
Abstract
Podocyte foot process (FP) is dysregulated in nephrotic syndrome. The effacement of podocyte FPs typically arises following perturbations in the actin cytoskeleton. Recent data suggest that the effects of calcineurin (CaN) inhibitor cyclosporine A (CsA) are independent of its effects on T-cells, and CsA has been identified as stabilizing the actin cytoskeleton through stabilizing synaptopodin in podocytes, and thereby directly reducing proteinuria. Other studies showed that CsA could regulate cofilin-1 directly within tubular epithelial cells. However, whether synaptopodin is the only target of CsA or whether the antiproteinuric role of CsA is played by regulating cofilin-1 in podocytes has not been studied. In the present study, changes in the expression and distribution of nephrin, synaptopodin, cofilin-1 and phosphorylated cofilin-1 (pho-cofilin-1) were detected in both puromycin aminonucleoside (PAN) induced nephrotic rats treated with CsA and cultured podocytes exposed to PAN with/without CsA. Cofilin-1, synaptopodin mRNA was knocked down or combined by siRNA to investigate whether cofilin-1 was critical for the protective effect of CsA and whether the effect of CsA on cofilin-1 was independent of its effect on synaptopodin. We found that CsA reduced proteinuria and repaired FP effacement of PAN-induced nephropathy, restored expression of nephrin, synaptopodin, cofilin-1, pho-cofilin-1 both in vivo and in vitro. CsA also repaired actin cytoskeleton impaired by PAN in vitro. The protective effect of CsA was diminished when cofilin-1 was knocked down compared to negative control. Synaptopodin knocked down had no effect on cofilin-1. The protective effect of CsA decreased significantly when cofilin-1 and synaptopodin were simultaneously knocked down compared to only cofilin-1 knock down. In conclusion, the antiproteinuric effect of CsA is derived from the stabilization of the podocyte actin cytoskeleton by upregulating expression of cofilin-1, which was independent of its effect on synaptopodin.
Introduction
Since its introduction in the 1980s as an immunosuppressive agent, cyclosporine A (CsA) has been widely used to prevent allograft rejection and dramatically improve outcomes of organ transplantation. CsA is a CaN inhibitor that mediates T-cell receptor signaling and suppresses inflammatory cytokine expression, and inhibits leukocyte migration. It is also a potent inhibitor of mitochondrial permeability protecting cells from death. For more than three decades, CsA has been widely used in the treatment of proteinuric diseases. The treatment of nephrotic syndrome with CsA was originally based on the assumption that the disease is immune mediated. Thus, the immunosuppressive effects of such drugs are likely to be helpful. Faul et al. made the novel observation that the antiproteinuric effects of CsA can be explained by its direct effects on the podocyte actin cytoskeleton and are independent of its effects on T lymphocytes. 1 The mechanism involves synaptopodin, a key stabilizer of the actin cytoskeleton in podocytes. Faul et al. found that CsA prevents degradation of synaptopodin, stabilizes the actin cytoskeleton, and protects against proteinuria. Faul’s study highlights a novel mechanism of CsA in treating nephrotic syndrome, which is the local effect of CsA within the podocyte. The effect of CsA in podocytes is also related to podocyte cytoskeleton, synaptopodin.
Podocyte foot process (FP) effacement is a common pathological phenomenon of proteinuric glomerular diseases and is usually a dynamic and reversible process. Cytoskeleton actin filaments are the structural backbone component of podocyte FPs. Protein complexes that regulating or stabilizing the actin cytoskeleton are therefore essential for the maintenance of podocyte FP structure as well as the intact glomerular filtration barrier. The dynamics of actin filament (F-actin) assembly/disassembly and organization in cells are regulated mainly by two actin-binding proteins, Actin Depolymerizing Factor (ADF)/cofilins and the Arp2/3 complex.2,3
ADF/cofilins are ubiquitous among eukaryotes and are essential proteins responsible for the turnover and reorganization of actin filaments. 4 The actin-binding protein cofilin-1 is expressed in podocytes and is necessary for maintaining normal podocyte architecture. Cofilin-1 as a cytoskeleton molecule is also involved in the induction and recovery of podocyte injury. 5 Ashworth et al. also found that cofilin-1 is an essential regulator for actin filament recycling, being required for the dynamic changes of podocyte FPs. 6 Interestingly, Martin-Martin et al. recently revealed that CsA could regulate cofilin-1 directly within tubular epithelial cells. They found that CsA can phosphorylate cofilin-1 and promote stress fiber generation in tubular epithelial cells. 7
Taking all these previous studies together, we proposed that CsA plays an antiproteinuric role in podocytes not only via stabilizing synaptopodin but also via regulating another cytoskeleton molecule cofilin-1. Cofilin-1 might be a crucial molecular target for CsA in maintenance of cytoskeleton stabilization and FP structure of podocytes. In this study, we found that the antiproteinuric and cytoskeleton-stabilizing effects of CsA were demonstrated directly by the upregulation of actin-binding protein cofilin-1.
Materials and methods
Animal model
Male Sprague-Dawley rats (n = 15, 120–140 g) were purchased from the Experimental Animal Center at Peking University Health Science Center and divided into three groups. The first group (n = 5) was treated with normal saline, the second group (n = 5) received a single intraperitoneal injection of puromycin aminonucleoside (PAN) (15 mg/100 g, Sigma-Aldrich, USA), as previously described. 8 The third group received PAN + CsA (Sandimmum; Sandoz Pharma, Basel, Switzerland).9,10 The exact procedure was that rats received CsA 5 mg/kg intraperitoneally daily for 11 days and received a single intraperitoneal injection of PAN on the second day. All animals were sacrificed at day 10 after PAN treatment. The animal studies were approved by the Animal Research Review Board of Peking University.
Urine collection and tissue preparation
Twenty-four hour urine was collected at day 10 for measuring urinary protein on an automatic biochemical analyzer (7170A, Hitachi, Japan) using a pyrogallol red-molybdate dye-binding method. All rats were sacrificed on day 10 and the kidneys were removed. One kidney was used for isolating glomeruli using the conventional sieving method. 11 The renal cortex of the other kidney was divided into four parts, one fixed in 3% glutaraldehyde for transmission electron microscopy, one embedded in opti-mum cutting temperature (O.C.T.) compound (Sakura, USA) for immunofluorescence, and two stored at −80℃ for protein isolation.
Transmission electron microscopy
The renal cortex kept in 3% glutaraldehyde was further fixed in 1% osmium tetroxide, followed by dehydration in graded ethanol and washing in acetone, and finally embedding in Epon 812. Ultrathin sections were stained with uranyl acetate and lead citrate and examined with a transmission electron microscope (JEM-1230, JEOL, Japan).
Cell culture and drug treatment
Immortalized human podocytes (AB8/13, gift from Moin Saleem, Bristol) were grown in RPMI-1640 cell medium containing 10% Fetal Calf Serum (FBS) (Gibco, USA), 1% penicillin/streptomycin (Invitrogen, California, USA), and insulin–transferrin–sodium selenite at 33℃ with 5% CO2 as previously described. 12 Murine podocytes (MPC5, a gift from Peter Mundel, USA) were cultured at 33℃ in RPMI-1640 containing 10% FBS, 1% of 100 units/mL penicillin/streptomycin, and 10 units/mL of γ-interferon at 33℃ with 5% CO2. Both podocyte cell lines were grown in a flask and incubated at 37℃ with 5% CO2 for a minimum of 10 days to differentiate. Cells were then plated at a density of 6 × 104 cells/well in six-well plates for western blot and plated 4 × 104 cells/well in six-well plates with glass coverslips for immunofluorescence staining. All experiments were undertaken at 60% confluence. Podocytes were pretreated with CsA (0.5 µg/mL) for 1 h before exposure to PAN for 24 h.
siRNA transfection
Podocytes were transfected with cofilin-1 siRNA (Santa Cruz, USA), and synaptopodin siRNA (Guangzhou RiboBio Co., China), using Lipofectamine 2000 Reagent (Invitrogen, USA) as per the manufacturer’s instructions. After transfection for 24–48 h, the podocytes were exposed to CsA and PAN as mentioned earlier.
Quantitative PCR analysis
Two micrograms of RNA extracted from cultivated human or murine podocytes with Trizol (Invitrogen, USA) were reverse transcribed to cDNA for quantitative real-time Polymerase Chain Reaction (PCR) with TransStartTM Green qPCRSuperMix (TransGen Biotech, China). PCR reactions were performed on a CFX CONNECT system (BIO-RAD, USA) at 95℃ for 10 s, followed by 35 cycles of 95℃ for 5 s, 61℃ for 31 s, with a dissociation stage at 95℃ for 15 s, 60℃ for 1 min, and then 95℃ for 5 s. All primers for the listed transcripts were designed using Primer 3 software: human-cofilin-1 left: TCAAGATGCTGCCAGATAAG, human-cofilin-1 right: TTGGAGCTGGCATAAATCAT, murine-cofilin-1 left: CAAGGATGCCATCAAGAA, murine-cofilin-1 right: GTCCTTGACCTCCTCGTA, human-synaptopodin left: GGACAGTTTGGGTTTGGGAC, human-synaptopodin right: TCATCTTCACAGCAGCAGGA, glyceraldehyde-3-phosphate dehydrogenase (GAPDH) left: AAACCCATCACCATCTTCCA, GAPDH right: GTGGTTCACACCCATCACAA.
Protein extraction and western blot procedure
The isolated glomeruli and podocytes were lysed with a Radio Immunoprecipitation Assay (RIPA) buffer containing protease inhibitors (1 mM phenylmethylsulfonyl fluoride, 1 µg/mL leupeptin and pepstatin). Thirty micrograms of the total tissue protein and 20 mg of total cell protein were subjected to 8–15% Sodium Dodecyl Sulfonate-Polyacrylamide gelelectrophoresis (SDS-PAGE) and transferred to nitrocellulose membranes (Amersham Life Science). After blocking with a 5% low-fat milk powder, the membranes were incubated with antibodies against rabbit anti-cofilin-1 (1 : 2000, Cell Signaling, USA), phosphorylated cofilin-1 (pho-cofilin-1) (1 : 2000, Cell Signaling, USA), nephrin (1 : 2000, gift from Karl Tryggvason, Sweden), synaptopodin (1 : 1000, Abcam, USA) antibody, and mouse anti-GAPDH (1 : 5000, Chemicon USA) antibody at 4℃ overnight (14–16 h). Subsequently, the membranes were rinsed four times for 5 min in Phosphate Buffered Saline (PBS) buffer with 0.05% Tween-20 and incubated with horseradish peroxidase-conjugated anti-rabbit (1 : 5000, ImmunoChemistry, USA) or mouse IgG (1 : 5000, Santa Cruz, USA) for 1 h at room temperature. After a final washing, the membranes were developed using an enhanced chemiluminescence reagent (Millipore, USA), and the specific protein bands were scanned and quantitated in relation to GAPDH.
Confocal immunofluorescence
Five micrometer cryosections were fixed in ice-cold acetone, subsequently permeabilized and blocked with 0.3% Triton X-100 and 10% goat serum. The following primary antibodies were used: rabbit anti-cofilin-1 (1 : 200, Abcam, USA), pho-cofilin-1 (1 : 200, Cell Signaling, USA), mouse antisynaptopodin (1 : 100, USBiological, USA). After washing three times, the slides were incubated with Fluorescein Isothiocyanate (FITC) goat anti-rabbit IgG (1 : 200) and Tetramethyl Rhodamine Isothiocyanate (TRITC) goat anti-mouse IgG (1 : 200). For cells, the coverslips were fixed with 4% paraformaldehyde, followed by permeabilization and blocking with 0.3% Triton X-100 and 5% bovine serum albumin. Primary and secondary antibodies, TRITC-Phalloidin (Invitrogen, USA) and Hoechst nuclear dye (Sigma-Aldrich, USA) were applied. The slides were mounted with 50% glycerol. Stained images for each antibody at the same light exposure were obtained by confocal laser-scanning microscopy (Zeiss Lsm510 Meta, Germany). Photographs of glomeruli and podocytes stained with cofilin-1 were selected randomly and analyzed by a person who was blinded to the study groups.
Statistical analysis
Statistical analysis was performed with Graphpad Prism 5 software (PRISM). Results were presented as means ± SD. Comparisons were performed by TTEST, with statistical significance set at a P value of <0.05.
Results
CsA reduced proteinuria level and restored FPs effacement
Proteinuria increased sharply in PAN-injected rats versus controls by day 10 (387.2 ± 32.3 mg/24 h versus 9.5 ± 2.8 mg/24 h, P < 0.01), and treatment with CsA significantly attenuated proteinuria (387.2 ± 32.3 mg/24 h versus 142.8 ± 15.5 mg/24 h, P < 0.01) (as shown in Figure 1, panel a). FPs of normal rats were long and thin (as shown in Figure 1, panel b), while 10 days after PAN injections, podocytes presented with a loss of FPs and diffuse, wide spread fusion (as shown in Figure 1, panel c). When treated with CsA typical FPs were observed again (as shown in Figure 1, panel d). These data showed that CsA protected podocyte from PAN injury.
Twenty-four hour urinary protein and ultrastructural changes of podocyte foot processes in rat model. (a) Compared to the control groups, proteinuria significantly increased at day 10 after PAN injection. Proteinuria level decreased significantly with the treatment of CsA. Data are presented as mean ± SD. N = 5. *P < 0.01. (b) The foot processes were long and thin in normal rats. (c) Ten days after PAN injection, foot processes were lost and present with diffuse and widespread fusion. (d) When treated with CsA, typical foot processes were observed again. Bar = 2 µm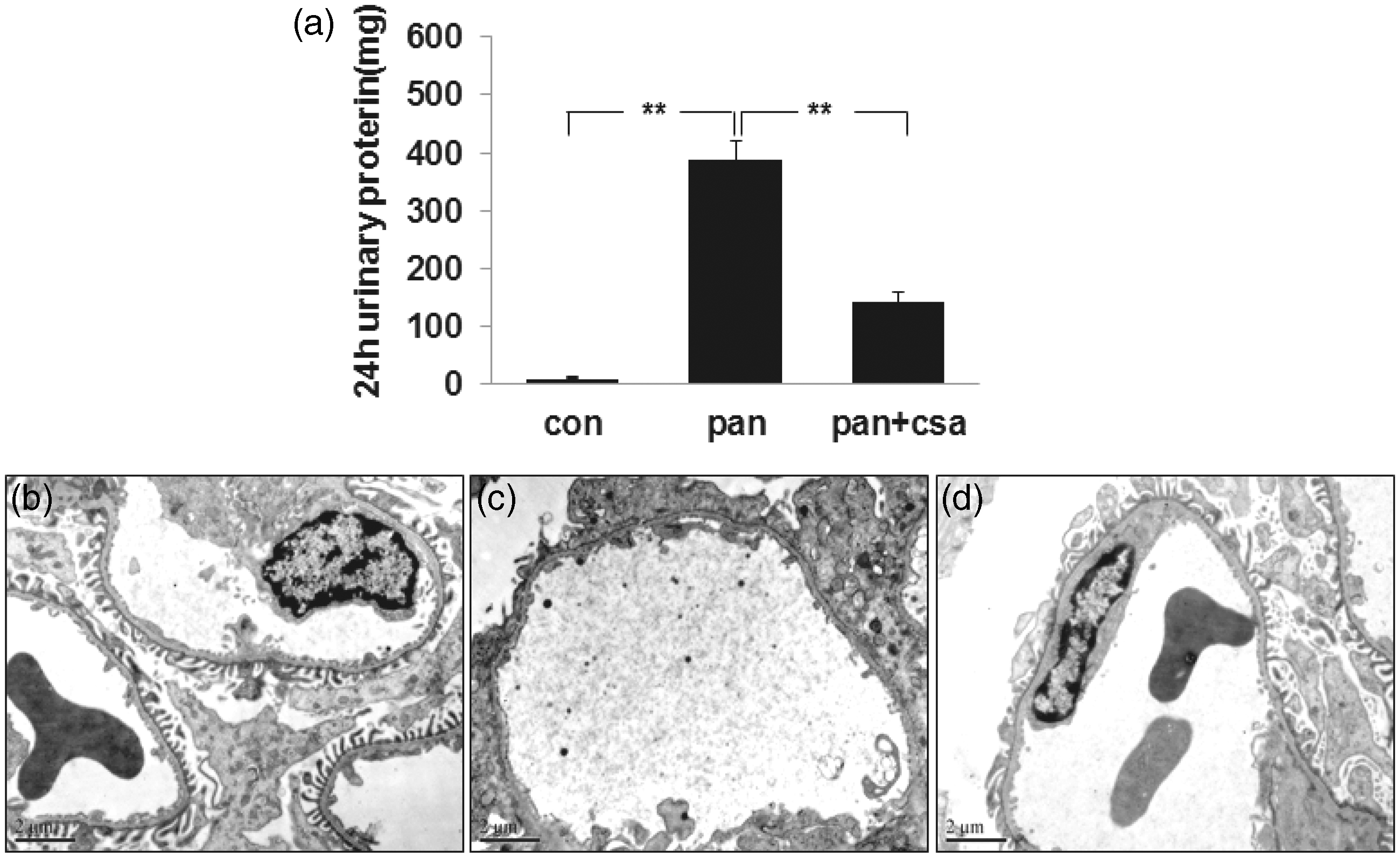
CsA restored both expression and distribution of cofilin-1 in rat glomeruli
We quantitated podocyte injury by assessing the expression of nephrin, a podocyte specific molecule that acts as a podocyte injury marker. The specific protein bands for nephrin were detected at 185 kDa by western blot, and this decreased markedly by day 10 in PAN-injected rats compared with controls (as shown in Figure 2, the first band of panels a and b). Meanwhile, another podocyte specific molecule synaptopodin decreased significantly by day 10 in PAN-injected rats compared with controls (as shown in Figure 2, the second band of panel a and c). However, both nephrin and synaptopodin levels were restored in CsA treatment rats. The expression of cofilin-1 at the size of 19 kDa was decreased significantly by day 10 after PAN injection compared with controls, and treatment with CsA restored its expression significantly (as shown in Figure 2, the third band of panels a and d). The relative expression of pho-cofilin-1 increased significantly by day 10 after PAN injection compared to control, and CsA partially restored its expression (as shown in Figure 2, the fourth band of panels a and e).
The expression level of nephrin, synaptopodin, cofilin-1, and pho-cofilin-1 in isolated glomeruli of rat nephrotic models. (a) Western blot analysis of nephrin, synaptopodin, cofilin-1, and pho-cofilin-1 in isolated glomeruli. Calibrated with GAPDH, the expression of nephrin (b), synaptopodin (c), cofilin-1 (d) decreased significantly by day 10 after PAN injection compared to the control. The expression of nephrin, synaptopodin, and cofilin-1 was restored with the treatment of CsA compared to PAN groups. (e) Calibrated with cofilin-1, the expression of pho-cofilin-1 was increased significantly by day 10 after PAN injection compared to control group, and CsA restored its expression partially. Data are presented as mean ± SD. N = 5. *P < 0.05, **P < 0.01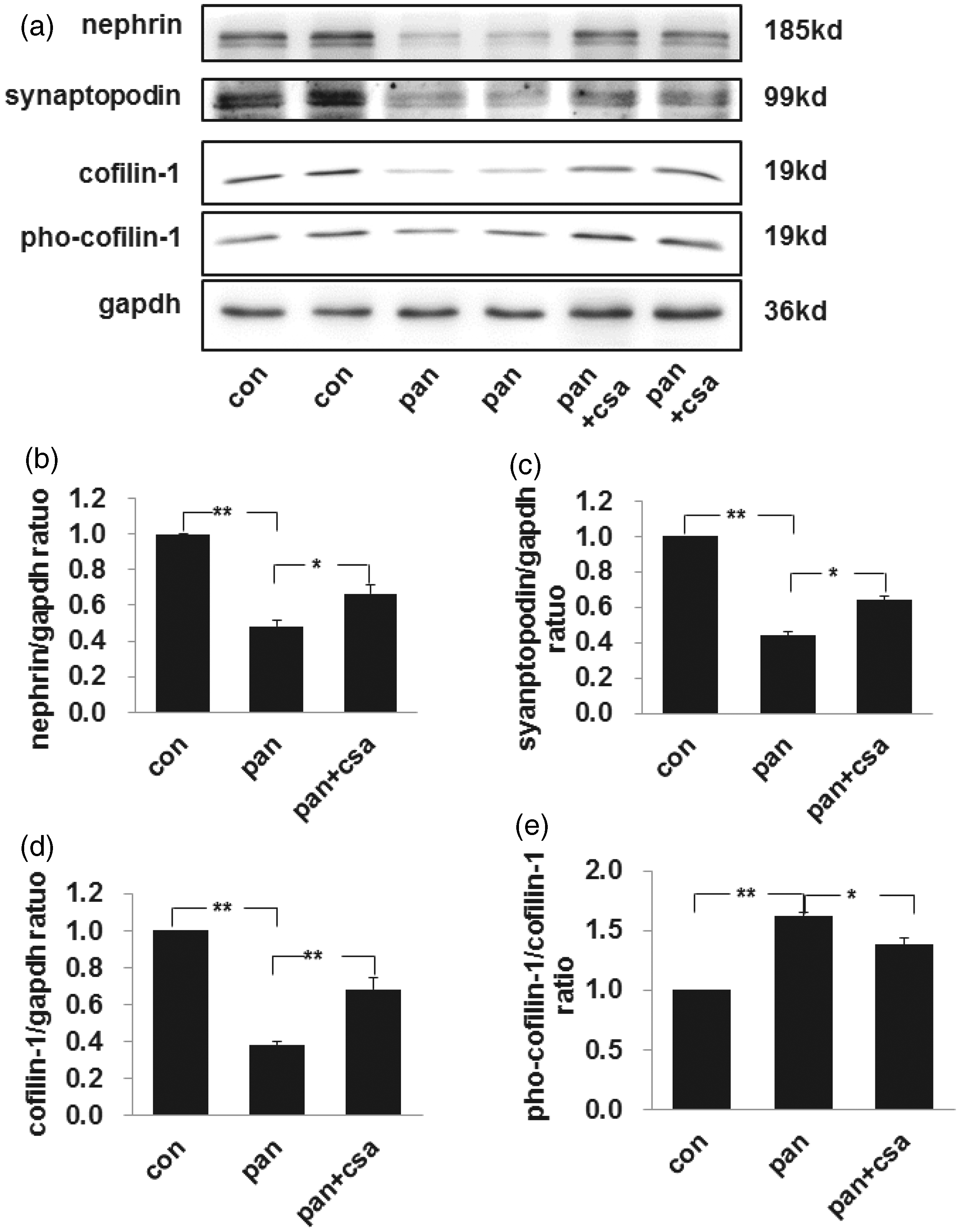
Cofilin-1 is known as one of the regulators of actin filament, and the function of cofilin-1 distributing in cytoplasm or nucleus was different. In the next experiments, we assessed the expression and distribution of cofilin-1 and pho-cofilin-1 by immunofluorescence. We found that the expression of cofilin-1 decreased significantly by day 10 after PAN injection compared with controls, and treatment with CsA significantly restored its expression, whereas no significant changes of distribution were found (as shown in Figure 3, panels a1 to a3). The expression of pho-cofilin-1 was weak in the glomeruli of control rats, but 10 days after PAN injection, the pho-cofilin-1 expression increased significantly, and treatment with CsA significantly restored its expression (as shown in Figure 3, panels d1 to d3). Synaptopodin, another actin cytoskeleton, which is considered as a podocyte-specific marker, significantly decreased by day 10 in PAN-injected rats compared with controls. Treatment with CsA significantly restored its expression (as shown in Figure 3, panels b1 to b3, e1 to e3). Double-labeling assays showed that both cofilin-1 and pho-cofilin-1 colocalized well with synaptopodin (as shown in Figure 3, panels c1 to c3, f1 to f3). These observations indicated that cofilin-1 was involved in PAN-induced podocyte injury and the beneficial effect of CsA. We next explored potential mechanisms of cofilin-1 in both podocyte injury and CsA therapeutic effect in vitro.
Immunofluorescence staining of cofilin-1, pho-cofilin-1, and synaptopodin in rats. Cofilin-1 was labeled green, its staining intensity was decreased in the glomeruli of PAN-injected rats (a2) as compared to control group (a1), treatment with CsA restored cofilin-1 expression (a3) compared to PAN-injected rats, and treatment with CsA restored cofilin-1 expression (b3) compared to PAN-injected rats. Pho-cofilin-1 was labeled green and its staining intensity was weak in the glomeruli of control rats (d1). Ten days after PAN injection, the pho-cofilin-1 staining intensity was increased (d2), treatment with CsA restored its expression partially (d3). Synaptopodin, another podocyte marker, was labeled red, which displayed a linear pattern along the glomerular capillary loops. Its staining was decreased in the glomeruli of PAN-injected rats (b2, e2) compared to control groups (b1, e1), and treatment with CsA restored its expression (e3). Double-labeling assays showed that both cofilin-1 and pho-cofilin-1 colocalized with synaptopodin (c1–c3, f1–f3). Bar = 20 µm. (A color version of this figure is available in the online journal.)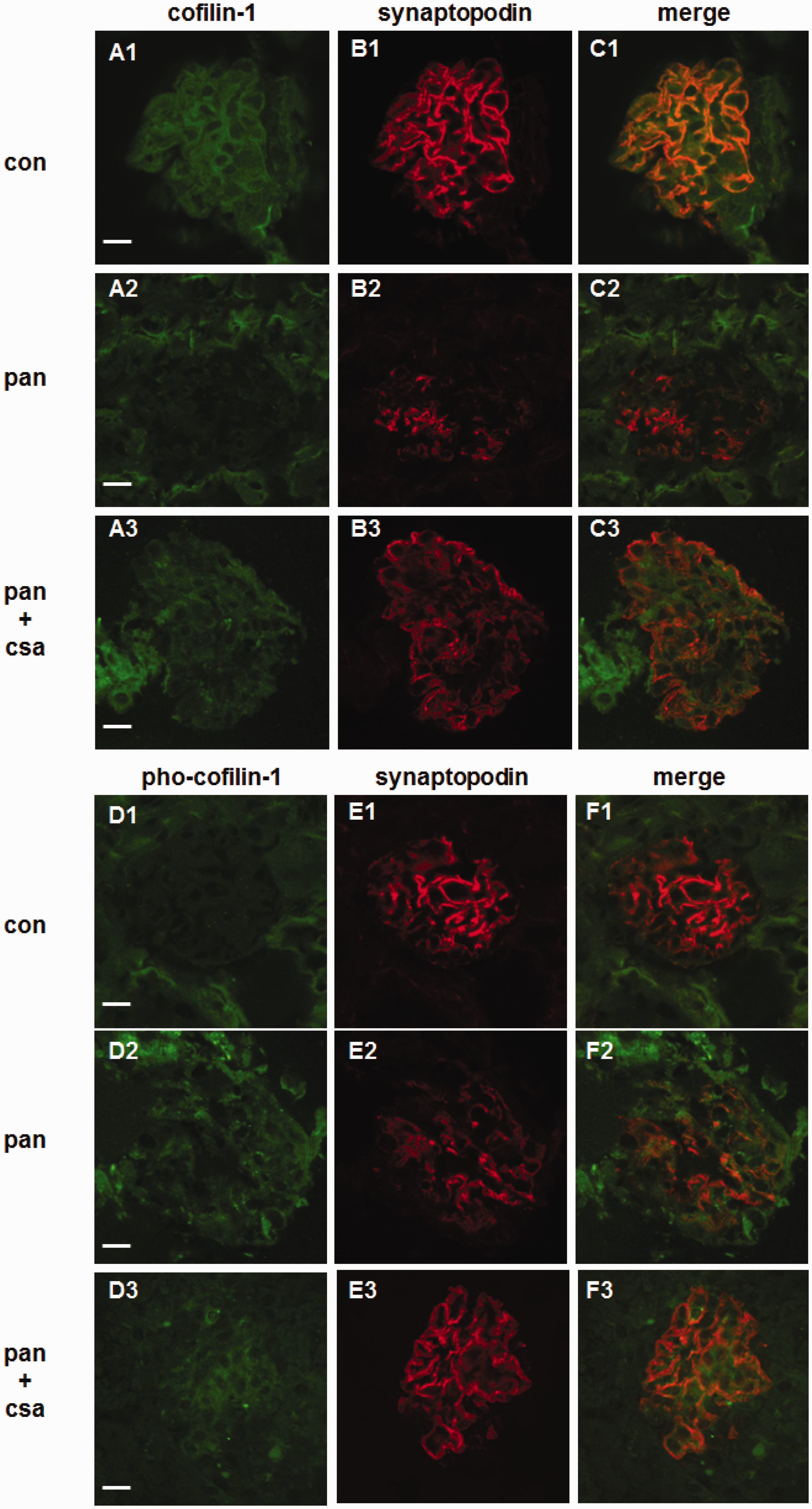
PAN downregulated the expression of cofilin-1 in dose-dependent manner in vitro
The expression of cofilin-1, nephrin, as well as synaptopodin decreased significantly after PAN treatment for 24 h, in a dose-dependent way, while the expression of pho-cofilin-1 increased (as shown in Figure 4, panels a to e). The same phenomenon was seen in murine podocytes (as shown in Figure 4, panels f to j). The more seriously podocyte injured, the more lower cofilin-1 expression declined. Our results also showed that the relative expression of pho-cofilin-1 increased significantly after PAN treatment for 24 h, also in a dose-dependent manner. Our data suggested that cofilin-1 was involved in the podocyte injury.
The changes of nephrin, synaptopodin, and cofilin-1 and pho-cofilin-1 expression when cultured podocytes (both human and murine) were exposed to PAN in a dose-dependent manner. Cells were treated with different dose of PAN for 24 h. Western blot analysis showed that nephrin, synaptopodin, and cofilin-1 expression decreased in a dose-dependent manner of PAN both in human podocytes (a–d) and murine podocytes (f–i) when values were calibrated with GAPDH. The expression of pho-cofilin-1 increased in a dose-dependent manner of PAN both in human podocytes (e) and murine podocytes (j), when values were calibrated with cofilin-1. Data are presented as mean ± SD. N = 3, *P < 0.05 versus control, **P < 0.01 versus control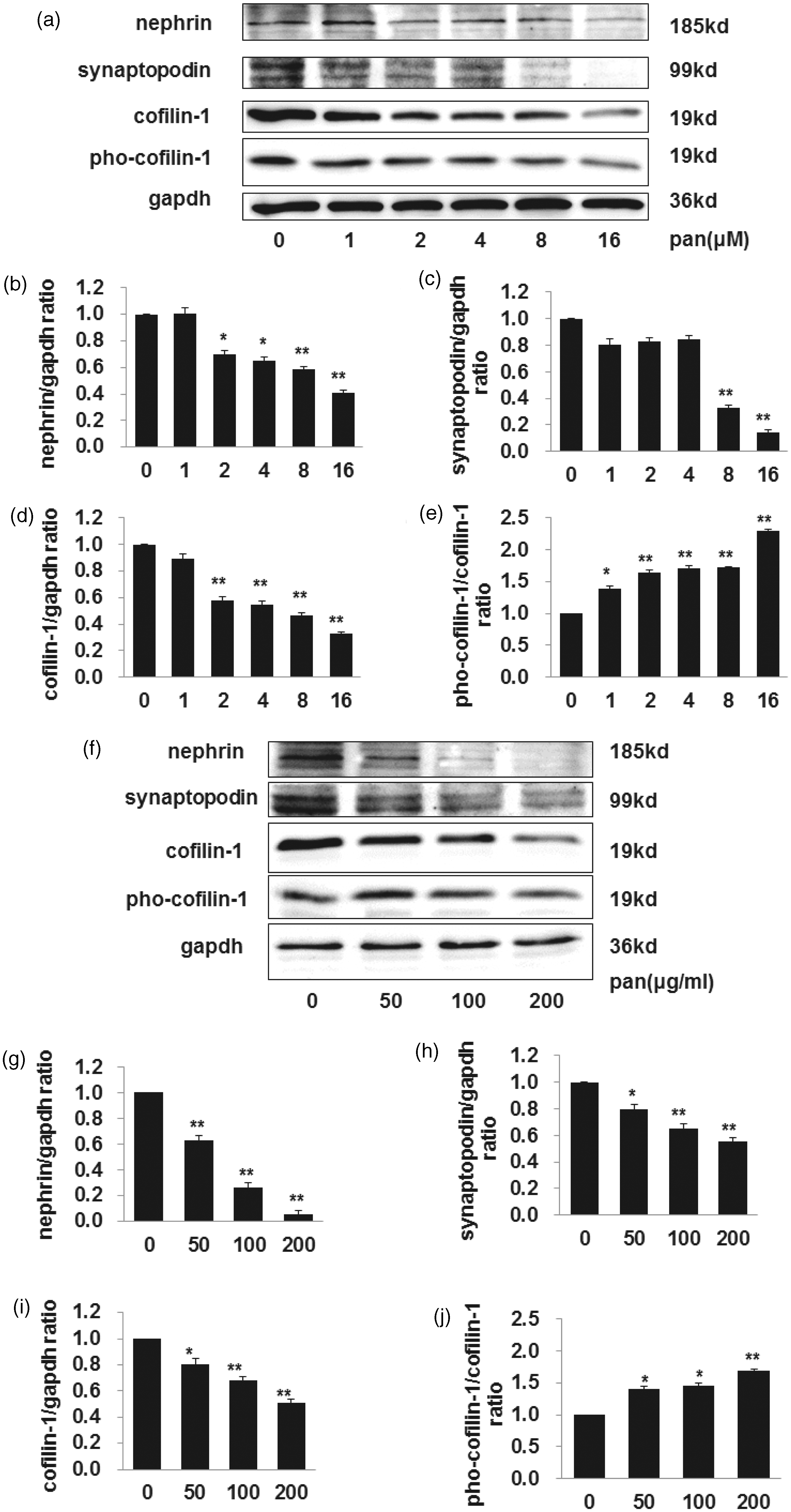
CsA restored the downregulation of cofilin-1 expression by PAN in vitro
Western blot results showed that the expression of cofilin-1 decreased significantly after treatment with PAN for 24 h versus the control, but administration of CsA significantly restored the expression of both cofilin-1 and pho-cofilin-1 (as shown in Figure 5, panels a to e).
Effects of CsA treatment on cofilin-1 and pho-cofilin-1 in cultured human podocytes. Cells were pretreated with Dimethyl Sulfoxide (DMSO) or CsA (0.5 µg/mL) for 1 h, then treated with vehicle control or PAN for 24 h. Western blot results of nephrin, synaptopodin, cofilin-1, and pho-cofilin-1 in cultured human podocytes (a). Calibrated with GAPDH, the expression of both nephrin and synaptopodin decreased significantly after being treated with PAN for 24 h compared to controls, and their expressions were restored with administration of CsA versus PAN group. However, when podocytes were exposed to CsA only, no significant changes of nephrin and synaptopodin expression were detected compared to controls (b, c). The expression of cofilin-1 was decreased in PAN group; CsA treatment restored its expression significantly (d). Calibrated with cofilin-1, the expression of pho-cofilin-1 was increased significantly after treatment with PAN for 24 h compared to control, and CsA partially restored its expression (e). Data are presented as mean ± SD. N = 3, *P < 0.05, **P < 0.01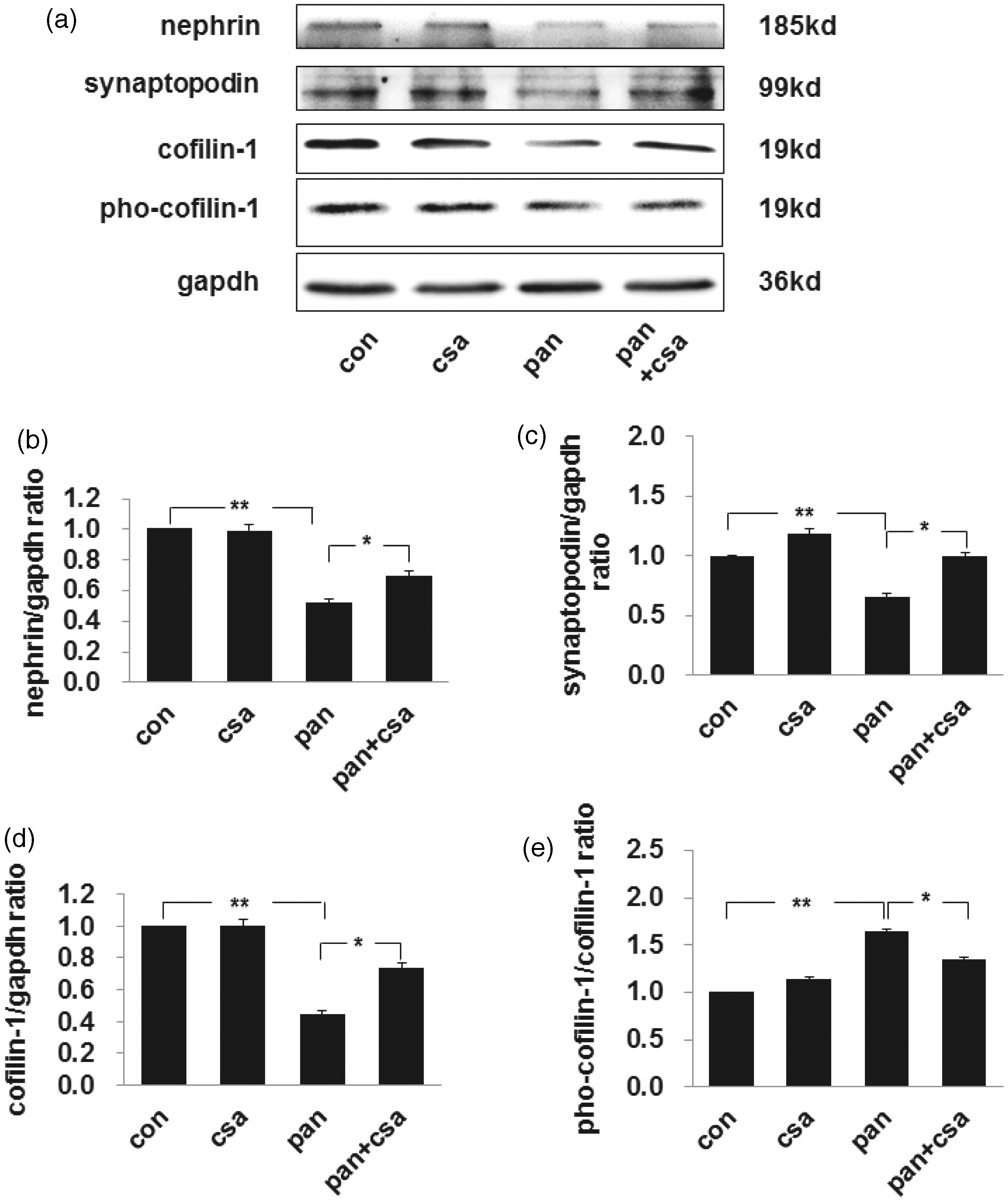
The immunofluorescence staining showed that the expression of cofilin-1 decreased significantly after treatment with PAN for 24 h versus control. However, administration of CsA significantly restored its expression. In normal cultured human podocytes, the staining of cofilin-1 was widely distributed in the cytoplasm (as shown in Figure 6, panel b1), whereas the fluorescence intensity decreased markedly for PAN-treated podocytes both in the cytoplasm and the nucleus (as shown in Figure 6, panel b2), and this was restored significantly following administration of CsA (as shown in Figure 6, panel b3). The immunofluorescence staining also showed that the expression of pho-cofilin-1 increased markedly in both the cytoplasm and the nucleus after PAN treatment for 24 h versus control, and administration of CsA restored its expression (as shown in Figure 6, panels f1 to f3).
Effects of CsA on expression and distribution of cofilin-1, pho-cofilin-1, and F-actin in cultured human podocytes. The nucleus was stained blue with Hoechst (a1–a3, e1–e3). Both cofilin-1 and pho-cofilin-1 were labeled green. Cofilin-1 was widely distributed in cytoplasm and nucleus in normal podocytes (b1), whereas their fluorescence intensity decreased in cytoplasm as well as nucleus in PAN-treated podocytes (b2). Administration of CsA restored the fluorescence intensity of cofilin-1 (b3). The fluorescence intensity of pho-cofilin-1 was weak in normal podocytes (f1). Pho-cofilin-1 fluorescence intensity increased in cytoplasm as well as nucleus in PAN-treated podocytes (f2). Treatment with CsA restored the fluorescence intensity of pho-cofilin-1 partially (f3). In normal podocytes F-actin was characterized by the presence of highly ordered parallel, contractile actin filament bundles (c1, g1), when injured with PAN, the cytoplasm was filled with reorganized, short, branched actin filaments in a disordered manner (c2, g2), the administration of CsA recovered the normal arrangement F-actin (c3, g3). Double-labeling assays showed that both cofilin-1 and pho-cofilin-1 colocalized with F-actin well (d1–d3, g1–g3). Bar = 20 µm. (A color version of this figure is available in the online journal.)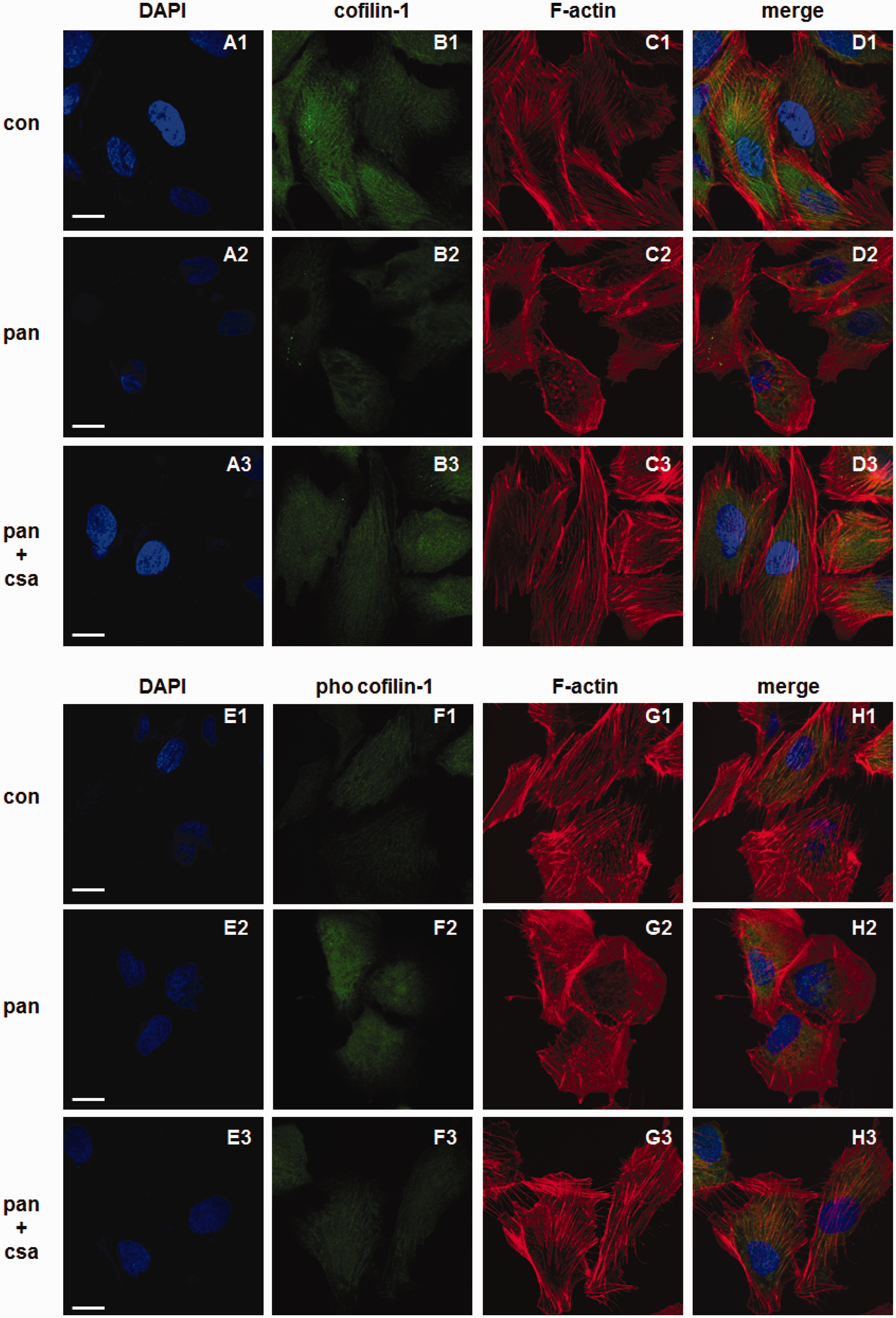
In normal podocytes F-actin was characterized by the presence of highly ordered parallel, contractile actin filament bundles (as shown in Figure 6, panels c1 and g1). Once injured with PAN, the cytoplasm was filled with reorganized, short, branched actin filaments in a disordered manner (as shown in Figure 6, panels c2 and g2), and administration of CsA recovered the normal arrangement of F-actin (as shown in Figure 6, panels c3 and g3).
Taken together, these data confirmed our hypothesis that cofilin-1 is crucial for PAN-mediated injury and CsA protected it by upregulating cofilin-1 expression. To examine whether the cofilin-1 mediates the protective effect of CsA after PAN-induced podocyte injury, we carried out a knockdown assay for cofilin-1 with siRNA.
Knocking down of cofilin-1 ameliorated the protective effect of CsA in vitro
To confirm cofilin-1 was critical for the treatment effect of CsA on podocytes, cofilin-1 expression was silenced with siRNA. Firstly, we tested the knockdown efficiency by using two different doses (30 and 60 pmol) of cofilin-1 siRNA. The real-time PCR results showed that cofilin-1 mRNA levels were silenced effectively by 39 and 73%, respectively (as shown in Figure 7, panel a), while western blot results showed that cofilin-1 protein levels were silenced by 67 and 85%, respectively (as shown in Figure 7, panels b and c). We defined the 60 pmol as the optimal dose to silence cofilin-1, and this dose was used in subsequent experiments.
Silencing of cofilin-1 in cultured human podocytes. The mRNA (a) and protein (b, c) expression of cofilin-1 decreased significantly after transfected with cofilin-1 siRNA by both concentrations (30 and 60 pmol, respectively) compared to negative control. Data are presented as mean ± SD. N = 3. *P < 0.05 versus negative control, **P < 0.01 versus negative control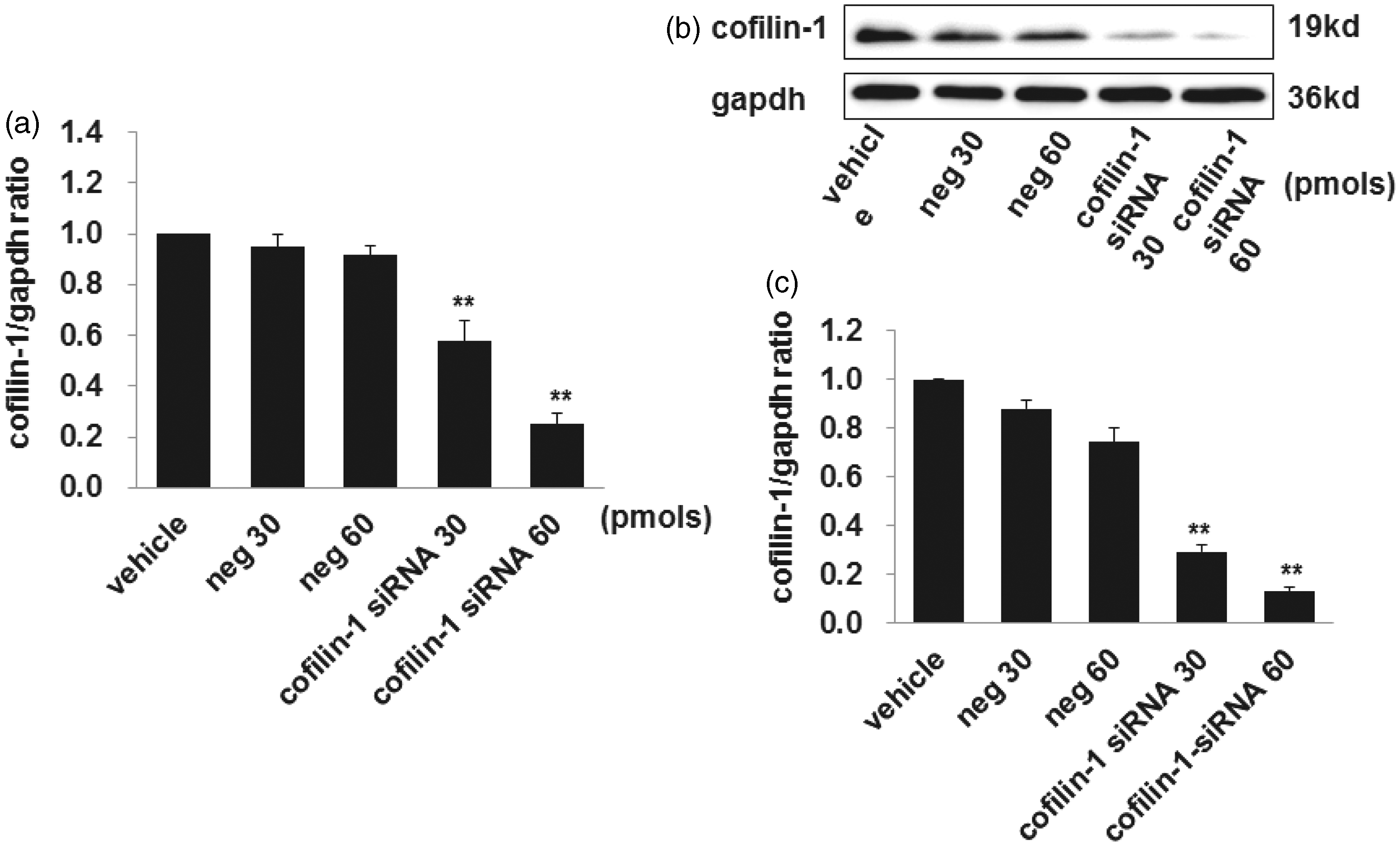
Secondly, podocytes were exposed to CsA with or without cofilin-1 siRNA exposure. Real-time PCR results showed that PAN decreased the expression level of cofilin-1, and administration of CsA restored the downregulation of cofilin-1 compared to the PAN group. The cofilin-1 mRNA level was decreased by 87% in the PAN + CsA + cofilin-1 siRNA group compared to the PAN + CsA group (as shown in Figure 8, panel a). The western blot results showed that PAN decreased the expression level of cofilin-1, and administration of CsA partially restored the downregulation of cofilin-1 compared to the PAN group (as shown in Figure 8, the third band of panels b and e). As a podocyte injury marker, nephrin decreased significantly after PAN injury compared to negative control and was restored after CsA treatment. However, CsA failed to restore nephrin expression when cofilin-1 was silenced (as shown in Figure 8, the first band of panels b and c).
Effect of CsA on cofilin-1 knocked down podocytes. The mRNA expression of cofilin-1 decreased significantly after PAN groups compared to negative control, and CsA restored its expression significantly, cofilin-1 decreased significantly in cofilin-1 siRNA group (a). Calibrated with GAPDH, both nephrin and synaptopodin protein levels were decreased significantly in PAN group compared to negative control, and administration of CsA significantly restored their expression (b, c, d). However, the ability of CsA was decreased significantly to restore nephrin expression when cofilin-1 was knocked down (c). Meanwhile there was no influence of CsA on synaptopodin expression whether cofilin-1 was knocked down or not (d). The protein expression of cofilin-1 was same as its mRNA expression (e). Data are presented as mean ± SD. N = 3. *P < 0.05, **P < 0.01. (A color version of this figure is available in the online journal.)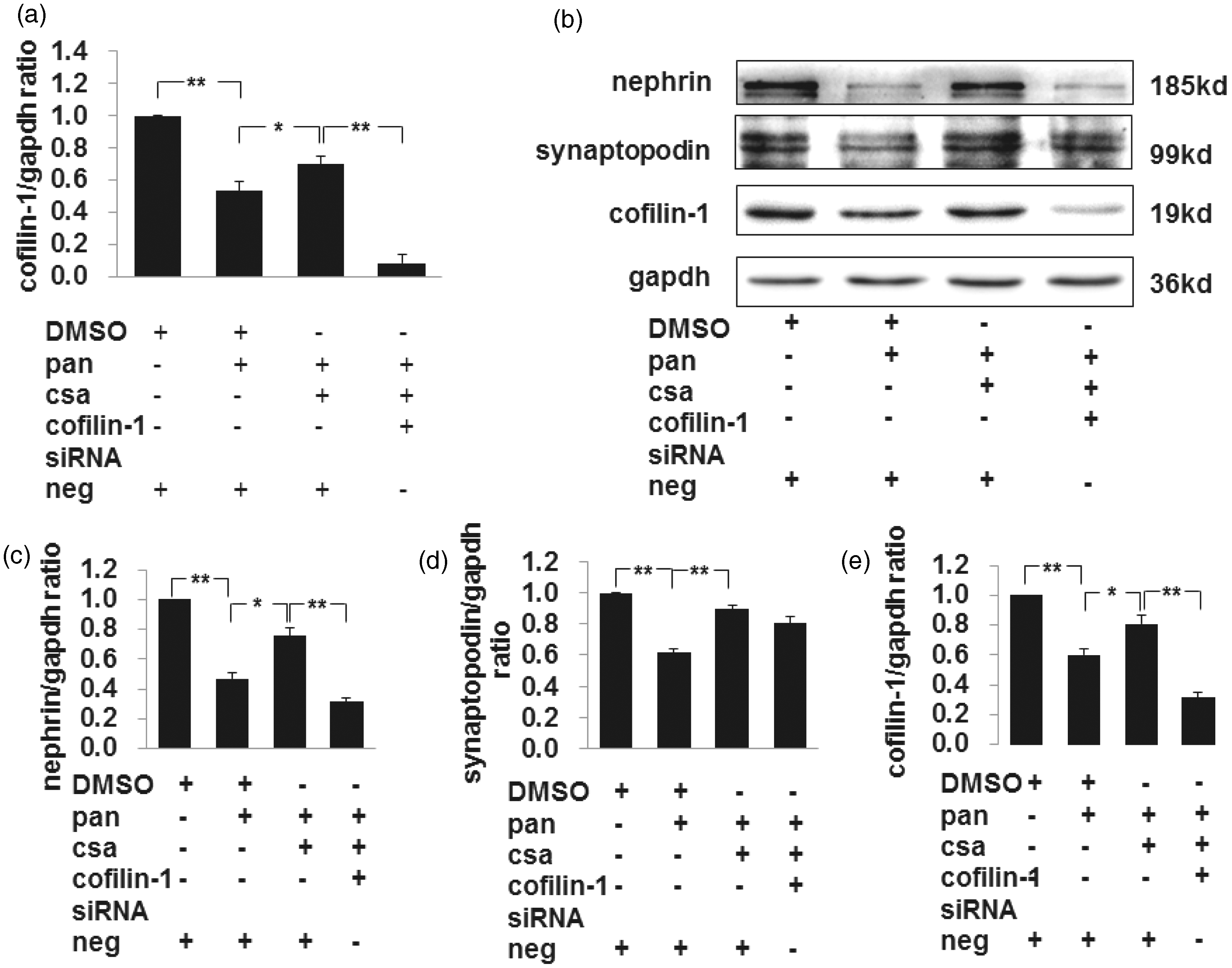
Finally, we attempted to detect the effect of cofilin-1 on the actin cytoskeleton of podocytes, in order to determine whether it participates in the effect of CsA treatment. The immunofluorescence staining showed the same change of cofilin-1 expression as real-time PCR and western blot results. In normal cultured human podocytes, the staining of cofilin-1 was widely distributed in the cytoplasm and nucleus (as shown in Figure 9, panel b1), whereas their fluorescence intensity decreased markedly in PAN-treated podocytes (as shown in Figure 9, panel b2). After administration with CsA, the fluorescence intensity restored significantly (as shown in Figure 9, panel b3). In normal podocytes F-actin was characterized by the presence of highly ordered parallel, contractile actin filament bundles (as shown in Figure 9, panel c1). When injured with PAN, the cytoplasm was filled with reorganized, short, branched actin filaments that were in a disordered manner (as shown in Figure 9, panel c2), and administration of CsA recovered the normal arrangement of F-actin (as shown in Figure 9, panel c3). Interestingly, CsA failed to recover normal actin arrangement when cofilin-1 was silenced (as shown in Figure 9, panel c4). Double-labeling assays showed that cofilin-1 was well colocalized with F-actin.
Effect of CsA on F-actin in cofilin-1 knockdown podocytes. The nucleus was stained blue with Hoechst (a1–a4), the cofilin-1 was labeled green and widely distributed in cytoplasm and nucleus in normal podocytes (b1), whereas their fluorescence intensity decreased in PAN-treated podocytes (b2). However, administration of CsA restored its fluorescence intensity (b3). In normal podocytes F-actin was characterized by the presence of highly ordered parallel, contractile actin filament bundles (c1). When injured with PAN, the cytoplasm was filled with reorganized, short, branched actin filaments in a disordered manner (c2), the administration of CsA recovered the normal arrangement F-actin (c3). However, CsA treatment failed to recover normal actin arrangement when cofilin-1 was knocked down (c4). Double-labeling assays showed that cofilin-1 colocalized with F-actin well (d1, d2, d3, d4). Bar = 40 µm. (A color version of this figure is available in the online journal.)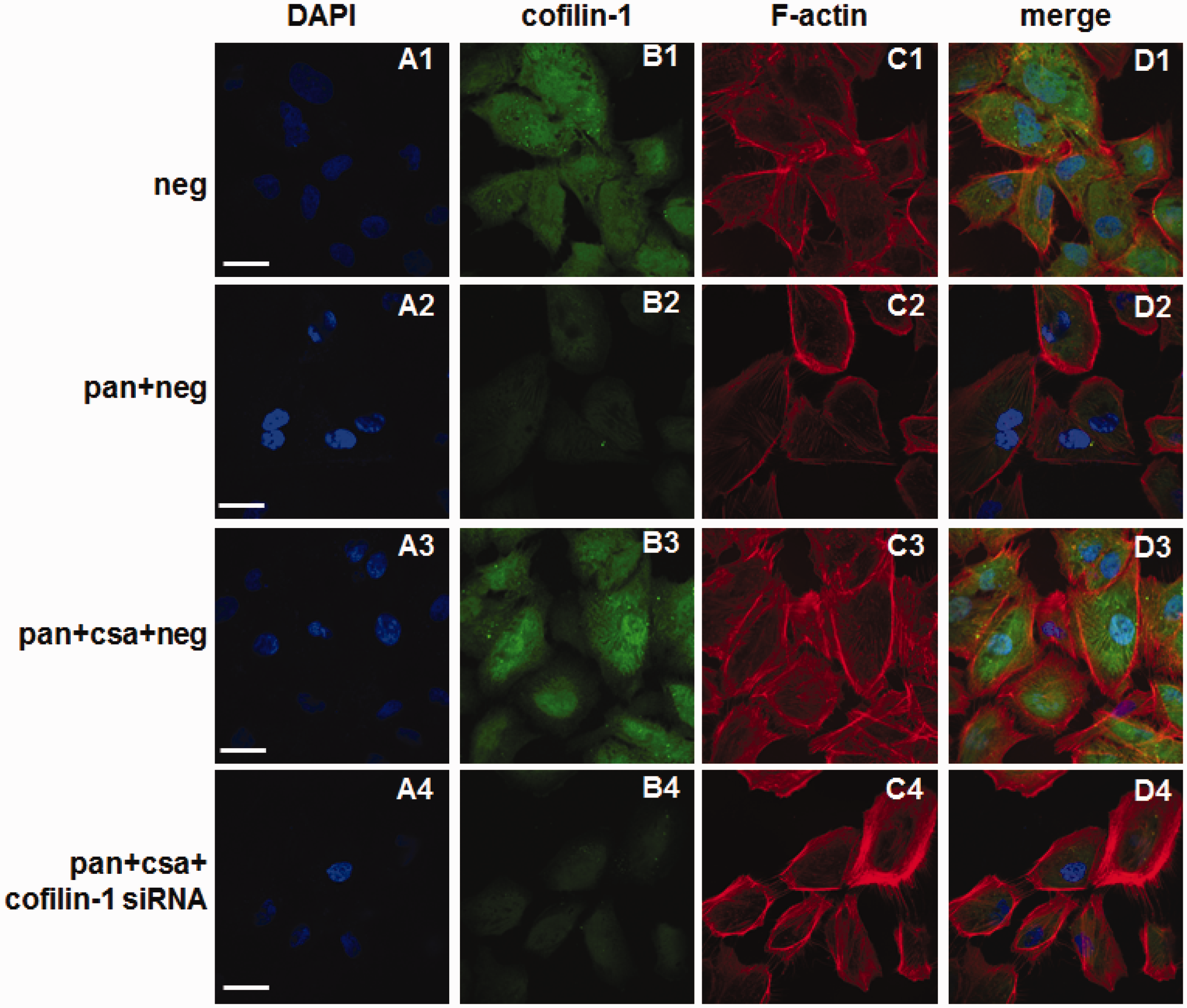
To further confirm whether CsA has a regulative effect on cofilin-1 in a synaptopodin independent manner, synaptopodin expression was silenced with siRNA. We tested the knockdown efficiency by using three synaptopodin siRNAs (synaptopodin siRNA-1, synaptopodin siRNA-2, synaptopodin siRNA-3). The real-time PCR results showed that the synaptopodin mRNA level was silenced effectively by 39, 74, and 65%, respectively (as shown in Figure 10, panel a), while western blot results showed that cofilin-1 protein level was silenced by 47, 83, and 77%, respectively (as shown in Figure 10, panels b and c). Synaptopodin siRAN-2 was therefore used in subsequent experiments.
Silencing of synaptopodin in cultured human podocytes. The mRNA (a) and protein (b, c) expression of synaptopodin decreased significantly after being transfected with three synaptopodin siRNAs compared to negative control; we choose synaptopodin siRNA 2 for the next experiment. Data are presented as mean ± SD. N = 3. **P < 0.01 versus negative control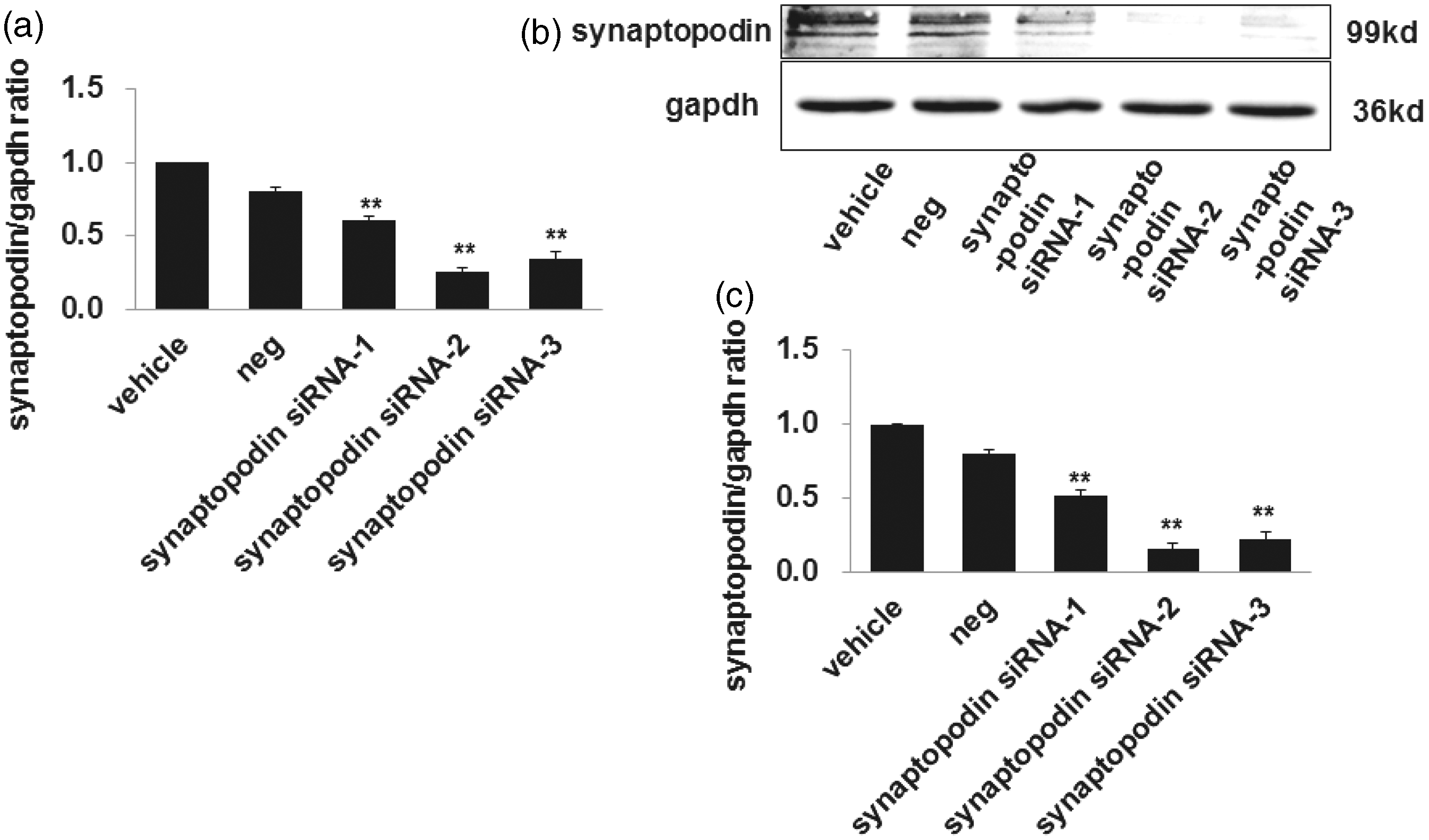
The podocytes were divided into four groups (PAN + CSA group, PAN + CSA + cofilin-1 siRNA group, PAN + CSA + synaptopodin siRNA group, and PAN + CSA + cofilin-1 siRNA + synaptopodin siRNA group). Our results showed that nephrin expression was decreased significantly in both the cofilin-1 silenced group and the synaptopodin silenced group compared to negative control. Meanwhile, the expression level of nephrin decreased markedly in the combined silenced group compared to the cofilin-1 silenced group and the synaptopodin silenced group, respectively (as shown in Figure 11, the first band of panels a and b). The synaptopodin expression was decreased significantly both in the PAN + CsA + synaptopodin siRNA group and the PAN + CsA + cofilin-1 siRNA + synaptopodin siRNA group. There were no significant differences of CsA on synaptopodin expression between the PAN + CSA group and the PAN + CsA + cofilin-1 siRNA group (as shown in Figure 11, the second band of panels a and c). The cofilin-1 expression was decreased significantly both in the PAN + CsA + cofilin-1 siRNA group and the PAN + CsA + cofilin-1 siRNA + synaptopodin siRNA group. There were no significant differences of CsA on cofilin-1 expression between the PAN + CSA group and the PAN + CsA + synaptopodin siRNA group (as shown in Figure 11, the third band of panels a and d).
Effects of CsA on nephrin, synaptopodin, and cofilin-1 in cofilin-1 silenced group, synaptopodin silenced group, and combined silenced group. Nephrin expression was decreased significantly in either cofilin-1 silenced group or synaptopodin silenced group compared to negative control. Meanwhile, the expression level of nephrin decreased significantly in combined silenced group compared to cofilin-1 silenced group and synaptopodin silenced group, respectively (a, b). The synaptopodin expression was decreased significantly both in PAN + CsA + synaptopodin siRNA group and PAN + CsA + cofilin-1 siRNA + synaptopodin siRNA group. There were no significant differences of CsA on synaptopodin expression between PAN + CSA group and PAN + CsA + cofilin-1 siRNA group (a, c). The cofilin-1 expression was decreased significantly both in PAN + CsA + cofilin-1 siRNA group and PAN + CsA + cofilin-1 siRNA + synaptopodin siRNA group. There were no significant differences of CsA on cofilin-1 expression between PAN + CSA group and PAN + CsA + synaptopodin siRNA group (a, d). Values were calibrated with to GAPDH. Data are presented as mean ± SD. N = 3, *P < 0.05, **P < 0.01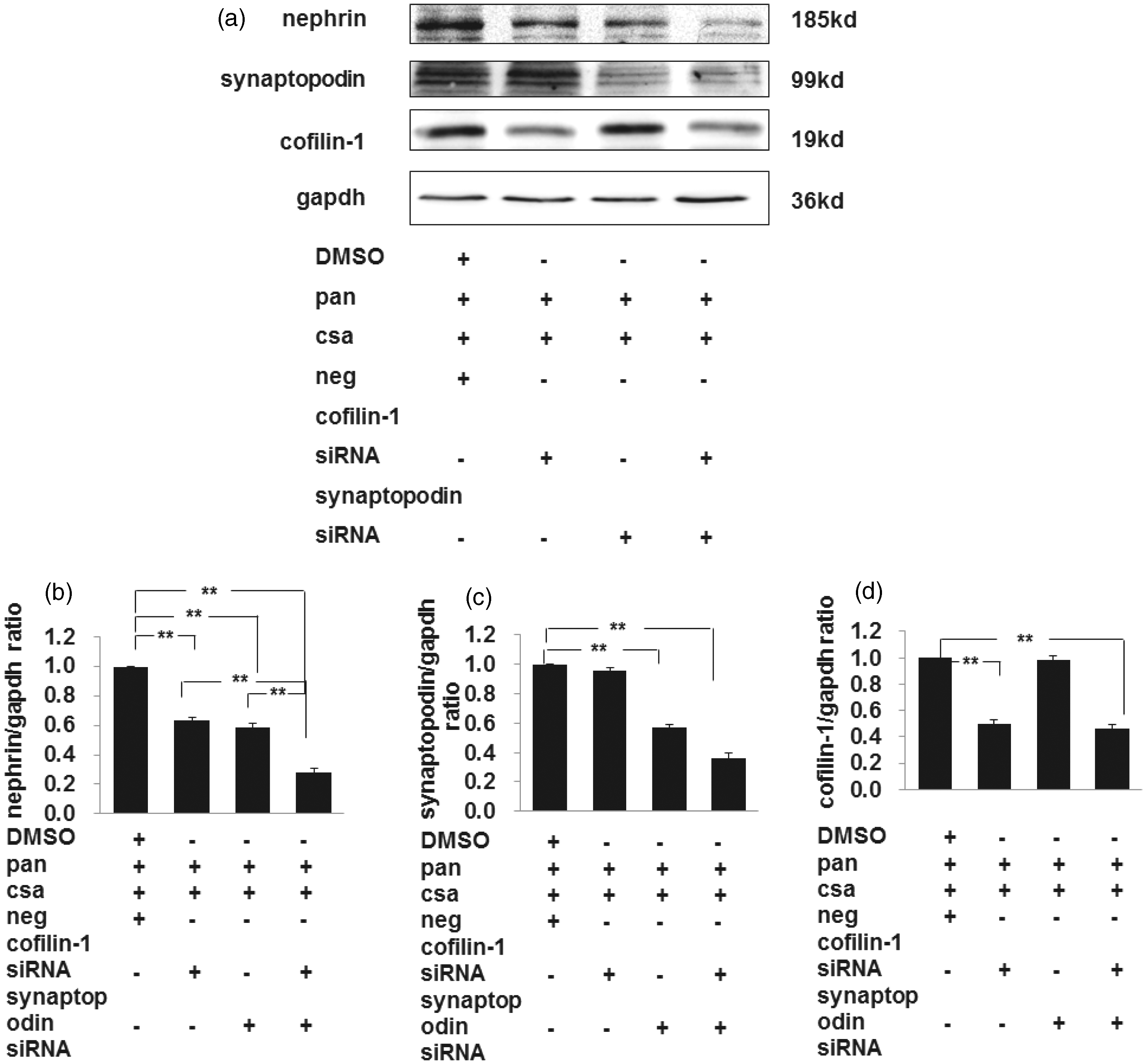
We also observed the reorganization of F-actin among the four groups. In the negative control group, F-actin was characterized by the presence of highly ordered parallel, contractile actin filament bundles (as shown in Figure 12, panel a). In the cofilin-1 silenced and synaptopodin silenced group, the cytoplasm was filled with reorganized, short, branched actin filaments in a disordered manner (as shown in Figure 12, panels b and c). In the cofilin-1 and synaptopodin combined silenced group, podocytes were turned round and had lost their normal structure, and F-actin reorganized in a disordered manner (as shown in Figure 12, panel d).
Effects of CsA on F-actin in cofilin-1 silenced group, synaptopodin silenced group, and combined silenced group. In negative control podocytes F-actin was characterized by the presence of highly ordered parallel, contractile actin filament bundles (a). In both cofilin-1 silenced and synaptopodin silenced group, the cytoplasm was filled with reorganized, short, branched actin filaments in a disordered manner (b, c). In cofilin-1 and synaptopodin combined silenced group, podocytes were turned round and structure lost (d). Bar = 20 µm. (A color version of this figure is available in the online journal.)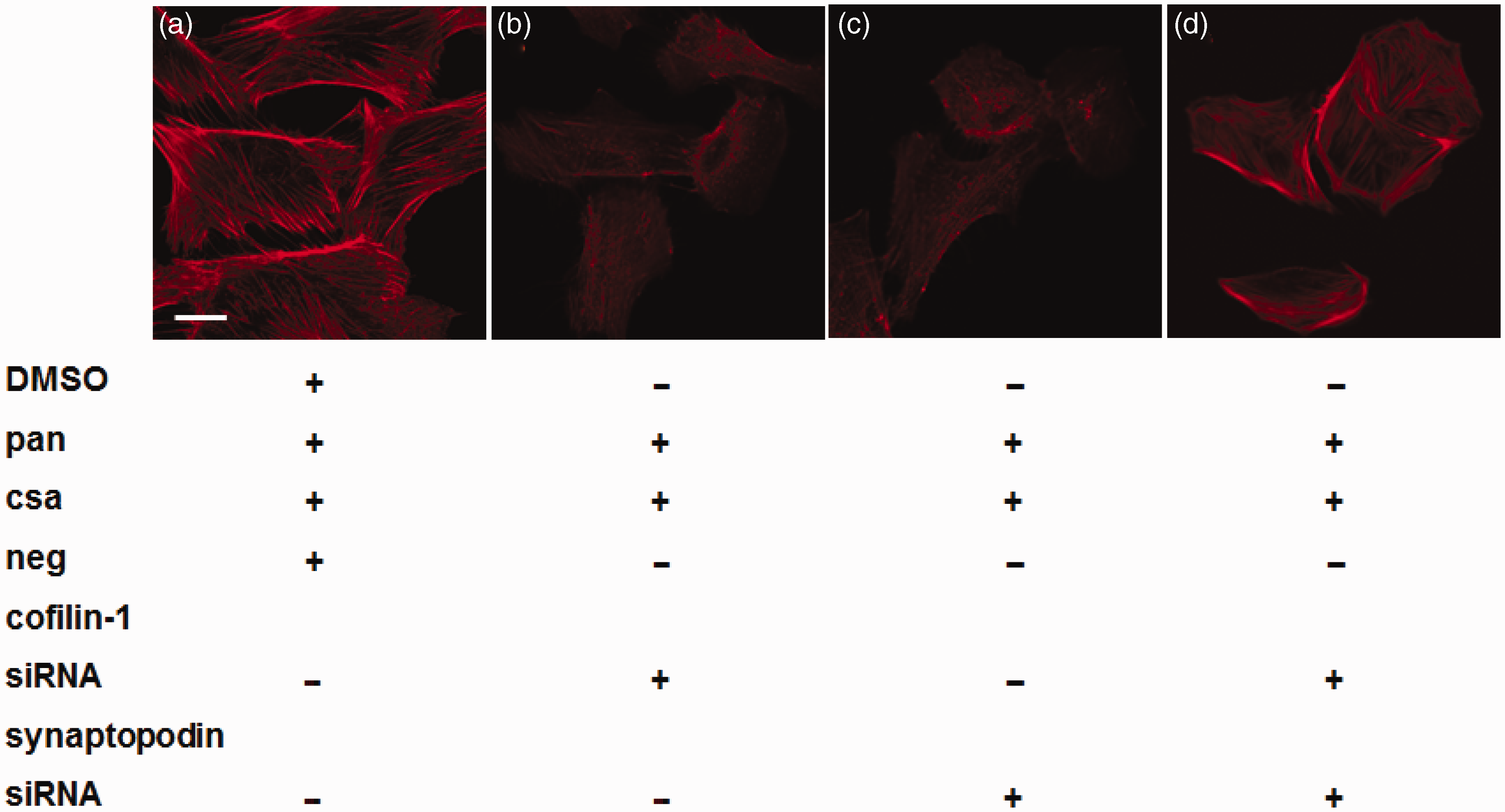
Both in vivo and in vitro data showed that the beneficial effects of CsA on PAN-induced podocyte injury was dependent, at least partially, on upregulation of cofilin-1 expression in podocytes, and this was independent of synaptopodin. Our data provided preliminary evidence that cofilin-1 seems to have a more dominant role in regulating podocytes than synaptopodin.
Discussion
It is now widely accepted that the podocyte is a key target of injury in a variety of renal diseases. Podocyte FP effacement is a common pathological phenomenon of proteinuric glomerular diseases. The effacement of podocyte FPs typically arises owing to perturbations in the actin cytoskeleton. 13 Since the FP effacement is a dynamic and reversible process, any drugs and proteins which regulate or stabilize the actin cytoskeleton are therefore crucial for sustained glomerular filter function. 14 CsA are widely used in the treatment of proteinuric diseases, and it is now realized that the effect of CsA is related to stabilization of the podocyte cytoskeleton. Cofilin-1, an actin-binding protein belonging to the ADF/cofilin family, plays a key role in actin filament dynamics in cells and has highly complex and interesting modes of regulation.2,15,16 In the present study, we firstly demonstrated that CsA protects podocyte injury via upregulating the expression of cofilin-1.
In our study, we firstly demonstrated that cofilin-1 expression was decreased in PAN-induced podocyte injury model both in vivo and in vitro. The more seriously podocyte injured, the more lower cofilin-1 expression declined. However, treatment of CsA could restore the expression of cofilin-1. We also found that CsA restored the disarrangement of F-actin after PAN injury. In our study, no significant change of cofilin-1 expression level was detected when CsA was administered alone. This result demonstrated that the protective effect of CsA mediated by cofilin-1 occurs in pathological conditions.
In the present study, we have shown that the expression of cofilin-1 was crucial for PAN-mediated podocyte injury, as supported by the following lines of evidence. Ashworth et al. have demonstrated that knockdown of cofilin-1 expression by both morpholino silencing and viral insertional mutagenesis resulted in altered zebrafish phenotypes, compromised glomerular filtration, and podocyte effacement. 6 Garg et al. have demonstrated that podocyte-specific cofilin-1 –/– mice developed FP effacement by 8 months of age. 5 These studies confirmed that cofilin-1 had critical functional activities in podocytes. We also found that phosphorylation of cofilin-1 was upregulated in PAN-induced podocyte injury both in vivo and in vitro. Cofilin-1 activity is regulated by many factors such as phosphorylation, pH, binding of phosphoinositides, and subcellular compartmentalization. It has been reported that cofilin-1 activity is inhibited by phosphorylation on serine 3 which leads to loss of actin binding and severing activities and subsequently results in decreased directed cell motility.17,18 The phosphorylation effect was regulated by LIM kinases 1 and 2, which are activated by Cdc42 and Rac and Rho-dependent kinase (ROCK).19–21 Cofilin-1 activity is also inhibited by binding phosphatidylinositol 4,5-bisphosphate (PIP2) at the plasma membrane and the scaffold protein 14-3-3.22,23 Stimulation of cofilin-1 activity by dephosphorylation of serine 3 is conducted by Slingshot1 (SSH1) and chronophin phosphatases.24,25 Therefore, both the phosphorylated and dephosphorylated forms of cofilin-1 have potential functional activities in podocytes. A previous study has shown that phosphorylated cofilin-1 was increased in the podocyte nuclei of patients suffering from Focal Segmental Glomrulosclerosis (FSGS), Membranous Glomerulonephritis (MGN), Minimal Change Disease (MCD). 6 Meanwhile, several groups reported that phosphorylation of cofilin-1 was involved in podocyte injury. Garg et al. found that after PAN injection for 3 days in female Sprague-Dawley rats, besides FP effacement being initially observed, cofilin Ser 3 phosphorylation was markedly attenuated in glomerular lysates compared with controls. Other studies also showed that CsA blocked Ca2+ induced cofilin dephosphorylation and SSH1L activation in Hela cell and induced cofilin phosphorylation and promoted stress fiber generation in proximal tubular cells.7,26 In contrast, our data showed that treatment with CsA partially decreased the abnormal upregulation of relative phosphorylated cofilin-1 expression, and this was different from its effect on tubular cells as reported previously by another group. We suggest that the different mechanisms of CsA may be involved in the regulation of cofilin-1 activities. Moreover, the different experimental model and different time points might be another reason. Furthermore, since the effect of CsA on podocytes and tubular cells was different, there may be different mechanisms of CsA on cofilin-1 regulation. Descazeaud et al. analyzed the reason for different roles of CsA on podocytes and tublular cells. They proposed that in glomerular cells, CsA inhibits CaN action on the actin filament, thus having a beneficial effect by restoring normal podocyte structure. In contrast, in the tubular structure, the stiffening of the actin network induced by CsA may alter the response of proximal cells to modifications of tubular flow, thus exerting a detrimental effect on tubular structure. 27 Our data demonstrated for the first time that the expression of cofilin-1 was important in the antiproteinuric effect of CsA. In the present study, we observed that the proteinuria induced by PAN was reduced by CsA treatment, and the FPs effacement was repaired after CsA treatment. However, the precise mechanism of how CsA reduces proteinuria in nephrotic syndrome and PAN-induced nephropathy remains to be further studied.
We report that cofilin-1 is distributed in both the cytoplasm and the nucleus. The function of cofilin-1 in the cytoplasm involves stabilizing the actin cytoskeleton, and this is in accordance with other studies.16,28,29 However, the function of cofilin-1 in the nucleus was not clear. Several studies presumed that cofilin-1 chaperoned actin to the nucleus, which is important in chromatin remodeling, the formation of heterogeneous nuclear ribonucleoprotein complexes, and gene expression.30,31 Some evidence indicates that cofilin forms a complex with actin and DNaseI and perhaps plays a role in DNA degradation and initiation of apoptosis. 32 In the present study, we also found that cofilin-1 was widely distributed in the cytoplasm and the nucleus in normal podocytes, and their fluorescence intensity decreased in the cytoplasm as well as in the nucleus after PAN injury. The treatment with CsA could restore cofilin-1 fluorescence intensity. We proposed that cofilin-1 plays an important role in both the cytoplasm and the nucleus. However, the role of cofilin-1 in the nucleus is not clearly understood.
Nephrin is a 185-kDa transmembrane molecule with Ig-like domains and a structural component of the slit diaphragm.33,34 Nephrin also serves as a signaling scaffold by recruiting proteins to the cytoplasmic face of podocyte FPs. 35 The absence of nephrin or inherited mutation of nephrin leads to proteinuria and rearrangement of the actin cytoskeleton. 13 Several studies have reported a modulation or correlation of nephrin expression with the development of proteinuria, including PAN, diabetes, and MCD.36–39 Our studies have uniquely demonstrated that CsA partially restored the decrease of nephrin expression and the disarrangement of F-actin induced by PAN in a cofilin-1 mediated manner. Garg et al. have demonstrated that cofilin-1 interacted with nephrin, and the activity of cofilin-1 was regulated by nephrin in a PI3K-mediated way. 5 However, the mechanism of cofilin-1 regulating nephrin and F-actin still needs further study.
Our data showed that both cofilin-1 and synaptopodin expression were decreased during PAN-mediated podocyte injury. It is reported that a key event in the development of podocyte FP effacement and proteinuria lies in the induction of a cytosolic form of protease cathepsin L (CatL) in podocytes, which triggering the disease process.1,40 Cytosolic CatL proteolyzes synaptopodin and results in disorganization of podocyte actin cytoskeleton and FP effacement. The CatL inhibitor E64 prevented the loss of synaptopodin expression. 1 However, it is still not clear how cofilin-1 was downregulated and perhaps the expression of cofilin-1 was also regulated by synaptopodin, since whose expression was also decreased after PAN injury. To testify the regulative role of synaptopodin on cofilin-1, the kinase inhibition experiment in the presence of the cathepsin inhibitor E64 together with PAN will be performed in our further study.
Since synaptopodin is a well-characterized target of CsA, it is important to identify whether CsA has a synaptopodin independent direct effect on cofilin-1 and the actin cytoskeleton. In the present study, we also found that nephrin expression was decreased significantly in either the cofilin-1 silenced group or the synaptopodin silenced group compared to the negative control. Meanwhile, the expression level of nephrin decreased significantly in the combined silenced group compared to the cofilin-1 silenced group and the synaptopodin silenced group, respectively. We also found that synaptopodin knocked down had no effect on cofilin-1. These results confirmed that the protective effect of CsA on podocytes via cofilin-1 is independent of its effect on synaptopodin. Our data showed that cofilin-1 seems to have a more dominant role in regulating podocytes than synaptopodin.
In conclusion, our study suggested that both cofilin-1 and its phosphorylation form were involved in PAN-induced nephropathy and podocyte injury. The antiproteinuric effect of CsA is directly from the stabilization of the podocyte actin cytoskeleton through upregulation of actin-binding protein cofilin-1, which was independent of its role on synaptopodin. Therefore, we consider that cofilin-1 may be an attractive molecular target for structural maintenance of podocyte FPs and for antiproteinuric treatment in the future.
Footnotes
Author contributions
All authors participated in the design, interpretation of the studies, and analysis of the data and review of the manuscript. XL conducted the experiment and wrote the manuscript, XZ and JD reviewed the whole manuscript and XL, XW, SW supplied technical support. XL and XZ contributed equally to this work.
ACKNOWLEDGEMENTS
We are grateful to Professor Peter Mundel (USA) and Professor Moin Saleem (Briston) for the podocyte clones, and Professor Karl Tryggvason (Sweden) for nephrin antibody. We thank Lixia Yu and Guohong Wu for the technical support in tissue sectioning. We thank Yinghong Tao and Guosheng Yang for their help during the animal experiment. This study was supported by the National Basic Research Program of China (973 Program, 2012CB517701) and National Nature Science Foundation of China (81170657/30830105/81200511).