
Abstract
Soy contains many bioactive molecules known to elicit anticancer effects. One such peptide, Lunasin, has been shown to selectively act on newly transformed cells while having no cytotoxic effect on non-tumorigenic or established cancer cell lines. While this effect on in vitro systems is promising, Lunasin’s efficacy in an in vivo system is yet to be assessed. In this review, we discuss the state of knowledge with respect to Lunasin and then review some of the powerful genetic tools available in Drosophila. The availability of a sophisticated genetic tool box makes Drosophila an excellent genetic model well suited to studying the biology of Lunasin and its effect on tumor progression in an in vivo model organism.
Keywords
Introduction
History of soy
Soy (Glycine max) has been a staple in the diets of many cultures for millennia. Originally cultivated in Asia, soy arrived in Europe in the 1700s CE and by the 1800s was being grown in the United States. There are many health benefits associated with soy rich diets. Soy consumption lowers bad cholesterol, LDL,1,2 and reduces risk of coronary disease and cancers, particularly breast, colon, and prostate. 3 Soybeans contain many bioactive compounds with demonstrated anticancer activity including isoflavones, protease inhibitors (most noted is the Bowman-Birk protease inhibitor, BBI), saponins, and more recently the peptide Lunasin. 4
Lunasin was discovered by Odani et al. while isolating the 2S cDNA which codes for a nutrient enhancing, methionine-rich albumin protein in soy.
5
The isolated gene, Gm2S-1, codes for the methionine-rich protein, and three additional peptides: a signal peptide, a linker peptide, and Lunasin (Figure 1) all of which are produced as a proprotein and then post-translationally processed.
6
Lunasin has also been found in wheat,
7
barley,
8
amaranth,
9
pepper,
10
rye,
11
oats,
12
and the herb Solanum nigrum.
13
As Lunasin has been isolated from a variety of angiosperm seeds, it is possible that a more thorough screening of plant seeds could reveal Lunasin from many additional sources. However, to date, soy remains the richest source of Lunasin.
Amino acid sequence of isolated Gm2S-1 from Glycine max. Various regions of the protein are color coded and amino acid numbers are provided in parenthesis. Gm2S-1 codes for a methionine rich albumin protein. The protein is made up of a signal peptide (amino acids 1–21), Lunasin (amino acids 22–64), a linker peptide (amino acids 65–81), and Alisin (amino acids 82–158). (A color version of this figure is available in the online journal.)
Structure of Lunasin
Lunasin is a 43 amino acid peptide, which is organized into four motifs; an N-terminal region of unknown function followed by a predicted helical region, a carboxyl terminal RGD cell adhesion motif, and a run of eight aspartic acid residues (poly-D tail) (Figure 2).
14
The helical region is homologous to chromatin binding proteins. The N-terminus, predicted helix, and the poly-D tail are essential for the bioactivity of Lunasin; however, there is conflicting evidence as to whether the RGD motif is essential for the anticancer effects (Table 1).14,15 The poly-D tail binds to deacetylated histones. Galvez and de Lumen found that the poly-D tail and RGD motif are necessary for the antimitotic effect of Lunasin. This was demonstrated in both murine hepatoma cells and Escherichia coli (E. coli) cells.
14
Transfection of murine hepatoma cells with full-length lunasin resulted in a diffused chromosomal mass, indicative of mitotic disruption, whereas transfection with a poly-D deletion mutant resulted in a loss of antimitotic activity (i.e. chromosomal DNA appeared normal and there were no morphological changes). This poly-D deletion also disrupts the RGD motif and so it is not possible to distinguish whether the loss of antimitotic activity is a result of poly-D deletion or RGD disruption. Moreover, E. coli cells expressing full-length lunasin exhibited reduced bacterial growth and abnormal filament formation. On the other hand, E. coli cells expressing a lunasin deletion mutant (resulting in disruption of poly-D and RGD motifs) exhibited normal septa formation and normal division,
14
supporting the claim that the poly-D tail and RGD motif are essential for the bioactivity of Lunasin.
Domain organization of Lunasin. Four distinct regions of Lunasin are shown; a domain of unknown function spanning amino acids 1–22, a predicted chromatin binding helical domain encompassing amino acids 23–32, an RGD cell adhesion motif covering amino acids 33–35 which allows for the internalization of Lunasin into cells, and a poly-D tail made up of amino acids 36–43 which binds to deacetylated histones. (A color version of this figure is available in the online journal.) Regions of Lunasin required for bioactivity Underlined amino acids represent the helical region, the RGD domain is in bold lettering, Poly-D tail is italicized and (−) represents deleted amino acid.

The region of unknown function at the N-terminus is also necessary for the full bioactivity of Lunasin. Lam et al. demonstrated this by creating a deletion mutant that resulted in the deletion of the N-terminus domain. This truncated protein was only about one-third as effective as full-length Lunasin at preventing foci formation in mouse embryonic fibroblasts (NIH 3T3). 15
The RGD cell adhesion motif is thought to be responsible for the internalization of Lunasin. Galvez et al. reported that an RGD-deletion mutant failed to prevent chemically induced transformation of mouse embryonic fibroblasts (C3H 10T1/2). The lack of bioactivity was due to an inability of Lunasin to be internalized by cells and thus it was concluded that the RGD motif is necessary for internalization of Lunasin. 16 In a later study by Lam et al., Lunasin lacking the RGD motif was only slightly less effective at preventing transformation of mouse embryonic fibroblasts (NIH3T3) compared with full length Lunasin. Additionally, they reported that RGD-deletion mutants are internalized into the nucleus as effectively as full-length Lunasin, demonstrating that the RGD motif is not required for internalization. 15 Lam et al. propose that this discrepancy in the data is due to differences in internalization mechanisms between cell lines (NIH3T3 and C3H 10T1/2).
Natural role in soy
Lunasin is found in abundance in developing soy cotyledons. There are three stages in the development of the cotyledon: (1) a phase of rapid cell division followed by (2) a cessation of mitosis, during which time cells grow larger and acquire and store nutrients, and (3) dehydration of the seed. The expression of Lunasin coincides with mitotic arrest. It is thought that Lunasin may play a role in triggering the arrest of cell division because of its demonstrated antimitotic activity in vitro. 17
Effects in tumorigenic and non-tumorigenic lines
Lunasin exhibits no cytotoxic or antimitotic effects on non-tumorigenic or established cancer cell lines.15,16,18,19 When treated or transfected with Lunasin, these cells show normal growth and cell division. In contrast, in cells newly transformed by chemical carcinogens or viral oncogenes, Lunasin has been shown to suppress transformation, arrest cell division, and induce chromosomal fragmentation and apoptosis.15,16,18,19
Proposed mechanisms of action
Inhibition of core histone acetylation
Under non-mitotic, steady-state conditions, core histones are typically in a repressed, deacetylated state. During a transformation event the histones are acetylated, which relaxes the chromatin, and results in the initiation of transcription. 16 Interruption of acetylation is perceived by the cell as abnormal, and in response the cell commits itself to apoptosis. Lunasin competes with histone acetyl-transferases (HATs) to bind the histones that are exposed during transformation events. 16 If HAT binds, transcription is initiated, S phase proteins are produced (like DNA polymerase α and proteins of the origin recognition complex), and mitosis progresses. However, binding of Lunasin to exposed, deacetylated histones disrupts the acetylation process. This results in commitment of the newly transformed cells to apoptosis, thereby preventing the replication of neoplastic cells. 20 In this way, de Lumen et al. liken Lunasin to a “watchdog”, whereby Lunasin sits ideally in the nucleus waiting for a transformation event to occur before jumping to action, inhibiting the replication of newly transformed cells. 20
Inhibition of mitosis by increasing expression of CDK-inhibitors, p21 and p27
Cyclin-dependent kinases (CDKs) are important in controlling cell cycle progression.
21
Deregulation of the cell cycle is attributed to low levels of CDK inhibitors, such as p21 and p27, and is a hallmark of transformed cells.
22
Additionally, low levels of p27 are correlated with a poor prognosis in breast cancer patients.
23
Lunasin has been shown to increase the expression of p21 and p27 (Figure 3), resulting in an arrest at the G2/M stage in transformed colon cancer cells (KM12L4) and mouse embryonic fibroblasts (NIH3T3).15,18
Known modes of action of Lunasin. Lunasin brings about its anticancer effects in a variety of ways. The effect of Lunasin on cancer cells is shown in white lettering within the circles. The mechanism through which Lunasin exerts a specific effect is denoted in yellow. (A color version of this figure is available in the online journal.)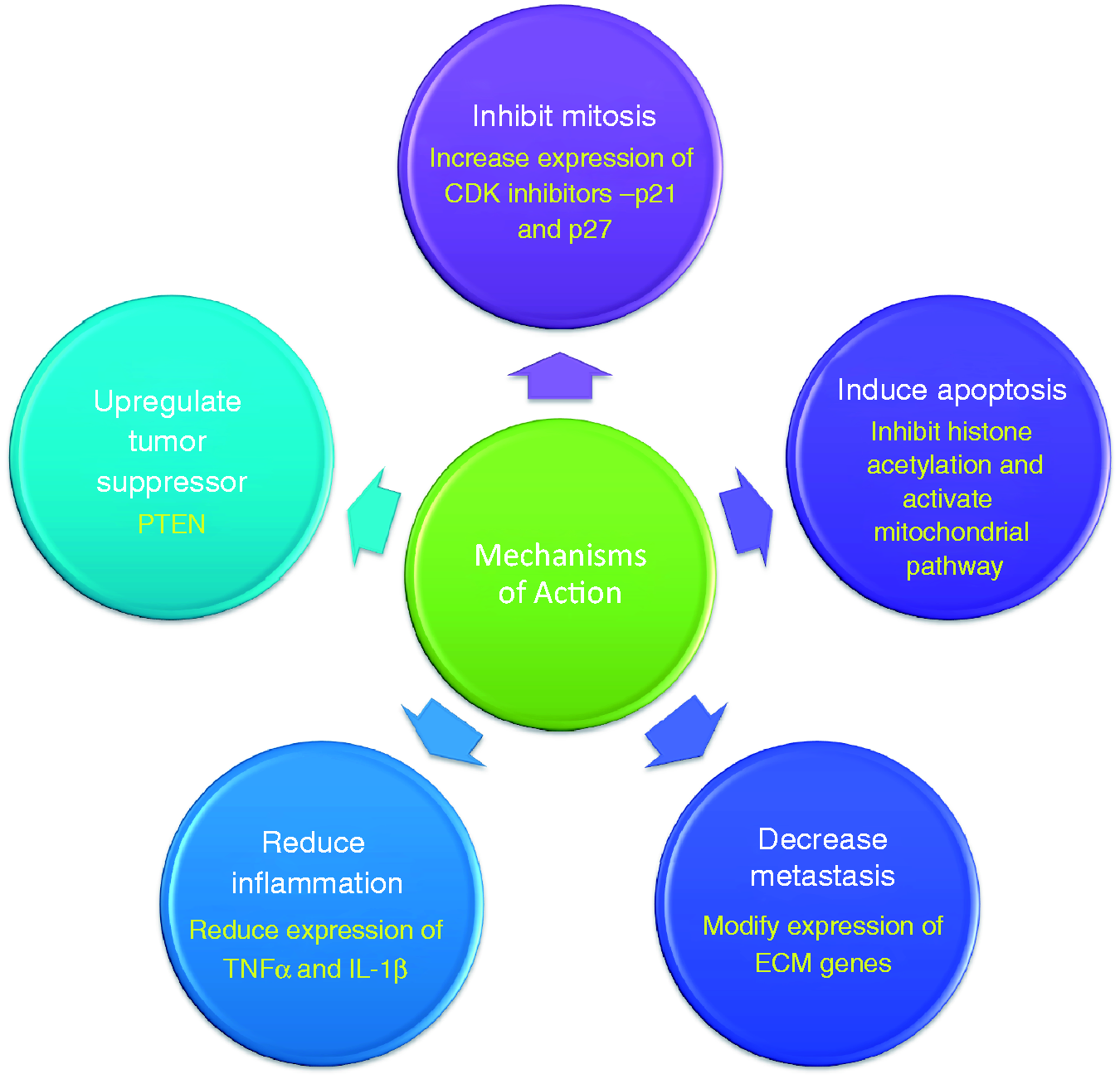
Additionally, earlier work has shown that a reduction in p21 and p27 gene expression correlated with an increase in drug resistance.24,25 It is possible that the increased levels of these CDK inhibitors, as induced by Lunasin, will reduce the prevalence and slow the rise of drug resistance in patients.
Decreased metastasis via modification of ECM gene expression
Effects of Lunasin on gene expression in human KM12L4 colon cancer (adapted from (Dia and Gonzalez de Mejia 18 )
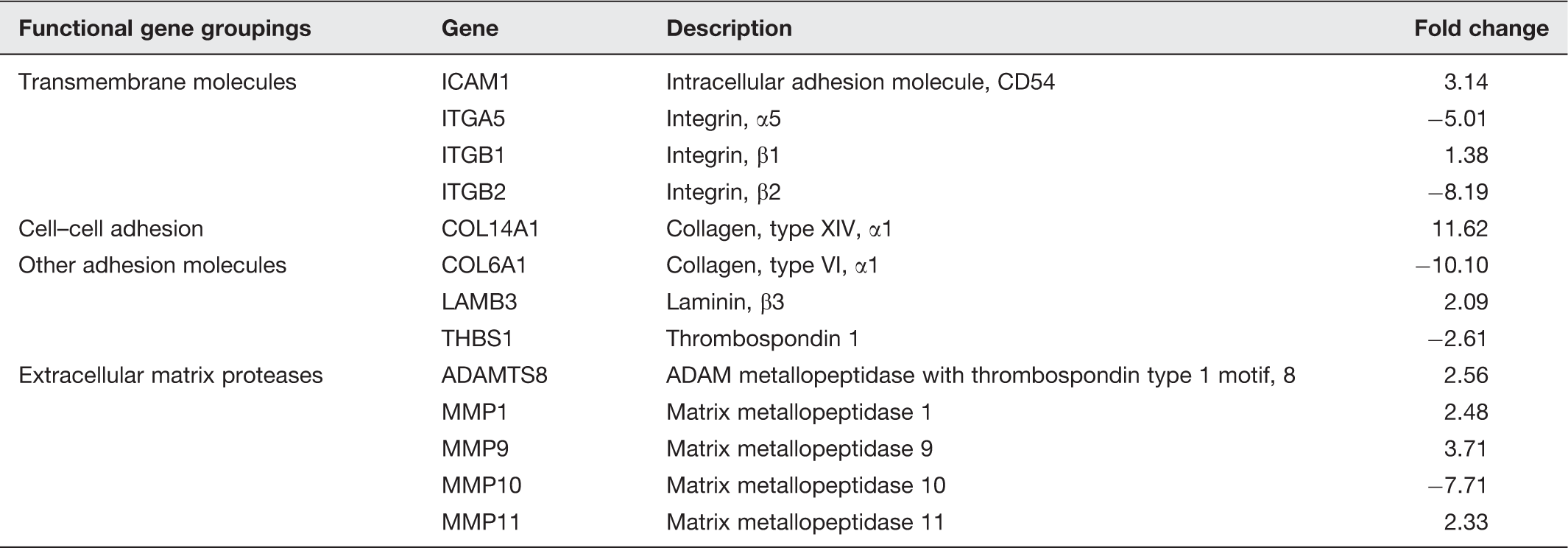
α5β1 integrin plays a key role in cell adhesion, growth, and proliferation. 27 Elevated levels of α5β1 are associated with increased invasiveness of colo-rectal cancer (CRC) cell lines 28 and loss of α5β1 function results in decreased metastasis in CRC cells. 29 Lunasin is able to downregulate the expression of α5 integrin and has the most potent antimitotic effect on cell lines which highly express the α5β1 integrin, 18 suggesting a role as an anti-metastatic molecule.
Induction of apoptosis by activating the mitochondrial apoptosis pathway
Mitochondrial-mediated apoptosis can be triggered by a change in membrane permeability, which is regulated by proteins in the Bcl-2 super family and cytochrome c. 30 Lunasin has been found to activate the mitochondrial apoptosis pathway in colon cancer cells (KM12L4) by increasing levels of pro-apoptotic molecules such as Bax, nuclear clusterin, and caspases 2, 3, and 9, while reducing the expression of Bcl-2, a pro-survival molecule. 18
Reduction of inflammation by inhibition of proinflammatory cytokines
Chronic inflammation has long been implicated as an initiating factor in many cancers (colon, liver, bladder, and stomach). 31 During times of inflammation, leukocytes induce DNA damage through the production of genotoxins including reactive oxygen species (ROS) and nitric oxide (NO). Recently, it has been suggested that Lunasin may help reduce inflammation by inhibiting the release of NO and pro-inflammatory cytokines like TNF-α and IL-1β. 3 The discovery of the anti-inflammatory effects attributed to Lunasin may be significant in the search for therapeutics for other ailments directly caused or amplified by inflammation including arthritis, heart disease, diabetes, and Alzheimer’s disease.
Upregulation of a tumor suppressor, PTEN
PTEN, a phosphatase and tensin homolog, is a tumor suppressor that plays a regulatory role in cell cycle progression and apoptosis, and is often found mutated or downregulated in cancers.32,33 Genistein, a soy isoflavone, has been found to impart antitumor effects via activation of PTEN pathways,34,35 leading researchers to wonder if Lunasin shares this antitumor mechanism. Pabona et al. found that human breast cancer cells (MCF-7) treated with Lunasin exhibited elevated levels of PTEN expression and nuclear localization resulting in the induction of apoptosis via the PTEN pathway. 19
Most of the studies reviewed in this article have utilized in vitro systems to study the effects of Lunasin, with no studies addressing the effects of in vivo expression of lunasin on tumor growth and metastasis in an intact organism. In order for Lunasin to progress to advanced stages of drug development, it is imperative that the in vivo effects of Lunasin expression be assessed in a living organism. Drosophila provides a powerful, economical choice for a model organism in which to test this. Here we present reasons why Drosophila is a good choice for studying the in vivo effects of Lunasin and provide an experimental strategy for how this can be achieved in Drosophila.
Drosophila as a model organism
Since the time of Thomas Hunt Morgan, Drosophila has been an important model organism. Morgan originally chose the fruit fly because of the ease of collection, fast generation time, and small size. Since his time, much information has been collected that leads researchers to continue to choose Drosophila as a model organism.
The power of Drosophila as a model lies in the simplicity of conducting genetic screens made possible by the ease and affordability of rearing multiple generations in a short period of time. Drosophila has an 8–10 day life cycle at 25℃ with females laying between 500 and 1000 eggs per animal. 36 Vast quantities of eggs allow researchers to analyse hundreds or even thousands of mutant lines over multiple generations, a feat matched only by bacteria and yeast. 37
The Drosophila genome is organized into four chromosomes, containing approximately 13,600 genes 38 ; a relatively low number as compared to the estimated 20–30,000 genes in humans. 39 Furthermore, Drosophila has relatively little genetic redundancy as compared to vertebrate models 37 allowing for easier genetic mapping and balancing (discussed below). Additionally, the low number of orthologs present in flies for human genes presents a unique opportunity and an ease in understanding the gene function in an intact organism. For example, the mammalian genome contains more than 20 MMPs, whereas flies have only two MMPs. 40 To study the effect of total MMP knockdown it is easier to knock down the two MMPs in Drosophila than to mutate or knockdown the more than 20 MMPs in mammals.
Perhaps the most important aspect of a model organism is the ability to translate information gained to other organisms. As many processes and biochemical pathways are conserved from flies to humans, 37 Drosophila provides an informative model with which to study human disease states. About 80% of human genes have homologs in Drosophila and about the same percentage of identified human disease genes have Drosophila cognates (75%).41,42
Availability of a sophisticated tool box in Drosophila
Selected genetic tools available in Drosophila.

Balancer chromosomes are chromosomes that contain multiple inversions capable of suppressing crossover events between homologous chromosomes during meiosis. These chromosomes are marked by dominant, homozygous lethal mutations (e.g. Tb, Sb, ScO, etc.) and allow investigators to (1) maintain recessive lethal mutations in a population and (2) to know the genotype of individuals without having to genotype each animal. 48 Balancer chromosomes are available for the X chromosome, as well as all three Drosophila autosomes. 49
Another important advance in fly biology happened with the identification and engineering of P transposable elements (P-element). The re-engineered P-element and its incorporation into DNA vectors like pCASPER made it possible for researchers to create transgenics and insertional mutations for and in various genes, respectively. 50 Furthermore, the P-element also facilitated the development of enhancer traps and other genetic tools used by the fly community today. 51
The GAL4/UAS system is one such tool in which the UAS and GAL4 elements are derived from yeast and, in combination with the P-element, results in a powerful gene expression technique. This technique is routinely utilized to achieve targeted overexpression of a gene of interest.
52
To achieve this targeted overexpression, the gene is placed under the control of a upstream activation sequence (UAS) enhancer element which is responsive to the transcription factor GAL4, and drives expression of downstream genes (Figure 4). GAL4 expression can be spatially restricted by placing GAL4 under the control of a tissue specific enhancer (e.g. the glass multiple reporter enhancer in GMR-GAL4 drives GAL4 expression to the eye). By forcing the ectopic expression of a gene, it is possible to study the biology of an otherwise lethal gene in vivo
53
in various tissue contexts. A number of GAL4 driver lines are available from the Bloomington Drosophila Stock Center (http://flystocks.bio.indiana.edu/) that allow for the expression of GAL4 in specific tissues..
The Drosophila GAL4/UAS binary overexpression system for targeted gene expression. Expression of a gene of interest (in this case green fluorescent protein, GFP) in a targeted manner is brought about by crossing flies bearing a UAS-GFP transgene with flies containing a GAL4 driver under the control of an enhancer element (in this example the GMR enhancer). Progeny flies express GAL4 only where the driver, GMR enhancer, is active in the developing eye. GAL4 will bind to UAS and force the expression of the target gene, GFP, resulting in Drosophila with green fluorescing eyes. (A color version of this figure is available in the online journal.)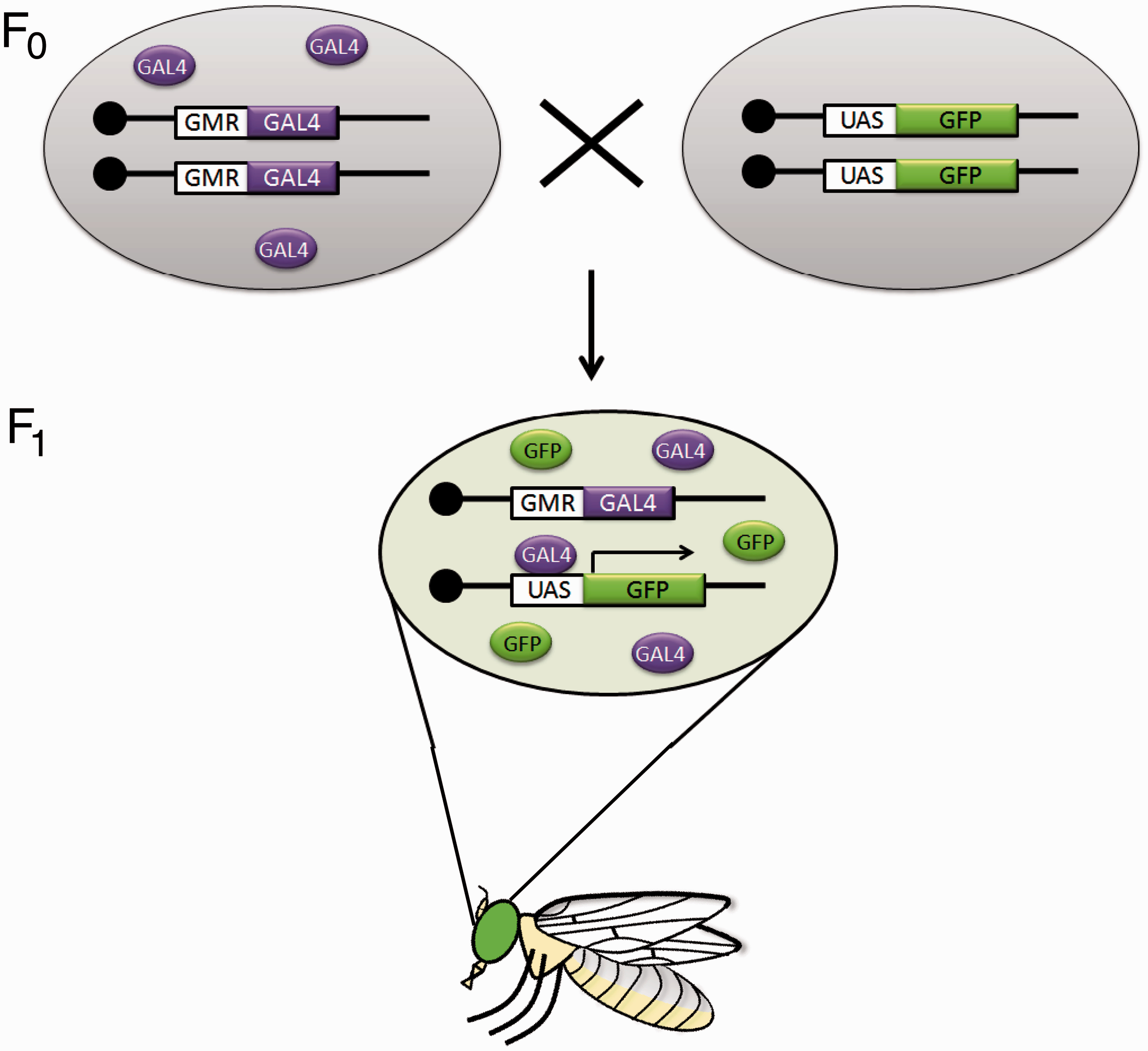
Genetic recombination between homologous chromosomes is typically not found in somatic cells but is restricted to meiosis. Thus a recessive lethal mutation cannot be studied in its homozygous state in an individual fly as this would lead to lethality. The flippase/flippase recombination target (FLP/FRT) technique circumvents this problem and allows for mitotic recombination between homologous chromosomes in Drosophila. The resulting products of mitosis consist of clones of cells bearing the mutation in a heterozygous state, clones of cells bearing the mutation in a homozygous state and clones of cells homozygous for the wild type allele. The creation of clones that carry the mutation in the homozygous state allows for its study without outright killing the individual fly. By placing this recombination system under the control of a heat shock promoter or other tissue specific enhancers, we can temporally and spatially control mitotic recombination 45 and clone generation in the fly.
Another powerful technique emerged when Liqun Luo’s group at Stanford combined the GAL4/UAS system with the FLP/FRT technique to develop the MARCM system short for ‘mosaic analysis with a repressible marker’. 47 MARCM allowed researchers to create discrete, fluorescently labeled clones that are homozygous for a target gene, in an otherwise heterozygous animal. 54 Due to the presence of the GAL4/UAS system in MARCM, one can also simultaneously overexpress a gene of interest. These properties of the MARCM system have been utilized to model tumor progression and to study the tumor microenvironment interaction.55,56
Biomedical advances attributed to fruit fly research
Drosophila has been used to model a variety of diseases and genetic disorders associated with aging, 57 neurological diseases such as Alzheimer’s and Parkinson’s,37,58 and cancers. Drosophila thus provides a valuable model to understand disease states in humans and has led to many advances in modern medicine.
One such advancement in medicine is the approval of a therapeutic for the treatment of tuberous sclerosis (TS). In patients with TS, tumors develop in multiple organs. Research on Drosophila elucidated the signaling pathway responsible for the misregulation of tissue growth, and furthermore led to the discovery that reducing the expression of a gene (S6 kinase) prevents this misregualtion. 59 Rapamycin is a drug known to inhibit a portion of the S6 kinase pathway, and was approved by the FDA for clinical trials in TS patients. In 2008, Bissler et al. published findings that a 12-month regimen of rapamycin resulted in a decrease in angiomyolipomas in 50% of patients. 60
Modeling of tumors in Drosophila
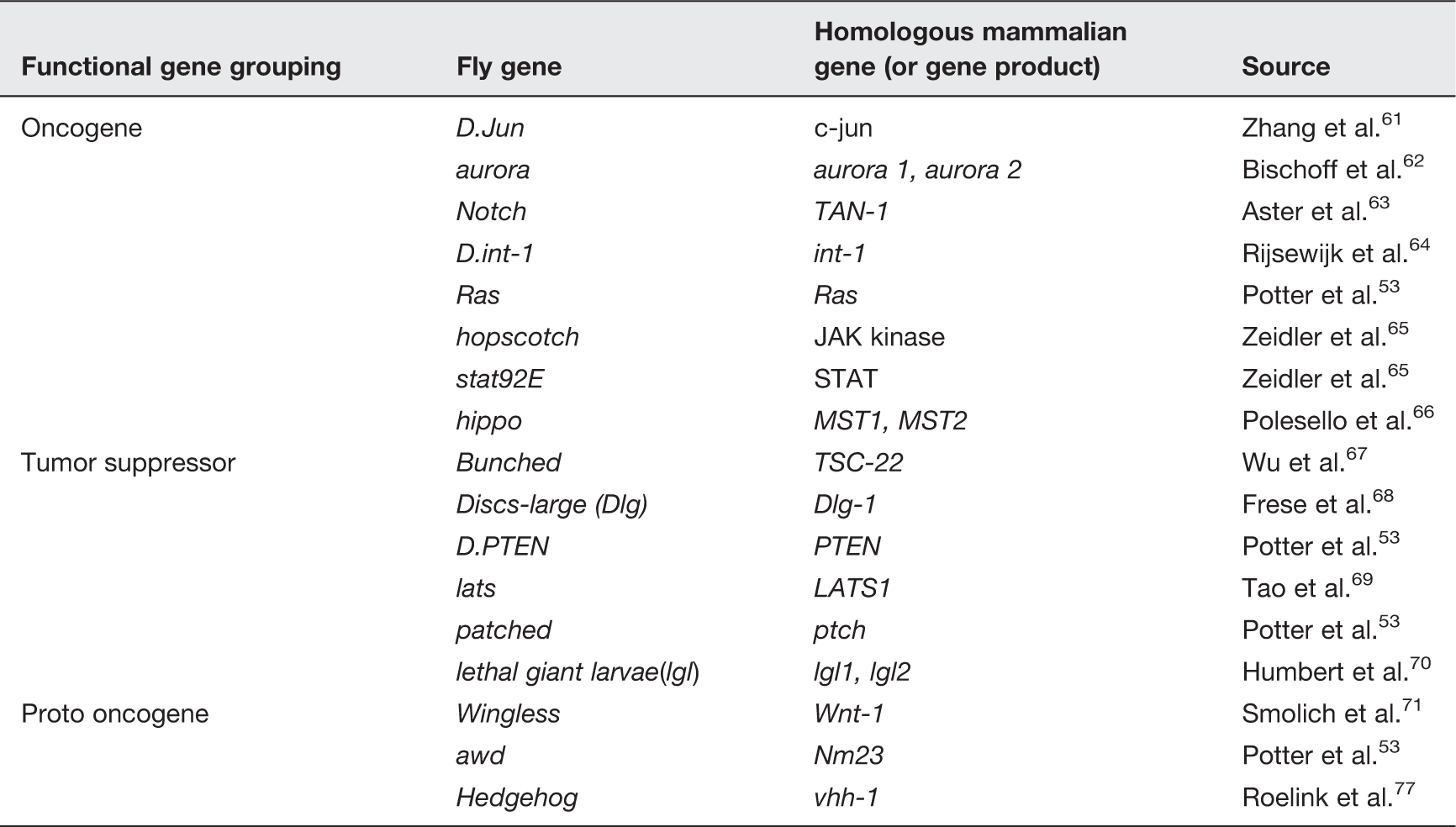
Mammalian homologs to Drosophila genes related to tumorigenesis.
Modeling of metastasis in Drosophila
The majority of cancer fatalities in humans are attributed to secondary metastatic growth. 74 As many biochemical mechanisms and pathways associated with metastasis are conserved between humans and flies (e.g. ECM turnover), 53 Drosophila provides a useful model for studying metastasis.
Features of tumor metastasis have been modeled in Drosophila. This modeling was achieved by utilizing the property of cooperation between an activated oncogene and loss of cell polarity function. Cooperation between these two functions results in massive overgrowth and migration of tumors. One such modeling makes use of the RasV12/lgl−/− genotype.56,75 Ras is an oncogenic protein and RasV12 codes for an activated isoform of the protein. Flies overexpressing RasV12 exhibit benign, non-metastatic tumors. However, when RasV12 clones are also mutant for lethal giant larvae, a tumor suppressor encoded by the gene lgl, aggressive metastatic tumors result.56,75
Limitations of Drosophila as a tumor model
Although there are structures in flies that perform similar functions to mammalian organs (e.g. Drosophila Malpighian tubules and mammalian kidneys 76 ), several human organs have no direct homologous organ in flies. This can make modeling of organ specific cancers difficult. Regardless, there is much to be learned about the highly conserved biochemical characteristics of tumors that can be studied by mimicking tumor environments in the fly.
Instead of blood vessels, Drosophila utilizes an open circulatory system. Hemolymph serves as the nutrient transport system and the hemocoel, an open cavity, houses and transports hemolymph. Although true angiogenesis does not occur in Drosophila, in tumors produced by mutations in the lats gene, ‘lumen like’ structures, resembling blood vessels, form and serve to distribute nutrients, 53 mimicking angiogenesis. In this way, it is possible to study angiogenesis in flies as it relates to tumors; however, care must be taken to ensure conclusions are not overstated.
Use of Drosophila to study the effects of Lunasin in vivo
Lunasin is currently a focus of investigation for its role as a potential anticancer therapeutic. Many studies have investigated the effects of Lunasin in vitro, but to date no studies address the effects of in vivo expression of lunasin. Drosophila provides a valuable tool in which to study the effects of Lunasin on tumor growth and progression in vivo. Transgenic Drosophila capable of expressing Lunasin under the control of the GAL4/UAS overexpression system in combination with RasV12 benign or RasV12/lgl−/− genetically induced invasive tumors can afford us the opportunity to assess how Lunasin affects tumor growth and progression in an in vivo model (Figure 5).
Modeling of benign and invasive tumors in Drosophila. (a) Drawing of a third instar larva is shown with an approximate location of the cephalic complex. A magnified view of the cephalic complex is presented on the right and various imaginal tissues are labeled. (b) Third instar larval schematics with genotypes utilized in modeling benign and invasive tumors are shown. Cephalic complexes are shaded gray except where tumor tissues marked green are present. No tumor tissue is present in wild type larva (first from left). Larvae overexpressing an oncogenic form of Ras, RasV12 results in benign overgrowth in the eye-antennal disc as shown (second from left). Larval eye antennal disc cells mutant for lgl result in small clones (third from left). Cells overexpressing RasV12 and simultaneously mutant for lgl result in overproliferation of eye-antennal disc cells (fourth from left). These cells are capable of invading other organs and display hallmarks of metastasis found in humans. The potential of Lunasin as an anticancer agent can be tested in these benign and invasive tumor cells by simultaneous overexpression from a UAS-lunasin transgene (fifth from left). In all panels tumor tissue is marked with GFP. The tumors were induced using the MARCM technique (see text for details). (A color version of this figure is available in the online journal.)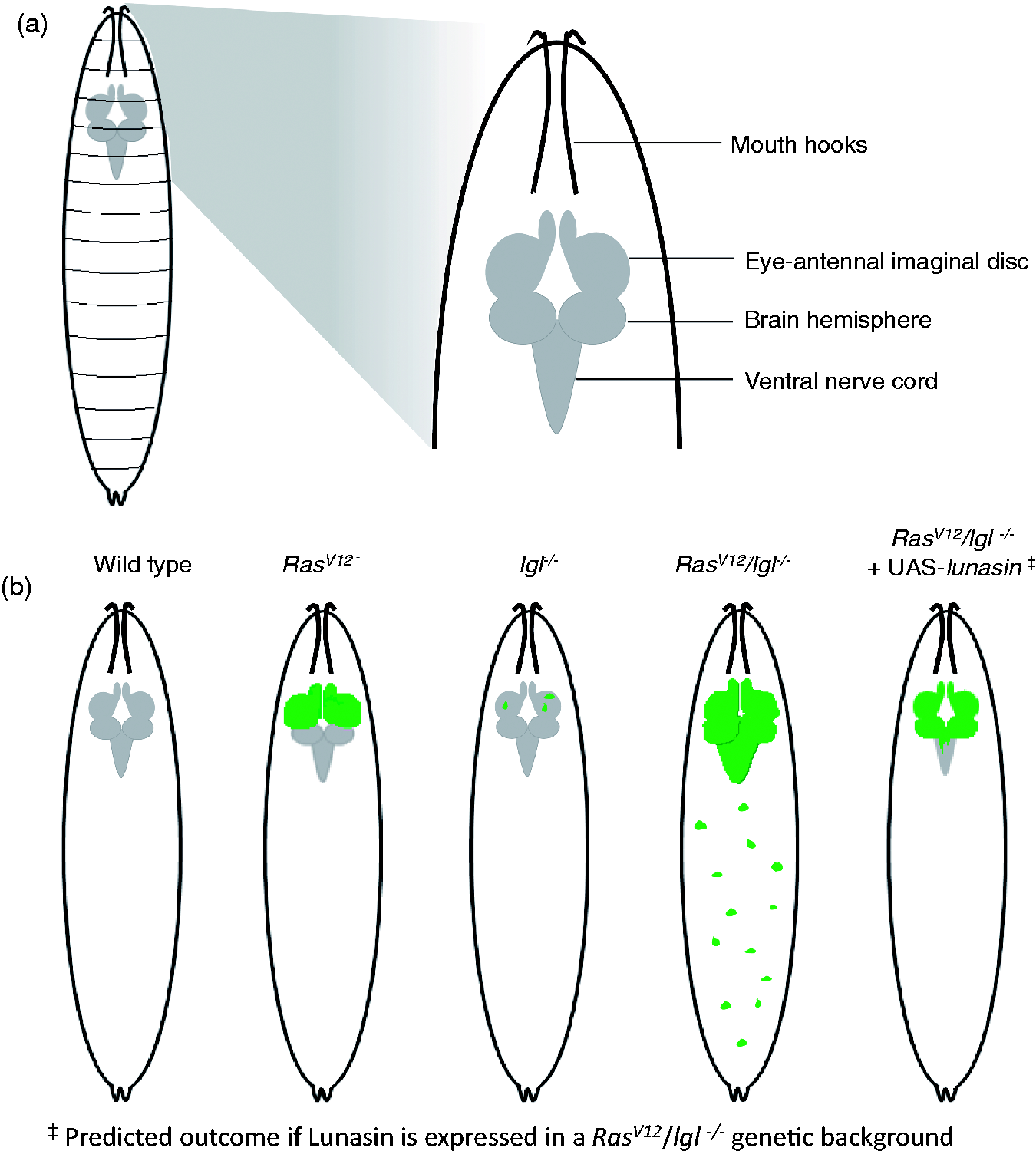
Concluding remarks: Lunasin as a natural chemotherapeutic
Aside from the demonstrated benefits of Lunasin, public opinion may very well play a role in the development of Lunasin as a therapeutic. Our society increasingly places a high value on natural products, as manmade chemicals often have unforeseen side-effects. It therefore makes sense for us to find ways to reduce our dependence on these chemicals. Because of the multitude of anticancer pathways influenced by Lunasin, this short soy bean derived peptide holds promise in combating numerous cancers, including those which have developed drug resistance. The next stage in Lunasin research is to test the efficacy of Lunasin in vivo, and Drosophila provides us a powerful model with which to do so.
Footnotes
Author contributions
AS conceptualized the article, AS and GJ researched and wrote the article.
ACKNOWLEDGEMENTS
The authors would like to thank Cheryl Davis and Rodney King for critically reading the manuscript and helpful suggestions. Part of the research presented in this article was funded through a WKU graduate studies grant to GEJ. Research in the laboratory of AS is supported by the WKU Department of Biology startup funds, WKU Research Foundation RCAP-I grant # 11-8032, and by a KBRIN-AREA grant funded through a parent grant from the National Institute of General Medical Sciences of the National Institutes of Health under award number 5P20GM103436-13.