSCs on intima hyperplasia of the injured carotid artery
After vascular injury, an excessive accumulation of cells at the intima was observed on day 14 and 28 in control animals and resulted in a significantly thickened intima (Figure 3(a) and (b), left panel). However, the increases in the intimal area of the EGFP-MSCs and EGFP-hRAMP1-MSCs groups were significantly lower than that of the control group. Moreover, 28 days after cell delivery, the neointimal area of the EGFP-hRAMP1-MSCs group was only 50% of that of the EGFP-MSCs group (Figure 3(a) and (b), left panel). As expected, the area ratio of I/M for both the EGFP-MSCs group and the EGFP-hRAMP1-MSCs group was lower than that of control group. Among the three groups, the EGFP-hRAMP1-MSCs group displayed the lowest I/M ratio (Figure 3(b), right panel). The results suggested that MSCs, especially EGFP-hRAMP1-MSCs, may reduce intima hyperplasia by inhibiting intimal cell proliferation.
Effects of transplanting EGFP-hRAMP1-MSCs on intima hyperplasia of the injured carotid artery. (a) Light micrographs of injured rabbit carotid arteries at 7, 14, and 28 days post-MSCs transplantation (n = 6). HE staining, magnification is 100 × Scale bar = 200 µm. (b) Quantitative analysis of the area of neointima (left panel) and the area ratio of neointima to media. The data are represented as the mean ± SD, *P < 0.05. (A color version of this figure is available in the online journal)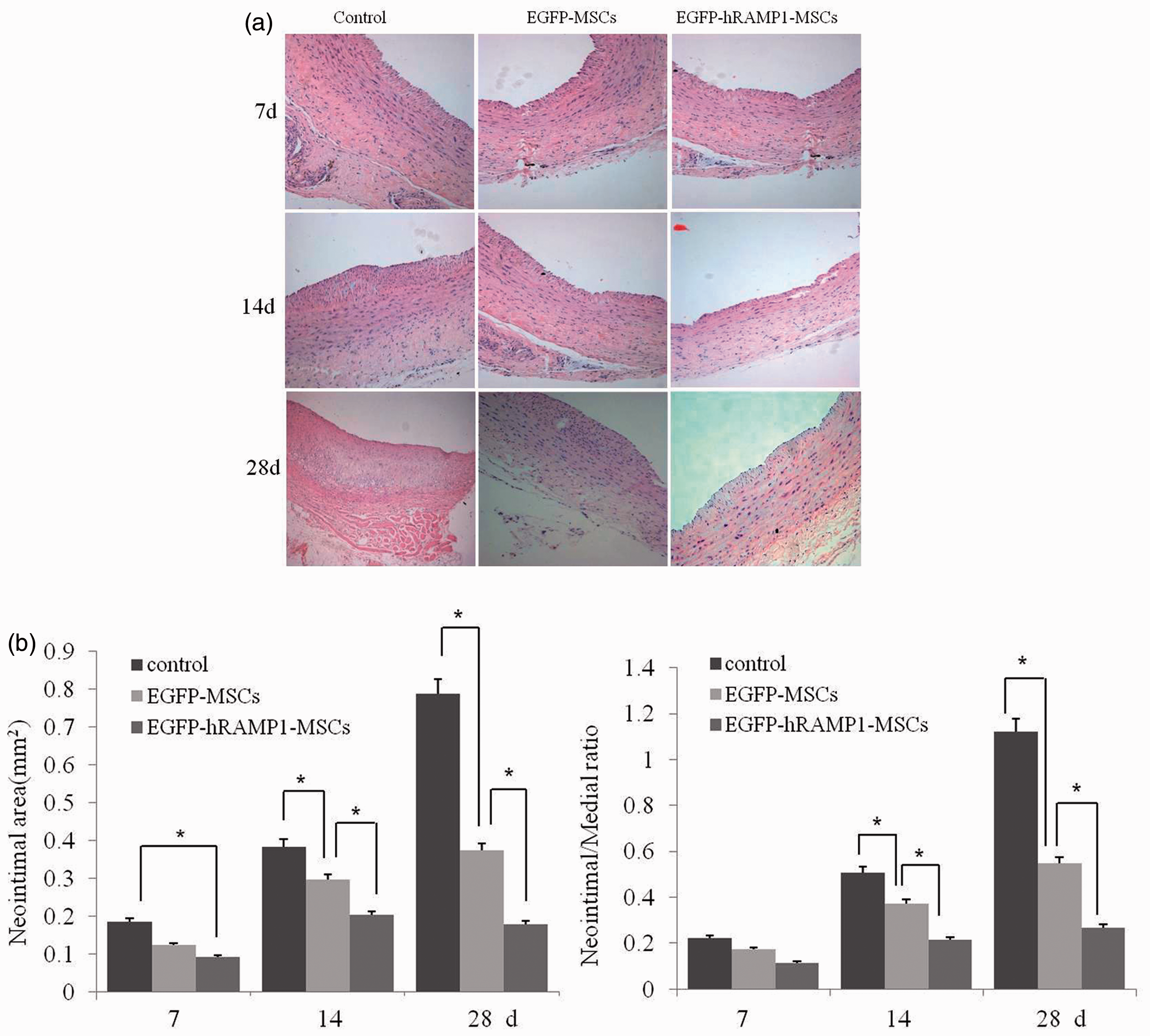
We further investigated the neointimal cells by immunostaining for α-SMA and PCNA in the neointima of the injured carotid artery. α-SMA, a specific marker of VSMCs, was stained for because a large number of smooth muscle cells had been found to immigrate and proliferate in the intimal area in response to injury. Both α-SMA and PCNA were stained sepia and α-SMA located in cytoplasm but PCNA in nucleus. In the control group, a great number of α-SMA- or PCNA-positive cells were observed in the neointima. Meanwhile, the PCNA-positive cells also existed in the tunica media of all groups (Figure 4(a)). Compared with the control group, the expressions of PCNA and α-SMA were reduced in the neointima of the EGFP-hRAMP1-MSCs and EGFP-MSCs groups, especially in the EGFP-hRAMP1-MSCs group (Figure 4(b)). These results indicated that the inhibition of VSMC proliferation contributed to the decrease in intima hyperplasia in the EGFP-hRAMP1-MSCs group.
Effects of transplanting EGFP-hRAMP1-MSCs on the smooth muscle cell proliferation in the intima of the injured carotid artery. (a) Immunohistochemistry staining for α-SMA and PCNA showed fewer labeled cells in the intimal of the carotid artery of the EGFP-hRAMP1-MSCs group (n = 6). Magnification is 400 × Scale bar = 50 µm. (b) Quantification of smooth muscle cell proliferation in the intima. The data are represented as the mean ± SD (n = 6), *P < 0.05. (A color version of this figure is available in the online journal)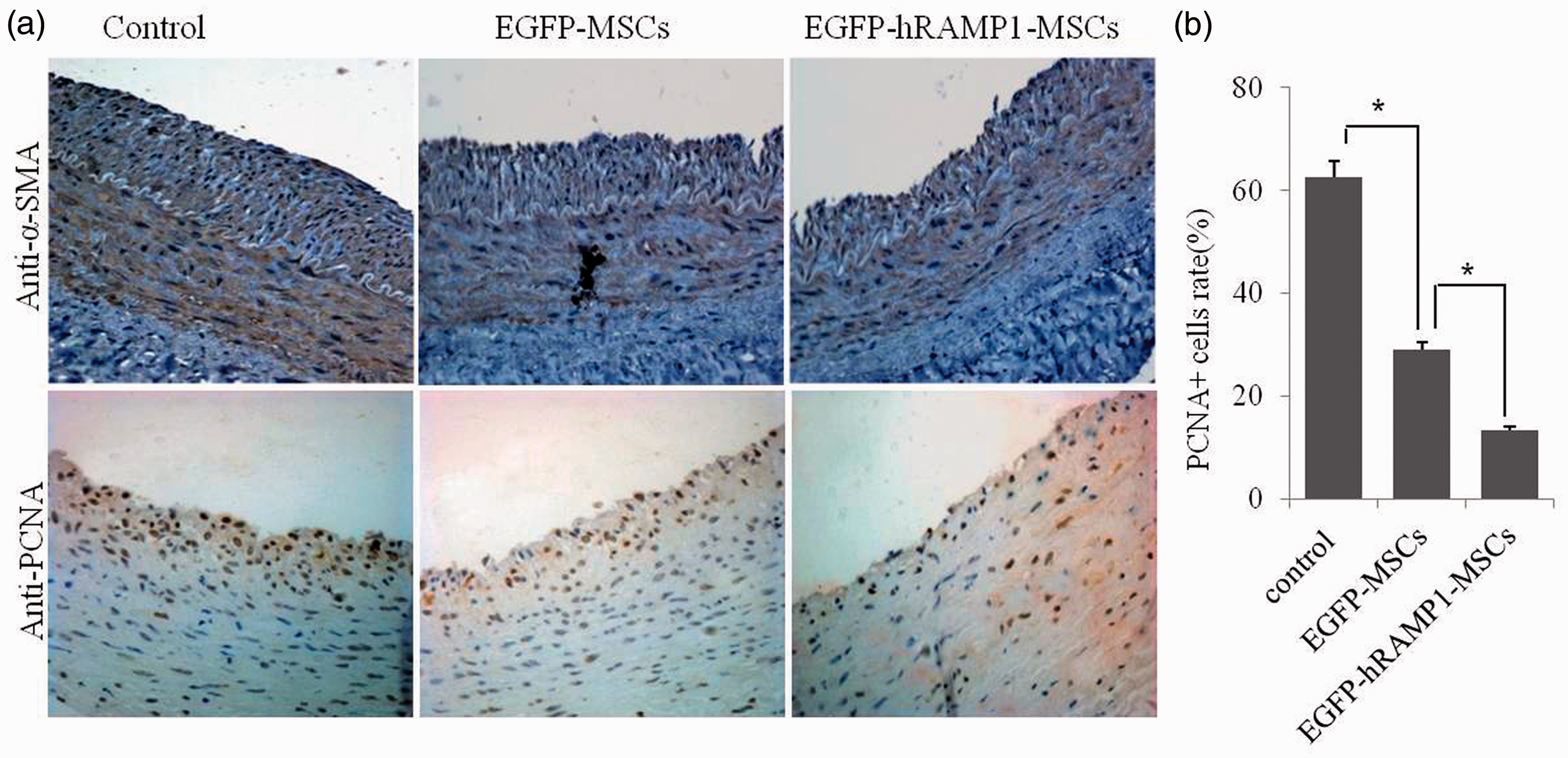
Improvement of cardiac function after transplanting EGFP-hRAMP1-MSCs by reducing myocardial fraction and fibrosis
We examined the effect of transplanting MSCs on cardiac function, as described in “Materials and methods” section. As shown in Figure 5, a 25–50% decrease in the LVDd and LVDs and an increase in the fractional shortening (FS) and the LVEF were observed in both the EGFP-hRAMP1-MSCs and EGFP-MSCs groups. Importantly, the heart function of the EGFP-hRAMP1-MSCs group was significantly improved compared with that of the other two groups. These results declared that transplanting MSCs, especially EGFP-hRAMP1-MSCs, has the potential for improving heart function after MI.
Effects of transplanting EGFP-hRAMP1-MSCs on cardiac function. Twenty-eight days after transplantation, the heart function of animals was evaluated with an ultrasonic cardiogram. LVEF: left ventricular ejection fraction; LVFS: left ventricular fractional shortening; LVDd: left ventricular diastolic dimension; LVDs: left ventricular systolic dimension. The data are represented as the mean ± SD (n = 6), *P < 0.05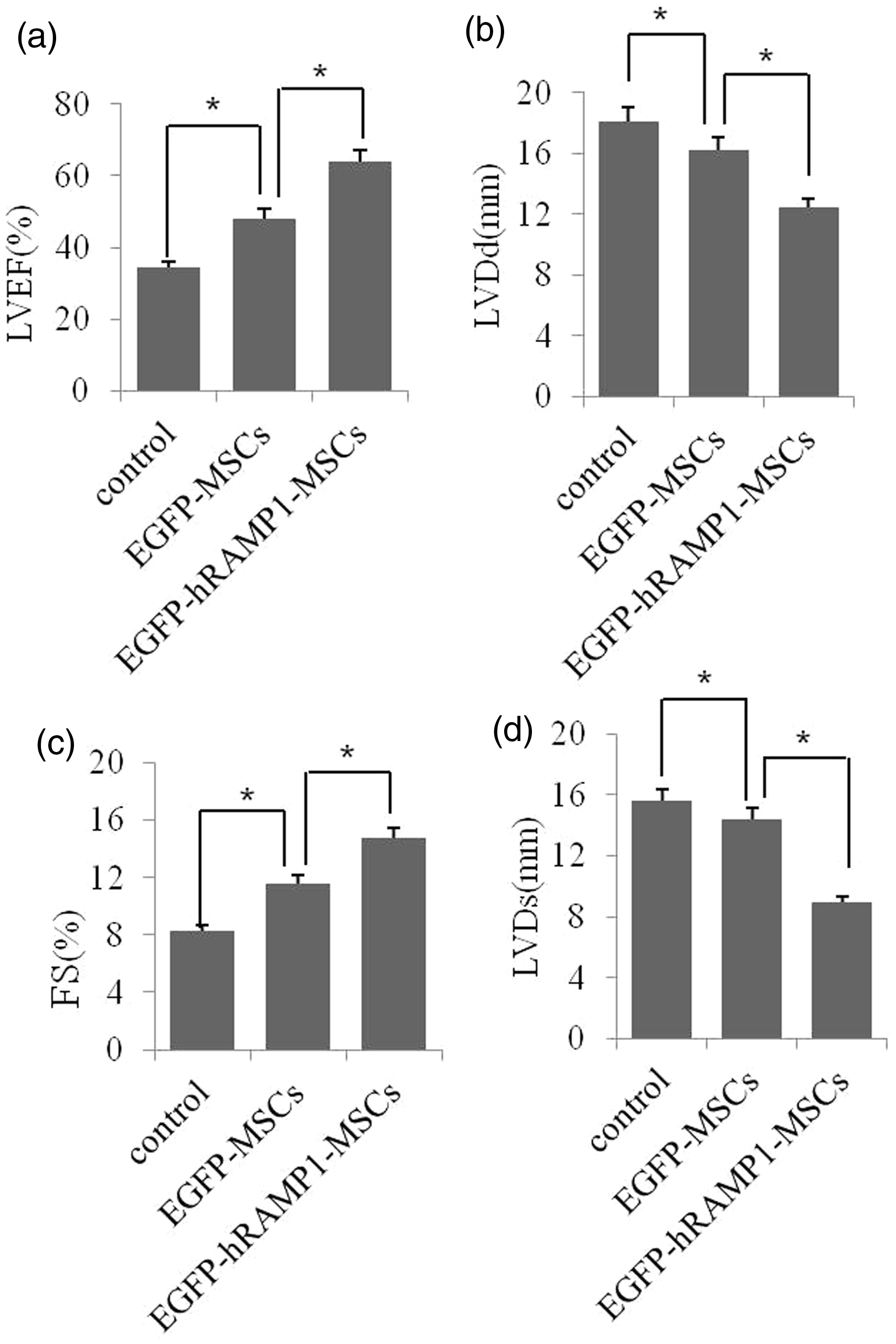
Twenty-eight days after cell delivery, TTC staining in the myocardium showed that the normal myocardium displayed a brick-red color whereas the infarcted tissue was white. Compared with the control group, the infarcted areas in both the EGFP-MSCs group and the EGFP-hRAMP1-MSCs group were smaller (Figure 6(a), upper panel). Quantitative analysis also demonstrated that the infarction volume of the EGFP-hRAMP1-MSCs group was significantly smaller than that of both the EGFP-MSCs and control groups (Figure 6(a), lower panel). HE staining and Masson’s trichrome staining revealed that the myocardial cells surrounding the infarcted myocardium were disorganized or disappeared, and a fiber scar formed (Figure 6(b), control group). However, compared with the control group, the areas of fiber scar tissue surrounding the infarcted zone were smaller, and the myocardial cells were highly organized in both EGFP-MSCs and the EGFP-hRAMP1-MSCs groups, especially in the EGFP-hRAMP1-MSCs group (Figure 6(b)). The results indicate that transplanting EGFP-hRAMP1-MSCs and EGFP-MSCs significantly attenuated the development of myocardial fibrosis and also increased the myogenesis of the infarcted region. As expected, these effects were more significant in the EGFP-hRAMP1-MSCs group.
Effects of transplanting EGFP-hRAMP1-MSCs on myocardial fraction and fibrosis. (a) Twenty-eight days after transplantation (n = 6), TTC staining in the myocardium demonstrated that the normal myocardium displayed a brick-red color while the infarcted tissue was white (upper panel). Quantitative analysis demonstrated that the infarct area of the EGFP-hRAMP1-MSCs group was significantly smaller than that in the EGFP-MSCs and control groups (lower panel). The data are represented as the mean ± SD (n = 6), *P < 0.05. (b) Photomicrographs show the representative myocardial sections stained with HE or Masson’s trichrome. Magnification is 200 × Scale bar = 100 µm. (A color version of this figure is available in the online journal)
Discussion
Here, we present data demonstrating that the transplantation of hRAMP1-overexpressing MSCs into the rabbit models of MI reperfusion and carotid artery injury has more potential than natural MSCs for improving heart function by attenuating the development of myocardial fibrosis and increasing the myogenesis of the infarcted region. Moreover, we demonstrated that modified MSCs can also significantly reduce intima hyperplasia of the injured carotid artery by inhibiting the proliferation of intimal cells including VSMCs.
MSCs have emerged as powerful candidates for cell therapy in cardiac repair after MI. MSCs are known to be present throughout the body and can differentiate into vascular endothelial cells,20,21 cardiomyocytes,22,23 and smooth muscle cells. 24 Interestingly, MSCs not only modulate the neighboring cells via Cx40- and Cx43-mediated intercellular junctions but also through paracrine actions.25,26 Previous studies have demonstrated that transplanted MSCs can improve cardiac function via multiple mechanisms, such as enhanced myocardial regeneration, angiogenesis, and decreased collagen deposition in the myocardium. 27 To improve the MSCs-based cardiomyoplasty approaches for clinical utilization, several strategies including combination therapies involving MSCs, preconditioned MSCs,28–31 and genetic modification methods11–13,32–34 have been reported to gain a better engraftment of the transplanted MSCs and significantly improved cardiac functions. Nevertheless, information about the effects of natural or modified MSCs on the repair of an atherosclerotic artery is limited. MSCs may deteriorate restenosis for the differentiation of MSCs into VSMCs, and our previous data indeed showed the differentiation of the induced MSCs into VSMCs in vitro. Therefore, in this study, we explored the effects of hRAMP1-overexpressing MSCs on animal models of both MI and carotid artery injury.
CGRP is known as a powerful vasorelaxant that exists throughout the cardiovascular system. RAMP1 is a component of the CGRP-receptor subunit and is required for the activation of the CGRP receptor. VSMCs proliferation was strongly inhibited when CGRP receptors were activated in vitro. Therefore, we speculated that VSMCs proliferation of the injured artery would be inhibited when the CGRP receptors were activated by transplanting hRAMP1-overexpressing MSCs. To test this hypothesis, a recombinant adenoviral vector, which has a few advantages including the capacity for infecting many dividing or quiescent mammalian cells, higher transfection efficiency, and lower genetic toxicity, was used in this study. This pAdxsi-EGFP-hRAMP1 vector efficiently infected the MSCs and expressed EGFP-hRAMP1. Importantly, we found that there were certain expression levels of EGFP-hRAMP1 in injured vessels after transplanting EGFP-hRAMP1-MSCs. Seven days after delivery of MSCs, the expressions of EGFP and CD31 were detected in the neointima of injured carotid arteries in both the EGFP-hRAMP1-MSCs group and the EGFP-MSCs group, which indicated that the MSCs have been targeted to the injured carotid artery and improved re-endothelialization probably by differentiating into endothelial cells after cell transplantation. Interestingly, Forte and his colleagues also showed that allogenic bone marrow-derived MSCs effectively reduced surgically induced stenosis by affecting the expression level of inflammation-related genes, decreased IL-1beta (IL-1β) and MCP-1 expression, and increased transforming growth factor (TGF-β) expression in injured rat carotids. 35 Moreover, the paracrine effects of MSCs may also contribute to the process of endothelialization. MSCs produce a variety of cytokines and growth factors, including vascular endothelial growth factor (VEGF), hepatocyte growth factor (HGF), Insulin-like Growth Factor 1 (IGF-1), and TGF-β,36,37 which act on endothelial cells existing around desquamation or remnant cells of the artery injury and promote their proliferation. It is noteworthy that we have no direct evidence to support the notion that overexpressed hRAMP1 protein has the capacity to induce MSCs differentiation into endothelial cells or boosting the paracrine function of MSCs. However, previous studies have suggested that CGRP and its receptor subunit, RAMP1, may play an important role in the growth and proliferation of endothelial cells.38,39
Natural MSCs themselves have few inhibitory effects on the proliferation and migration of VSMCs; instead, they may facilitate cell proliferation by a paracrine mechanism or cell differentiation.40,41 In this study, we provided evidence to support that overexpressing hRAMP1 protein in MSCs likely changed the effect of MSCs on the proliferation and migration of VSMCs. The experimental data showed that among the EGFP-hRAMP1-MSCs, EGFP-MSCs, and control groups, the lowest I/M ratio was found in the injured artery after EGFP-hRAMP1-MSCs transplantation. Furthermore, the expressions of α-SMA (a specific marker of VSMCs) and PCNA in the carotid neointima of the EGFP-hRMAP1-MSCs animals were the lowest among the three groups, indicating that EGFP-hRAMP1-MSCs more strongly inhibited VSMCs proliferation than natural MSCs and thus resulted in a more significant reduction of the neointima area and the area ratio of I/M. Interestingly, similar results were also found in the study by Deng et al. 15 These investigators demonstrated that bone marrow stromal cells transfected with an adenovirus containing CGRP had the potential to inhibit VSMC proliferation in vitro by secreting CGRP. Moreover, transferring the RAMP1 gene to the VSMCs may have activated the CGRP receptor and therefore increased the effectiveness of the CGRP-induced decrease in restenosis via cAMP-dependent pathways after aortic angioplasty. 42 Although the distinct mechanisms, such as how the overexpressing hRAMP1 in MSCs acts on the VSMCs of the neointima of the injured artery, remain unknown, the above information strongly suggests that overexpressing hRAMP1 in MSCs can significantly inhibit the proliferation and migration of VSMCs into the neointima of the injured artery by activating the CGRP receptor.
Many studies have shown that natural MSCs or genetically engineered MSCs can improve heart function by regenerating myocardial cells and blood vessels and by decreasing myocardial fibrosis after MI.43,44 While MSCs privilege found in animal experiments cannot be tested randomly for there are many obstacles to overcome before its clinic use 45 such as the optimal route for delivery, appropriate dose for efficient engraftment, and the best preconditioning or genetic modification of MSCs. In this study, we demonstrated that a single intravenous injection of 100 million EGFP-hRAMP1-MSCs into the carotid angioplasty and MI rabbit model also could significantly improve heart function by reducing the infarct size and the formation of fiber scar tissue. The intravenous injection of MSCs not only was an efficient noninvasive route for MSCs delivery but also allowed the MSCs to be targeted to both the infarcted myocardium and injured artery. After engraftment in the infarcted myocardium, the MSCs should proliferate and replace the dying cells, improve infarcted myocardium perfusion, and reduce fibrosis. Importantly, the paracrine effect or trophic effect of MSCs played a favorable role in cardiovascular repair by releasing a plentitude of angiogenic, antiapoptotic, and mitogenic factors. 46 Our previous data showed that MSCs modified with human RMAP1 gene distinctly promote re-endothelialization of injured carotid artery and lessen the proliferation of neointima and subsequently make for vascular impair in the model of sacculus-injured carotid rabbit, which CGRP and its receptors have the accelerating effect of endothelial regeneration. Toda et al. reported that CGRP involved in the vascular development and tumor grow by the way of nerve-dependent mechanism. 39 Recently, two studies showed38,47 that CGRP may enhance the vasculogenesis and angiogenesis by the activation of activated protein kinase -endothelial nitric oxide synthase (AMPK-eNOS) pathway or the expression increase of VEGF, TGF-β, and basic fibroblast growth factor (bFGF) in lower limb ischemia tissues in the model. RAMP1 modified MSCs compared with MSCs have more significant effect of improvement of heart function than angiogenesis, which may be related to the enhanced angiogenesis of CGRP induced by exogenous RAMP1.
From the clinical point of view, the rabbit model of MI perfusion and carotid artery injury in the present study is the best pseudotype to the human patients who suffered from acute MI and received complete PCI. Takaoka and his colleagues 48 also reported a similar double-injury model, showing that conditions after acute MI could aggravate postangioplasty restenosis by stimulating signaling through TNF-α, TNFR1, and IL-6. There are several possible causes for the differences between the results of both studies, including the following: (1) the difference between the injured carotid artery in our study and the femoral in his report; and (2) natural or modified MSCs were used for allogeneic transplantation in our research and no engraftment in his models. The MSCs, whether natural or modified, have a great potential in cardiac repair by many mechanisms discussed earlier. However, our research requires obtaining more direct evidence using the larger animal models.
Conclusively, in our study, the rabbit models for both MI and carotid artery injury were successfully created and transplanted with EGFP-hRAMP1-MSCs or EGFP-MSCs, and we found that MSCs overexpressing the hRAMP1 gene greatly improved both infarcted myocardium and injured carotid artery recovery, which would provide useful information for the development of clinical cell-based therapeutic approaches for the treatment of MI and artery injury.
Footnotes
Author contributions
BS and XL contributed equally to this study. BS designed the study and critically revised the paper. XL performed the research, analyzed the data, and wrote the paper. RZ conducted statistical analyses and revised the paper. ZL performed the research and collected the data. DW performed the research and analyzed the data. GX designed the study and revised the paper. All authors approved the final version of the paper.
ACKNOWLEDGEMENTS
We thank Dr Andrew F Russo for providing the pOTB7-hRAMP1 vector. This work was supported by the Natural Science Foundation of China (NSFC30860100) and the International Cooperative Project of Guizhou Province (Qiankehewai G [2010]0732).