
Abstract
Obesity represents a route to broad physiological dysfunction affecting major organs including male urogenital system. Hyperglycemia, hyperlipidemia, and oxidative stress associated with obesity augment the formation of reactive metabolic by-products, namely advanced glycation end products (AGEs), leading to increased tissue deposition and damage. The exogenous intake and the endogenous accumulation of AGEs contribute to metabolic and reproductive abnormalities in both women and men. The present study assessed the effects of a diet high in saturated fatty acids (SAFA) on the lipid and metabolic profile (AGE levels, oxidative stress) as well as pathogenic (AGE, receptor for AGEs [RAGE] expression, apoptosis) and morphometric parameters of male reproductive system in vivo. Effects of switching to a diet rich in monounsaturated fatty acids (MUFA) or equal in the proportion MUFA to SAFA were further investigated. SAFA-fed animals were characterized by increased serum lipid concentrations (p < .05) compared to controls, but AGEs and peroxide levels were not significantly different across the different experimental groups. Elevated AGE deposition was detected for the first time in germ cells with a higher staining intensity in animals on the SAFA diet, compared to MUFA or MUFA–SAFA-fed animals or the control samples (p = .018). In Leydig cells, AGE localization was higher in the entire cohort of high-fat-fed animals compared to controls (p < .05). High-fat-fed mice displayed enhanced apoptosis compared to controls (p < .005). Furthermore, prostatic tissue demonstrated reduction in epithelial folding, an effect which was significantly reversed after MUFA diet administration. Our findings provide the basis for further investigation of AGE–RAGE axis in testicular and prostatic disturbances associated with diet-induced obesity. Simple dietetic intervention has beneficial effects on metabolic dysfunction of reproductive system before overt manifestations, indicating glycation as a promising therapeutic target.
Keywords
Introduction
Obesity is becoming one of the most serious current epidemics in countries of the Western world as a result of diets high in simple carbohydrates and saturated fat that are associated with sedentary lifestyle.1,2 Metabolic disturbances associated with obesity such as type 2 diabetes, hypertension, hypercholesterolemia, hypertriglyceridemia, and nonalcoholic fatty liver disease form a pathogenic cluster known as metabolic syndrome (MetS). Emerging evidence in recent years is linking obesity and MetS to urogenital system disorders, 3 gaining research interest due to potential reversibility of the effects through lifestyle manipulation.
Although there are strong links with hypogonadism 4 and erectile dysfunction, 5 published data are conflicting both on the effects on various parameters of spermatogenesis as well as on the risk of infertility among couples with overweight or obese male partner. 6 Additionally, cumulative data, arising initially from epidemiological observations but confirmed by clinical and pathophysiological studies, suggest that development of benign prostatic hyperplasia and progression of prostate cancer are associated with obesity per se or in the context of MetS. 7
Excess of metabolic glycation by-products (advanced glycation end products, AGE) occurs in diverse settings, including diabetes and aging. 8 The degree of hyperglycemia and the extent of oxidant stress associated with obesity and MetS lead to their accumulation and increased tissue deposition. 9 Previous studies from our group have demonstrated augmented AGE deposition in the ovaries of women with polycystic ovary syndrome, 10 as well as of normal female rats exposed to AGE-enriched diet, 11 associated with metabolic and hormonal abnormalities, suggesting their implication in female reproductive perturbations. In males, Mallidis et al. have shown that Nɛ-carboxymethyl-lysine (CML), a prominent AGE, accumulates in the reproductive tract of patients with diabetes 12 as well as in animal models of both diabetes 13 and MetS. 14 Furthermore, the receptor for AGEs (RAGE) was immunolocalized in testes, epididymides, and sperm, coinciding with increased sperm nDNA fragmentation.13–15 Given that RAGE is expressed in invasive and noninvasive prostate cancer cells, it has been proposed as a central factor in prostate cancer tumorigenesis and metastatic potential through its interaction with other ligands as well as with AGE. 16
The aim of the present study was to explore the differential pathogenic impact of diet in the biochemical and metabolic profile as well as in the reproductive system of an established animal model of diet-induced obesity and MetS. It was postulated that diets differing in saturated fatty acid (SAFA) and monounsaturated fatty acid (MUFA) content after the induction of obesity and MetS in a genetically intact C57BL/6 mouse model would affect, apart from the biochemical and lipid profile, morphometric and pathogenic factors of the urogenital system. To gain insight into the pathophysiologic role of AGE–RAGE interaction in male reproductive system, their expression was investigated in parallel with assessments of histologic alterations of the prostate and induction of apoptotic pathways.
Materials and methods
Animals
Fifty-eight 3-month-old male C57BL/6 mice were obtained from Pasteur Institute, Athens, Greece. The animal protocol was approved by the Veterinary Service of the Prefecture of Athens, Animal Care and Use Committee (permit no. K/6030/18-10-2005) according to the Greek legal requirements for animal experimentation and the University of Athens Medical School.
The animals were housed in cages under stable environmental conditions (i.e. temperature 18–20℃, relevant humidity 55 ± 10%, 10 h of light and 14 h of darkness) throughout the duration of the experiment and were given pelleted food and water ad libitum at the animal house facility of the Department for Experimental Surgery and Surgical Research “N.S. Christeas,” University of Athens Medical School.
Experimental design
Nutrient composition of experimental diets
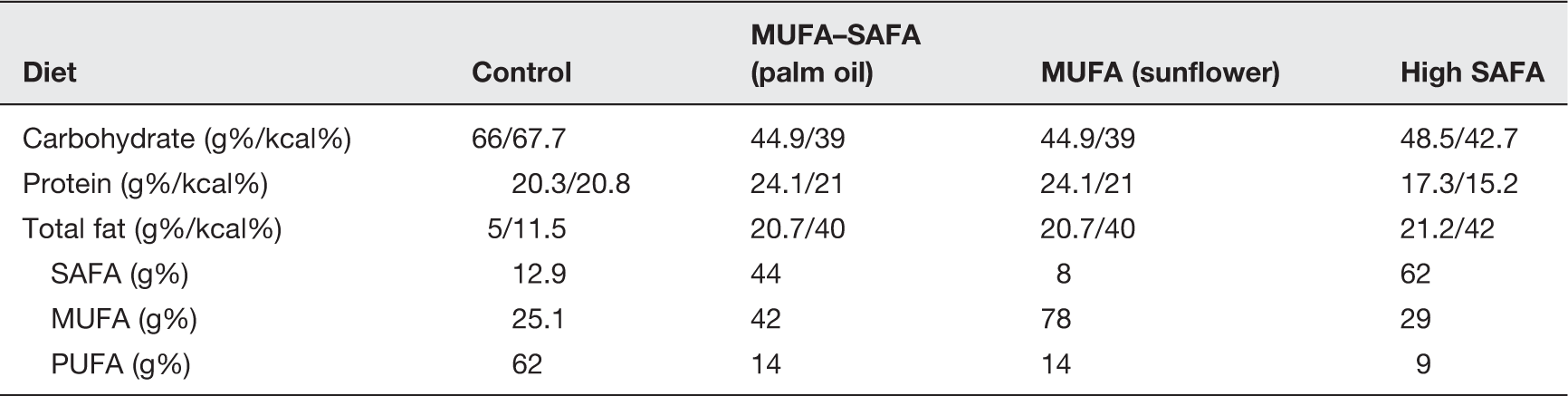
After the development of hypercholesterolemia, animals were administered diets equal in fat content but differing in proportion of SAFA and MUFA concentration for 16 more weeks (Figure 1). Sixteen animals in each group were fed either a diet rich in MUFA in the form of sunflower oil (fatty acid composition: 78% MUFA, 14% poly-unsaturated fatty acid [PUFA], 8% SAFA; D05101205 from Research Diets) or a diet with an equal MUFA and SAFA content in the form of palm oil (fatty acid composition: 42% MUFA, 14% PUFA, 44% SAFA; D05101204 from Research Diets) (MUFA–SAFA Group) or continued the previous high SAFA diet (SAFA Group). At the end of 24 weeks all animals were sacrificed for blood collection and prostatic and testicular tissue were removed.
Outline of experimental design
Biochemical and hormonal measurements
Biochemical measurements for lipid profile assessment were performed previously and independently of this project. 17 Serum testosterone levels (ng/mL) were estimated using the commercially available immunoassay kit from IBL Immunoassays (Hamburg, Germany).
Evaluation of serum AGE levels
Serum AGE levels (U/mL) were measured by AGE-specific competitive Enzyme-linked immunosorbent assay (ELISA) as described previously by Diamanti-Kandarakis et al. 18 Briefly, sera were thawed and diluted 1:5 with dilution buffer (phosphate-buffered saline [PBS], 0.02% Tween-20, 1 mM sodium azide [NaN3]). All measurements were performed at the Chemwell analyser (Awareness, Palm City, Florida, USA). Ninety-six-well ELISA plates were coated with 100 µL/well of µg/mL AGE-bovine serum albumin (BSA) in coating buffer overnight at 4℃. Wells were washed three times with 150 µL washing buffer and then blocked with 100 µL Superblock™ blocking buffer in PBS at room temperature for 1 h. After three rinses with washing buffer, 50 µL of competing antigen in dilution buffer was added, followed by 50 µL of anti-AGE mAb (6D12, Cosmo Bio, Tokyo, Japan—dilution 1:1000) in dilution buffer containing 2% Normal Goat Serum (NGS). Plates were incubated at room temperature for 2 h with gentle agitation on a horizontal rotary shaker. Wells were subsequently rinsed three times with washing buffer. Secondary antibody in dilution buffer with 1% NGS was then added to each well and the plates were incubated at 37℃ for 1 h. After rinsing three times with washing buffer, 100 µL p-nitrophenyl phosphate (p-NPP) substrate was added to each well. Optical density (OD) at 405 nm was determined by a SPECTRA Fluor Plus TECAN reader after 30–60 min. Results were expressed as B/Bo, calculated as (experimental OD − background OD [i.e. no antibody])/(total OD [i.e. no competitor] − background OD).
Evaluation of oxidative status in serum samples—Oxystat assay
The oxidative status in serum samples was evaluated by measuring the total peroxide concentrations using the Biomedica OxyStat assay (Biomedica Gruppe, Vienna, Austria), according to the manufacturer’s instructions. Briefly, the kit detects total peroxide concentration of a sample based on the reaction of the circulating biological peroxides with peroxidase and a subsequent color reaction using 3,3′,5,5′-tetramethylbenzidine as substrate. After addition of stop solution, the colored liquid is measured photometrically at 450 nm. For the assay, a calibrator is used to calculate the concentration of biological peroxides in the sample. The concentration is expressed as H2O2 equivalents (µmol/L). Results show a direct correlation between free radicals and circulating biological peroxides, which allows the characterization of the oxidative status in biological samples.
Immunohistochemical staining
Paraffin-embedded sections of formalin-fixed testicular and prostate tissue were deparaffinized by xylene and dehydrated in graded ethanol. Sections were treated in 3% H2O2 in PBS for 15 min and then rinsed in PBS. Antigen retrieval was performed in 0.01 M citric acid-buffered solution (pH 9.0) and microwaving at 500 W for 5 min. After thorough washing, the sections were incubated with normal rabbit serum for 20 min at room temperature to avoid nonspecific binding of the antibodies. The sections were then incubated overnight at 4℃ with the anti-AGE mAb 6D12 (0.25 mg/mL stock, dilution 1:50) or goat monoclonal anti-RAGE-antibody (Research Diagnostics Inc., Flanders, NJ, USA; 0.25 mL, dilution 1:400) in PBS containing 1% BSA.
Immunoreactivity was detected by the streptavidin–biotin–peroxidase method according to the manufacturer’s protocol. The final reaction product was visualized with 3,3′-diaminobenzidine tetrahydrochloride (LSAB detection kit; Dako, Carpinteria, San Diego, CA, USA). Lung tissue sections from diabetic rats were used as positive controls for AGE and RAGE antibodies. Negative controls (e.g. prostatic or testicular tissue in which the primary antibody was substituted with nonimmune serum) were also stained in each run. The percentage of positive cells was estimated using light microscopy. The evaluation of the immunostained slides was performed blindly and independently by PK and GL. AGE and RAGE staining intensity were semiqualitatively assessed and categorized in four levels: +/–, very weak; +, weak; ++, moderate; and +++, strong. AGE and RAGE expression were categorized in four levels according to the percentage of positive cells, as follows: minimum (1–10% of cells), low (11–30% of cells), moderate (31–60% of cells), and extensive (>60% of cells) immunostaining. Semiquantitative expression data were presented as an H-score that is produced by average intensity of positive staining multiplied by the percentage of cells showing positive staining.
Measurement of apoptosis
Apoptotic cells were quantified immunohistochemically by means of a mAb recognizing exposed single-stranded (ss) regions in the DNA of apoptotic cells during heating, as previously described. 19 This reagent (MAb F7-26; Apostain, Alexis Biochemicals, San Diego, CA, USA) allows the specific discrimination between apoptotic and necrotic cells. Briefly, according to the manufacturer’s protocol, formaldehyde-fixed, paraffin-embedded sections were incubated for 20 min with 0.1 mg/mL saponin in PBS, followed by treatment with 20 µg/mL proteinase K for 20 min at room temperature and then washing with distilled water. Sections were then incubated in a 50% aqueous solution of formamide at 56℃ for 20 min, washed for 5 min in cold PBS, then exposed to 3% H2O2 in PBS for 5 min, and rinsed in distilled water. Nonspecific binding was blocked with 3% skim milk at room temperature. Sections were incubated with the mAb F7-26 for 30 min at room temperature and then with an anti-mouse IgM conjugate (1:100 dilution) for another 30 min at room temperature. Application of diaminobenzidine as chromogen and subsequent counterstaining with hematoxylin completed the procedure. The quantification of apoptotic cells was based on the total number of ssDNA-positive cells with apoptotic morphology counted in five randomly selected optical fields at ×200 magnification. A case of Burkitt’s lymphoma was used as a positive control. The positive cells were counted without previous knowledge of the experimental condition.
Morphometric procedures
A segment of the prostatic complex from each mice, comprising a representative sample of the dorsolateral lobes and the urethra, cross-sectional cuts from the midportions of both seminal vesicles with the adhering coagulating gland (anterior lobe of the prostate), as well as ventral lobes in close proximity to urinary bladder was fixed in 10% buffered formalin. Likewise, testicular tissue was processed by routine histological methods. Serial sections 4 - to 5-mm thick of the fixed prostatic and testicular tissue embedded in paraffin were stained by hematoxylin–eosin for standard microscopic evaluation. Epithelial, luminal, and stromal areas in the ventral lobe of the prostate were assessed for structural analyses in the sections stained for light microscopy in all animals of each group, using ×200 magnification. The degree of mucosal folding was assessed in three levels, i.e. 0, low or absent; 1, moderate; 2, normal.
Measurement of seminiferous tubule diameter
The 10 most circular seminiferous tubules were randomly identified in each section of the testis and their diameters were measured with an ocular micrometer using the ×10 lens. The mean seminiferous tubule diameter (in micrometers) was then determined for each testis.
Measurement of seminiferous tubule basement membrane (STBM) thickness
Five-micrometer-thick sections obtained from each animal were viewed under a microscope with an attached video camera and image analyzer system using Image-Pro Plus v.6.0 software (Media Cybernetics, Rockville, MD, USA). The measurement was performed on 10 randomly selected STBMs and averaged.
Statistical analysis
Quantitative normally distributed data are expressed as mean ± 1 standard deviation (SD) and as median Interquartile Range (IQR) in cases of asymmetric distributions. The Kolmogorov–Smirnov test was used to assess the normality of distribution. Comparisons between more than two groups were performed with analysis of variance (ANOVA) and Kruskal–Wallis test. In the latter cases, Mann–Whitney’s U test and False Discovery Rate (FDR) were used for post hoc multiple testing. Comparisons between more than two measurements were performed using repeated neasures ANOVA, Friedman’s test, and Wilcoxon’s signed rank test with FDR, as appropriate. Pearson’s correlation coefficient and Spearman’s rho were calculated in order to examine linear relationships between variables. In all cases of multiple hypotheses testing, FDR was utilized in order to assess between-group differences, as well as to control family-wise error to < 0.05. All tests were two sided. Statistical significance level was set at 0.05.
Results
Somatometric and biochemical parameters
Somatometric and metabolic parameters of all groups throughout the different phases of the experiment are presented in Table 2.
Before randomization into the four study groups, there were no significant differences between the groups in total cholesterol (TC), low-density lipoprotein cholesterol (LDL-C), high-density lipoprotein cholesterol (HDL-C), and triglyceride (TG) levels or body weight.
After the first 8 weeks, a significant increase in weight of animals both in the control and the high SAFA-fed group was observed. TC and HDL-C concentrations were lower compared with baseline readings (p < .05) in control animals, whereas in mice belonging to the remaining three groups on the high SAFA diet they were significantly increased compared to prediet levels (p < .05).
At the end of the study (week 24), body weight was significantly increased in controls (p < .05) compared to 8th week measurements at levels not different from MUFA and MUFA–SAFA diet groups (p > .05). Administration of both MUFA-rich diet and MUFA–SAFA diet resulted in a significant decrease in TC, HDL-C, and LDL-C levels (p < .05) compared with the 8th week measurements, while MUFA–SAFA diet group exhibited a significant increase in serum TG levels (p < .05) at the same period. Both low SAFA diets implementation restored low TC and LDL-C values at levels not significantly different from control group.
Interestingly, animals were found to have significantly higher TC and LDL-C levels compared to control as well as to MUFA and MUFA–SAFA groups (p < .05). At the same time, HDL-C concentrations were significantly elevated in SAFA group in comparison with control and MUFA–SAFA-fed animals, while the MUFA-rich diet group exhibited the lowest TG levels (p < .05 versus SAFA).
Moreover, testosterone levels were not statistically significantly different among the four experimental groups at the time of euthanasia. Serum AGE levels in high-fat-fed animals were comparable across the different experimental groups. Notably, similar to the lack of statistical significance in comparison to CML levels, no difference was observed in the peroxide concentrations at the end of experimentation.
Morphometric evaluation of seminiferous tubule diameter and basement membrane thickness
Somatometric, metabolic, hormonal, and oxidative stress parameters
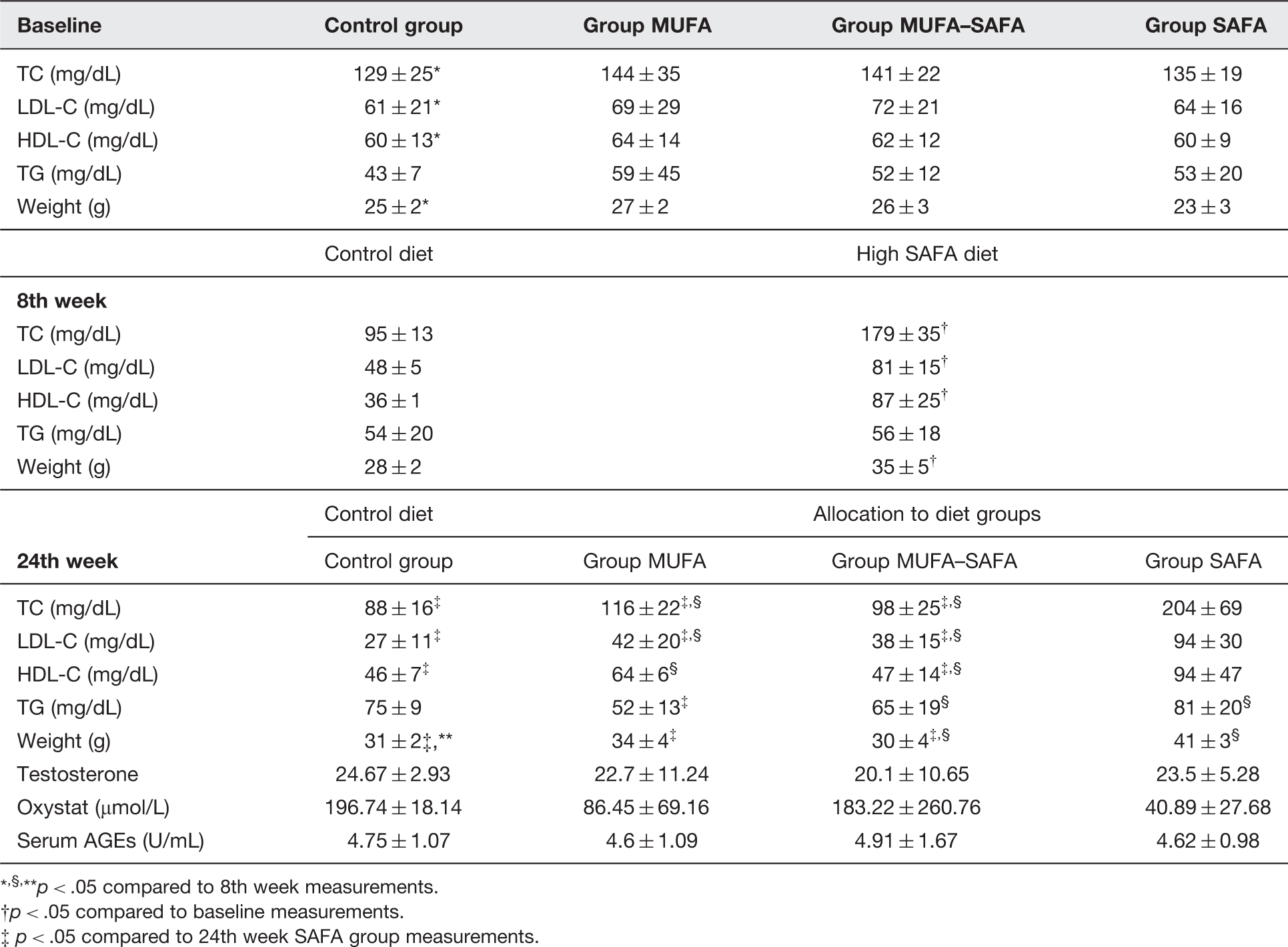
,§,**p < .05 compared to 8th week measurements.
p < .05 compared to baseline measurements.
p < .05 compared to 24th week SAFA group measurements.
Seminiferous tubule diameter, STBM thickness, prostate histologic changes, and apoptotic cell counts in the various experimental groups
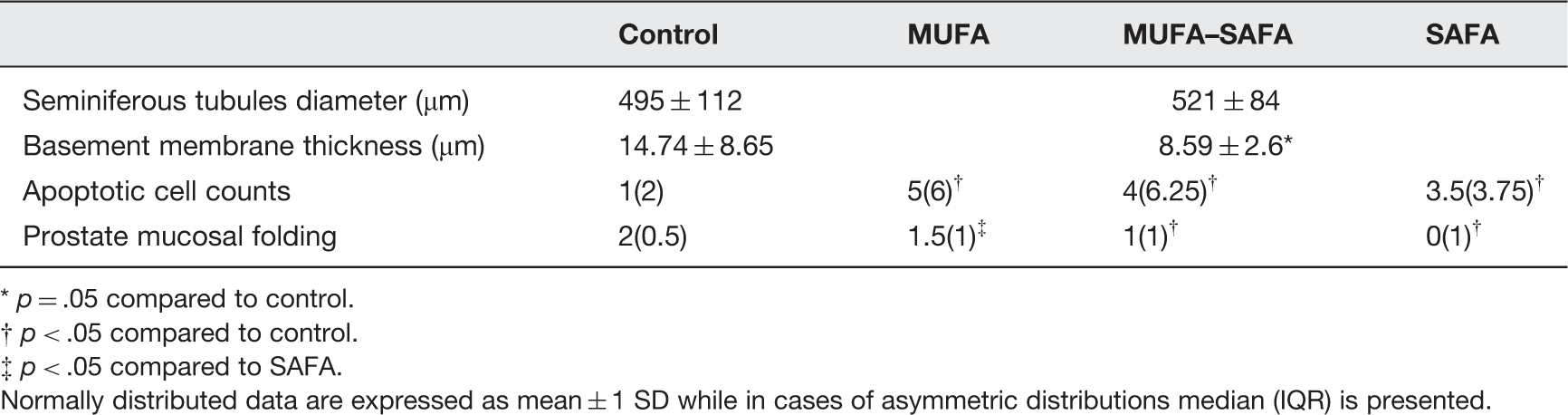
p = .05 compared to control.
p < .05 compared to control.
p < .05 compared to SAFA.
Normally distributed data are expressed as mean ± 1 SD while in cases of asymmetric distributions median (IQR) is presented.
Histological evaluation of prostate
In control animals, the glandular portion was of the tubuloalveolar type, presenting acini of different sizes with a normal folded mucosa showing tall columnar secretory epithelial cells, enriched by fibromuscular stroma.
The only difference among the various groups was the significantly decreased mucosal folding in the ventral lobe of all the high-fat-fed animal groups (p < .01) Figure 2. More specifically, the glandular mucosa was found less folded in MUFA–SAFA and SAFA group than that observed in control mice (p = .032 and p = .012, respectively), while the adherence to MUFA diet resulted in significantly increased level of folding compared to SAFA group (p = .015, Table 3). The secretory epithelium consisted mostly of tall columnar cells, with basal (peripheral acini) or apical (distal acini), round-shaped, clear nuclei, while no atrophy of the cytoplasm of the cells lining the lumen of the gland was observed. No alteration in the ratio of epithelial-to-stromal component of prostate glands was observed in high-fat-fed animals compared to control group (Figure 2, Table 3). Furthermore, prostate mucosa folding was positively correlated with lipid parameters and in particular TC levels (r = –0.28, p < .05).
Representative photomicrographs of hematoxylin–eosin-stained sections of mice prostate. (Aa) Control group ventral lobe prostatic section. (Bb) SAFA group ventral lobe prostatic section. Decreased mucosal folding in SAFA mice compared to control is observed (×200 magnification). (A color version of this figure is available in the online journal.)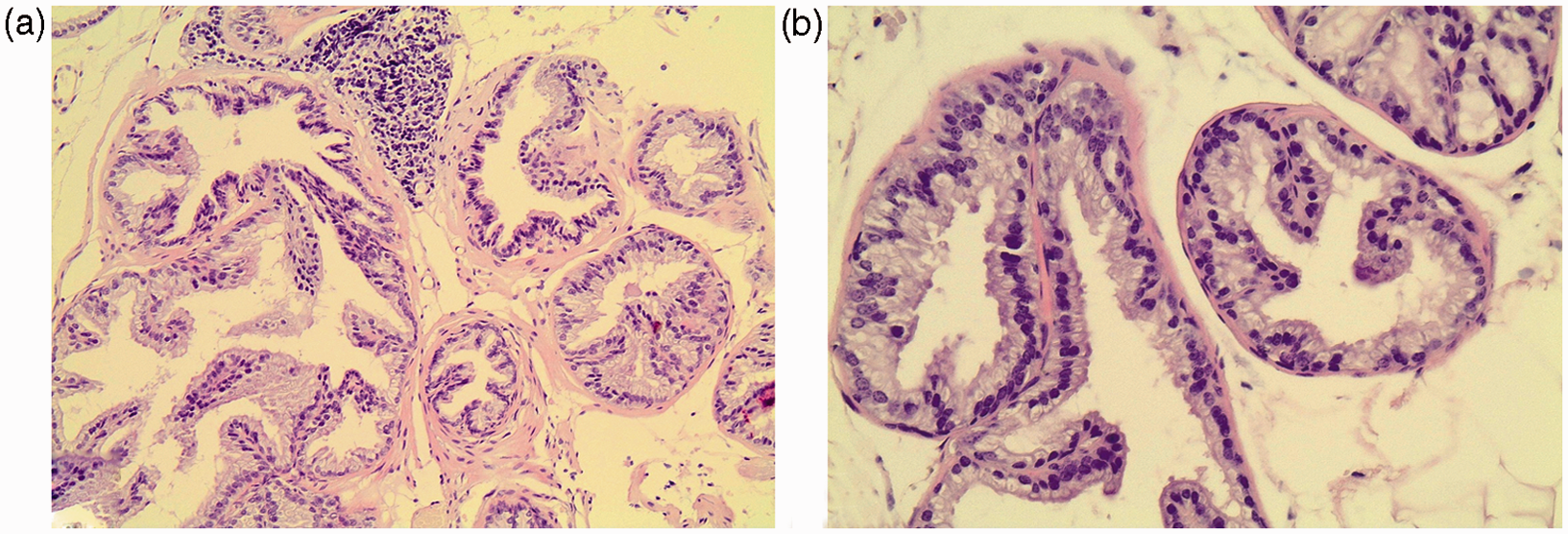
Evaluation of apoptosis
Overall, in the control group the number of apoptotic cells was rather small. In the entire population of high-fat-fed animals, apoptotic cells showed a predilection for the early stages of spermatogenesis and their number was significantly higher compared to control mice testis (p < .005) as well as in each experimental group with respect to the control mice (MUFA p = .012, MUFA–SAFA p = .026, SAFA p = .012, Figure 3, Table 3).
Representative photomicrographs of anti-ssDNA-stained sections of mice testes. (a) Control group. (b) MUFA group. (c) MUFA–SAFA group. (d) SAFA group. Increased number of apoptotic cells was identified in high-fat-fed animals compared to controls (×200 magnification). (A color version of this figure is available in the online journal.)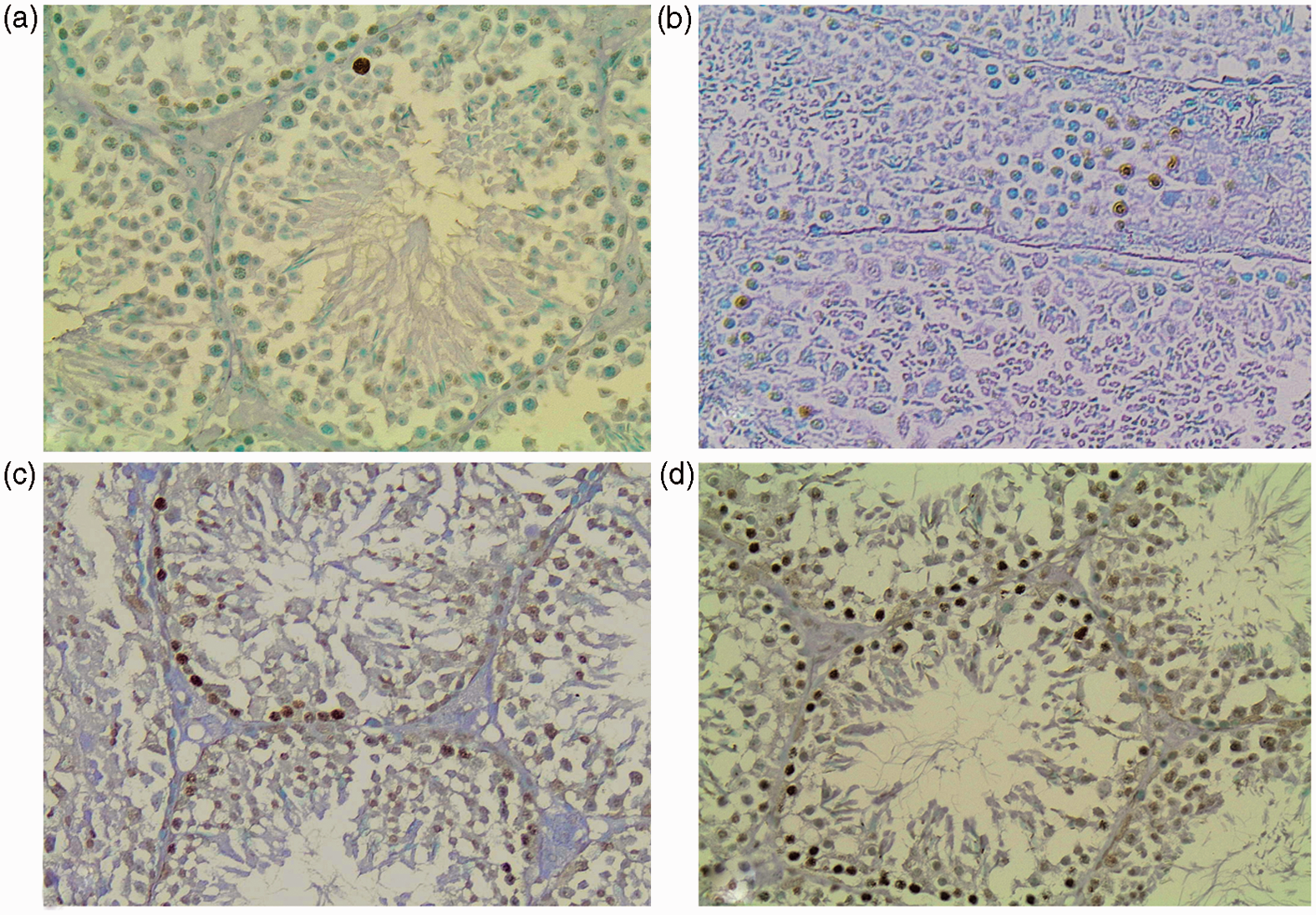
Immunohistochemical evaluation of AGE in testes, epididymis, and prostate
The immunocytochemical localization of AGE in the testicular tissue revealed a uniform staining pattern in the cytoplasm of all types of germ cells and Leydig cells of both control and high-fat-fed groups. The intensity of AGE staining was stronger in the spermatic epithelium close to the basement membrane.
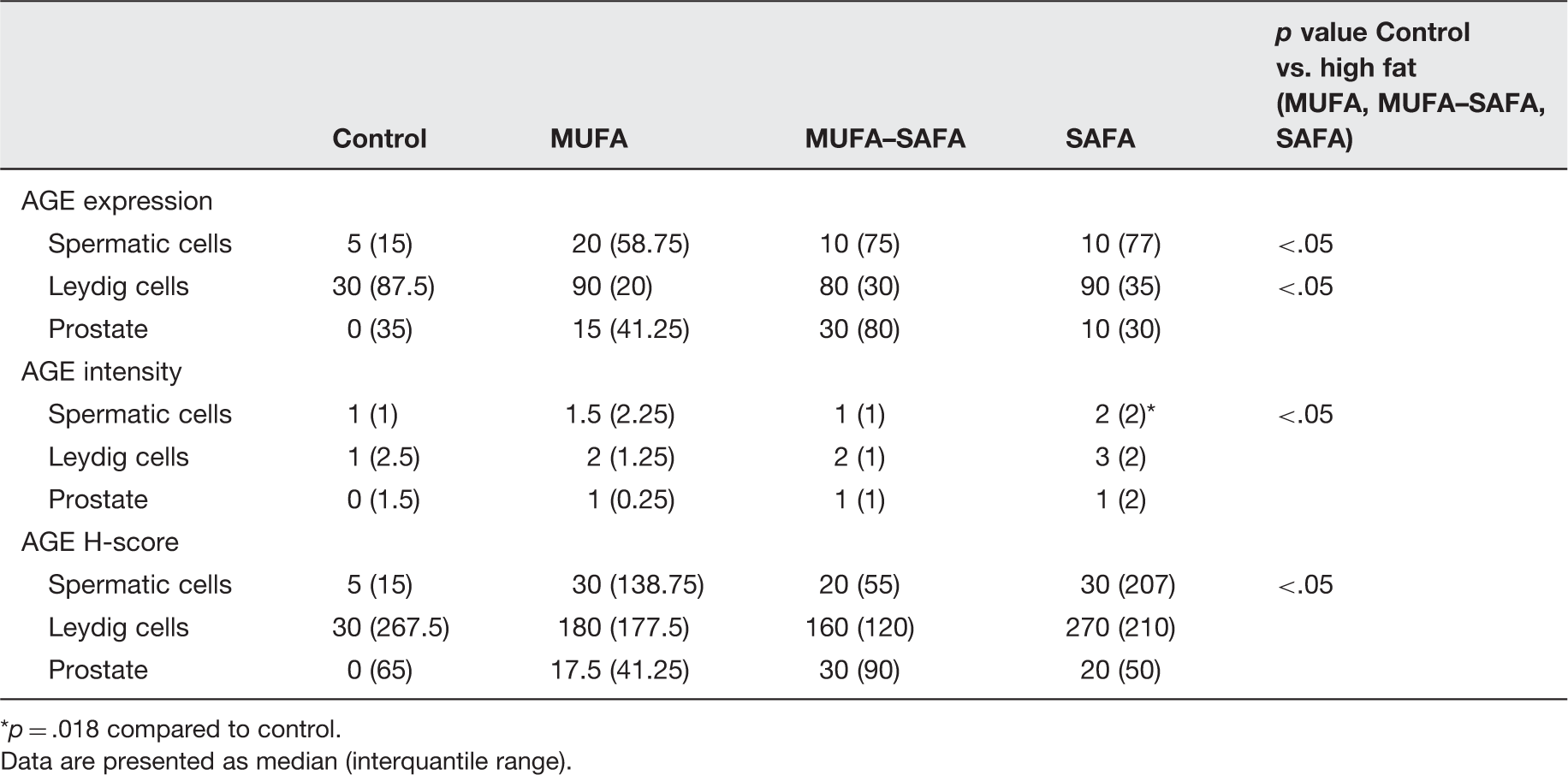
AGE expression in spermatic epithelium was strongest in SAFA followed by MUFA and MUFA–SAFA groups compared to control samples. Intensity of AGE staining was more potent in SAFA versus control (p = .018). However, overall in high-fat-fed mice considered as one group, AGE expression was more prominent both in terms of intensity and H-score (p < .05 for both comparisons) compared to control mice (p < .05). Furthermore, the percentage of AGE-positive Leydig cells was higher in the entire cohort of high fat diet animals compared to controls (p < .05, Figure 4, Table 4). Upon completion of 16 weeks of MUFA–SAFA diet, a reduction of AGE levels comparable to controls was observed.
Representative photomicrographs of AGE staining in testicular and prostatic tissue of mice. (a) Control group testes. (b) SAFA group testes. AGE staining in spermatic cells is more prominent in SAFA, compared to control animals. (c) Control group prostate. (d) SAFA group prostate. AGE positive secretory cells are seen only in high-fat-fed animals (×200 magnification). (A color version of this figure is available in the online journal.) Immunohistochemical data of AGE staining in testicular and prostate tissue of experimental groups p = .018 compared to control. Data are presented as median (interquantile range).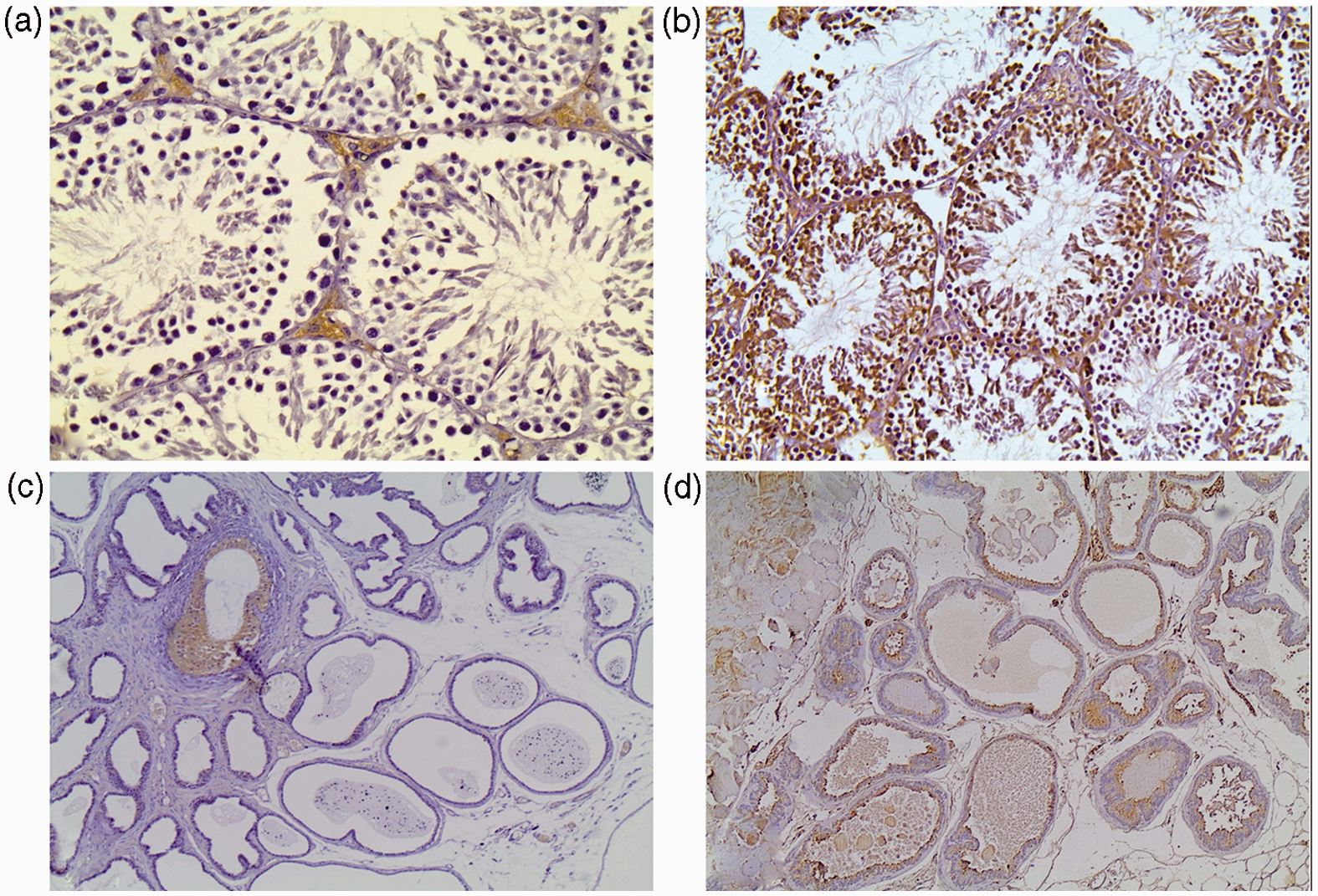
Epithelial epididymal cells were distinctively immunoreactive for AGE, with a cytoplasmic pattern and weaker intensity close to the basement membrane. A faint immunoreactivity was also noted in the muscle coat of epididymal duct.
The intensity of AGE immunostaining in germ cells showed positive correlation with different facets of the lipid profile. In particular, significant correlations were observed with TC (r = .37, p < .01), HDL-C (r = .35, p < .01), and LDL-C (r = .35, p = .01). Moreover, a significant correlation of AGE expression in spermatic epithelium with the body weight of the experimental animals was noted in terms of intensity (r = .42, p < .001) and H-score (r = .27, p < .05). AGE immunoreactivity in Leydig cells was negatively correlated with TG (r = −0.39, p = .005 and r = −0.36, p < .01, respectively) and positively with body weight (r = .28, p = .05).
In prostate tissue, the epithelial compartment displayed a uniform cytoplasmic pattern of AGE staining with higher intensity in luminal cells (Figure 4, Table 4). Various degrees of immunoreactivity among different groups were observed with a trend of increased deposition in high-fat-fed animals compared to controls.
Immunohistochemical evaluation of RAGE in testes, epididymis, and prostate
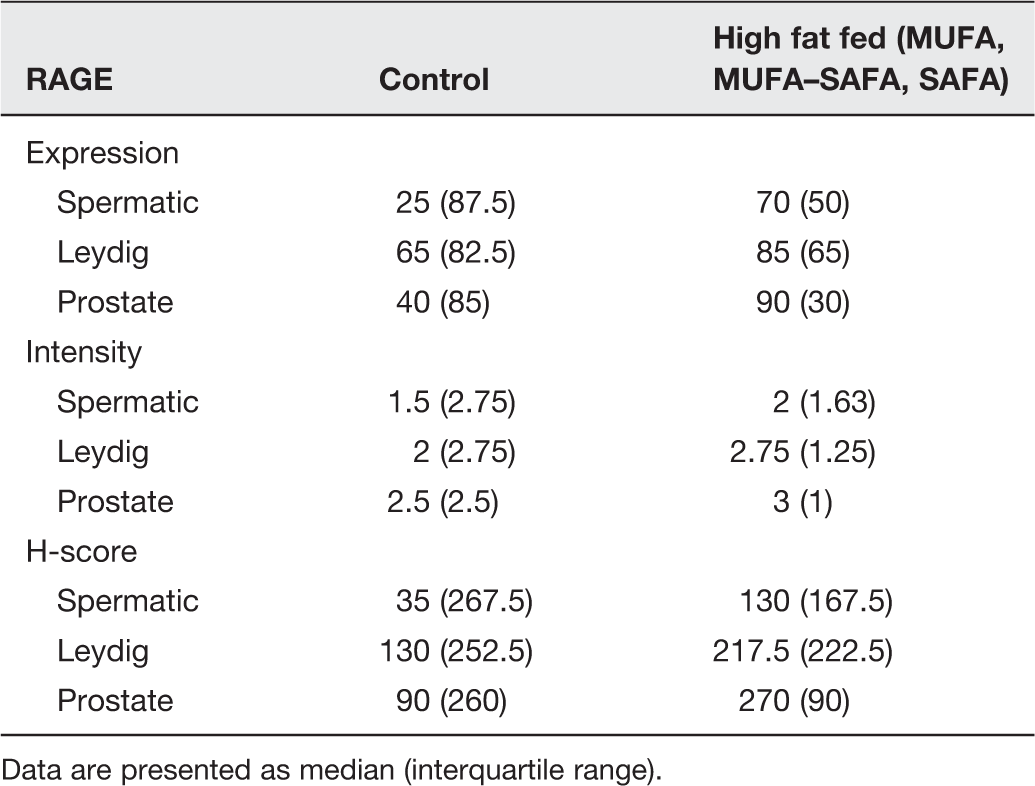
Similar to AGE localization in testis, cytoplasmic RAGE expression was observed in Leydig cells and the spermatic epithelium. Leydig cells also displayed strong membranous expression. RAGE expression levels in both testicular and epididymal epithelium did not significantly differ across the various diet groups.
In prostatic tissue RAGE immunoreactivity, similar to AGE accumulation, was more prominent along the luminal side (Figure 5, Table 5).
Representative photomicrographs of RAGE staining in testicular and prostatic tissue of mice. (a) Control group testes. (b) SAFA group testes. RAGE expression is observed in both the spermatic epithelium and in Leydig cells. (c) Control group prostate. (d) SAFA group prostate. RAGE is detected along the luminal border of the secretory cells (×200 magnification). (A color version of this figure is available in the online journal.) Immunohistochemical data of RAGE staining in testicular and prostate tissue of experimental groups Data are presented as median (interquartile range).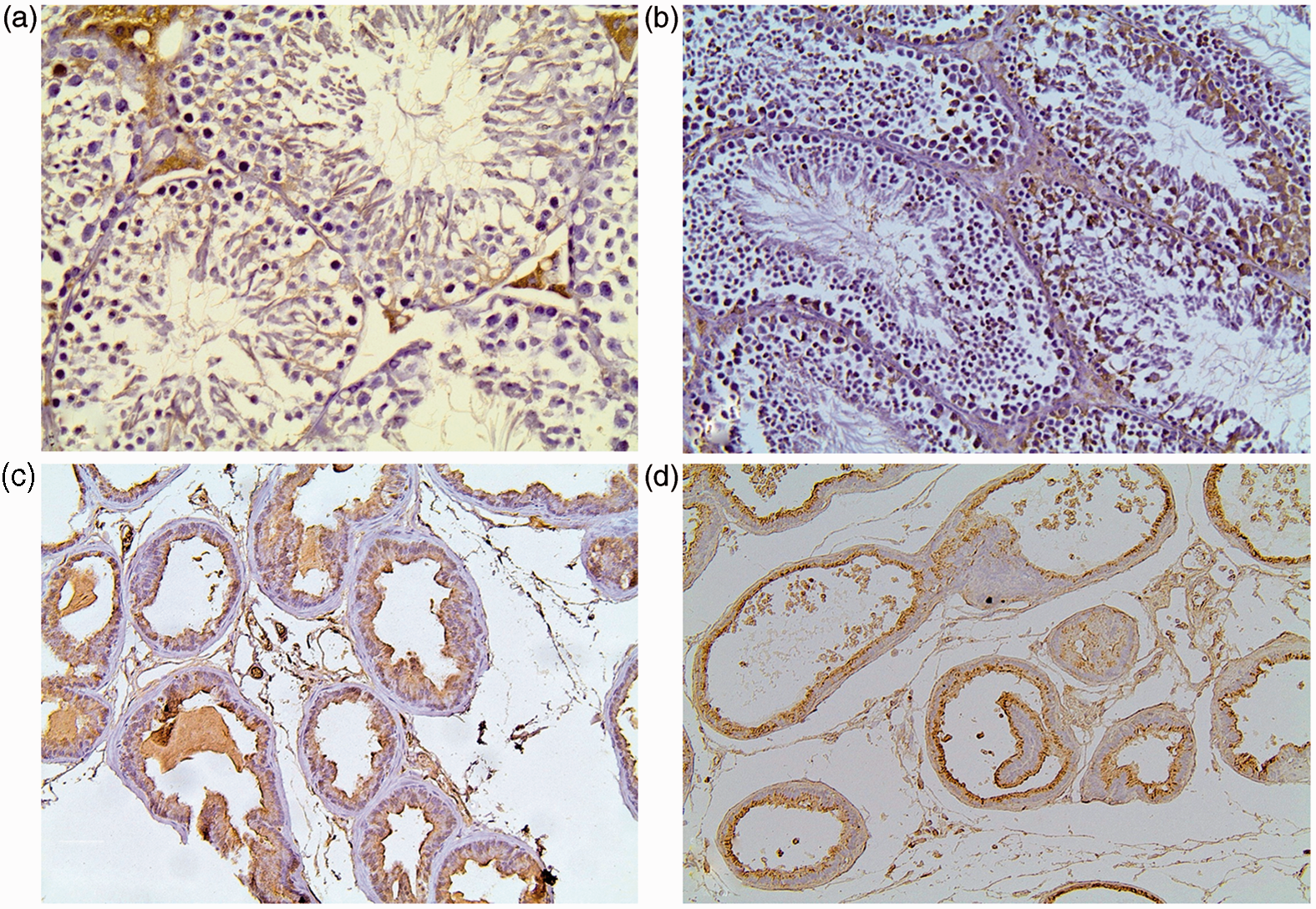
Discussion
The present study demonstrated for the first time that normal C57BL/6 mice exposed to Western diet for 24 weeks, known to induce obesity, exhibit increased levels of reactive metabolic by-products (AGE) deposited in spermatic epithelium and Leydig cells. Although spermatogenic activity was not affected, enhanced apoptosis was observed. Interestingly, administration of healthier diets, after high SAFA chow for 8 weeks did not manage to reduce AGE levels. Furthermore, morphologic alterations were also detected in prostatic tissue, mainly involving reduction in prostatic epithelial infolding of the glands, which was significantly reversed after MUFA diet administration.
The genetically unmodified C57BL/6 mouse model was used in our study, as it was previously established to be particularly sensitive in developing features of human obesity and MetS 20 when given ad libitum access to chow comprising high levels of SAFA representative of Western diet. The metabolic profile of these animals resembles the complex human disorder characterized by abdominal fat accumulation, gradual onset of insulin resistance, and increased feed efficiency (weight gained/kcal consumed), named passive overconsumption. In accordance, mice fed with a high SAFA diet in our study had significantly higher body weight, TC, and LDL-C levels compared to controls. A diet rich in MUFA instead of SAFA or with balanced MUFA–SAFA composition resulted in lipid profile restoration to control group values. The adverse effects of diet-induced obesity on the male reproductive system were extensively studied in C57BL/6 mice, which were found to have decreased sperm concentration and motility, 21 reduced fertilization rate, 22 number of embryos, and pregnancy rate. 23 The subfertility phenotype reportedly seems to be transferred in two subsequent generations of offsprings when raised on a control diet. 24
A plausible explanation for reduced fertility in the context of obesity could be hypogonadism. However, testosterone levels did not seem to differ between our experimental study groups. This observation is in agreement with the results of Bakos et al. 21 who found that paternal diet-induced obesity affected multiple aspects of spermatogenesis as well as fertility status in the absence of altered androgen levels. Human studies are somewhat inconclusive on that issue with the majority reporting hypogonadism related to obesity, while others show increased bioavailable testosterone and estradiol on obese nonfertile male patients 25 or reproductive hormone levels not statistically affecting the association between body mass index and semen quality. 26 It is, however, possible that either the harmful effects of obesogenic diet are not discernible on endocrine profile or other endocrine abnormalities (e.g. leptin and insulin levels) are involved in reproductive dysfunction. 22
Differences in seminiferous tubules diameter was not identified between the various diet groups. Nonetheless, our data showed decreased basement membrane thickness in testes of high-fat-fed animals compared to controls without an effect on spermatogenesis. The aging effects on control animals and the lack of deleterious diabetic environment in obese mice could be a reasonable explanation for this observation. 27
Interestingly, investigation of the subcellular effects of diet-induced obesity οn the spermatic epithelium of the experimental mice revealed increased number of apoptotic nuclei in the early stages of spermatogenesis compared to controls. This was performed by immunohistochemical detection of ss DNA regions in apoptotic nuclei that enabled us to assess apoptosis in the testicular microenvironment reliably, without the need for sophisticated in situ hybridization techniques. Apoptosis, a fundamental induced cell death process, is involved in every step of germ cell development. 28 The inherent sensitivity of germ cells to apoptosis renders them particularly vulnerable to environmental triggers, like pollutants or oxidative stress. Apoptosis during spermatogenesis has been assumed to play a role in male infertility, 29 and increased apoptotic index has been previously observed in the seminiferous tubules of several animal models.27,30
Oxidative stress is a major factor underlying the systemic effects of obesity and MetS, thereby contributing to the generation of reactive AGE. These highly reactive compounds are generated under physiological conditions by the irreversible reaction of proteins, lipids, or nucleic acids with carbohydrates in metabolically active cells. AGE exert their deleterious effects by directly modifying proteins through extensive crosslinking or indirectly through interaction with their specific receptor (RAGE) in the cell surface. This receptor–ligand interaction leads to increased reactive oxygen species formation and cell activation inducing a vicious cycle of inflammation, cellular dysfunction, and tissue damage.
Our data revealed increased AGE deposition in germ cells of SAFA mice compared to controls. In accordance with previous studies,12–14 AGE were detected throughout the cytoplasm of spermatic epithelium. The abundance of fructose and glucose as energy source for the maturing gametes and the high levels of fatty acids in spermatogenic cell membrane render them highly susceptible to glycation. This was further confirmed in our study by the significant correlation of AGE staining in germ cells with TC, HDL-C, and LDL-C levels as well as the body weight of experimental animals. A marked reduction in the intensity of AGE staining was observed along with the maturation of germ cells, AGE immunoreactivity being absent in the luminal space of the tubules. Nuclear AGE immunoreactivity was only noted in a small number of germ cells close to the basal membrane, while extensive staining of premitotic cells nuclei has been reported in human tissue. 12 AGE accumulation, featuring one of the most important posttranslational modifications of proteins, lipids, and nucleic acids, could represent one of the epigenetic sperm alterations previously suggested to be responsible for the transmission of the impaired reproductive potential related to high fat diet in two generations of descendants. 24
In the interstitial testicular tissue, accumulation of AGE was distinctively recognized in Leydig cells. Taking into consideration the fundamental role of Leydig cells in the endocrine function of male gonad, a possible effect of glycation on their functional integrity could be postulated. This finding is consistent with previous reports showing increased Nɛ-carboxyethyl-lysine (CEL) and CML localization being associated with reduced numbers of Leydig cells in diabetic rats compared to age-matched control, presumably due to AGE intoxication. 31 In contrast, Mallidis et al. detected only RAGE in Leydig cells but not CML.12,15 Notably, in our study AGE staining was higher in Leydig cells of SAFA and MUFA animals compared to controls, while MUFA–SAFA diet after 8 weeks restored levels comparable to control. The intracellular supplies of lipids necessary for steroidogenesis may be the source of lipid-derived glycation adducts interrelated with the circulating lipids as a result of the variability in fatty acid composition of diet regimens.
The co-localization of RAGE with accrued AGE in seminiferous tubules, especially in early stages of germ cell maturation, and interstitial compartment of testes implies interaction of receptor with ligands. Activation of AGE–RAGE axis results in perturbation of cellular processes, initiation of pro-inflammatory and pro-apoptotic signaling, and induction of further AGE production through oxidative stress. It would be tempting, therefore, to speculate that AGE accumulation in testes of obese mice through binding to RAGE results in apoptosis of germ cells in initial phases of their development, affecting pluripotent stem cells and thereby reproductive capacity. Nevertheless, the potential relationship remains to be elucidated in future studies employing molecular analysis approaches.
Regarding the prostate of high-fat-fed mice, reduction in epithelial folding observed in SAFA and MUFA–SAFA animals implies that the content of SAFA in diet induces histological alterations. In support of this notion, a positive correlation of mucosal infolding with facets of the lipid profile was demonstrated. The observed structural changes, given the absence of epithelial atrophy or alteration in epithelium-to-stroma ratio, could be perceived as early morphological modifications involved in organ’s aging. 32 The acceleration of aging process is well known to interfere with carcinogenesis in various tissues. Our present observation is in harmony with existing evidence that not only the excess of fat but also the fatty acid composition of diet affects the microenvironment of prostate.33–35
Possible mechanisms underlying abnormal prostate growth and structure, given the absence of hypogonadism as a result of obesity, could involve sustained systemic inflammation and oxidative stress response locally. AGE, a common denominator of metabolically triggered tissue stress were for the first time found to be deposited in prostate epithelium corresponding to the distribution reported for RAGE. The implication of AGE–RAGE axis in prostate homeostasis and pathophysiology remains elusive.
The nonsignificant elevation of peroxides and AGE found in serum may explain the lack of robust difference in AGE accumulation or morphologic alterations of both testes and prostate of experimental animals. Without their contribution, an oxidative environment may not be established or adequately augmented to the extent necessary to cause systemic damage and subsequently alterations in prostate gland. However, the higher than usual metabolic rates in testicular microenvironment that are extremely favorable for the accrual of AGE induce increased level of local stress affecting structural and functional status of the male gonad.
In conclusion, high fat diet for 24 weeks hastened AGE deposition in germinal epithelium and Leydig cells of mice. Moreover, it increased apoptosis of spermatic epithelium. The cytoarchitecture of mice prostate significantly changed as a result of high SAFA or MUFA–SAFA diet compared to control, while MUFA diet restored the control phenotype. Further studies are required to clarify the role of AGE–RAGE interplay in the reproductive system of individuals with chronic metabolic disturbances. The clinical importance of our preliminary findings, if confirmed in large human studies, could obviate refinement of management of reproductive abnormalities and prostatic disorders as a result of obesity. Simple dietary intervention toward a Mediterranean regimen with high content of MUFA could reverse the metabolic perturbation caused by Western diet, thereby ameliorating gonadal function and fertility. Finally, targeting AGE–RAGE signaling pathway in tissues may prove beneficial in either treating or even preventing obesity effects before their overt manifestation without affecting hormonal milieu.
Footnotes
Author contributions
CP and PK contributed equally to this work. All authors participated in the design, interpretation of the studies, and analysis of the data and review of the manuscript. Study design: DPM, DP, DM, EDK, AGP. Conducted experiments: DPM, CP, IV, CA, PK, GL, EAT. Data collection, analysis, interpretation: DPM, IV, GL, PK, CP. Manuscript writing: DPM, CP, PK. Manuscript editing and study supervision: DP, DM, EDK, AGP.
ACKNOWLEDGEMENTS
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.