
Abstract
Freshwater stingray accidents cause intense pain followed by edema, erythema, and necrosis formation. Treatment for stingray envenomation is based on administration of analgesic, antipyretic, and anti-inflammatory drugs. This report evaluated the local inflammatory reaction—including edema formation, leukocyte recruitment, release of inflammatory mediators, and histopathological changes—after the intraplantar injection of Potamotrygon motoro stingray venom in mice. Edema was observed as soon as 15 min after venom injection, peaking at 30 min, and lasted up to 48 h. In addition, P. motoro venom increased neutrophil counts in the site of injection, at all time periods and venom doses analyzed. Increased eosinophil and lymphocyte counts were detected mainly at 24 h. Moreover, monocytes/macrophages were observed in large amounts at 24 and 48 h. Microscopically, the venom induced leukocyte migration to the injured tissue, edema, mast cell degranulation, angiogenesis, and epidermal damage. Inflammatory mediator release (IL-6, MCP-1 and KC) was detected as soon as 1 h after venom injection, and it increased significantly at 4 h. At 24 h, the venom induced only the production of MCP-1. These results show that this stingray venom evokes a complex inflammatory reaction, with rapid and persistent edema formation, leukocyte recruitment, and release of cytokines and chemokines.
Introduction
Rays are elasmobranchs (Chondrichthyes) found worldwide, especially in tropical and temperate seas, and in river waters. These animals are characterized by a flattened disk-shaped body with large pectoral fins used for locomotion. Some rays, known as “stingrays,” possess one or more stings in the base of their tail, with a very sharp tip.1–4 The sting is composed of a retroserrated mineralized structure covered by an integumentary tissue, and toxins are produced by specialized cells located all over the epithelium.5–8 In fact, there is no specific venom-producing gland, and the supernatant of the extract obtained from the macerated tissue that covers the sting is called venom.4,9
Clinical symptoms of stingray envenomation, due to mechanical trauma and the sting toxic components (venom) action, involve an intense pain followed by edema, erythema and, in freshwater stingray accidents, necrosis formation. Systemic symptoms, e.g., fever, myalgia, dizziness, vomit, and headache, are also described.3,10,11 Despite being rare, severe accidents and death are also reported.1,11–16 There is no specific treatment for stingray envenomation so far and therapy consists of analgesic, antipyretic, and anti-inflammatory drugs administration,10,11,14,17 besides the use of antibiotic to prevent secondary infection, that may occur due to bacteria found in the mucus covering the sting as well as in the river water. 18
There are few documented studies on the characterization of stingray venoms. This lack of data is mainly due to the difficulty of capturing the animal to obtain the venom and for its thermolability. However, some studies have shown that the venom of stingrays is quite complex, with components distributed in a wide electrophoretic spectrum.9,19,20 Experimental studies have identified enzymes that may contribute to the local effects of envenomation, which include proteases, hyaluronidases, phospholipases, acid phosphatase, gelatinases, elastases, and proteases.9,21 Freshwater stingray venoms also induce nociception, edema, myotoxic and lethal activities, severe local inflammation, and necrosis in mice.9,19,21,22 Besides, two peptides with biological activities, named orpotrin and porflan, were also identified from a freshwater stingray venom.23,24
The studies with stingray venoms are scarce in the literature and, thereby, the mechanism of action of these venoms is not fully understood, impairing precise treatment for accidents. Since freshwater stingray venom causes an intense and long-lasting local reaction, the objective of this study was to characterize some aspects of the inflammation induced by Potamotrygon motoro venom in mice, evaluating edema formation, leukocyte recruitment, the histology of the injured tissue, as well as the production of inflammatory mediators at the injured site.
Materials and methods
Animals and venom
Swiss mice (males, 18–20 g) were provided by the Institute Butantan animal house and animals received fresh water and food ad libitum. Stings of specimens of P. motoro (identified according to Rosa 25 ) were collected in Paraná river (Mato Grosso do Sul State), and transferred immediately to laboratory. Tissue extracts were obtained from the integumentary tissue covering the stinger as previously described. 9 The protein content of P. motoro tissue extract pools (named as venom) was determined by bicinchoninic acid method, 26 using bovine serum albumin (BSA) as a standard. The procedures involving animals were conducted according to national laws and policies (protocol numbers: CEUAIB 635/09, CGEN 02001.005111/2008, and SISBIO 15702-3).
Evaluation of paw edema
Mice (n = 6–9) were injected (i.d., 30 µL) with P. motoro venom (2, 8 or 32 µg) or Phosphate Buffered Saline (PBS) (negative control) in the right hind paw. After 15 min, 30 min, 1, 4, 24, and 48 h of injection, edema was measured by plethysmometry (7140 Plethysmometer, Ugo Basile, IT). Results expressed the difference in paw volume (µL) prior to (control) and after (experimental) injection (mean ± SEM).
Leukocyte count in footpad pellets
Mice (n = 5–6) were injected (i.d., 30 µL) with P. motoro venom (2, 8 or 32 µg) or PBS (negative control) in the right hind paw. After 30 min, 1, 4, 24, and 48 h of injection, the right paws were removed at the level of the tibiotarsal joint and tissue was disrupted and homogenized in 2 mL in a solution containing PBS pH 7.4, EDTA 2 mM, NaHCO3 5%, and indomethacin 10 µM. Following centrifugation at 4℃ for 10 min, supernatants were used to assay the release of cytokines and chemokines by enzymatic immunoassay. Cell pellets were recovered in PBS and BSA 0.1% solution to perform cell counts. Total leukocyte counts from footpad homogenates were performed by Trypan blue 0.1% exclusion (Sigma-Aldrich, USA) using a hemocytometer chamber; differential counts were accomplished using cytocentrifuge slides stained with HEMA-3 (Fischer Scientific Company, MI, USA). For differential cell counts, 100 leukocytes were enumerated and identified as monocytes/macrophages, lymphocytes, neutrophils, or eosinophils, based on staining and morphologic features. The results represent the mean ± SEM per milliliter of cell suspension.
Histopathological analysis
Mice (n = 3) were injected (i.d., 30 µL) in the right paw with P. motoro venom (2, 8, or 32 µg) or PBS (control group). Venom-injected group were euthanized at 30 min, 1, 4, 24, and 48 h and PBS-injected mice at 4 and 24 h. The right paws were removed and footpad tissue was fixed in paraformaldehyde 4% in PBS for 24 h. After dehydration in ethanol 95%, tissue was embedded in glycol methacrylate (Leica Microsystems Nussloch GmbH, Heidelgerg, GR) and, after polymerization, sections of 4 µm were obtained in a Microm HM340 microtome and stained with toluidine blue and fucsine and examined under a light microscope. Photomicrographs were obtained in an Olympus B×51 microscope and in an Olympus SZ stereomicroscope, equipped with a digital camera and Image-Pro Express software (Media Cybernetics).
Quantification of cytokines and chemokines
Levels of cytokine (IL-6) and chemokines (KC and MCP-1) were measured in supernatants of paw injected with P. motoro venom tissue extracts (as described above) using commercial specific two-site sandwich enzyme linked immunosorbent assay (ELISA), according to manufacturer's instructions (IL-6 and MCP-1—B&D Biosciences, CA, USA; KC—R&D Systems, MN, USA). As positive control, mice were i.pl. injected (10 µg/paw) with Loxosceles gaucho spider venom (Barbaro et al. 27 ) and paw homogenate supernatants of control groups were obtained 2 h after injection. Samples were quantified using standard curves of recombinant murine cytokines and chemokines. Detection limits were 15.625 pg/mL for all assays. Results were expressed as mean ± SEM for duplicate samples.
Statistical analyses
Results were expressed as means ± SEM. One-way or two-way analysis of variance (ANOVA), followed by Holm-Sidak test, was used to analyze data, employing SigmaStat 3.5 software. Values with P< 0.05 were considered statistically significant.
Results
Edema induced by P. motoro venom in mice footpad
It was verified that P. motoro venom caused edema as soon as 15 min after samples injection, in relation to control group, peaking at 30 min (Figure 1). After this time period, the edema decreased, but it was still detected at 4 h in all venom doses tested. In 24 and 48 h after injection, edema was observed in animals injected with 8 and 32 μg of venom. The following experimental protocols were initiated at 30 min, since the peak of edematogenic activity induced by P. motoro venom occurred in this time period.
Edematogenic activity induced by P. motoro venom. Aliquots of 2, 8, or 32 µg of venom were injected i.d. into the paw of mice (n = 6–9) and edema formation was measured by plethysmometry at 15 min, 30 min, 1, 4, 24, and 48 h time periods. ANOVA test *P< 0.05 in relation to control group (PBS)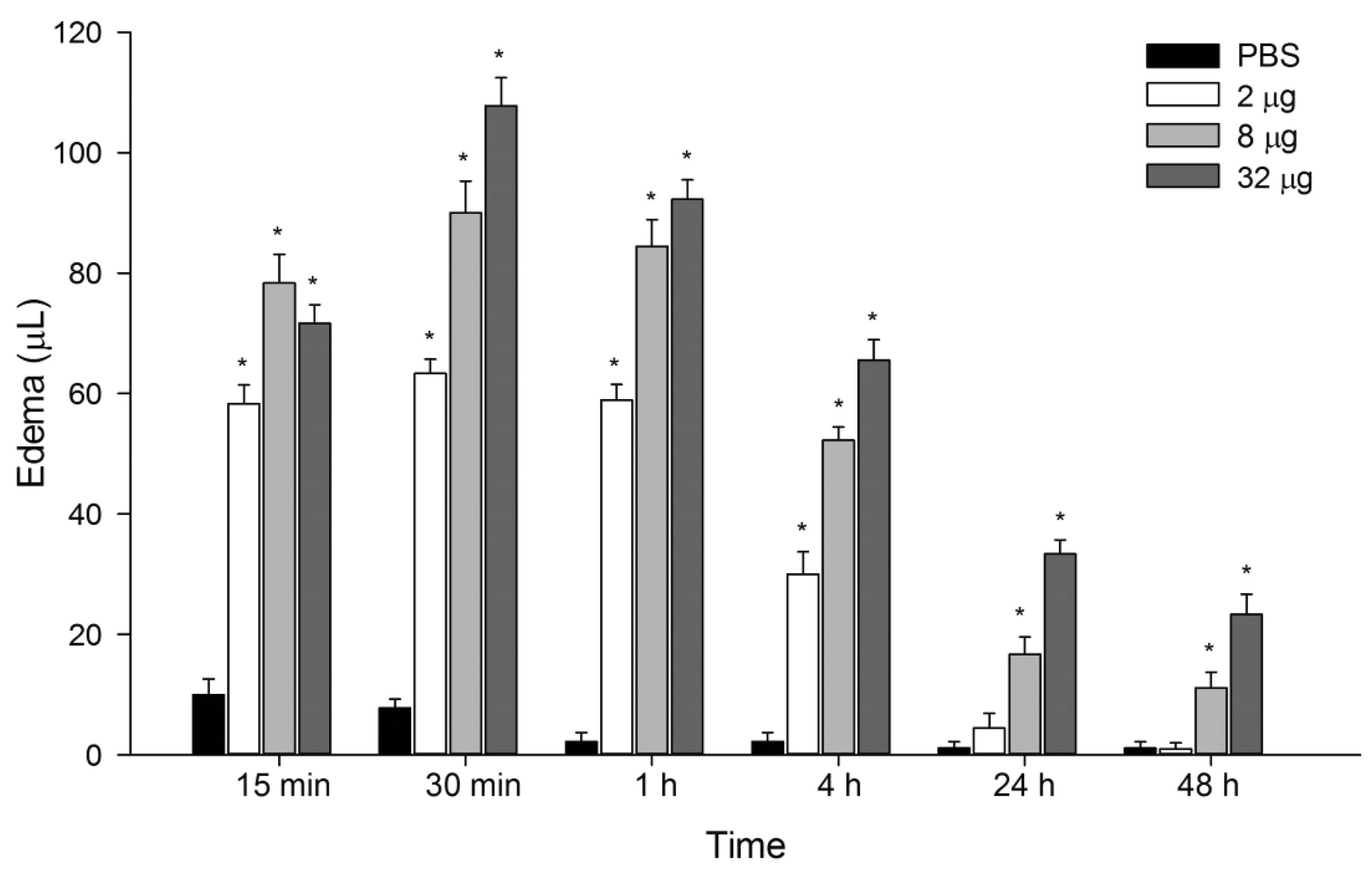
Effect of P. motoro venom in the number of leukocytes on mice footpad
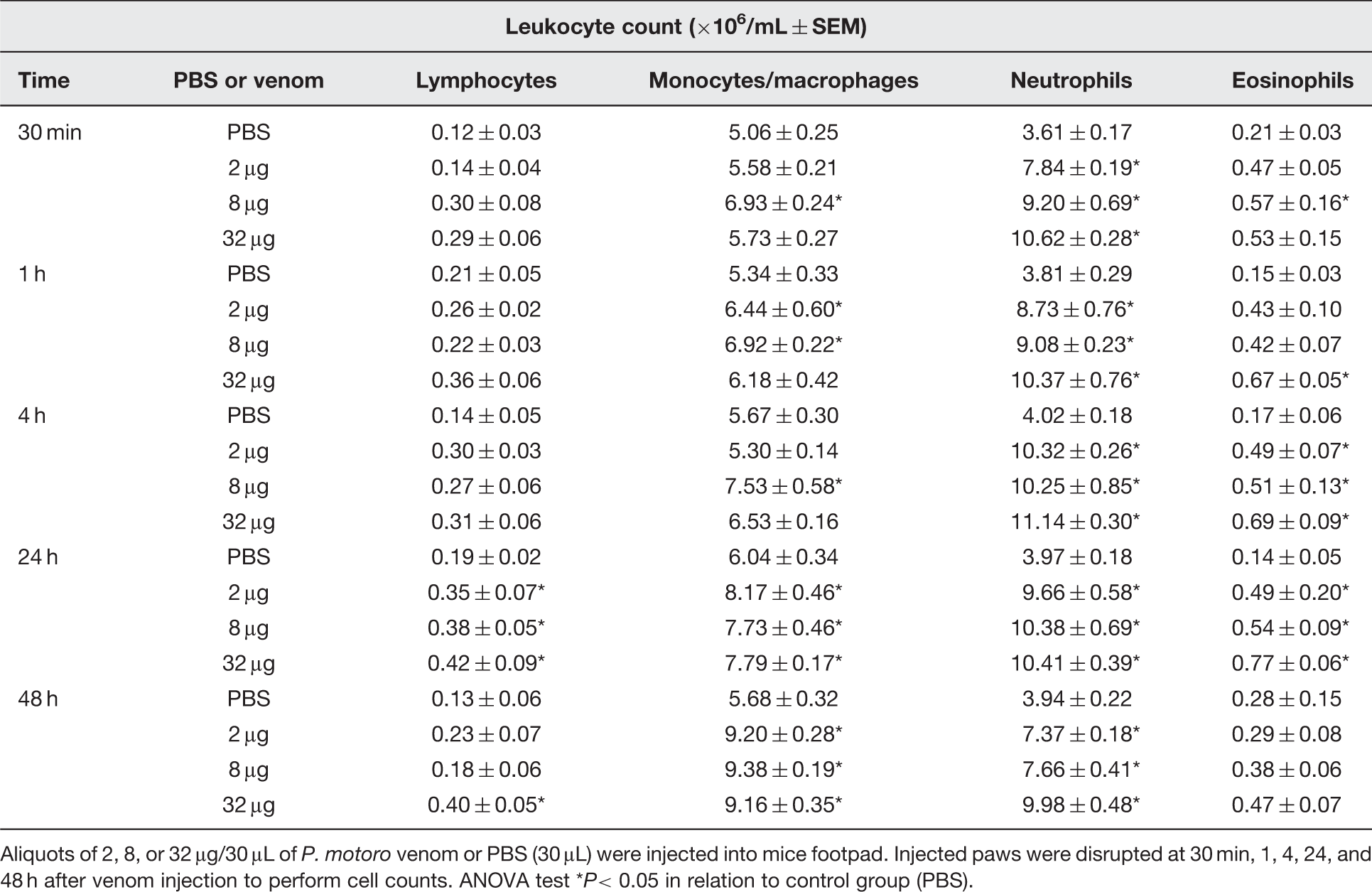
All doses of venom tested increased the number of leukocyte from 30 min up to 48 h after injection, in relation to control group (Figure 2). Differential cell counts showed an augmentation in neutrophil count (about twofold), which remained up to 48 h after injection, induced by all venom doses studied (Table 1). Eosinophil counts increased mainly in 4 and 24 h after injection in all doses of venom tested. In relation to mononuclear cells, it was seen that P. motoro venom quantitatively increased monocyte/macrophages influx to mouse footpad in all doses tested after 24 and 48 h of injection, when compared to PBS-injected group. Besides, an increase in lymphocyte count was detected, especially at 24 h after venom injection.
Effect of P. motoro venom in total leukocyte count at the site of injection. Mice were injected into footpads with P. motoro venom (2, 8, or 32 μg) or PBS (control) (n = 5–6). After 30 min, 1, 4, 24, and 48 h of venom injection, animals were euthanized and tissue footpads were processed for leukocyte counts. ANOVA test *P< 0.05 in relation to control group (PBS) Differential count of leukocytes present in mice footpad after P. motoro venom injection Aliquots of 2, 8, or 32 μg/30 μL of P. motoro venom or PBS (30 μL) were injected into mice footpad. Injected paws were disrupted at 30 min, 1, 4, 24, and 48 h after venom injection to perform cell counts. ANOVA test *P< 0.05 in relation to control group (PBS).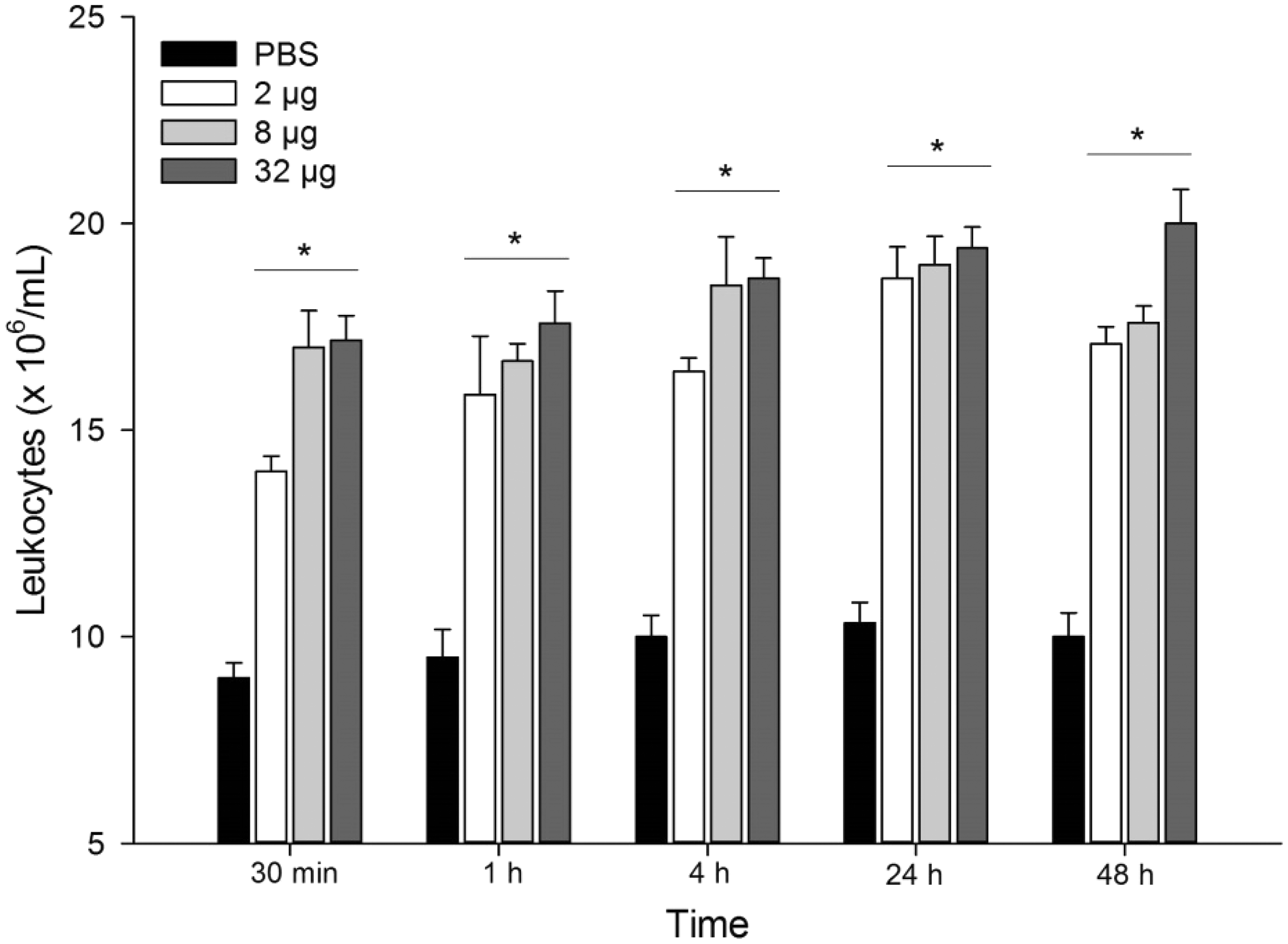
Local histopathological changes induced by P. motoro venom
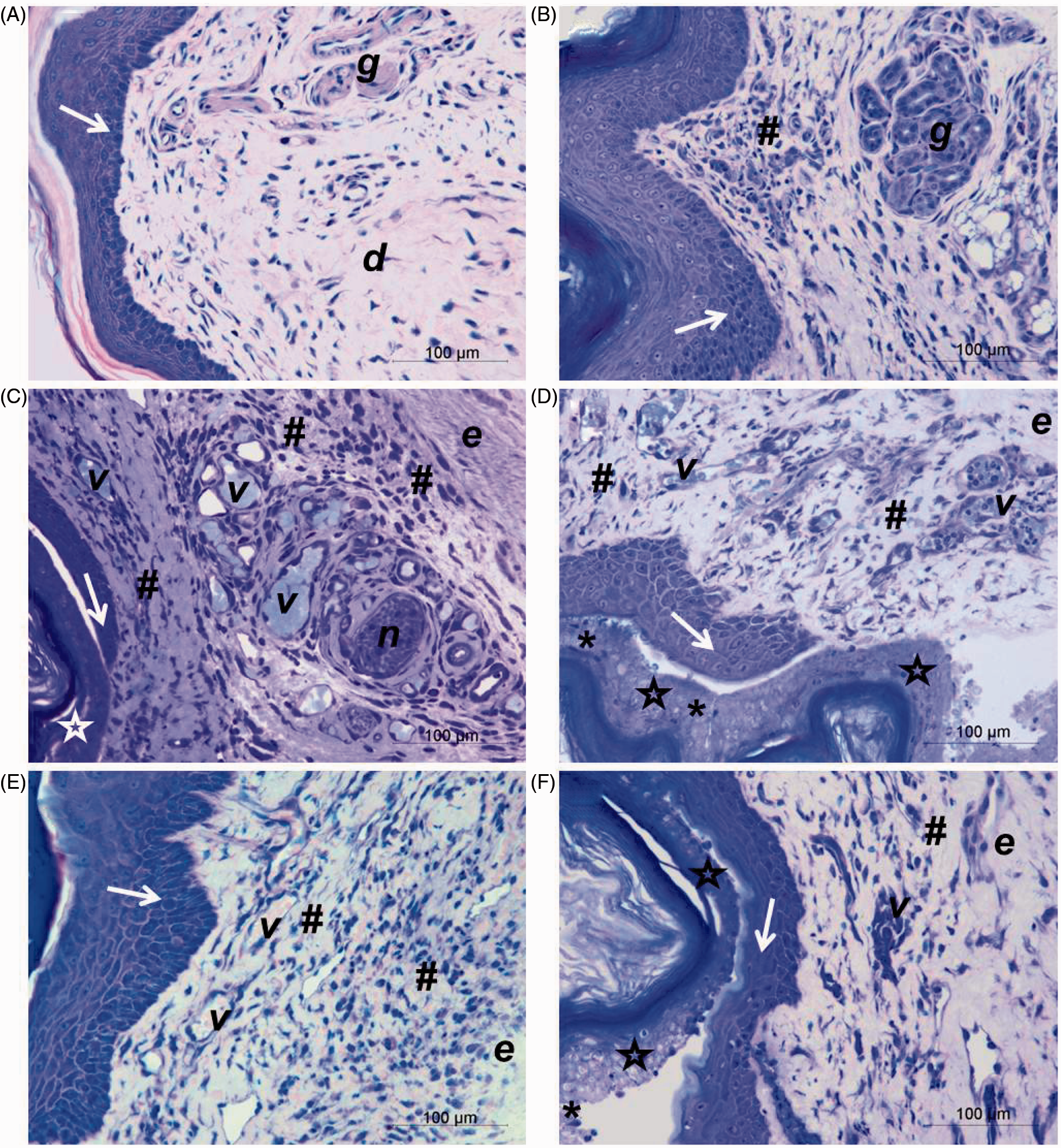
It was observed that connective tissue, at 30 min after venom injection, was already disorganized with a slight presence of polymorphonuclear leukocytes in injured tissue, edema, mast cell degranulation, and alterations in the epidermis basal layer (Figure 3(B)) when compared to PBS-injected group (control) (Figure 3(A)). After 1 h of injection, besides edema and degranulation of numerous mast cells, a mild cell infiltration was noticed, injury overrunning epidermal parabasal layers and blood vessels filled with leukocytes (Figure 3(C) and (D)). In the course of time, three studied venom doses caused a large leukocyte influx and increased the injury in epidermal layers after 4 h of injection. Furthermore, at 2 μg venom dose, polymorphonuclear cells were observed among injured epidermal cells layers (Figure 3(E)) and degranulated mast cells were still present in tissue injected with 32 μg of venom (Figure 3(F)). At 24 h, it was verified regeneration of epidermis, which was apparently complete in the group injected with 2 μg of venom (Figure 4(B)). However, in tissue injected with 8 or 32 μg, new layers of epithelial cells were observed beneath the injured epidermis and the presence of numerous blood vessels in the inflamed region (Figure 4(C) and (D)) when compared to the control group (Figure 4(A)). At 48 h, structural changes in the dermis and connective tissue, and inflammatory infiltrate were still present in the footpads for three venom doses. In relation to epidermis, it was almost completely regenerated within 48 h in animals injected with 8 μg (Figure 4(E)), whereas in tissue injected with 32 μg of venom, epidermal cell layers affected by venom, containing polymorphonuclear cells were still detected (Figure 4(F)).
Histological analyses of mice footpad injected with P. motoro venom. Mice footpad were injected with PBS (control) (A) or 2 (C, E), 8 (D) or 32 μg (B, F) of venom and animals were euthanized after 30 min (B), 1 h (C, D), or 4 h (A, E, F). Epidermis (arrows), dermis (d), adipocytes (a), glandular tissue (g), edema (e), degranulation of mast cells (arrow heads), mild inflammatory influx (*), intense inflammatory influx (#), and blood vessels (v). Tissue was embedded in historesin and stained with toluidine blue and fuchsin. (A color version of this figure is available in the online journal.) Histological analyses of mice footpad injected with P. motoro venom. Mice footpad were injected with PBS (control) (A) or 2 (B), 8 (C, E) or 32 μg (D, F) of venom and animals were euthanized after 24 h (A, B, C, D) or 48 h (E, F). Epidermis (arrows), injured epidermis (stars), dermis (d), glandular tissue (g), edema (e), mild inflammatory influx (*), high inflammatory influx (#), nerve (n), and blood vessels (v). Tissue was embedded in historesin and stained with toluidine blue and fucsine. (A color version of this figure is available in the online journal.)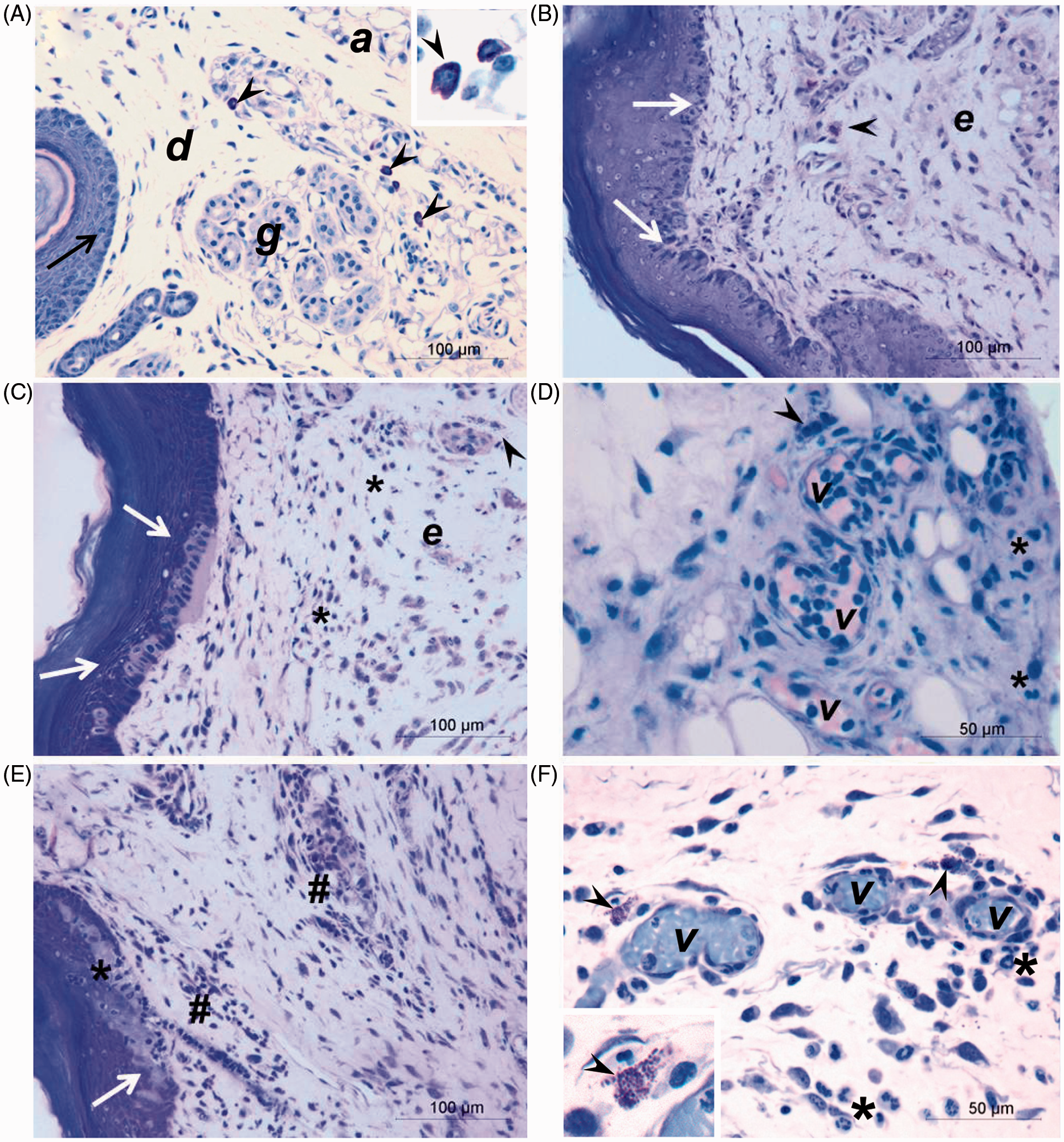
Detection of inflammatory mediators at the site of injection of P. motoro venom
Although P. motoro venom stimulated the production of inflammatory mediators (IL-6, MCP-1 and KC) as soon as 1 h after venom injection, these were significantly increased at 4 h, when compared to the control group (PBS) (Figure 5). At 24 h, only MCP-1 was detected in samples from animals injected with 32 µg of venom (Figure 5(b)).
Effect of P. motoro venom on production of IL-6 (a) and the chemokines MCP-1 (b) and KC (c). Mice were injected into footpads with P. motoro venom (2, 8, or 32 μg) or PBS (control) (n = 5–6). After 30 min, 1, 4, 24, and 48 h of venom injection, animals were euthanized and footpads were removed and processed to assess inflammatory mediators release using EIA kits. ANOVA test *P< 0.05 in relation to control group (PBS)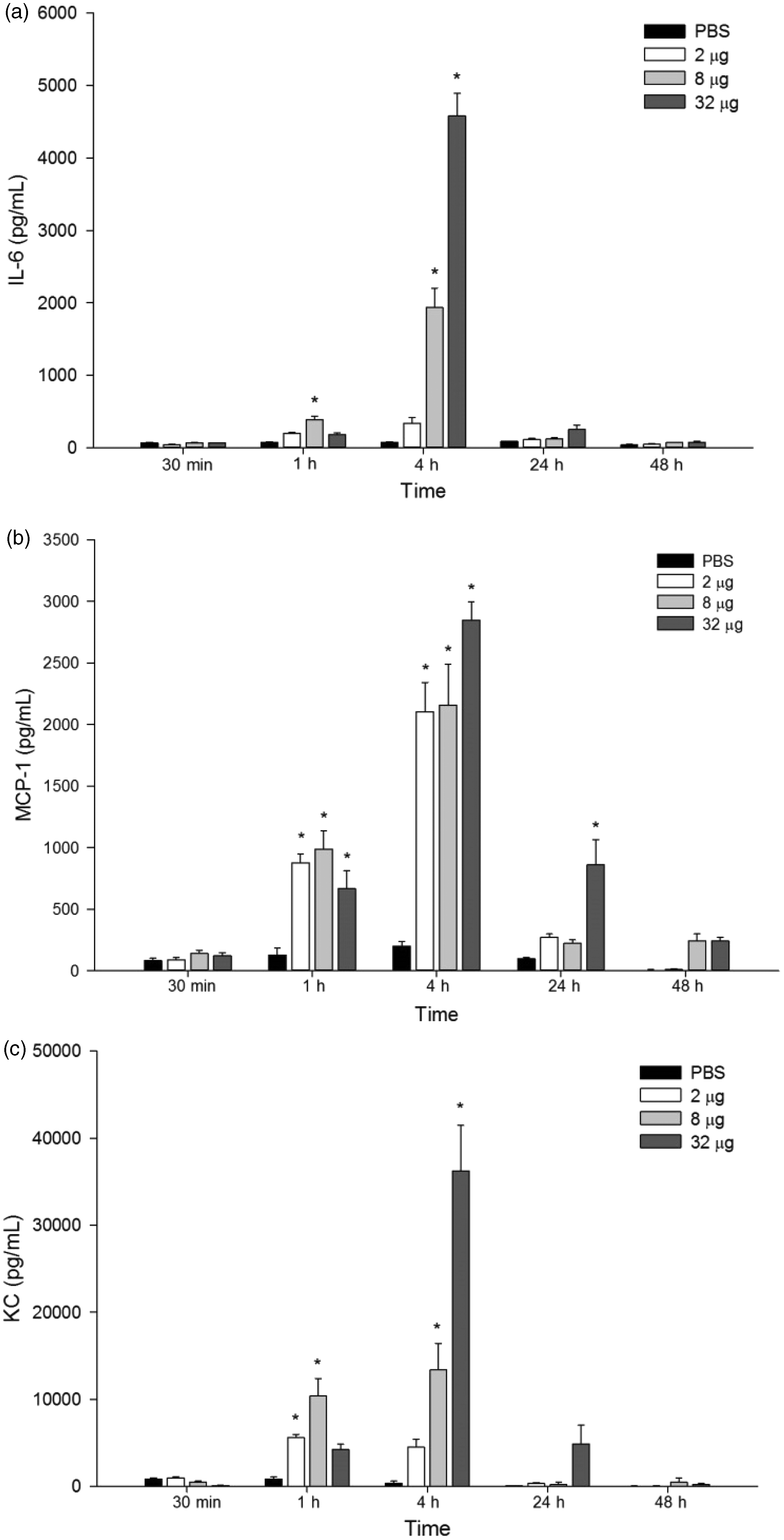
Discussion
The freshwater stingray envenomation causes an intense local reaction, which can lead to necrosis formation. The characterization of inflammatory activities induced by stingray venoms is rarely described in the literature. Thus, this study first aimed to verify the ability of P. motoro stingray venom to induce edema in an animal model.
It was found that animals presented edema already in 15 min after venom injection, with a peak of activity at 30 min, and remained up to 48 h, indicating the rapidity and persistence of this activity. These results were also observed in animals injected with marine (Dasyatis guttata) and freshwater (Potamotrygon falkneri, Potamotrygon cf. scobina, and Potamotrygon gr. orbignyi) stingray venoms.9,19 However, in these studies, the edema was no longer present at 24 h after venom injection. Such variation in relation to the duration of edema response induced by different stingrays venom may be, besides interspecific differences between venoms and/or differences in measurement techniques, due to the loss of venom activity caused by thermolability, 19 demonstrating the need of extreme care in the storage of these toxins. Edema activity induced by animal venoms in experimental models is reported in the literature; however, edema duration and intensity may also vary depending on the venom studied.27–31
After edematogenic activity analysis, our next step was to further characterize the inflammatory reaction induced by P. motoro venom, by counting the leukocytes at the injection site. In order to mimic envenomation in humans caused by stingrays, which usually strike hands or feet, the experimental model used in this study was the intraplantar injection of P. motoro venom instead of intraperitoneal injection, which is the most common method used for the analysis of venom-induced inflammatory aspects. Our results showed that the venom induced an increase in the number of leukocytes at the site of injection at all times and venom doses evaluated. This recruitment, in all periods analyzed, consisted mostly of neutrophils and in later times, the count of monocyte/macrophages also augmented. This fact confirms the cellular mechanism of the inflammatory response described in the literature, in which there is a predominance of neutrophils in the acute phase response and the appearance of monocytes/macrophages in the early stage of inflammation resolution.32,33 The presence of leukocytes at the inflammation site plays multiple roles, such as phagocytosis, release of toxic components, tissue repair, and production and release of some inflammatory mediators that induce the recruitment and accumulation of additional cells at the site. In this study, we quantified the presence of some of these mediators, which attract mononuclear cells (MCP-1), polymophonuclear cells (KC), and a cytokine that is a mark of acute inflammation (IL-6) at the site of P. motoro venom injection in mouse. High production of KC was verified at the initial time (1 and 4 h), which probably provided chemotaxis for neutrophils, since this function for this chemokine is widely described in literature.32,34,35 The level of chemokine MCP-1, which mainly attracts monocytes to the site, was increased in 1 and 4 h after venom injection, and it was still detected at 24 h at the highest dose of venom tested. In addition, high levels of IL-6 at 4 h after P. motoro venom injection provide neutrophil recruitment in early inflammatory response and promotes influx of monocytes, neutrophil apoptosis, and removal of cellular debris during the response, contributing as a regulator for transition in recruitment among these cell types.32,33 In addition to neutrophils and monocytes/macrophages, P. motoro venom also induced an increase in lymphocyte and eosinophil count in the injection site. This may be due to production of other chemokines, probably belonging to subtype C-C, which are responsible for recruiting these cells, and/or also by the presence of certain cytokines (e.g., TNF and IL-1) secreted by many types of cells.34–36
Our results corroborate to the data obtained by Monteiro-dos-Santos et al. 20 The authors studied inflammatory aspects induced by Potamotrygon cf. henlei stingray venom species found in rivers of the Araguaia-Tocantins basin. The authors observed an increased leukocyte count, especially neutrophils, in mice paw homogenized in 6 h after i.pl. venom injection. Furthermore, it was found that P. cf. henlei venom induced the release of IL-6, MCP-1, IL-1, and KC in 2 h after venom injection in mouse paw. Leukocyte migration and production and release of inflammatory mediators, as a result of local injury caused by other animal venoms, are already described in literature.27,29,37 The presence of leukocytes in the injured site may be an essential condition to promote tissue repair as described by Teixeira et al. 38 who observed that the depletion of neutrophils in experimental animals caused less efficient regenerative response and delayed cleaning of necrotic tissue in the pathological changes induced by Bothrops asper snake venom. This suggestion was also proposed by Pareja-Santos et al. 39 for the envenomation caused by Thalassophryne nattereri fish venom, where the absence of leukocyte recruitment in early times and the reduction in mononuclear cells count in 48 h hindered the process of local inflammatory resolution reaction.
We also studied the alterations on the tissue injected with P. motoro venom by microscopic analysis of the injured site. It was observed that the venom provoked an intense local inflammatory reaction, promoting mast cell degranulation, irreversible epidermal damage and angiogenesis, and causing edema and cell migration. Furthermore, we found that most of these reactions, such as edema, injury to the epidermis, and mast cell degranulation were dependent on the dose of injected venom. However, the presence of inflammatory cell infiltrate was intense in all doses of venom studied. Similar result was also observed after injection of P. falkneri venom on the back of mice. This venom induced severe local reaction with edema formation and cell migration. Furthermore, the dose used (400 μg) promoted necrosis 24 h from injection, affecting both the epidermis and dermis, and the subcutaneous tissue and skeletal muscle. After this period, regenerative hyperplasia of epidermal layers was observed in 96 h. 22 Recently, Lameiras et al. 40 also described systemic rhabdomyolysis induced by P. motoro venom, and inflammatory infiltrate at the site of venom inoculation in the gastrocnemius muscle of mice.
In conclusion, the results of this study show for the first time some local inflammatory phenomena related to envenomation caused by P. motoro stingray venom. This venom induced rapid and acute inflammation in mice with persistent edema activity, significant leukocyte migration, and release of inflammatory mediators at the site of injection. Also, histological analysis showed that the venom caused mast cell degranulation and tissue injury. There is no species-specific treatment for stingray accidents, and clinical treatment and wound healing in patients stung by freshwater stingrays are still a challenge.4,41 The results presented in the manuscript were the first step to understand the inflammatory mechanism of action of P. motoro venom. However, our study did not evaluate the possible treatments to diminish the local signs of envenomation, because other parameters, e.g., venom dose, evolution of local reaction, time interval to receive medical care, as well as the different intensity of trauma must be accounted and used to provide the most appropriate treatment for patients. Therefore, further studies are being carried out in our laboratory to determine inflammatory pathways that provide targets for therapies for stingray accidents.
Footnotes
Author contributions
All authors participated in the design, interpretation of studies, analysis of the data, and review of the manuscript. LFK, JPPN, and KCB conducted the in vivo experiments, MMA and SGSJ conducted histological processing, and MLS contributed to statistical analysis. The data of the present paper are part of the MD thesis of LFK.
ACKNOWLEDGEMENTS
The authors thank Marcos Teixeira da Silveira and Antenor Pires do Nascimento for helping in stingrays capture, and Pollyanna Fernandes, Pedro Fontana, Vanessa Mendes, Dr Domingos Garrone-Neto and Bianca Távora, for their support. This paper was supported by São Paulo Research Foundation (FAPESP) (2007/55272-4, 2012/00166-3) and INCTTOX. We also thank Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for the grant to K.C.B. (305359/2010-0) and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for the grant to L.F.K. (PROAP 680058).