
Abstract
Perivascular adipose tissue (PVAT)-derived leptin is a detrimental adipocytokine and plays a critical role in the development of cardiovascular diseases in metabolic syndrome (MetS). During vascular remodeling, vascular smooth muscle cells (VSMCs) undergo phenotypic switching into a synthetic phenotype characterized by decreased expression of differentiation markers (smooth muscle myosin heavy chain, α-smooth muscle actin, and calponin) and increased proliferation. We aimed to determine whether PVAT-derived leptin influences VSMC phenotypic switching and to explore the underlying mechanisms in MetS rats. In vivo, 32 Wistar rats were divided into two groups that received either a normal diet (control rat) or a high-fat diet (MetS rats). After 16 weeks, rat aortas were stained using hematoxylin–eosin and imaged. VSMC differentiation markers and proliferating cell nuclear antigen (PCNA), PVAT-derived leptin, aortic leptin receptor (ObR), and p38 mitogen-activated protein kinase (MAPK) expression were detected. In vitro, aortic VSMCs were incubated with MetS rat PVAT conditioned medium (PVAT-CM) to mimic in vivo conditions and were pretreated with a p38 MAPK inhibitor (SB 203580) or leptin antagonist. Differentiation marker expression, including PCNA and p38 MAPK, was detected. MetS rats exhibited pronounced insulin resistance, hyperglycemia, hyperlipidemia, hypertension, obesity, and an associated increase in PVAT weight. VSMCs underwent phenotypic switching in MetS rat aorta and contributed to vascular remodeling. PVAT-derived leptin expression was higher in MetS rats than in control rats (P < 0.01). ObRa expression and p38 MAPK phosphorylation were upregulated in MetS rat aorta. In vitro, VSMCs incubated with MetS rat PVAT-CM underwent phenotypic switching, associated with increased p38 MAPK phosphorylation. This VSMC phenotypic switching was inhibited by pretreatment with SB 203580 or a leptin antagonist. These results suggest that in MetS rats, PVAT-derived leptin promotes VSMC phenotypic switching via a p38 MAPK-dependent pathway to exacerbate vascular remodeling.
Keywords
Introduction
As the body’s largest endocrine and paracrine organ, adipose tissue not only stores excess body energy but also secretes a variety of bioactive adipocytokines, such as leptin, resistin, and adiponection. 1 Adipocytokine production also influences the development of cardiovascular diseases. 2 Although adipocytokines link cardiovascular disease to metabolic syndrome (MetS), the exact mechanisms are poorly understood. 3
It is well documented that perivascular adipose tissue (PVAT), which normally surrounds large arteries, plays a critical role in the regulation of vascular function. Importantly, a causal link between PVAT function and vascular lesions has been demonstrated. For example, PVAT-derived leptin exacerbates vascular endothelial dysfunction in MetS animals. 4 In MetS, PVAT becomes dysfunctional and production of PVAT-derived protective factors diminishes while production of detrimental adipocytokines, such as leptin, increases, ultimately resulting in vascular lesions.5,6
Leptin is predominantly produced by adipocytes and plays an important role in MetS-associated cardiovascular disease. 7 Leptin is a proliferative and migratory factor for vascular smooth muscle cells (VSMCs) and acts via mitogen-activated protein kinase (MAPK) and phosphatidylinositol 3-kinase signaling pathways. 8 Although recent studies in MetS animals show that locally elevated leptin levels may promote neointima formation and endothelial dysfunction, little is known about the effect of local leptin on VSMCs in MetS.4,9
VSMCs exhibit multiple phenotypes. In normal mature blood vessels, VSMCs exhibit a ‘contractile’ or differentiated phenotype characterized by increased expression of differentiation markers, such as smooth muscle myosin heavy chain (SM-MHC), α-smooth muscle actin (α-SMA), and calponin. During pathogenic vascular remodeling, VSMCs exhibit a ‘noncontractile’ or synthetic phenotype characterized by decreased expression of differentiation markers, an increased rate of proliferation, and synthesis of extracellular matrix components.10,11 However, the mechanisms regulating VSMC phenotypic switching are not fully understood. Previous studies have demonstrated that MAPK signaling pathways play a vital role in phenotypic switching. 12 Furthermore, multiple in vitro experiments have demonstrated that leptin promotes VSMC proliferation via the p38 MAPK signaling pathway.8,13 Based on this evidence, we hypothesized that PVAT-derived leptin promotes VSMC phenotypic switching in MetS via the p38 MAPK signaling pathway. In the present study, we examined VSMC phenotypic switching induced by PVAT-derived leptin in vivo and in vitro and sought to explore the potential mechanisms underlying this effect.
Materials and methods
Preparation of the MetS animal model and experimental groups
Experiments were approved by the Ethical Committee for Animal Experimentation of the First Affiliated Hospital of Xi’an Jiaotong University (Shaanxi, Xi’an, China). The study conforms to the Guide for the Care and Use of Laboratory Animals published by US National Institutes of Health (NIH Publication No. 85-23, revised 1996). In total, 32 specific pathogen-free male Wistar rats (8 weeks old, weighing 200 ± 50 g) were obtained from the Zoological Research Center of Peking Union Hospital, Chinese Academy of Medical Science (animal certification number: SCXK, Beijing 2011-0003). Rats were individually housed in a constant air-conditioned environment with a 12-h day–night cycle. Room temperature was maintained at 25 ± 0.5℃, and humidity was maintained between 55 and 60%. All animals were allowed free access to food and water. Rats were randomly divided into two groups and habituated to research assistants for 1 week before experiments. Each group was fed either a high-fat diet (HFD) or normal diet (ND). Rats in the control group (n = 10) were fed a ND containing 50% carbohydrate, 20% protein and 8% fat, and 0% cholesterol. Rats in MetS group (n = 22) were fed a HFD containing 30% carbohydrate, 4.8% protein and 12% fat, 20% sucrose, 10% lard, 2.5% cholesterol, and 1.4% sodium cholate. The duration of the diet period was 16 weeks for both MetS and control rats. All laboratory chows were purchased from the Xi’an Jiaotong University Laboratory Animal Center (Shaanxi, Xi’an, China).
Physiological and biochemical evaluation
The mental state, fur luster, water intake, and urine output of the two groups were monitored every day, and body mass was determined once a week. Fasting blood glucose, plasma triglyceride (TG), total cholesterol (TC), high-density lipoprotein cholesterol (HDL-C), and low-density lipoprotein cholesterol (LDL-C) were measured every 4 weeks. An oral glucose tolerance test and an insulin release test were performed at week 0 and week 16 (corresponding to 25-week-old rats), respectively. At both time points, fasting insulin (FINS) was measured using a Rat Insulin ELISA Kit (Elabscience, Wuhan, China) and insulin resistance was assessed through homeostasis model assessment (HOMA), as described in previous studies. 14 At the end of the experiment, carotid blood pressure was measured by right carotid artery catheterization. Blood samples were collected and plasma was removed by centrifugation and then snap frozen at −20℃. Plasma TC, HDL-C, TG, and LDL-C were determined using a Hitachi 7600-010 Automatic Biochemical Analyzer (Hitachi High Technologies Corp., Tokyo, Japan). Plasma glucose concentrations were measured in duplicate using the glucose oxidase method. Carotid arterial blood pressure was measured using a BL-420 F biological signal collecting and processing system (Taimeng Biotech. Co., Sichuan, Chengdu, China), and systolic blood pressure (SBP), diastolic blood pressure (DBP), and mean arterial pressure (MAP) were calculated using the software provided by the manufacturer.
Pathological evaluation of aorta
At the end of the experiment, animals were killed and thoracic aorta and PVAT were carefully dissected and weighed. Segments (5 mm thick) of aortic tissue resected near to the aortic arch were fixed with 4% paraformaldehyde for 12 h, then dehydrated, treated with a clearing agent, wax dipped, and embedded into paraffin blocks. Tissues were then sectioned for hematoxylin–eosin staining. Aortic morphology was observed using an Olympus BX51 microscope (Olympus, Japan). Aortic medial thickness (MT), lumen diameter (LD), and MT/LD ratio (MT/LD) were measured using computer-assisted image analysis software (QWin, Leica Imaging Systems Ltd, Cambridge, UK). Fresh PVAT was used to make PVAT conditioned medium (PVAT-CM) as described below. Other tissue samples were cleaned of blood in cold physiological saline, blotted, and weighed immediately and then frozen in liquid nitrogen and stored at −80℃ until protein or mRNA extraction. All operations were in RNAse free conditions.
Semi-quantitative real-time polymerase chain reaction (RT-PCR) of PVAT-derived leptin and aortic leptin receptor mRNA
PVAT-derived leptin (Ob) mRNA, long-form (ObRb) and short-form (ObRa) leptin receptor mRNA expression was determined by semiquantitative RT-PCR as previously described. 15 Total RNA was extracted from aortic and PVAT using TRIZOL reagent (Invitrogen, Carlsbad, CA, USA), and the quantity and quality of isolated RNA was determined. RT-PCR was performed using SYBR® Premix Ex TaqII (TaKaRa Biotechnology, Tokyo, Japan) following the manufacturer’s instructions. Expression of β-actin mRNA was used as a control. Primers used for RT-PCR were based on a previous study and are listed below: Ob, 5′-TCACACACGCAGTCGGTATCC-3′ (sense), and 5′-GTCTCGCAGGTTCTCCAGGTC-3′ (antisense); ObRa, 5′-TGATATCGCCAAACAGCAAA-3′(sense), and 5′-AGTGTCCGCTCTCTTTTGGA-3′ (antisense); ObRb, 5′-AGCAGTCCAGCCTACACTCTT-3′ (sense), and 5′-ACCACATACCTCCTCACACTACAC-3′ (antisense); β-actin, 5′-CTATCGGCAATGAGCGGTTCC-3′ (sense), and 5′-TGTGTTGGCATAGAGGTATTTACG-3′ (antisense). 16 The relative quantities of cDNA were calculated from cycle thresholds (Ct) and were normalized to the housekeeping gene β-actin using the 2-ΔΔCt method. 17
Western blot analysis
Leptin, leptin receptor (ObRb), α-SMA, SM-MHC, calponin, proliferating cell nuclear antigen (PCNA), total p38 MAPK (t-p38 MAPK), phosphorylated p38 MAPK (p-p38 MAPK), and β-actin protein expression were determined by western blotting. Western blotting was performed as previously described. 18 Briefly, aorta and PVAT were homogenized, centrifuged, and the resulting supernatants were collected for analysis. Equivalent amounts of protein were loaded onto 10% acrylamide gels for electrophoresis and blotting. Immunoreactivity was visualized using an enhanced chemiluminescence western blotting detection system. Western blot bands for the target protein and the internal reference β-actin were scanned and analyzed using Image-Pro Plus 6.0 software (Media Cybernetics Inc., Rockville, MD, USA); results are shown as the ratio of integrated optical density for both proteins, and statistical analysis was performed. The following primary antibodies were used: antileptin (Abcam, Cambridge, MA, USA), anti-ObR (B-3: sc-8391, Santa Cruz Biotechnologies, CA, USA), anti-α-SMA (Epitomics Inc., CA, USA), anti-SM-MHC (Bioss Inc., Woburn, MA, USA), anticalponin (ab46794, Abcam, Cambridge, MA, USA), anti-PCNA (Epitomics Inc., CA, USA), anti-phospho-p38 MAPK (Thr180/Tyr182), and anti-total-p38 MAPK (Cell Signaling Technology, Beverly, MA, USA).
Materials
Fetal bovine serum, antibiotics, trypsin,
Cell culture and treatments
To determine whether MetS rat PVAT-derived leptin induces VSMCs phenotypic switching via the p38 MAPK signaling pathway, we performed in vitro experiments. Primary VSMCs were obtained by enzymatic digestion of the thoracic aortic media of normal male Wistar rats (weighing 200 ± 50 g, 8 weeks old) purchased from Chi Scientific Inc. (Catalog No. 1-5067, Jiangyin, Jiangsu, China). Since passaged VSMCs show a dedifferentiated phenotype even under quiescent culture conditions, we prepared primary VSMCs, which maintain a differentiated phenotype as previously described. 19 VSMCs were identified by immunofluorescence staining. Cells were cultured at 37℃ in a humidified atmosphere of 95% air and 5% CO2. Before each experiment, cells were serum starved for 24 h in α-MEM containing 5.5 mM glucose and antibiotics (streptomycin 100 mg/mL and penicillin 100 U/mL).
PVAT-CM was prepared and concentrated as described previously. 20 Four-hundred milligrams of MetS rat PVAT were collected and conditioned at 37℃ for 24 h in 1 mL of serum-free DMEM-Ham’s F-12 medium containing 0.2% BSA. The PVAT-CM was centrifuged, frozen, and stored at −80℃ until use. As PVAT-derived leptin may be involved in VSMC phenotypic switching in MetS rats, we incubated VSMCs with MetS rat PVAT-CM for 48 h to mimic in vitro conditions (MetS group). PVAT from control rats was collected and prepared as described previously, and VSMCs were exposed to control rat PVAT-CM for 48 h (control group). To prevent adipokine depletion, the medium was changed every 8 h. To determine whether PVAT-derived leptin promotes VSMCs phenotypic switching via activation of p38 MAPK signaling pathway in MetS, cells were pretreated with the p38 MAPK inhibitor SB 203580 (10 µM for 24 h, SB203580 group) or leptin tA (10 µM for 24 h, leptin tA group) before incubation with MetS rat PVAT-CM for 48 h. Expression of α-SMA, SM-MHC, calponin, PCNA, t-p38 MAPK, and p-p38 MAPK was then determined by western blot analysis as described previously.
Further proof-of-principle studies were performed to examine the concentration-dependent response of normal rat aortic VSMCs to leptin administration (in the absence and presence of p38 MAPK inhibitor). The expression of α-SMA, SM-MHC, calponin, PCNA, t-p38 MAPK, and p-p38 MAPK were determined by western blot analysis as described previously.
Detection of leptin in PVAT-CM
Leptin protein levels in PVAT-CM were determined using a Rat Leptin ELISA Kit (ChrystalChem, Chicago, IL, USA) according to the manufacturer’s instructions.
Statistical methods
Statistical analysis was performed with SPSS version 16.0 (SPSS Inc., Chicago, IL, USA). Data are the mean ± standard error of mean, and differences between two groups were determined using an independent t-test. Statistical differences between multiple groups were determined using analysis of variance followed by Scheffe’s multiple comparison test. P < 0.05 was considered statistically significant.
Results
MetS rat model
The basic data of metabolic parameters of two groups
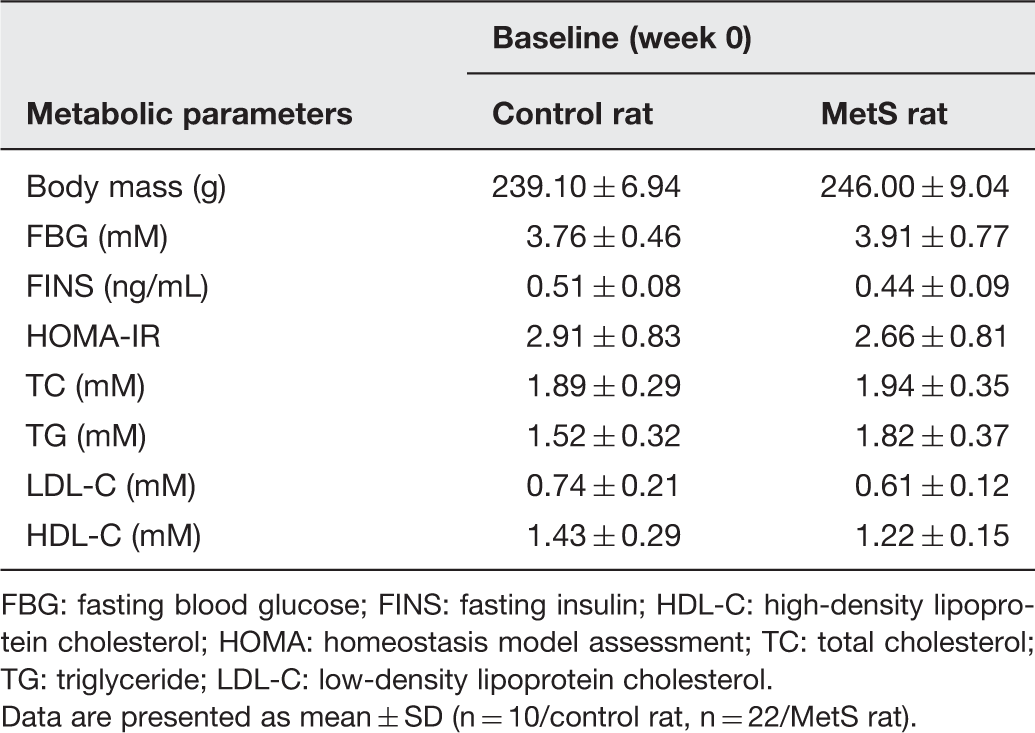
FBG: fasting blood glucose; FINS: fasting insulin; HDL-C: high-density lipoprotein cholesterol; HOMA: homeostasis model assessment; TC: total cholesterol; TG: triglyceride; LDL-C: low-density lipoprotein cholesterol.
Data are presented as mean ± SD (n = 10/control rat, n = 22/MetS rat).
The data of metabolic parameters of two groups at the 16th week (at the end of the experiment)
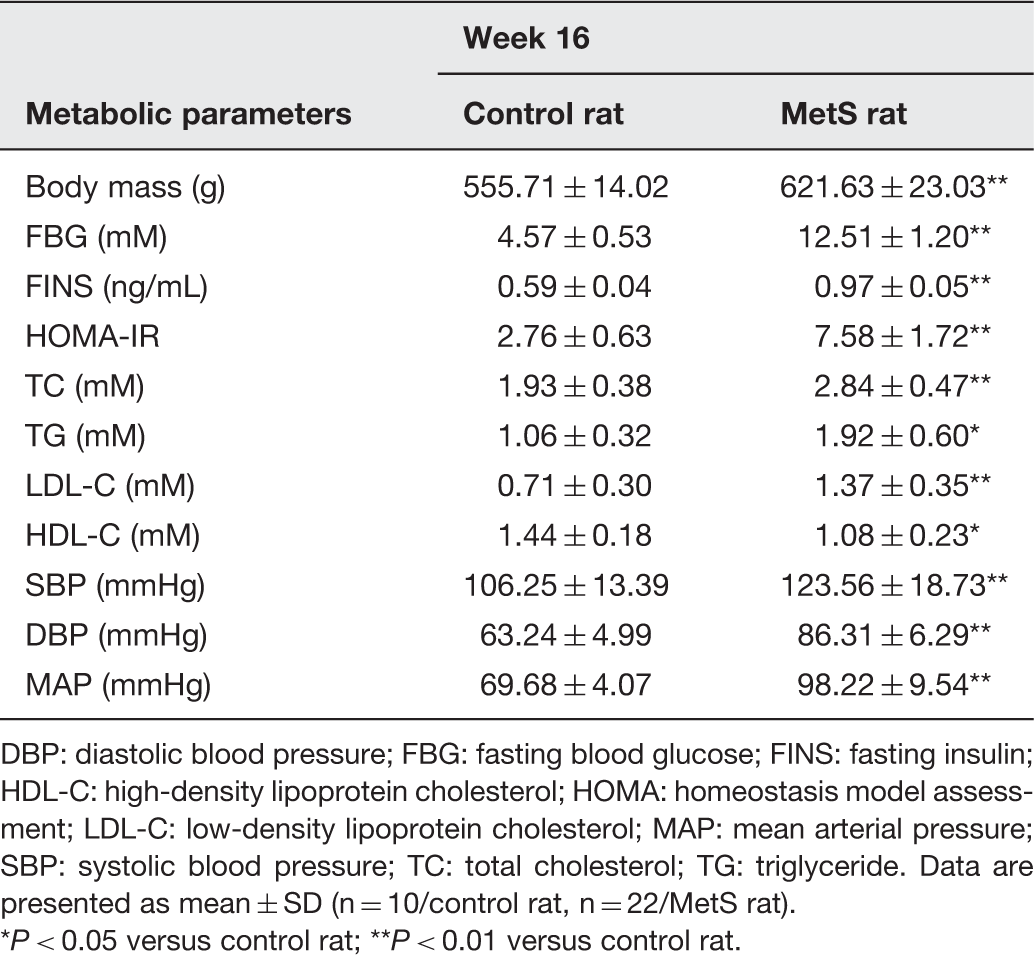
DBP: diastolic blood pressure; FBG: fasting blood glucose; FINS: fasting insulin; HDL-C: high-density lipoprotein cholesterol; HOMA: homeostasis model assessment; LDL-C: low-density lipoprotein cholesterol; MAP: mean arterial pressure; SBP: systolic blood pressure; TC: total cholesterol; TG: triglyceride. Data are presented as mean ± SD (n = 10/control rat, n = 22/MetS rat).
P < 0.05 versus control rat; **P < 0.01 versus control rat.
Vascular remodeling of MetS rat aorta
We next examined structural changes in the aorta to determine whether vascular remodeling had occurred. In the MetS rats, MT and MT/LD were significantly higher than in control rats (P < 0.01, Table 3). LD was lower in MetS rats than in control rats; however, this was not statistically significant. Hematoxylin–eosin staining demonstrated that aortic tissue structure was normal in control rats (Figure 1(a)). By contrast, there was significant aortic medial thickening in MetS rats (Figure 1(b)). The above results indicate that aortic remodeling occurred in MetS rats. The MetS rat model used herein is therefore a useful model of cardiovascular disease, as vascular remodeling plays a critical role in the development of cardiovascular disease. As VSMC phenotypic switching plays a key role in vascular remodeling, phenotypic switching was investigated in MetS and control rats.
Hematoxylin–eosin staining of aorta (original magnification: 400×). (a) Normal control rat’s aorta. Note that the intima, media, and adventitia layer boundaries are distinct and medial VSMCs are arranged in an orderly manner. No medial thickening or aortic lesions were observed. Scale bar represents 50 µm. (b) MetS rat’s aorta. Here, medial VSMCs are arranged in a disorderly manner and medial thickening is observed. The structure of each layer is unclear, and endothelial cells are discontinuous. Scale bar represents 50 µm. (A color version of this figure is available in the online journal.) The morphological measure results of aortic structure LD: lumen diameter; MT: medial thickness; MT/LD: the MT and LD ratio. Data are presented as mean ± SD (n = 10/control rat, n = 22/MetS rat). P < 0.01 versus control rat.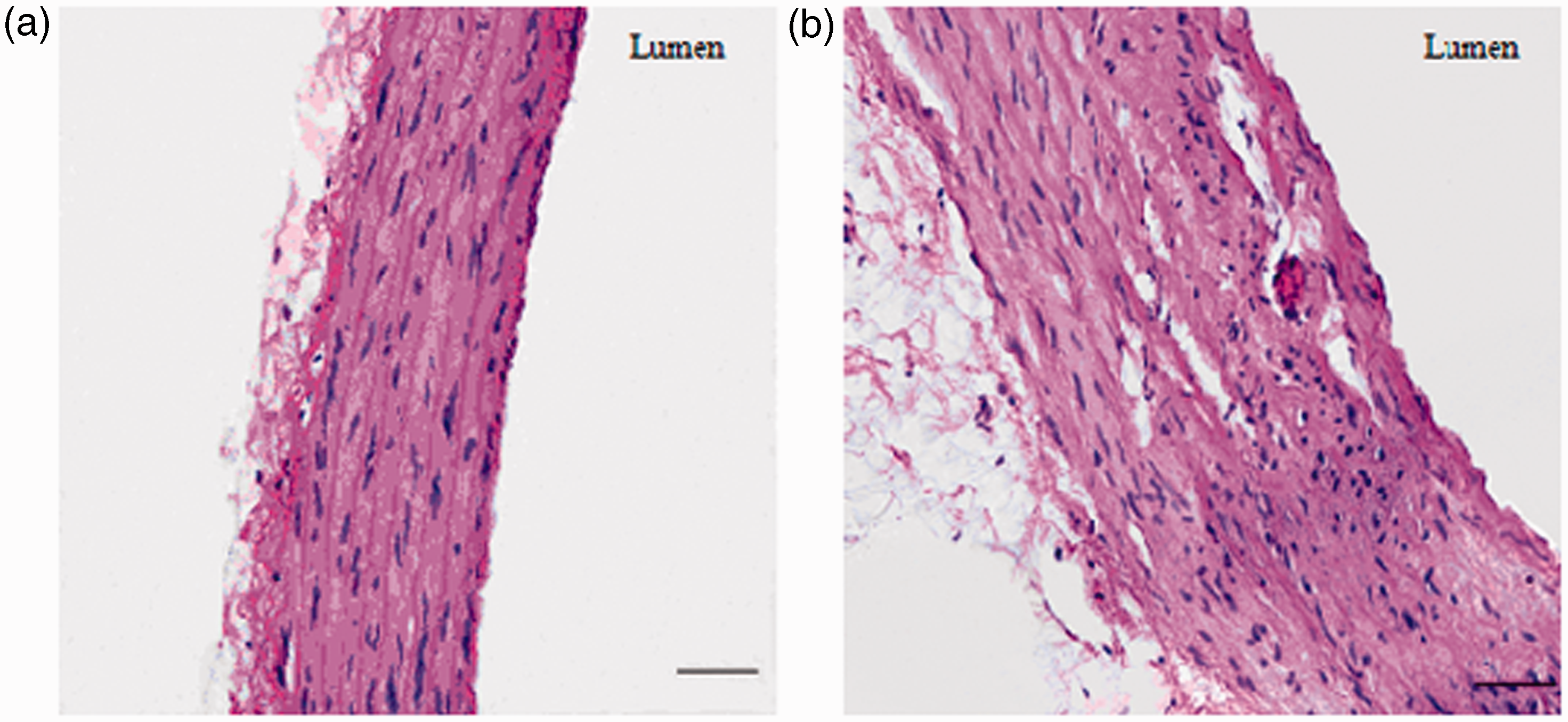

VSMC phenotypic switching in MetS rat aorta
As expected, after feeding with a HFD for 16 weeks, MetS rat aortas exhibited significantly lower expression of VSMC differentiation markers, including α-SMA, SM-MHC, and calponin, than the aortas of control rats (P < 0.01, Figure 2). By contrast, PCNA expression was significantly higher in MetS rat aorta than in control rat aorta. PCNA is a nuclear protein that acts as a cofactor for DNA polymerase in DNA synthesis during the S phase of the cell cycle.
21
PCNA is expressed in most proliferating cells, so it is a reliable biomarker for evaluation of cell proliferation.
22
Thus, in MetS rat aorta, VSMCs underwent phenotypic switching from the contractile phenotype to the synthetic phenotype, which participates in the pathogenesis of vascular remodeling, as described previously.
The VSMC differentiation markers and PCNA expression in rat aorta. After fed with HFD for 16 weeks, in MetS rat aorta, the VSMC differentiation markers, including α-SMA, SM-MHC, and calponin expression, were significantly decreased, while the PCNA expression was significantly increased. *P < 0.01 versus control rat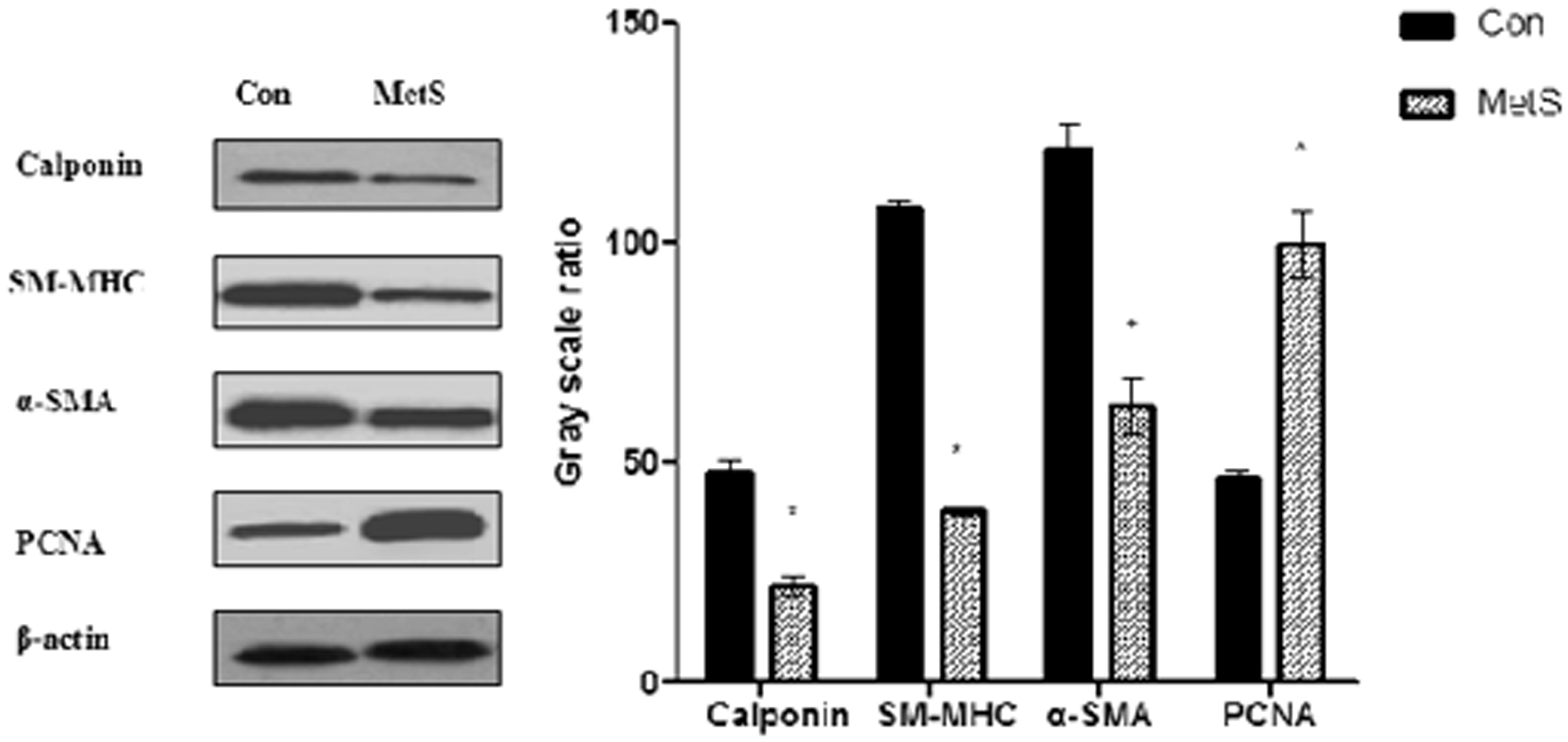
Elevated PVAT weight is associated with upregulation of PVAT-derived leptin and aortic ObRa expression in MetS rats
To exclude possible effects of body weight, PVAT weight was adjusted for the body weight of each animal. PVAT content was significantly higher in MetS rats than in control rats (P < 0.01, Figure 3(a)). As expected, PVAT-derived leptin mRNA and protein expression were significantly higher in MetS rats than in control rats (P < 0.01, Figure 3(b) and (d)). Interestingly, ObRa mRNA expression in aortic tissue was significantly higher in MetS rats than in control rats (P < 0.01, Figure 3(c)). However, there was no difference in ObRb mRNA expression between the two groups (Figure 3(c)). Furthermore, in MetS rat aorta, ObR protein expression was significantly higher in MetS rats than in control rats (P < 0.01, Figure 3(d)). Notably, the ObR (B-3: sc-8391) antibody used in the present study can detect both short- and long-form ObR. The molecular weight of ObRa and ObRb are 100 and 125 kDa, respectively. However, only one band (located at approximately 105 kDa) was detected in the western blots herein. Overall, the results indicate that ObRa expression was increased in MetS rat aorta.
(a) PVAT weight in two groups. The amount of PVAT was significantly increased in MetS rat. (b) PVAT-derived leptin mRNA expression in two groups. PVAT-derived leptin mRNA expression was significantly increased in MetS rat. (c) Expression of ObRa and ObRb mRNA in two groups. The ObRa mRNA expression was significantly increased in MetS rat aorta. (d) PVAT-derived leptin and aortic ObR expression in two groups. PVAT derived-leptin protein expression and ObR protein expression in aortic were significantly increased in MetS rat. *P < 0.01 versus control rat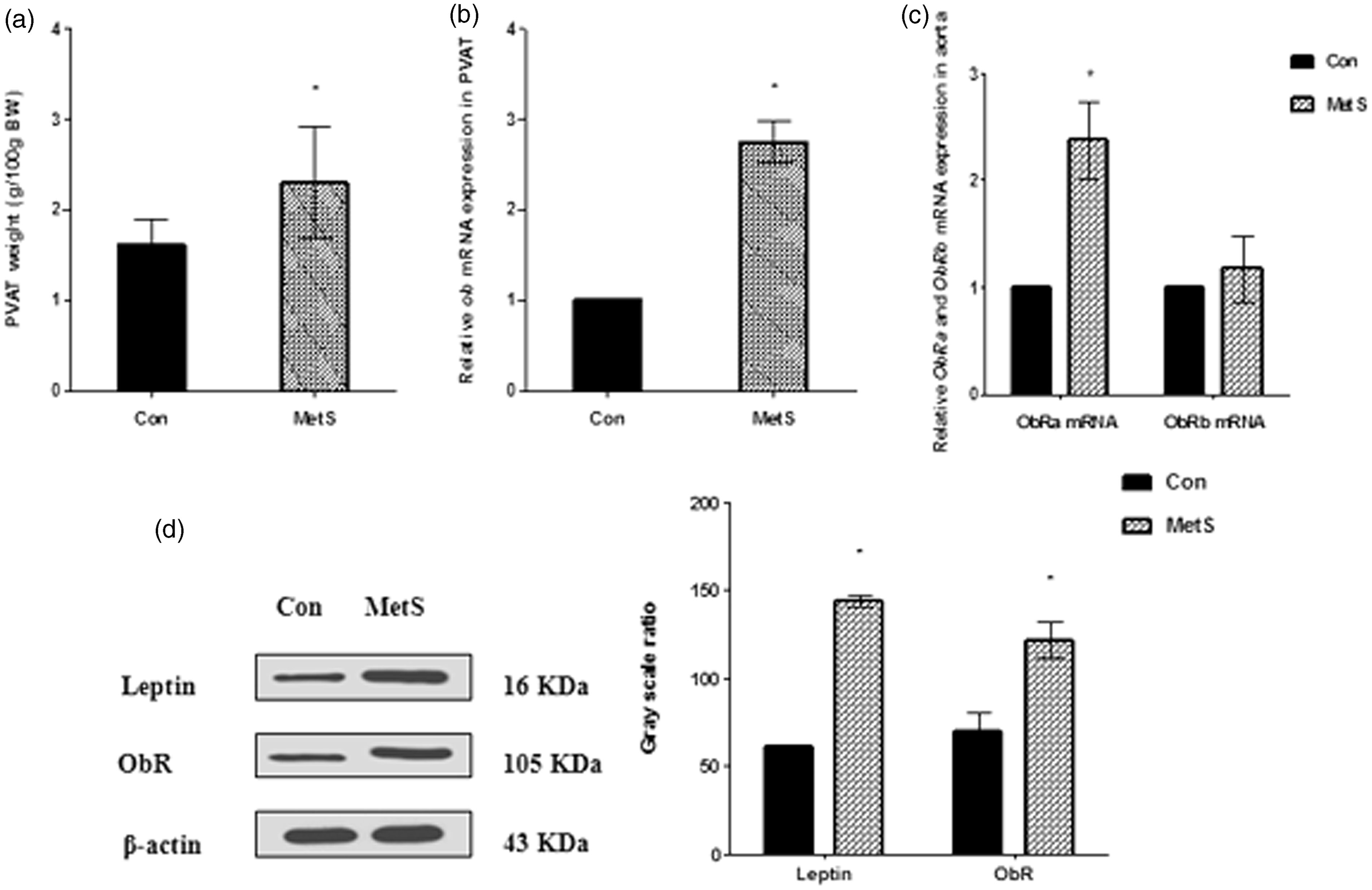
Increased p38 MAPK phosphorylation in MetS rat aorta
Western blot analysis was used to determine the expression of p-p38 MAPK and t-p38 MAPK in rat aorta. As shown, there was no significant difference in t-p38 MAPK expression between control rats and MetS rats (Figure 4). However, p-p38 MAPK expression was significantly higher in MetS rats than in control rats (P < 0.01, Figure 4). The results indicate that the p38 MAPK signaling pathway was activated in MetS rat aorta. However, a number of other factors are also associated with activation of p38 MAPK signaling during MetS. Hence, further experiments were performed in vitro to confirm our hypothesis.
The p38 MAPK expression in rat aorta. After fed with HFD for 16 weeks, in MetS rat aorta, the p-p38 MAPK expression was significantly increased. *P < 0.01 versus control rat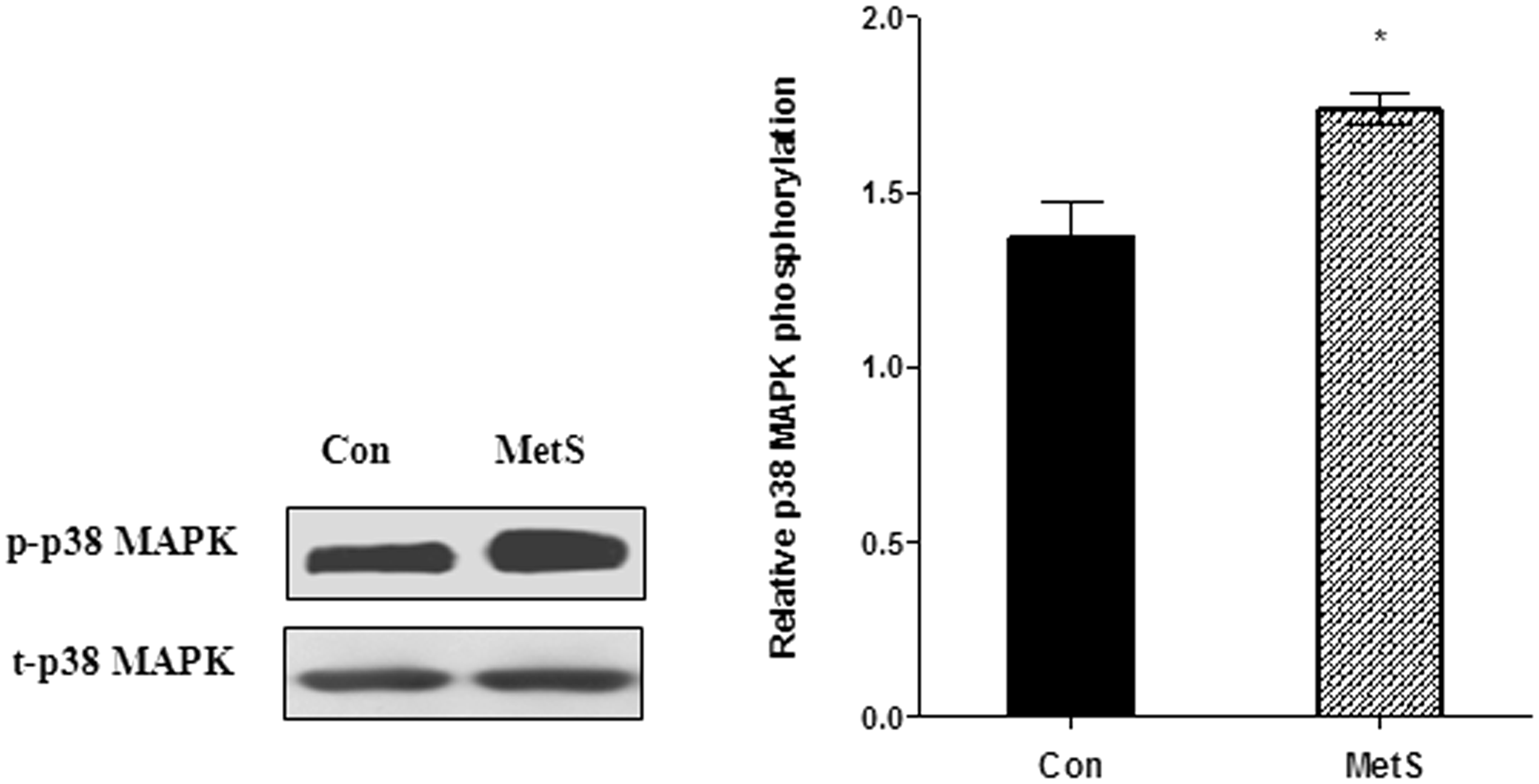
Immunofluorescence staining to identify VSMCs
Calponin is a marker of the contractile phenotype of VSMCs and appears to be restricted almost exclusively to differentiated VSMCs.
23
VSMCs in present study were not contaminated with fibroblasts or endothelial cells as evidenced by >98% positive immunofluorescence staining for calponin, as determined using a Fluorescein-5-isothiocyanate (FITC)-conjugated calponin antibody (Figure 5).
Immunofluorescence staining for identification of VSMCs. Images were captured using a laser confocal microscope (Leica TCS SP5 II, Germany) and processed with Image-Pro Plus 6.0 software (Media Cybernetics Inc., Rockville, MD, USA). (a) Immunofluorescence staining for calponin in VSMCs (original magnification: 200×). Green fluorescence represents calponin. Cell nuclei were counterstained with 4',6-diamidino-2-phenylindole (DAPI) (blue). VSMCs show a spindle-like shape and calponin is highly expressed in cytoplasm. Scale bar represents 100 µm. (b) Negative control with secondary antibody (original magnification: 200×). Cell nuclei were counterstained with DAPI (blue). Scale bar represents 100 µm. (A color version of this figure is available in the online journal.)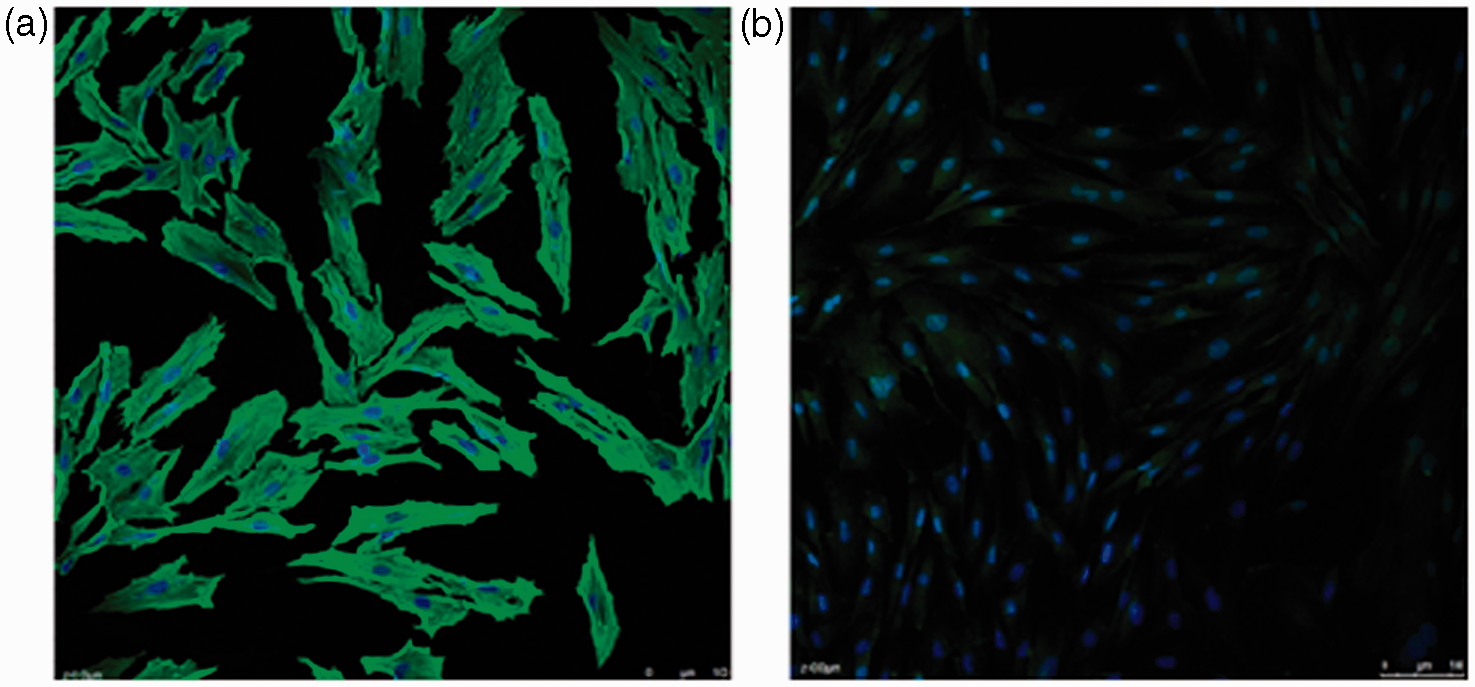
Detection of leptin levels in PVAT-CM
In contrast to control rat PVAT-CM, MetS rat PVAT-CM contained significantly higher levels of leptin (P < 0.01, Figure 6), which is consistent with the in vivo results. This suggests that PVAT-CM may be a good in vitro model of the PVAT microenvironment. Hence, the effect of PVAT-derived leptin on VSMCs phenotypic switching was examined using this conditioned medium.
The PVAT-CM leptin level in two groups. In MetS rat PVAT-CM, leptin level was significantly increased. *P < 0.01 versus control group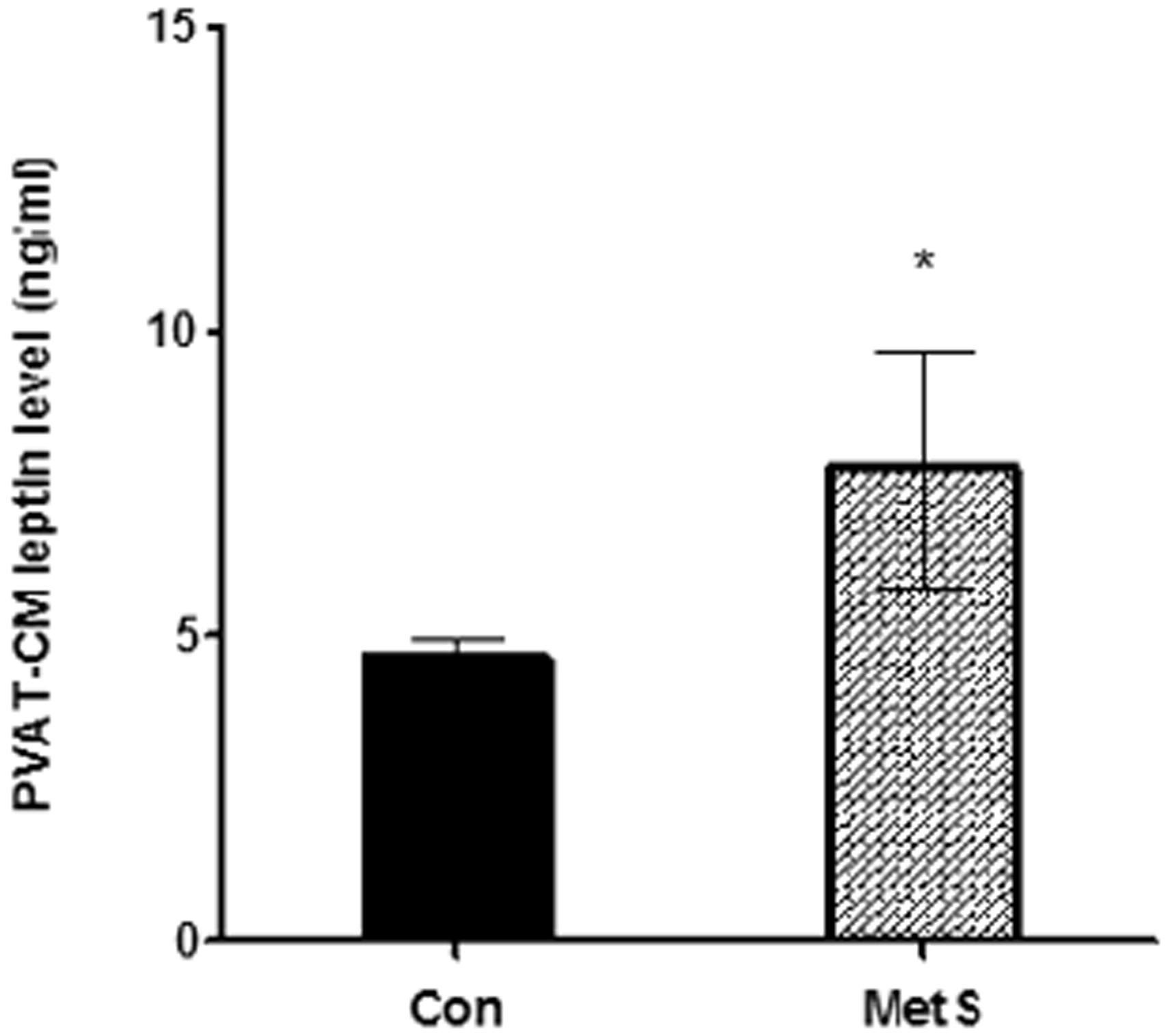
P38 MAPK phosphorylation is upregulated in VSMCs incubated with MetS rat PVAT-CM and downregulated in VSMCs pretreated with leptin tA
As expected, there was no difference in t-p38 MAPK expression between each of the four groups examined (Figure 7). However, p-p38 MAPK expression was significantly higher in the MetS group than in the control group (P < 0.01, Figure 7). The leptin tA-pretreated group exhibited significantly lower levels of p-p38 MAPK expression than the MetS group (P < 0.01, Figure 7). This indicates that PVAT-derived leptin from MetS rats stimulates p-p38 MAPK expression and activates the p38 MAPK signaling pathway. Interestingly, p-p38 MAPK expression in the leptin tA group was higher than in the SB203580 group (P < 0.01, Figure 7), which suggests that leptin tA only partially blocks the p38 MPAK signaling pathway.
The p38 MAPK expression in four groups. The p-p38 MAPK expression was significantly higher in the MetS group than in the control group, and after pretreatment with a leptin antagonist, p-p38 MAPK expression was significantly decreased. * P < 0.01 versus control group; # P < 0.01 versus MetS group; Δ P < 0.01 versus SB 203580 group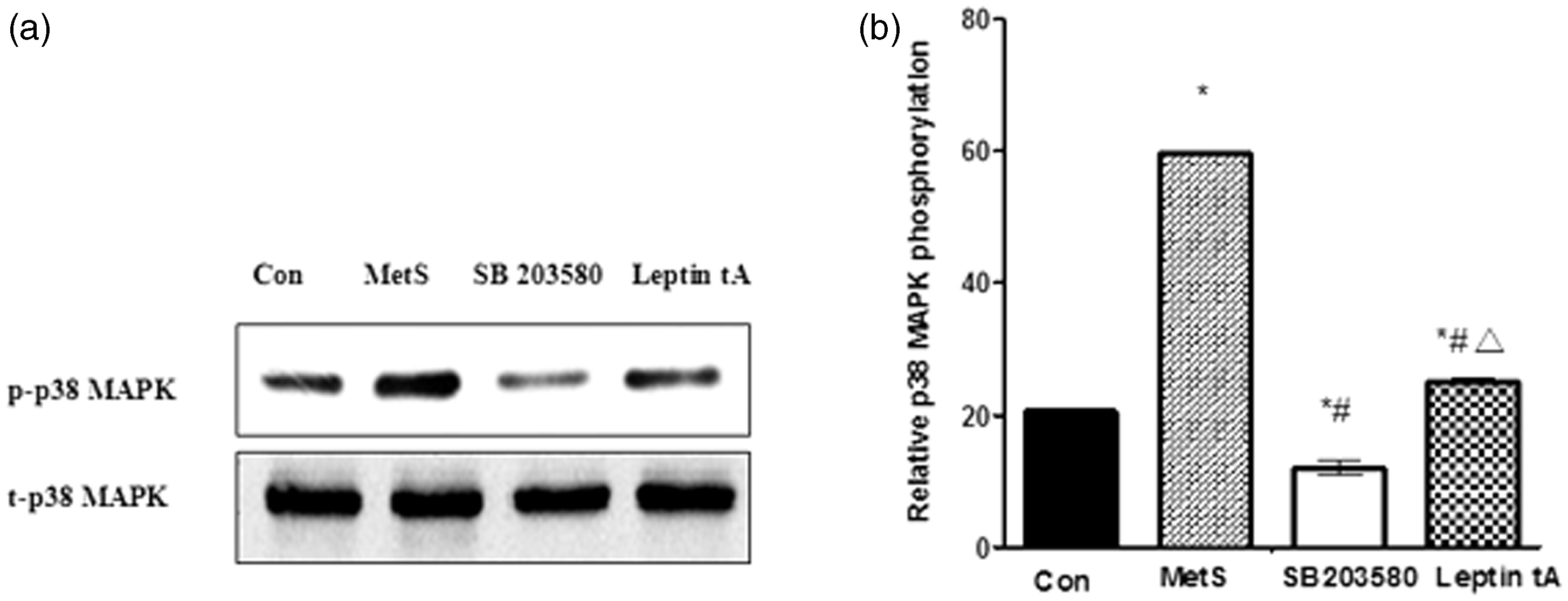
Effect of MetS rat PVAT-CM on differentiation markers and PCNA expression in VSMCs
Differentiation markers, such as α-SMA, SM-MHC, and calponin were highly expressed in VSMCs incubated with control rat PVAT-CM. In contrast to control cells, incubation with MetS rat PVAT-CM for 48 h (i.e. the MetS group) caused a significant decrease in differentiation marker expression (P < 0.01, Figure 8(a)). As expected, differentiation marker expression was significantly higher in leptin tA group than in the MetS group (P < 0.01, Figure 8(a)). This suggests that MetS rat PVAT-derived leptin promotes VSMCs phenotypic switching, which is consistent with the in vivo results earlier.
(a) The VSMC differentiation markers expression in four groups. The differentiation markers including α-SMA, SM-MHC, and calponin expression were significantly lower in the MetS group than in the control group, and after pretreatment with leptin antagonist, differentiation markers expression was significantly increased in comparison with MetS group. (b) The PCNA expression in four groups. The PCNA expression was significantly higher in the MetS group than in the control group, and after pretreatment with leptin antagonist, PCNA expression was significantly decreased in comparison with MetS group. *P < 0.01 versus control group; #P < 0.01 versus MetS group; ΔP < 0.01 versus SB 203580 group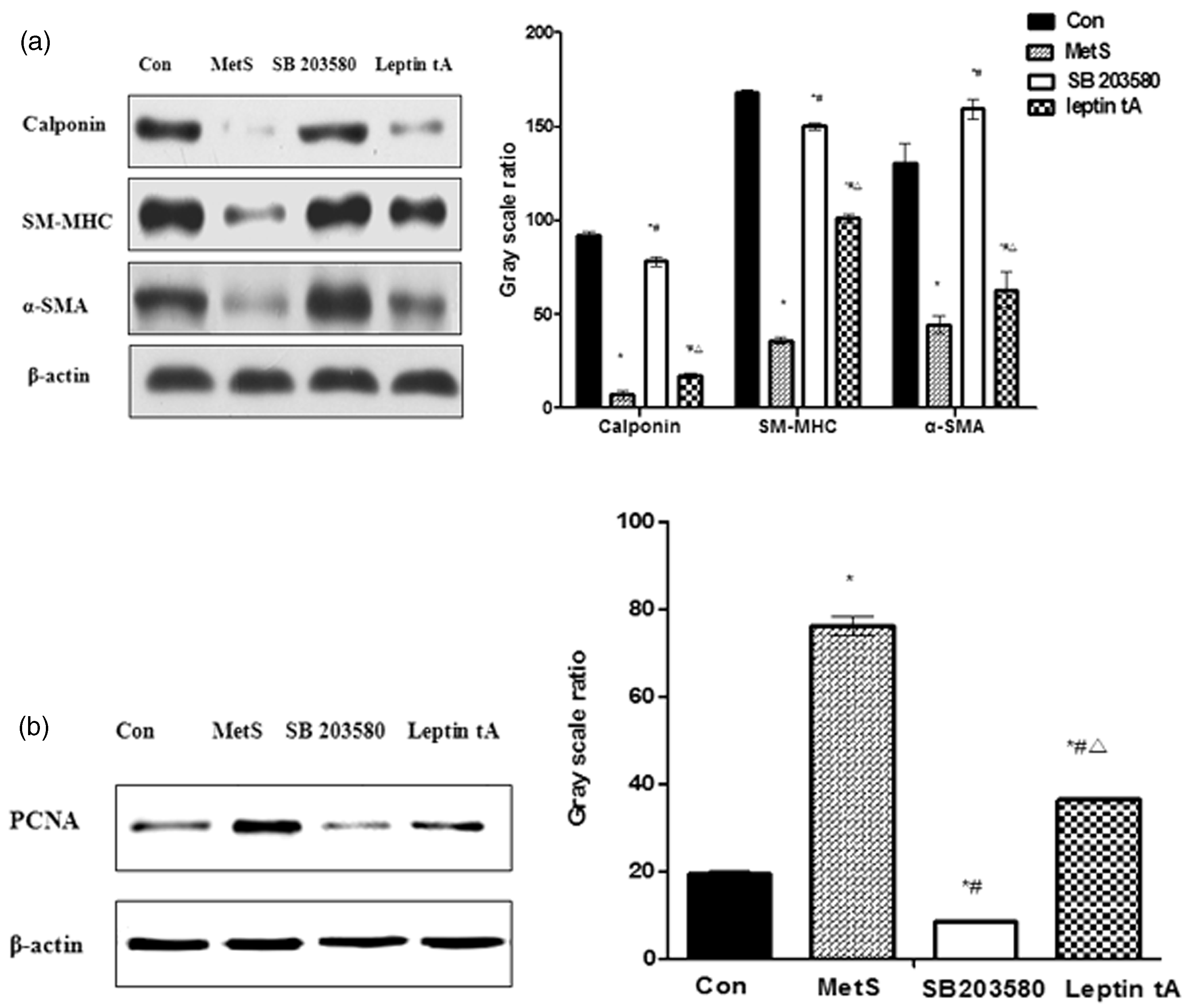
PCNA expression was significantly higher in the MetS group than in the control group, whereas the leptin tA-pretreated group exhibited significantly lower levels of PCNA expression than the MetS group (P < 0.01, Figure 8(b)). This indicates that MetS rat PVAT-derived leptin stimulates VSMCs proliferation. Again, this is consistent with the in vivo results. Combined with the p-p38 MAPK and t-p38 MAPK expression data (Figure 7), these results indicate that PVAT-derived leptin from MetS rats stimulates VSMC phenotypic switching via activation of the p38 MAPK signaling pathway.
Interestingly, differentiation marker expression was lower in the leptin tA group than the SB 203580 group, while PCNA expression was higher in leptin tA group than the SB 203580 group (P < 0.01, Figure 8(a) and (b)). These results indicate that MetS rat PVAT-CM-induced VSMC phenotypic switching can be partly blocked by pretreatment with a leptin antagonist. These results, combined with the p-p38 MAPK and t-p38 MAPK expression data from the in vitro experiments (Figure 7), suggest that, in addition to leptin, other PVAT-derived adipocytokines may promote VSMC phenotypic switching via activation of the p38 MAPK signaling pathway.
Leptin stimulates VSMC phenotypic switching in a concentration-dependent manner
Additional principle studies were performed to examine the concentration-dependent response of normal rat VSMCs to leptin administration. Rat VSMCs were treated with various concentrations (0–100 ng/mL) of leptin for 48 h, and differentiation markers and PCNA expression were determined by western blot. As we expected, leptin decreased the differentiation markers (α-SMA, SM-MHC, calponin) expression and increased PCNA expression in a concentration-dependent manner, and the maximal and minimal differentiated effect were at 100 and 80 ng/mL of leptin, respectively (Figure 9). It is reasonable to indicate that leptin stimulates VSMC phenotypic switching in a concentration-dependent manner.
Leptin induces VMSC phenotypic switching in a concentration-dependent manner. VSMCs were incubated with different concentrations of leptin for 48 h. PCNA, α-SMA, SM-MHC, and calponin expression were determined by western blot analysis. Leptin decreased the expression of differentiation markers and increased PCNA expression in a concentration-dependent manner. The maximal and minimal differentiated effect was at 100 and 80 ng/mL of leptin, respectively. *P < 0.01 versus leptin 0 ng/mL. (A color version of this figure is available in the online journal.)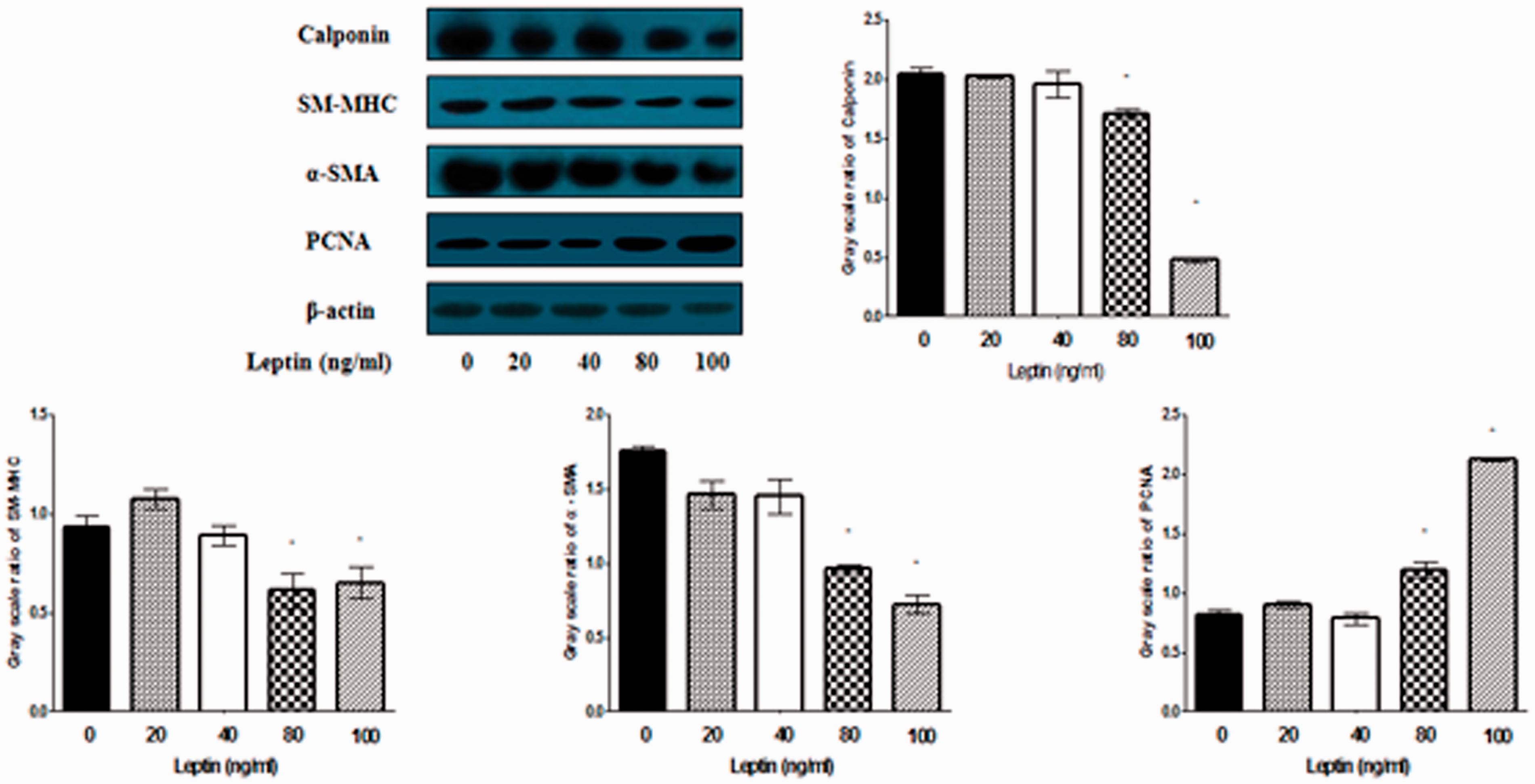
Leptin induces VSMC phenotypic switching via activation of the p38 MAPK signaling pathway
Since the maximal effect of leptin on VSMC phenotypic switching was at 100 ng/mL, VMSCs were treated with 100 ng/mL leptin for 48 h, either with or without pretreatment with the p38 MAPK inhibitor SB 203580 (10 µM for 24 h). Cells were then assayed for expression of the differentiation markers, PCNA and p38 MAPK, to determine whether leptin induces VSMCs phenotypic switching via activation of the p38 MAPK signaling pathway. As expected, leptin treatment had no significant effect on t-p38 MAPK expression in VSMCs. However, p-p38 MAPK expression was markedly increased, which is consistent with previous studies (Figure 10(a)). As shown in Figure 10(b), leptin-induced phenotypic switching was accompanied by activation of the p38 MAPK signaling pathway, which could be inhibited by pretreatment with SB 203580. Thus, leptin stimulates VSMC phenotypic switching via activation of the p38 MAPK signaling pathway.
Leptin stimulates VSMC phenotypic switching via activation of the p38 MAPK signaling pathway. VSMCs were incubated with leptin (100 ng/mL for 48 h) with or without pretreatment with the p38 MAPK inhibitor SB 203580 (10 µM for 24 h). PCNA, α-SMA, SM-MHC, calponin, t-p38 MAPK, and p-p38 MAPK expression were determined by western blot analysis. (a) Leptin treatment had no significant effect on t-p38 MAPK expression. However, p-p38 MAPK protein expression was markedly increased. *P < 0.01 versus leptin (–)/SB 203580(–); #P < 0.01 versus leptin (+)/SB 203580(–). (b) Leptin-induced VSMC phenotypic switching was inhibited by pretreatment with SB 203580. *P < 0.01 versus leptin (–)/SB 203580(–); #P < 0.01 versus leptin (+)/SB 203580(–)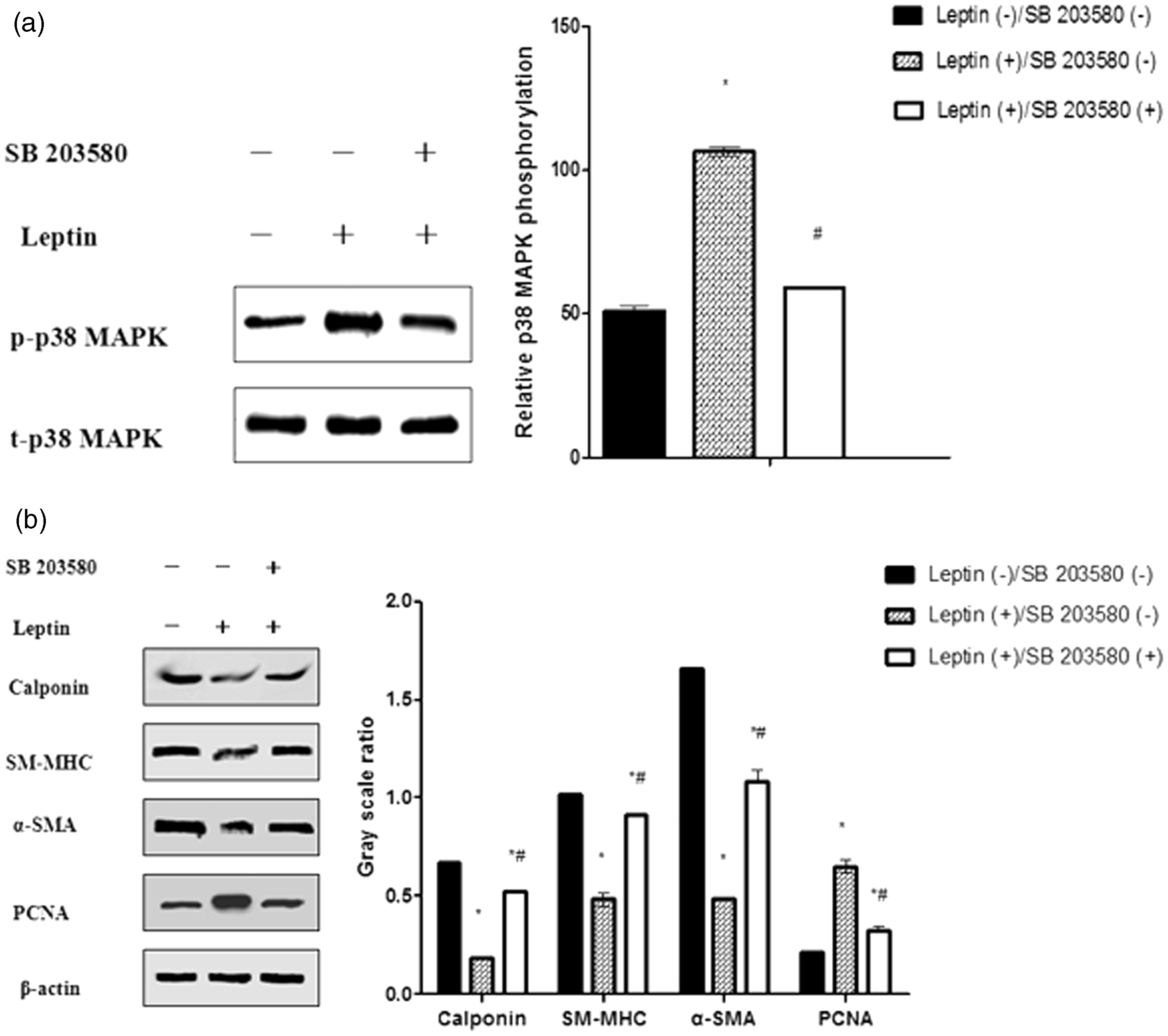
Discussion
The main findings of this study were: (i) PVAT-derived leptin and aortic ObRa expression were upregulated in MetS rats; (ii) PVAT-derived leptin from MetS rats promotes VSMC phenotypic switching via activation of the p38 MAPK signaling pathway; and (iii) PVAT-derived leptin from MetS rats promotes VSMC phenotypic switching at a low concentration.
The results showed that PVAT-derived leptin levels were significantly higher in the MetS rats than in the control rats (P < 0.01, Figure 3(b) and (d)). Furthermore, ObRa expression was upregulated in MetS rat aorta, concomitant with vascular remodeling (Figures 1(b) and 3(c) and (d)). In the HFD-induced MetS rats, PVAT weight was significantly higher than in normal control rats (P < 0.01, Figure 3(a)), which is consistent with the results of previous studies. 24 Leptin is an adipose tissue-derived hormone, thus PVAT-derived leptin levels were also increased as a result of the increase in PVAT weight. Recent studies show that PVAT-derived adipocytokines, such as leptin, are bioactive at very low concentrations, and 10% receptor occupancy is often sufficient to trigger a response. 25 It is therefore likely that in MetS rats, upregulation of both PVAT-derived leptin and aortic ObRa may exacerbate the adverse effects of leptin on the aorta.
There are six different isoforms of the leptin receptor, including ObRa-f. 26 However, leptin primarily exerts its biological effects through the long-form receptor ObRb, distributed mainly in hypothalamus, where it mediates neural signaling.13,27 Interestingly, in the present study, ObRa (short-form ObR) was highly expressed in MetS rat aorta (P < 0.01, Figure 3(c) and (d)). However, there was no statistically significant difference in aortic ObRb mRNA expression between control rats and MetS rats (Figure 3(c)). Although ObRb is referred to as the ‘signaling isoform,’ there is evidence of signaling by other short-form isoforms. Notably, ObRa binds leptin and induces activation of JAKs and the extracellular signal-regulated kinase (ERK)/MAPK signaling pathway. 28 We speculate that, in MetS rat aorta, activation of ObRa may indirectly or directly activate multiple pathways and may have a pathophysiological effect on VSMCs. Future experiments are planned involving small interfering RNA knockdown of ObRa in VSMCs to test this hypothesis. Furthermore, in the present study, only two isoforms of ObR mRNA were detected, further studies have been planned to detect expression of other ObR isoforms in MetS rat aorta.
In the present study, we demonstrated that MetS rat PVAT-derived leptin promotes VSMC phenotypic switching via activation of the p38 MAPK signaling pathway. To the best of our knowledge, this is the first time that this has been demonstrated. Our data showed that PVAT-derived leptin was upregulated in MetS rats, and aortic VSMC phenotypic switching was associated with activation of the p38 MAPK signaling pathway. Furthermore, in VSMCs incubated with MetS rat PVAT-CM, VSMC phenotypic switching was dependent upon activation of the p38 MAPK signaling pathway, as both the p38 MAPK inhibitor SB203580 and a leptin antagonist inhibited VSMC phenotypic switching. VSMC phenotypic switching is critical during the onset of vascular diseases, such as vascular remodeling, hypertension, and atherosclerosis. Our results indicate that PVAT-derived leptin is a critical pathogenic factor in MetS vascular remodeling. Notably, a recent study showed that PVAT, which is in close proximity to vascular walls, modulates vessel function through a paracrine mechanism. 24 A similar paracrine mechanism may be in place for the local leptin-induced phenotypic switching observed herein. This paracrine mechanism provides a novel insight into vascular disease and may be a target for therapeutic intervention.
Previous studies have shown that leptin stimulates VSMC proliferation in a concentration-dependent manner via activation of the p38 MAPK signaling pathway, and that these effects of leptin on VSMC proliferation occur at a supraphysiological dose in vitro.8,29,30 Similarly, we have demonstrated that leptin induces VSMC phenotypic switching in a concentration-dependent manner via activation of the p38 MAPK signaling pathway in vitro (Figures 9 and 10). The minimum concentration at which the effects of leptin could be observed was 80 ng/mL. Rat plasma leptin levels are modulated by multiple factors, including insulin administration, cold exposure, fasting, and body weight.31,32 However, existing research shows that rat plasma concentrations of leptin are typically between 1.75 and 25 ng/mL.31,32 Thus, as demonstrated herein, leptin stimulates VSMC phenotypic switching at supraphysiological doses in vitro. Interestingly, in the present study, MetS rat PVAT-derived leptin promoted VSMC phenotypic switching at low concentrations (7.74 ± 1.94 ng/mL, Figure 6). Local leptin is known to be bioactive at very low concentrations and can directly modulate VSMC function through a paracrine mechanism. 25 However, in addition to leptin, MetS rat PVAT likely releases multiple detrimental adipocytokines, such as resistin and visfatin, which may be involved in the regulation of VSMC phenotypic switching.33,34 However, further research is needed to confirm this hypothesis.
Interestingly, our study showed that in VSMCs incubated with MetS rat PVAT-CM, p-p38 MAPK expression was partially inhibited by pretreatment with a leptin antagonist (Figure 7), and that MetS rat PVAT-CM-induced VSMC phenotypic switching was also partially blocked by pretreatment with a leptin antagonist (Figure 8). As demonstrated herein, activation of the p38 MAPK signaling pathway is a critical factor for VSMC phenotypic switching, because when p-p38 MAPK expression was partially blocked VSMC phenotypic switching was also partially inhibited. It is well known that, in addition to leptin-mediated activation, the p38 MAPK signaling pathway is activated by multiple adipocytokines and inflammatory cytokines, including visfatin, TNF-α, and IL-β.34,35 In MetS, adipocytokine production is in a state of imbalance, as detrimental cytokines such as leptin, visfatin, and TNF-α are upregulated, whereas protective factors are downregulated. This suggests that there may be positive cross-talk between multiple detrimental adipocytokines at the level of p38 MAPK. Therefore, inhibition of leptin signaling alone may not completely inhibit p38 MAPK signaling and, in turn, p38 MAPK-mediated VSMC phenotypic switching. In addition, previous studies have demonstrated that ERK1/2 and phosphoinositide-3-kinase signaling pathways are involved in the regulation of VSMC function. 19 Since the molecular mechanisms behind phenotypic switching modulation of VSMC are very complex, further studies are needed to elucidate the adipocytokines and signaling pathways involved in VSMC phenotypic switching in MetS animal.
This study demonstrates that in MetS rats, PVAT-derived leptin promotes VSMC phenotypic switching via activation of a leptin-p38 MAPK signaling pathway. This paracrine action of local leptin provides a novel insight into the link between the vasculature and PVAT, and may be of particular significance in vascular diseases in MetS.
Footnotes
Author contributions
All authors participated in the design, interpretation of the studies, analysis of the data, and review of the manuscript. HL and GT designed the studies, performed laboratory experiments, analyzed the data, and wrote the manuscript. Y-P W and L-N Z performed laboratory experiments and analyzed the data.
ACKNOWLEDGEMENTS
This study was supported by the Nature Science Foundation of China (No. 30871042) and the international cooperation in science and technology key projects of Shaanxi province (No. 2012 kw-40-01).