
Abstract
Preclinical toxicity testing in animal models is a cornerstone of the drug development process, yet it is often unable to predict adverse effects and tolerability issues in human subjects. Species-specific responses to investigational drugs have led researchers to utilize human tissues and cells to better estimate human toxicity. Unfortunately, human cell-derived models are imperfect because toxicity is assessed in isolation, removed from the normal physiologic microenvironment. Microphysiological modeling often referred to as ‘organ-on-a-chip’ or ‘human-on-a-chip’ places human tissue into a microfluidic system that mimics the complexity of human in vivo physiology, thereby allowing for toxicity testing on several cell types, tissues, and organs within a more biologically relevant environment. Here we describe important concepts when developing a repro-on-a-chip model. The development of female and male reproductive microfluidic systems is critical to sex-based in vitro toxicity and drug testing. This review addresses the biological and physiological aspects of the male and female reproductive systems in vivo and what should be considered when designing a microphysiological human-on-a-chip model. Additionally, interactions between the reproductive tract and other systems are explored, focusing on the impact of factors and hormones produced by the reproductive tract and disease pathophysiology.
Introduction
Microphysiological modeling attempts to mimic multiple organs in vitro in a single bioengineered system. 1 By recapitulating the various cell and tissue interactions that occur in organs and body systems, these ‘organ-on-a-chip’ or ‘human-on-a-chip’ models aim to accurately replicate the physiology of the human body, with the goal of improving the speed and accuracy of toxicity testing in preclinical drug development. 2 To better mimic the in vivo setting, cells and tissues within microphysiological models are interconnected via microfluidic devices to produce intact anatomical systems for the study of normal function, disease, and response to novel therapeutics. 3 Traditional approaches to preclinical analysis, including the use of animal models and human cell culture methods, do not accurately predict drug toxicity, efficacy, and metabolism in human patients and often delay the drug development process. 4 Furthermore, the ethical concerns of testing drugs in pregnant women has greatly limited our knowledge of the effects of these drugs during gestation and on the developing fetus. 5 Microphysiological models of human physiological systems hold promise to increase the efficiency, accuracy, and safety of toxicity testing in drug development.
Systemic effects of hormone signaling

The development of microphysiological models of the male and female reproductive tracts will not only advance our study of fertility and disease, but also provide a new way to test the effects of therapeutics prior to human trials. Moreover, the ability to link reproductive models to those of other systems will allow for accurate recapitulation of the far-reaching effects of steroid hormones throughout the body. This review will briefly summarize the male and female reproductive organs and their functionality as a system, including the hormone signals that govern reproductive activity. Further, we will highlight the importance of paracrine communication within the male and female reproductive tract and with other organs and systems to illustrate the challenge of developing microphysiological models of the human reproductive system.
Female reproductive tract
The female reproductive tract is a dynamic system. During each menstrual cycle, the uterus sheds and regenerates tissue, the ovary undergoes scarless healing after ovulation, and the entire tract gains and loses the capacity to support the development of offspring. Regulation of these processes requires the coordination of hormones and growth factors that act on and between the ovarian, fallopian, uterine, cervical, and vaginal tissues that make up the reproductive tract (as summarized in Figure 1). The cyclical variations in the hormonal milieu make the female system challenging to investigate, and researchers continue to search for ways to recapitulate this system in cell, tissue, or organ culture.
Reproductive anatomy and steroid hormone signaling. (A color version of this figure is available in the online journal)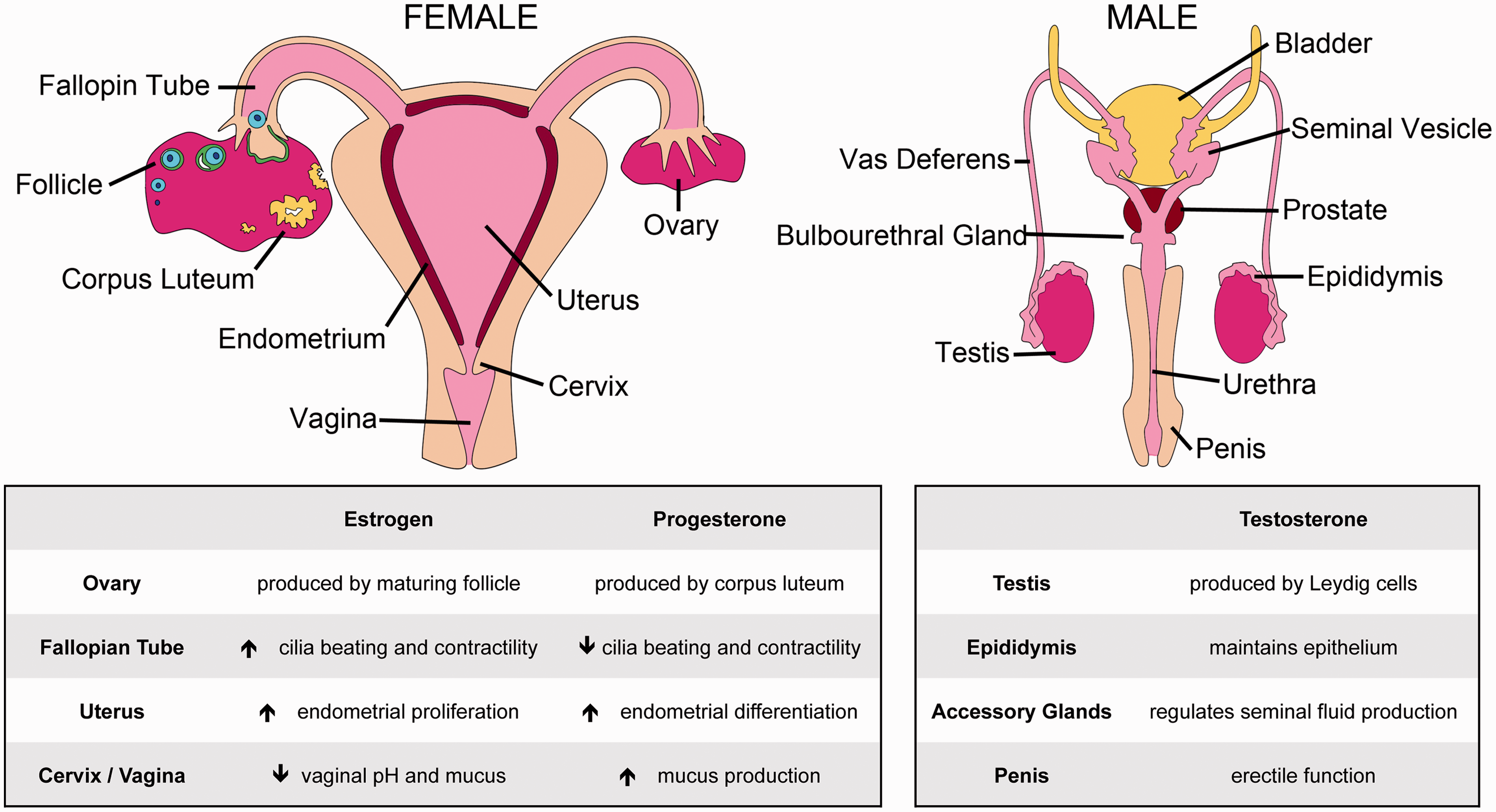
Ovary
The ovary has two interrelated functions: steroidogenesis and production of gametes. Ovarian follicles are composed of a single gamete (oocyte) surrounded by somatic cells, the granulosa and theca cells. Primordial follicles are formed during fetal life and are held in meiotic arrest; with each reproductive cycle, a few primordial follicles resume growth to produce a mature oocyte that is able to be ovulated and fertilized. 16 Although several primordial follicles are recruited in each cycle, in the human, most activated follicles undergo atresia, leaving a singular dominant follicle to reach ovulation. Within the growing follicle, the steroidogenic granulosa and theca cells proliferate and support oocyte maturation and sensitization. Theca cells produce androstenedione, which is converted into estrogen by adjacent granulosa cells. 17 Levels of estrogen and the peptide hormone inhibin rise as the granulosa cell population proliferates, which initiates a negative feedback loop that represses follicle-stimulating hormone (FSH) release from the pituitary and sensitizes the follicle for the impending luteinizing hormone (LH) surge. 17 Upon release of LH from the pituitary, the supporting theca cells produce collagenase enzymes 18 that aid in the rupture of the layer of epithelium surrounding the ovary and the release of the mature oocyte into the fallopian tube.
After ovulation, the ovary undergoes a swift restructuring of the surface epithelium. Granulosa cells from the ovulated follicle differentiate to form the corpus luteum, which produces progesterone 17 that regulates downstream reproductive function and maintains crosstalk with the pituitary. Progesterone from the ovary signals to the rest of the reproductive tract to induce changes that facilitate oocyte transport and implantation. 19 The luteal phase lasts approximately 12 days in women, during which time the corpus luteum degrades and the next set of primordial follicles is selected for activation. In the event of successful fertilization and subsequent implantation, human chorionic gonadotropin (hCG) produced by the blastocyst signals to the ovary to retain the corpus luteum. In turn, progesterone from the ovary preserves the uterine lining to support pregnancy.
Fallopian tube
The distal opening of the fallopian tube consists of finger-like projections called fimbriae that facilitate uptake of the ovulated oocyte. The distal fallopian fimbriae may also be a source of high-grade serous carcinoma. 20 The fallopian fimbriae consist primarily of secretory or ciliated epithelium. In response to the ovarian estrogen increase during ovulation, cilia beat towards the uterus 21 and fallopian smooth muscle peristalsis is stimulated 22 promoting oocyte transport towards the uterus. Ovarian estrogen also stimulates production of essential mucins from the fallopian tube secretory epithelium, such as oviductal-glycoprotein 1 (OVGP1), which promotes fertilization. 23 During the luteal phase, increasing levels of progesterone from the corpus luteum cause a decrease in tubal motility as the fallopian musculature relaxes and cilia movement slows.21,24
Uterus
If an oocyte is fertilized in the fallopian tube, the blastocyst migrates to the uterus where pregnancy is established. The uterus proliferates in response to rising estrogen levels created by developing follicles, allowing for renewal of the endometrial lining that was shed during the previous menses. 19 During the proliferative phase of the menstrual cycle, estrogen also stimulates production of progesterone receptors in the uterus, which will allow the endometrial lining to respond to rising levels of progesterone from the corpus luteum. Progesterone increases endometrial thickening as a result of increased glandular secretion and glycogen accumulation in the uterine stroma. 19 The progesterone-dominated secretory phase produces a nutrient-rich lining where the blastocyst can embed. In the absence of a fertilized egg, the corpus luteum degenerates and progesterone levels drop sharply, which initiates endometrial lining degeneration and discharge during menses. If fertilization occurs, hCG from the blastocyst maintains the corpus luteum in the ovary and the progesterone-regulated development of the uterine lining. Continued progesterone signaling prevents menstruation by suppressing uterine contractility 25 and promotes decidualization of the endometrium, 26 allowing for placental attachment and fetal development.
Cervix and vagina
The neck of the uterus terminates in the cervix, which acts as a barrier between the vagina and the rest of the reproductive tract. During most of the reproductive cycle, the cervix secretes cloudy, thick mucus that obstructs passage of pathogens and sperm into the internal tract. Mucus thickening is particularly evident during progesterone-dominated pregnancy, when the production of an impenetrable mucus plug and cervical remodeling help to protect the developing fetus. 27 In the non-pregnant woman, rising estrogen levels during the proliferative phase of the menstrual cycle change the consistency of cervical secretions to a more fluid-like mucus that promotes sperm migration from the vagina through the cervix. 28 In addition, estrogen regulates vaginal pH29; while the vaginal environment is acidic through most of the menstrual cycle, rising estrogen levels make it more alkaline prior to ovulation, which promotes sperm survival.
Female reproductive endocrine signaling
The organs and tissues of the female reproductive tract are connected via a luminal space as well as via lymphatic ducts and vasculature, which supports crosstalk between organs. The ovary, uterus, and vagina are all supplied by their respective major arteries (ovarian, uterine, and vaginal) 30 ; however, the blood supply is not segregated between these organs, but instead interlinked via anastomosis. 30
As discussed briefly, the female reproductive system is tightly regulated by hormonal signals but is also affected by other secreted proteins and lipids whose actions are more fully discussed elsewhere. 31 It is important to note that the signaling patterns that maintain the reproductive cycle in woman are finite, with folliculogenesis, activation of the fallopian tube and cervix, and uterine menstruation terminating when a woman undergoes menopause. In the absence of follicular development, ovarian production of estrogen and progesterone cease, creating a vastly different steroidogenic microenvironment compared to the cycling woman.
Male reproductive tract
Unlike the female, the male reproductive system functions in perpetuity rather than in a cycle. 32 However, as in the female, male reproductive function also relies on communication between physically and vascularly connected reproductive organs (summarized in Figure 1), making it possible to recapitulate the system and its signaling in vitro with microphysiological modeling.
Testis
As in the ovary, the testis regulates both gamete production and steroidogenesis. The testis consists of two compartments: the interstitium, composed primarily of steroidogenic Leydig cells, and the seminiferous tubules, which contain germ cells undergoing spermatogenesis and their supporting Sertoli cells. Leydig cells provide nutrients for the seminiferous tubules including testosterone, which is produced in response to LH. 32 FSH from the pituitary stimulates Sertoli cells inside the seminiferous tubules to produce several factors including androgen binding protein (ABP) and inhibin. 33 ABP has a high affinity for testosterone and creates a gradient of the testosterone within the seminiferous tubules. Inhibin provides negative feedback to downregulate FSH secretion from the pituitary, keeping the male system in equilibrium. 34 Sertoli cell secretion of ABP, inhibin, and other factors regulates the production and maturation of gametes within the seminiferous tubules. 32
Male gametes proliferate via mitosis initiated by testosterone and FSH signaling starting at puberty. 32 Immature spermatagonia undergo meiotic division to produce spermatids and then spermiogenesis – a transformation that involves elongation, flagellum formation, and nuclear condensation – before being released into the luminal space of the seminiferous tubules. 32 After testosterone-mediated maturation, spermatozoa are propelled along the seminiferous tubules to the efferent ducts connecting the testis to the epididymis.
Epididymis and accessory glands
The epididymis is a coiled tube located posterior to the testis lined with columnar epithelium and microvilli. 35 Spermatozoa from the testis are neither motile nor capable of fertilization, and require large volumes of fluid for propulsion through the seminiferous tubules; this fluid is resorbed in the epididymis via the specialized microvilli. 36 concentrating the sperm. Epididymal secretions partially activate fertilization capacity and motility. 37 a process which is governed by steroidogenic signaling from the testis. ABP synthesized by the Sertoli cells in the testis shuttles testosterone from the testis to the epididymal ducts 38 to maintain the epididymal secretory epithelium. Withdrawal of androgen signaling results in the apoptosis of this epithelium39,40 and leads to infertility in animal models. 41
The epididymis also serves as a storage site for sperm until the time of ejaculation, 42 when sperm move into the vas deferens. The vas deferens transports sperm to the seminal vesicles and other ejaculatory glands prior to expulsion via the urethra. 43 The vas deferens is lined with epithelium which further concentrate sperm and functions to propel, 43 and accessory ejaculatory glands (seminal vesicles, bulbourethral gland, and prostate) add protective seminal plasma and activation factors, which promote further capacitation of the sperm to support fertilization. The prostate gland produces 30% of the ejaculate and is a common site of cancerous transformation in later life, development of which is thought to be androgen-dependent, 44 and thus is treated with anti-androgen therapy. 45 Accessory gland development and secretory abilities are androgen-dependent, 46 and the alteration or loss of steroid signaling in these tissues leads to suboptimal semen quality and impaired fertility. 47
Urethra and penis
The urethra connects the male urinary and reproductive tracts, serving as a continuation of the reproductive lumen leading from the accessory glands through the penis to the outside of the body. Surrounding the urethra is the penis, the male copulatory organ. The penis consists of muscular and connective tissue with large vascular sinuses that fill with blood during erection. 48 Testosterone is required for the formation of the penis during fetal development49,50 and for erectile function in adulthood. 51 A loss or decrease in testosterone signaling prevents enervation via the pelvic ganglia, reducing penile blood flow and diminishing penile smooth musculature, which leads to erectile dysfunction and substandard fertility. 52
Male reproductive endocrine signaling
Steroidogenic signaling in the male reproductive tract is essential for tissue development and reproductive function. Male reproductive vasculature consists of the testicular, prostatic, and penile arteries, and although it is not as interwoven and the female reproductive vasculature, the male has an extensive lymphatic system that facilitates endocrine communication. 53 It is also important to note that although the reproductive tissues are connected, the male gametes are shielded by specialized anatomical barriers – the blood-testis barrier and the blood–epididymis barrier – that prevent blood and lymphatic infiltration. 54 Tight gap junctions between cells prevent the free transfer of paracrine signaling factors and allow for a highly regulated luminal milieu in which gamete maturation can be orchestrated. Further, these gap junctions prevent autoimmune response directed against sperm and allow for evasion from cytotoxic agents. 55 Transporters are located along the blood–testis and blood–epididymal barriers and allow for necessary signaling to occur. 55 However, these barriers greatly complicate the delivery of therapeutic interventions to the male reproductive system. 56 The development of a microphysiological model of the male reproductive tract will allow us to examine mechanisms of transmission across these barriers and improve the design of drugs targeting the testis.
Unlike menopause in women, men do not normally undergo age-related changes in reproductive endocrine function. However, some men experience a decline in testosterone production as a result of illness, for example diabetes, 57 referred to as ‘low T.’ The number of men suffering from low T is rising as diabetes and other illnesses become more prevalent, 57 resulting in the need for novel testosterone therapies. However, prostate cancer is also a common morbidity in men and is often treated with androgen deprivation, as testosterone signaling is linked to disease formation and progression. Therefore, microphysiological modeling of the male reproductive tract would be highly beneficial for development of androgen altering therapies, to clarify how they might affect other tissues and potential disease states.
The value of a ‘repro-on-a-chip’ model
Microphysiological modeling of any anatomical system provides invaluable insight into physiological function, disease processes, and the potential toxicity and efficacy of therapeutic agents; however, modeling the reproductive system (‘repro-on-a-chip’) would be particularly informative, as many aspects of the human reproductive tract cannot be easily studied in patients. For example, preterm births are responsible for 75% of perinatal death, and the prevalence of preterm birth is rising, estimated to be approximately 12% of all US births. 58 Although conclusions about risk factors can be made in observational studies and using animal modeling, these studies cannot fully clarify the mechanism of preterm labor, nor do they provide a human correlate in which therapeutics can be safely examined. Disorders in implantation, ectopic pregnancy, and low T are other examples of pathological conditions that are difficult to study in human patients, but that could be readily modeled ex vivo in a repro-on-a-chip system to clarify the mechanisms underlying these disorders and identify potential therapeutic targets. A repro-on-a-chip system could also be used to elucidate the mechanism of action of postcoital or emergency contraceptive agents, thereby directly addressing the ethical controversy surrounding their impact on implantation. Although these models would not be appropriate for the study of pregnancy in its entirety, due to both ethical and technical challenges, aspects are readily testable, such as remodeling, cytokine production, and cellular function. Microphysiological modeling of the human reproductive tract is therefore an optimal approach to study not only normal physiologic processes, but also infertility and gynecological malignancy, and the effects of investigational therapeutics without the need for human trials or patient exposure. Finally, when integrated into a larger ‘human-on-a-chip’ model,’ repro-on-a-chip modeling could provide an ideal platform for the study of reproductive hormonal regulation of other anatomical systems.
Integrating the reproductive system into a human-on-a-chip model
The reproductive sex steroid hormones influence many other organ systems; therefore microphysiological models of these systems should incorporate the reproductive organs or the steroid hormones they produce. Unfortunately, in current pharmaceutical testing and animal studies, the default is to use male subjects to reduce experimental variability associated with ovarian hormone cycles.14,15 This methodology has resulted in many unintended morbidities in certain populations 5 and overall inefficiency in pharmaceutical development, as male-centric trial designs exclude cycling, pregnant, and menopausal women, as well as men with low T. Differences in drug efficacy and side effects are a result of sexually dimorphic physiology and the wide-reaching effects of reproductive steroid hormone signaling in several systems, including the respiratory, digestive, nervous, muscular, and skeletal systems (detailed in Table 1). Reproductive steroid hormones modulate normal function, impact disease progression, and alter pharmacokinetics of drugs in these tissues.
Hormonal regulation of the cardiovascular system
Cardiovascular disease is the leading cause of death in the United States, 59 and is being examined in several ‘organ-on-a-chip’ systems.6,60,61 However, reproductive signaling regulates certain cardiovascular functions and should be considered in these microphysiological models. Reproductive hormone signaling within the cardiovascular system can be both protective and detrimental, and leads to distinct sexual differences in cardiovascular health, development of disease, and strategies to mitigate disease. Estrogen is known to upregulate angiotensin, the major ligand of the renin–angiotensin system (RAS) 62 ; however, estrogen simultaneously downregulates both angiotensin receptor and angiotensin-converting enzyme. 63 The overall effect appears to be beneficial, leading to a reduction in RAS signaling and blood pressure that results in improved health of endothelial cells lining the vascular network. 64 Estrogen also reduces certain inflammatory responses, particularly release of interleukin (IL)-1, -6, and -8 and C-reactive protein, which are involved in the formation of atherosclerotic plaques. 65 In men, testosterone increases renin activity, which can be detrimental to vascular tone, 64 and amplifies oxidative stress, which causes endothelial cell dysfunction and apoptosis. 66
Sexually dimorphic variation in cardiovascular disease
Reproductive steroid hormones modulate normal cardiovascular physiology and also play a role in the sexually dimorphic development of cardiovascular disease. Estrogen signaling reduces the risk of heart disease in premenopausal women 67 through various mechanisms, including enhanced vasodilation 68 and the prevention of atherosclerotic plaque formation. 69 However, as previously discussed, the levels of estrogen decrease with the onset of menopause, and the protective effect of estrogenic signaling is lost, 70 the risk of cardiovascular disease is enhanced and it surpasses the risk seen in age-matched men. 71
Recent work has found that the anatomical localization of coronary artery disease is different in men and women. In men, atherosclerotic plaques often build up in main coronary vessels over time. The formation of occlusive plaques likely accounts for the more pronounced heart attack symptoms seen in men, including sharp chest pains. Symptoms of coronary heart disease in women are not as evident and are described as episodes of fatigue and shortness of breath. 72 These alternative symptoms likely reflect the formation of small, longitudinal plaques in the peripheral microvasculature and in the smaller vessels that supply the heart. 73 Rather than forming plaques that occlude arteries, postmenopausal women develop lipid and calcium accumulation around vessels that causes them to stiffen 74 ; this prevents smaller vessels from vasodilating in times of vascular need, such as during exercise or stress, leading to cardiac ischemia and heart failure. Diagnostic tests of coronary heart disease based on symptom presentation were developed primarily based on observations in males, and female symptoms often go undetected, leading to an increase in mortality. 75 In fact, coronary heart disease is the leading cause of death in women, accounting for nearly one in four deaths. 59 Therefore model systems for pharmaceutical discovery of new drugs need to incorporate reproductive sex hormones in order to address these sexually-dimorphic processes in biology that impacts disease and then use these models to identify targets for sex-specific drug development.
Hormonal effects on the cardiovascular system
To address the devastating effects of cardiovascular disease in postmenopausal women, the women’s health initiative (WHI) was launched to examine if hormone replacement therapy (HRT) in postmenopausal women could prevent cardiovascular disease. Counterintuitively, the WHI results suggested that estrogen therapy in postmenopausal women was detrimental to cardiovascular health 76 ; however, reexamination of the data found that the timing of HRT therapy is a factor. 77 HRT initiated prior to onset or during perimenopause appears to be protective against cardiovascular disease, whereas HRT in women more than 10 years after the start of menopause has no benefit or may increase venothrombolic events. The switch in HRT efficacy is thought to be due to arterial stiffening soon after menopause in the immediate absence of estrogen, thus women who did not receive HRT early after menopause were predisposed to cardiovascular events. 77
Hormone therapy is also being examined for men for the reduction of cardiovascular disease risk and other morbidities. Although testosterone is thought to have adverse effects on cardiovascular health, evidence demonstrates that men with cardiovascular disease often have lower serum testosterone levels, even when adjusted for age, BMI, and metabolic status.78,79 The association between cardiovascular disease and lowered androgen levels appears to be due to fewer circulating endothelial progenitor cells, 80 which aid in maintaining the vasculature. Further, testosterone may act as a vasodilator, which would improve blood flow. 81 However, as with women, timing and risk analysis of potential comorbidities appears to be crucial for HRT therapy in men, with conflicting evidence of testosterone efficacy determined at different ages.82,83 These studies suggest ‘low T’ and menopause, both times of dynamic change in reproductive hormone signaling, are also periods in which new therapies for cardiovascular heath are needed. Microphysiological modeling of the cardiovascular system in the presence and absence of normal gonadal steroidogenesis would help to decipher the mechanism by which hormones modulate the development of cardiovascular disease and how this changes with age.
Sex-specific modulation of hepatic function
Reproductive hormone signaling is also essential for hepatic function, which governs drug clearance and bioactivation of new pharmaceutical agents. Steroid hormone influence on drug pharmacokinetics and liver metabolism is a primary justification for integration of hepatic and reproductive tract microphysiological models as part of a larger ‘human-on-a-chip’ model. Recent studies in the mouse demonstrate that more than 1000 genes display sexually dimorphic regulation in the liver. 84 This vast change in expression is partially attributed to the predominate expression in females of CYP3A4, a member of the cytochrome P450 enzyme family that is important for drug metabolism and the synthesis of cholesterol. 85 Expression of CYP3A4 is regulated by both progesterone and estrogen. 86 Additionally, growth hormone (GH) influences hepatic function, and while GH is produced by both sexes; in men it is pulsatile, whereas in women GH secretion is almost continuous. 87 As with CYP3A4, GH levels are also modulated by reproductive steroid hormone signaling, namely testosterone and estrogen. 88
Sex steroid effects in hepatic disease
One of the most common diseases of the hepatic system is alcoholic liver disease, which can lead to the development of cirrhosis and liver failure. There is strong evidence that women are more prone to deleterious hepatic effects of alcohol abuse. 89 In heavy drinkers, the risk of developing alcoholic liver disease and subsequent cirrhosis was doubled in women compared to men exposed to the same amounts of alcohol. 89 Although this may be due in part to women often having a lower body weight and thus a higher systemic percentage of alcohol, these changes alone do not account for the disparity in the disease development. 90 Two possible reasons for differences in the development of alcoholic liver disease have been proposed: a lower production of the metabolic enzyme alcohol dehydrogenase in women 90 and estrogen sensitization of hepatic Kupffer cells, which in turn cause hypoxia–reoxygenation injury. 91 Despite having reduced damage as a result of alcohol abuse, men are 3–5 times more likely to develop hepatocellular carcinoma, 89 which is in part due to the lack of protective estrogen signaling that prevents the inflammatory progression of hepatitis into carcinoma in women. Estrogen downregulates Kupffer cell production of a pivotal proinflammatory cytokine, IL-6, thereby reducing risk of developing hepatocellular carcinoma in a sex-specific manner. 92 Further, there is evidence FoxA1 and 2, which are influenced by both estrogen and androgen signaling, play a role in the sexually dimorphic formation of hepatic cancer. 93 Development of a microphysiological model recapitulating both reproductive and hepatic tissues would clarify the role of steroid hormone regulation of Kupffer cells.
Reproductive hormone-dependent pharmacokinetics in the liver
As discussed, several cytochrome P450 enzymes, namely CYP3A4, are differentially expressed between the sexes, which affects hepatic metabolism of various molecules, including pharmacologic agents. 94 These variations in enzymatic concentration affect key metabolic processes including oxidation, reduction, and hydrolysis, resulting in differential activity and breakdown of pharmaceuticals. Difference in P450 enzyme expression and activity is partially regulated by fluctuations in steroid hormone signaling. P450 expression is altered during the menstrual cycle, pre- and postmenopause, 95 and in response to changes in testosterone levels in men. 96 Additionally, hepatic transporters, such as MDR1 (hepatic p-glycoprotein), are increased in men, which accelerate drug clearance rates. 97 Altered hormone signaling during pregnancy can also greatly influence the pharmacokinetics of drug absorption and metabolism, which can easily lead to adverse effects. 98 These studies suggest a need to model P450 enzyme expression and ability to metabolize drugs – and the consequences on drug efficacy and toxicity – more effectively across different patient groups.
Drugs that may initially target the liver can also impact reproductive signaling and function. For example, statins are a widely used class of drugs that inhibit cholesterol production (which primarily occurs in the liver). However, by lowering cholesterol, which is a precursor to steroid hormone production, statins also impact reproductive function, as evidenced by incidence of erectile dysfunction in men 99 and abnormal development of the placenta in pregnancy in women 100 who are taking statins. Although, statins are not recommended for use during pregnancy, ∼50% of pregnancies are unplanned, making the likelihood of exposure high, especially in the first trimester. 101 Accidental fetal exposure and effects on placental formation are also a concern for other therapeutic agents used to treat chronic conditions such as epilepsy, 102 arthritis, 103 and depression. 104 Further, many women take vitamin supplements during pregnancy, but how these factors affect the pregnancy and what metabolites reach the fetus remains unclear. Better models of drug action in pregnant women are necessary to avoid toxicities reported previously in maternal use of drugs such as diethylstilbestrol, thalidomide, and many others. 5
Significance of microphysiological modeling of the reproductive tract and incorporation into human-on-a-chip models
As highlighted in this review, reproductive function and health is dependent on the coordination of multiple reproductive organs, which share a common microenvironment and several key signaling components, primarily the sex steroids. Reproductive cells and tissues are often studied in isolation, which has led to important discoveries in specific reproductive mechanisms, but does not allow for the study of the system as a whole or the effects of a disease or drug. New models and techniques are needed that recapitulate the complexity of the reproductive system ex vivo and the changes that occur in this system throughout an individual’s lifespan. For these reasons, developing ‘repro-on-a-chip’ models, either alone or as part of a ‘human-on-a-chip’ system, is a necessary step forward in our ability to examine reproductive function as a whole, and to measure and analyze downstream effects of changes in the system, whether they be age-related, disease-related, or drug-induced changes. 3 Such a system would be highly beneficial, as vulnerable populations that are not often examined in clinical trials, 105 are also those that experience dynamic differences in reproductive signaling. Some of the most vulnerable populations include children (prepubertal), pregnant women, and the elderly (menopause and ‘low T’), who each have unique hormone profiles. The physiological microenvironment in these patients is often vastly different from the middle-aged men that are typically used to examine drug efficacy and safety; the resulting lack of data in specific populations can lead to adverse outcomes. 105 In women in particular, unforeseen side effects of drug exposure can be long-lasting or even permanent, affecting fertility 106 and fetal health. 5 Additionally, these models can be utilized to examine the systemic effects of reproductive signaling disruption, in the cases of prostatectomy, oophorectomy, or hysterectomy, which have widespread effects on normal physiological function and potentially on therapeutic efficacy and toxicity.
There are many challenges to address when creating microphysiological models, such as finding a common media that provides the appropriate nourishment for each organ in the system, designing accurate microfluidics between organs, and providing the necessary substrate(s) for the production of signaling molecules. 1 Although progress has been made in recapitulating individual reproductive organs,107,108 there are currently no models of the reproductive system as a whole. However, technology developed for modeling other anatomical systems could be modified for development of a repro-on-a-chip system. For example, uterine and fallopian contractility could be mimicked utilizing technology designed to examine heart and smooth muscle contraction, 109 fallopian cilia function could be modeled after cilia systems developed for the study of lung function 110 and cervical barrier function could be studied utilizing epithelial models made for dermal research. 111 Further, reproductive organs could be connected and grown in vitro utilizing microphysological devices which allow for necessary tissue crosstalk, including bioreactors or micropumps utilized for the study of hepatic co-cultures 112 or porous microchannel devices developed for gut-on-a-chip models. 113
Producing ‘organs-on-a-chip’ and ‘humans-on-a-chip’ which are physiologically accurate and function in a reproducible, viable manner is no small task, but will yield significant rewards for the research community. Not only will these models provide a safer, more accessible way to examine vulnerable populations that cannot (or are not) included in traditional clinical trials, but development and testing of novel therapeutics can also be completed in a more efficient, cost-effective manner. ‘Human-on-a-chip’ provides a platform for investigating the toxicities and pharmacokinetics of new therapeutics using human tissues long before the agents are used in clinical trials. Perhaps early testing in human tissue-based models would prevent some of the obstacles currently encountered when using primarily cell-based and animal models. 114 There is a high rate of failure in the translation of preclinical data into new drug therapies, with an estimated cost of ∼$802 million per approved drug. 115 Moreover, 8 of 10 drugs removed after market are due to adverse events in women. 116 By testing these drugs in microphysiological models that can recapitulate sex-specific physiologies earlier in the drug testing process, less time and money would be spent on development of toxic or ineffective therapeutics.
Additionally, microphysiological systems can be modified to mimic disease, and thus can be utilized to examine specific aspects of a disorder isolated from the full disease state. This is particularly important in cases of comorbidity, when the side effects of a drug cannot always be easily delineated from symptoms of co-existing disease. Conversely, diseases that often occur together can be modeled simultaneously, as can the effects of co-administration of multiple therapies. This would be especially useful in elderly patients who take several therapies, often have several comorbidities, and have altered reproductive signaling, making them vulnerable to toxicity.
Unfortunately, current cell-based and animal models of the reproductive tract and other anatomical systems poorly recapitulate the human system, which leads to inaccuracies in preclinical data and inefficiencies in drug development. Furthermore, clinical testing is performed primarily in men, which overlooks the effects of sex-based differences in physiology and disease progression that can affect drug efficacy and safety. Data are also scarce for specific populations in which drug testing is not feasible, such as pregnant women. Microphysiological models of the reproductive tract are being developed to overcome these gaps and improve the ability to test new drugs on human tissues earlier in the development process. The reproductive steroid hormones are known to have systemic effects on various anatomical systems (Table 1), which are linked through endocrine hormone crosstalk beyond that discussed in this review. Therefore microphysiological models that recapitulate the reproductive milieu and model systems that incorporate the reproductive tract and its endocrine communication with the rest of the body are necessary tools that will benefit the biomedical and pharmaceutical fields. These models will provide platforms for studying normal physiology and disease mechanisms, as well as for more effective drug development, which will result in better outcomes for patients.
Footnotes
Author contributions
SLE, JJK, TKW, and JEB prepared the manuscript. SLE drafted figures.
Acknowledgments
We thank Stacey Chapman Tobin for critical reading of the manuscript.
This review was funded by NIH (UH2TR000498-01).