
Abstract
The tumor necrosis factor receptor-associated death domain protein (TRADD) regulates cell proliferation and apoptosis via tumor necrosis factor alpha (TNF-α)-mediated signaling pathways. Low levels of TRADD expression may result in the excessive proliferation of hypertrophic scar fibroblasts (HSFb). This study investigated the effects of a lentiviral vector carrying the human tradd gene on the proliferation, apoptosis and type I collagen synthesis of HSFb and embryonic fibroblasts (EFb) and further explored the resulting effects on hypertrophic scars (HS). We utilized cytoimmunofluorescence and Western blotting to confirm the expression of TRADD in HSFb and EFb. A PLVX-TRADD-EGFP lentivirus was prepared and transfected into EFb and HSFb, and then the expression of a TRADD-GFP-FLAG fusion protein was detected in HSFb and EFb. After stimulation with 10 ng/mL TNF-α, cell proliferation, apoptosis, and the synthesis of type I collagen were assessed. Our results show that the expression level of TRADD was significantly lower in HSFb than in EFb. A biologically active PLVX-TRADD-EGFP lentivirus was constructed and transfected into HSFb and EFb. The TRADD-GFP-FLAG fusion protein was effectively expressed in HSFb and EFb. Either alone or in combination with 10 ng/mL TNF-α, the PLVX-TRADD-EGFP lentivirus inhibited proliferation, caused a G2/M phase arrest, induced the appearance of a sub-G1 apoptotic peak and inhibited the secretion of type I collagen by HSFb without significantly affecting EFb. These results suggest that the low expression of TRADD in HSFb is a principal reason for their excessive proliferation. The transfection of a PLVX-TRADD-EGFP lentivirus led to the normal expression of TRADD in HSFb. When combined with 10 ng/mL TNF-α, a PLVX-TRADD-EGFP lentivirus transfection could inhibit cell proliferation, promote apoptosis, and reduce the secretion of type I collagen in HSFb, thereby reducing HS formation.
Keywords
Introduction
Hypertrophic scars (HS) are relatively common pathological scars in clinical practice. HS not only affect the appearance of damaged tissues but can also cause deformities and dysfunction, such as skin keloids and esophageal anastomotic strictures. HS-associated pathological changes mainly include fibroblast activation, excessive proliferation, excessive extracellular matrix secretion, excessive collagen synthesis, and the sedimentation of large quantities of collagen fibers. 1 Currently, there are two main categories of treatment approaches for HS: surgical therapy and non-surgical therapy. Surgical resection is the common method of clinical treatment for HS. The application of soft tissue dilation, skin grafts, flaps, and other repair technologies can significantly improve the appearance and functionality of lesioned areas. However, the recurrence rate is relatively high, most likely due to uncontrollable postoperative wound healing, leading to the reformation of pathological scars. Non-surgical treatments include compression therapy, local drug injections, laser therapy, and cryotherapy. Any non-surgical therapies either directly or indirectly act on regulating the critical processes of fibroblast proliferation and collagen metabolism. 2 Although there are many treatment approaches for HS, the results have not been satisfactory. Once formed, scars cannot be completely eliminated. Therefore, the prevention of HS formation is particularly important. The key to HS prevention is identifying scar-associated genetic defects and correcting them at the genetic level. This correction inhibits fibroblast proliferation and collagen synthesis at their source to prevent or reduce the occurrence and development of HS.
The tumor necrosis factor receptor-associated death domain protein (TRADD) regulates cell proliferation and apoptosis via tumor necrosis factor alpha (TNF-α)-mediated signaling pathways. Furthermore, the low expression of TRADD by hypertrophic scar fibroblasts (HSFb) 3 may inhibit the TNF-α-mediated apoptotic pathway, leading to the excessive proliferation of HSFb and the formation of pathological scars. TNF-α plays a critical role in fibroblast collagen synthesis. Following its induction by specific factors, collagen anabolism may begin to exceed collagen catabolism in fibroblasts, resulting in the excessive synthesis and deposition of type I collagen in the cellular matrix; this process forms the cellular basis of fibrotic diseases. 4 TNF-α maintains a homeostatic balance between collagen synthesis and degradation by increasing collagenase synthesis, thus promoting collagen degradation5,6 and antagonizing fibrosis.7,8 There are numerous positive and negative regulatory sequences upstream of the procollagen gene. This gene can be upregulated by factors such as transforming growth factor-beta 1, interleukin (IL)-4, IL-13, and insulin, and it is downregulated by factors such as TNF-α, interferon (IFN)-α, IFN-γ, and basic fibroblast growth factor (bFGF).9,10 Therefore, stable expression of the tradd gene after integration into the HSFb genome and subsequent culture of the cells under TNF-α inducing conditions, may increase HSFb apoptosis, reduce the secretion of type I collagen, and eventually suppress HS formation.
Lentiviral vectors are derived from human immunodeficiency virus-1 (HIV-1) and exhibit the characteristics of high infection efficiency, stable expression, weak autoantigenicity, strong controllability, and the accommodation of large fragments of exogenous target genes. 11 Lentiviral vectors have been considered the ideal gene transfer vector in recent years. For example, lentiviral-mediated introduction of the β-globin gene into mouse hematopoietic stem cells is highly efficacious for the treatment of β-thalassemia. 12 Research on cancer gene therapy has confirmed that lentiviral vectors possess advantages over other viral vectors. 13 Currently, antiproliferative stents have been generated by use of an antitissue proliferation coating to treat vascular scar stenosis, and they have been commercialized and applied clinically.14,15 Therefore, the precise positioning and transduction of a lentivirus carrying the tradd gene into the host cell genome, leading to stable expression of the exogenous gene and inhibition of proliferation and collagen secretion by HSFb, can be safely and effectively used in clinical practice.
For the first time, we used the lentiviral vector system to successfully construct a lentivirus carrying and overexpressing the human tradd gene. 16 However, its function requires further investigation. This study examined the effects of a lentiviral vector carrying the human tradd gene in combination with TNF-α on the proliferation, apoptosis, and synthesis of type I collagen by HSFb and embryonic fibroblasts (EFb), and it further explored the impact of this treatment on the inhibition of HS formation.
Materials and methods
Plasmids and cells
The tradd gene template was purchased from OPEN (Pittsburgh PA, United States), the lentiviral expression vector pLV X-EGFP-3FLAG-Puro was purchased from Shanghai SBO Medical Biotechnology (Shanghai, China), the ViraPower™ Lentiviral Packaging Mix was purchased from Invitrogen (Beijing, China), and DH5α competent cells, 293FT cells, and the EFb cell line were all from our laboratory. HSFb cells of passage 5 were derived from humans scar tissue in the popliteal fossa. The EFb and HSFb cells were cell lines and were cultured in DMEM medium containing 10% fetal bovine serum (FBS).
Main reagents
The DL2000 DNA Marker (D501A), agarose gel DNA extraction kit (DV805A), Taq enzyme, and dNTPs (DR001B) were purchased from TaKaRa (Dalian, China), EcoRI (R0101S) was purchased from New England Biolabs (Beijing, China), the Bicinchoninic Acid (BCA) Protein Assay Kit (23227) was purchased from HyClone-Pierce (Pittsburgh PA, USA), the prestained protein marker (SM1811) was purchased from Fermentas (Shenzhen, China), the rabbit antiTRADD antibody (Ab74320) was purchased from Abcam (Cambridge, MA, USA), the goat antirabbit IgG (Sc-2004) was purchased from Santa Cruz Biotechnology (Dallas, Texas, USA), and the Type I Collagen (Collagen I) ELISA Kit (F5700) was purchased from Shanghai Westang Biotechnology (Shanghai, China). The primers were synthesized by Shanghai Generay Biotech (Shanghai, China) and the positive clones were sequenced by the Beijing Genomics Institute (BGI).
Verification of TRADD expression in HSFb and EFb
Cytoimmunofluorescence
Coverslips were placed in each well of a 24-well plate. HSFb were resuspended in DMEM medium containing 10% FBS at 3.6 × 104 cells/mL, and 500 µL of this suspension was inoculated into each well (1.8 × 104 cells/well). Likewise, EFb were resuspended in DMEM medium containing 10% FBS at 1.56 × 105 cells/mL, and 500 µL of this suspension was inoculated into each well (7.8 × 104 cells/well). Then, the culture was terminated when the confluence rate reached 80%. The cells were washed for 3–5 min in phosphate buffered saline (PBS) three times, fixed in 4% paraformaldehyde for 20 min at room temperature, and washed three more times in PBS for 3–5 min. The cells were permeabilized in 0.5% Triton X-100 for 15 min, washed for 3–5 min in PBS three times, and blocked in 5% bovine serum albumin (BSA) for 30 min. Subsequently, the cells were incubated overnight at 4℃ with primary antibodies diluted 1:100 in 1% BSA and then washed for 3–5 min in PBS three times. Secondary antibodies diluted 1:50 in 1% BSA were added, the cells were incubated in the dark for 1 h at 37℃. After three washes in PBS for 3–5 min, the cells were stained with 5 µg/mL 4′,6-diamidino-2-phenylindole for 3 min, rinsed three times with PBS for 3–5 min, mounted in 95% glycerol and stored in the dark. The results were observed under a laser scanning confocal microscope. This procedure was performed in duplicate.
Western blotting
HSFb and EFb were harvested, transferred to Eppendorf tubes, lysed on ice for 10–15 min, and disrupted using a sonicator; the supernatant was removed after centrifugation. After the protein concentration was determined by the BCA method, the final protein concentration of each sample was adjusted to 2 µg/µL. Following conventional protein denaturation, the samples were subjected to gel electrophoresis and were wet transferred to a polyvinylidene difluoride (PVDF) membrane. The membrane was blocked in 5% fat-free milk at room temperature for 1 h, incubated with a primary antibody (1:200 rabbit antiTRADD) overnight at 4℃ and washed three times for 10 min in TBST (0.05% Tween-20). The PVDF membrane was incubated with a secondary antibody (1:3000 goat antirabbit IgG) at room temperature for 2 h, followed by three 10-min washes in TBST. Finally, a chromogenic reaction was performed using the ECL Plus Kit, then the X-ray film was exposed, developed, fixed, and analyzed. Quantity One 4.6.2 image processing software was used to measure the integrated optical density (IOD) of the electrophoretic bands to calculate the ratio of the optical density of the target protein to that of the internal reference. This value was used to represent the relative expression level of the target protein. The procedure was performed in duplicate.
Acquisition of the tradd target gene and the construction and identification of the lentiviral expression vector 16
Expression vector linearization and vector map in Figure 1 show that the expression vector was digested with EcoRI, and a vector fragment of approximately 8.8 kb was recovered after the digested products were electrophoresed. The tradd target gene was polymerase chain reaction (PCR) amplified using primers with the following sequences: tradd-EcoRI-F, 5′-CTCAAGCTTC Vector map. (A color version of this figure is available in the online journal.)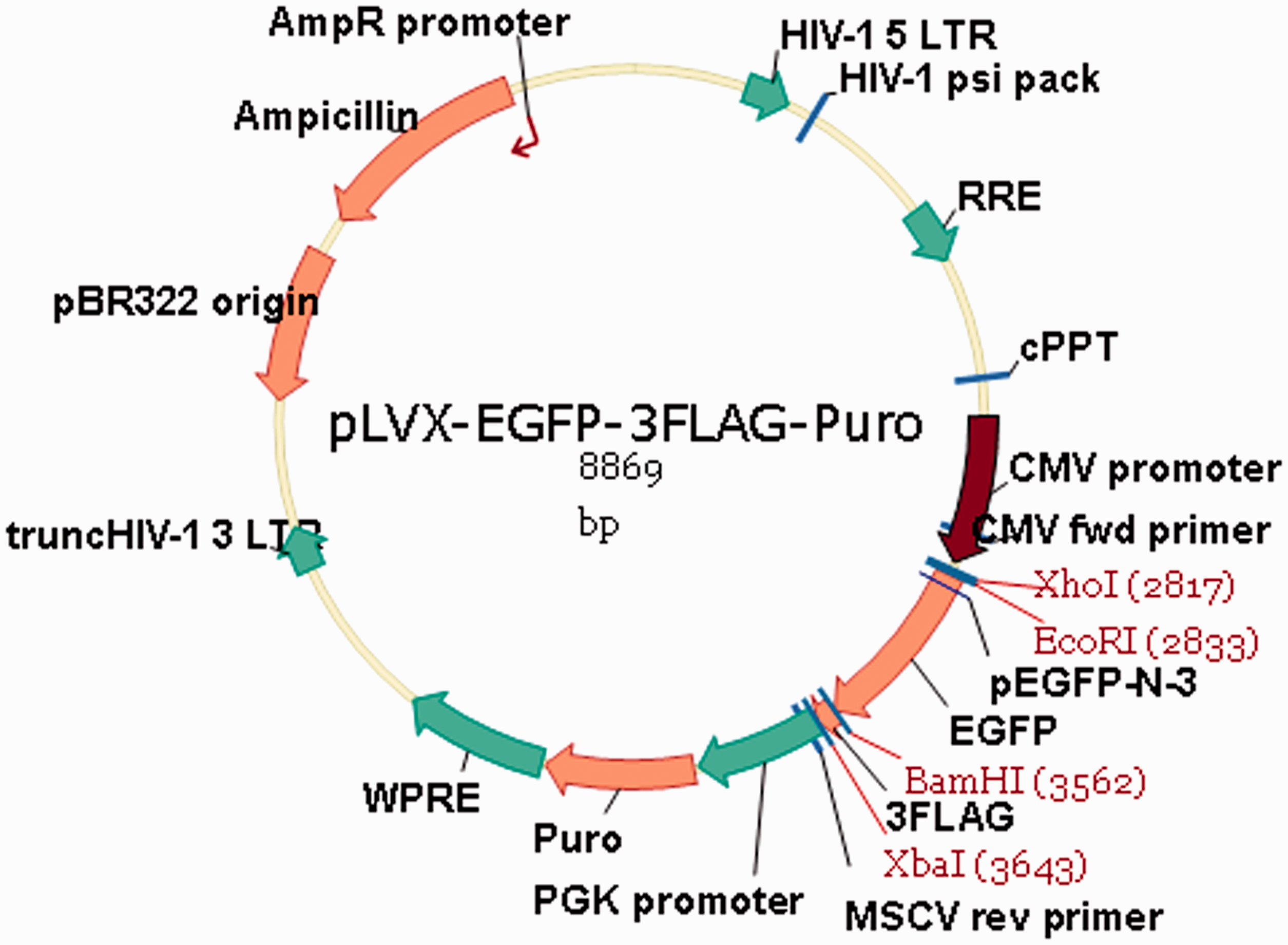
Western blot detection of the TRADD-GFP-FLAG fusion protein in HSFb and EFb after lentiviral transfection
Preliminary results showed that a multiplicity of infection of 100 was required for the infection efficiency of PLVX-TRADD-EGFP lentivirus and PLVX-EGFP lentivirus to reach 80%. Three groups of cells were included in this experiment: untransfected cells (blank control group), cells transfected with PLVX-TRADD-EGFP lentivirus (experimental control group), and cells transfected with PLVX-EGFP lentivirus (experimental group). Cell collection, protein extraction, and Western blotting were performed as described above. The procedure was performed in duplicate.
MTT assay of HSFb and EFb proliferation
Black control, experimental control, and experimental HSFb were resuspended in DMEM medium containing 10% FBS at 1.11 × 104 cells/mL, and 90 µL of this suspension was inoculated into each well of five 96-well plates (1.0 × 103 cells/well). After the cells became adherent 12 h later, 10 µL of 100 ng/mL TNF-α was added to each corresponding well (blank control group + TNF-α and experimental group + TNF-α) for a final concentration of 10 ng/mL. Each group included six duplicate wells. EFb cells (from both the control and experimental groups) were resuspended in DMEM medium at 2.22 × 104 cells/mL, and 90 µL of this suspension was inoculated into each well (2.0 × 103 cells/well). After the cells became adherent, 10 µL of 100 ng/mL TNF-α was added to each corresponding well (blank control group + TNF-α and experimental group + TNF-α) for a final concentration of 10 ng/mL. Each group included six duplicate wells. The OD values were measured at 490 nm. The procedure was performed in duplicate.
Flow cytometry detection of HSFb and EFb cell cycle and apoptosis
HSFb and EFb were infected with the PLVX-TRADD-EGFP and PLVX-EGFP lentivirus. After three days, TNF-α was added to the cells at a final concentration of 10 ng/mL. Six groups of cells were included in this experiment: the blank control group, blank control cells treated with TNF-α (blank control group + TNF-α), the experimental control group, experimental control cells treated with TNF-α (experimental control group + TNF-α), the experimental group, and experimental cells treated with TNF-α (experimental group + TNF-α). Cells exhibiting active proliferation capacity were harvested after 72 h of treatment. After the cell numbers were counted, 106 cells from each group were resuspended and fixed in precooled 70% ethanol at 4℃ for 2 h. After filtering through 300-mesh screen, the cell suspensions were transferred to a flow cytometry tube and examined. The procedure was performed in duplicate.
ELISA detection of collagen synthesis in HSFb and EFb
HSFb and EFb were infected with the PLVX-TRADD-EGFP and PLVX-EGFP lentivirus. After three days, TNF-α was added to the cells at a final concentration of 10 ng/mL, and the culture was continued. Six groups of cells were included in this experiment: the blank control group, blank control cells treated with TNF-α (blank control group + TNF-α), the experimental control group, experimental control cells treated with TNF-α (experimental control group + TNF-α), the experimental group, and experimental cells treated with TNF-α (experimental group + TNF-α). Cell culture medium was collected from each group 24 h, 48 h, and 72 h after treatment. The human type I collagen (Collagen I) ELISA kit was used to measure the OD at 450 nm. A standard curve was generated by plotting the standard concentration on the vertical axis (logarithmic scale) and the standard OD value on the horizontal axis (logarithmic scale). The standard curve was used to calculate the collagen I concentration of the sample based on its OD value. The procedure was performed in duplicate.
Results
Verification of TRADD expression in HSFb and EFb
Cytoimmunofluorescence
Cells growing on coverslips were examined by confocal microscopy. TRADD, which was labeled with a green fluorescent antibody, was scattered throughout the cytoplasm of HSFb and EFb, indicating its effective expression and revealing its location within the cell. The fluorescence intensity was measured directly by scanning confocal microscopy. The results showed that the TRADD expression was significantly lower in HSFb than in EFb (Figure 2).
TRADD expression in EFb and HSFb. (a) Fluorescence intensity of TRADD expression in EFb, (b) fluorescence intensity of TRADD expression in HSFb. (A color version of this figure is available in the online journal.)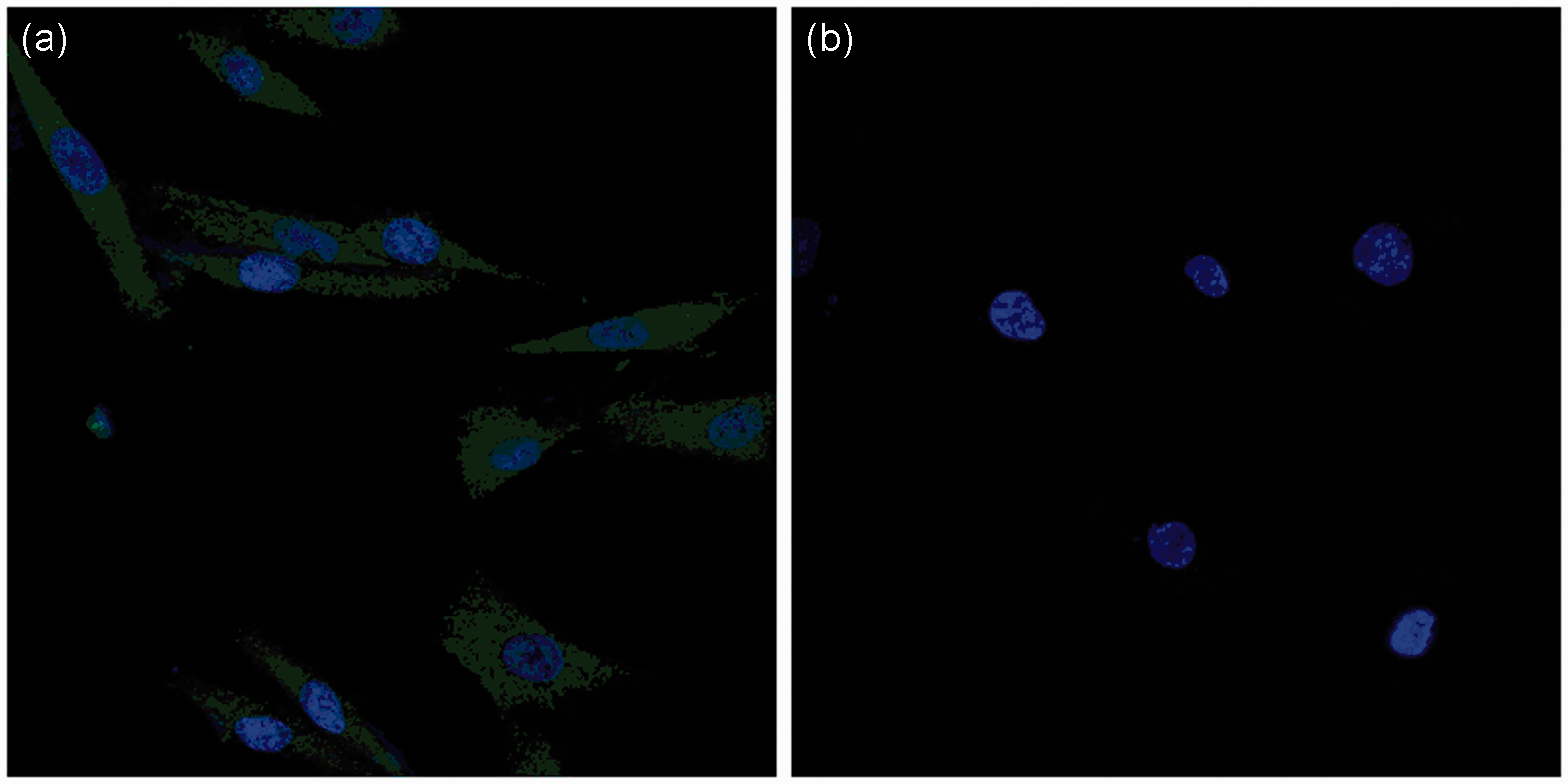
Western blot
The ratio of the TRADD target protein IOD to that of the internal reference, β-actin was calculated to indicate the relative expression level of TRADD. The experimental results were consistent with the cytoimmunofluorescence results, and the difference between the two groups was statistically significant (P < 0.05) (Figure 3).
Differences in TRADD expression levels between EFb and HSFb. Total protein was extracted from EFb and HSFb. The samples were prepared and run on an SDS-PAGE gel, transferred to a membrane, incubated with antibody, developed and fixed. The IOD value was measured using an image processing system, and the ratio of the TRADD electrophoretic band IOD to that of the internal reference, β-actin, was calculated to represent the relative expression level of TRADD. TRADD expression in HSFb was significantly lower than in EFb (aP < 0.05)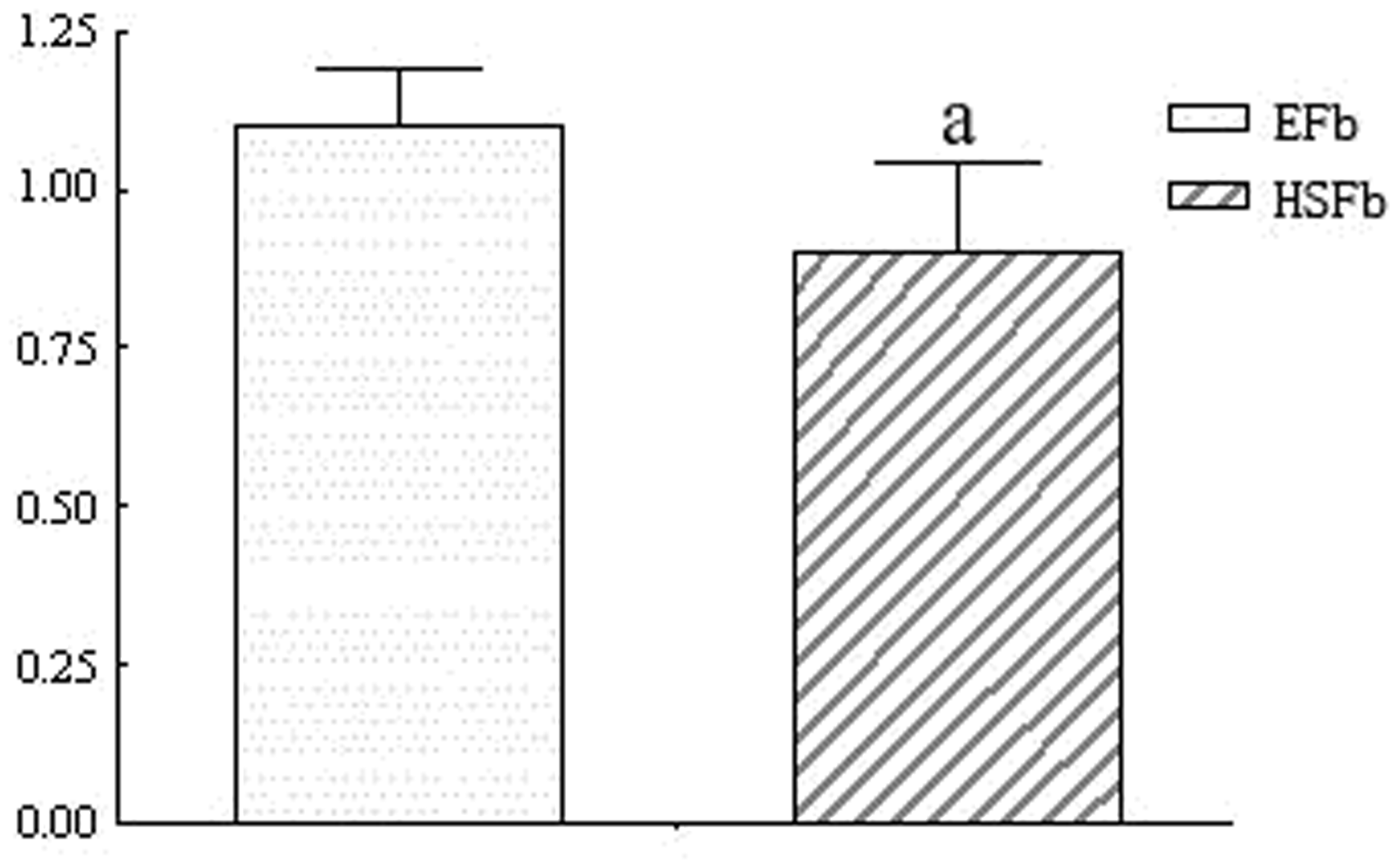
Western blot detection of fusion protein expression after lentiviral infection of HSFb and EFb cells
The tradd target gene was fused to gfp and flag. The theoretical size of the co-expressed TRADD-GFP-FLAG fusion protein is 64 kDa. With β-Actin as an internal reference, Western blotting demonstrated that the experimental group had a positive band at 55–72 kDa, which was consistent with the theoretical value of the fusion protein. The blank control group and the experimental control group had no positive bands. Therefore, we determined that the PLVX-TRADD-EGFP lentivirus was effectively expressed in HSFb and EFb cells (Figure 4).
Western blot detection of fusion protein expression after lentiviral transfection of EFb and HSFb. (1) Blank control group, (2) experimental control group, (3) experimental group. β-actin was used as an internal reference protein, and a prestained protein ladder was used. In the experimental group, a positive band was observed at 55–72 kDa, which was consistent with the predicted size of the TRADD-GFP-FLAG fusion protein (34 kDa + 28 kDa + 2 kDa = 64 kDa). (a) Western blot detection of fusion protein expression after PLVX-TRADD-EGFP lentivirus transfection of EFb. (b) Western blot detection of fusion protein expression after PLVX-TRADD-EGFP lentivirus transfection of HSFb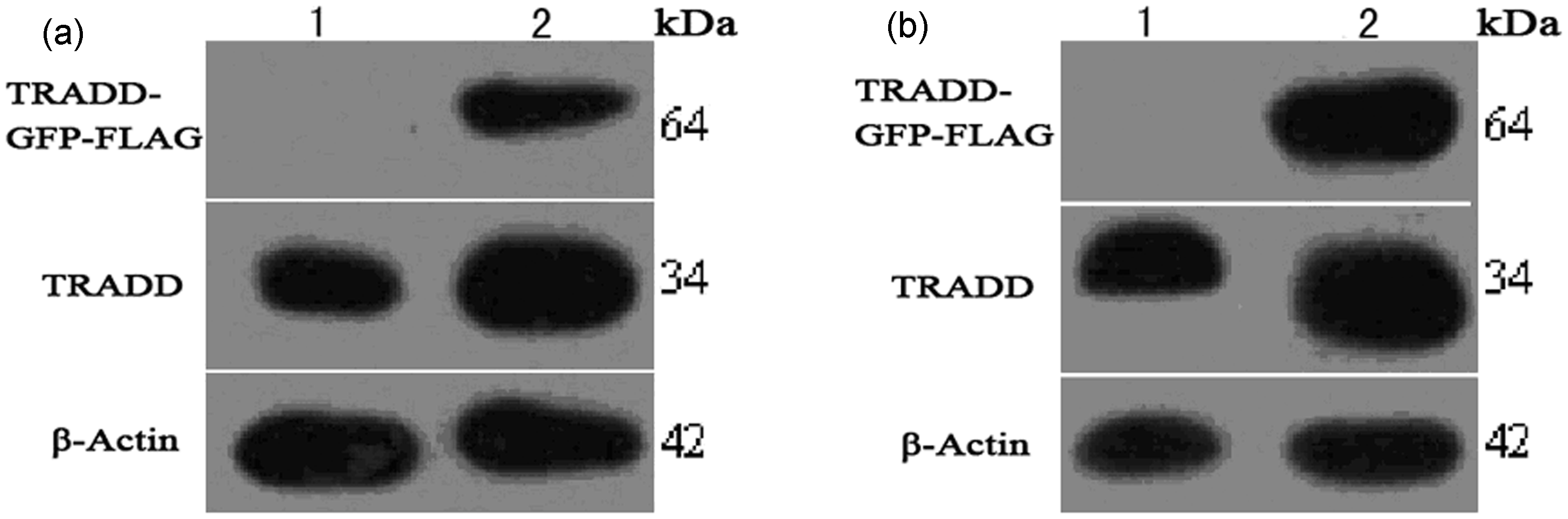
MTT assay detection of cell proliferation
The PLVX-TRADD-EGFP lentivirus and PLVX-EGFP lentivirus alone as well as those treated with 10 ng/mL TNF-α independently had positive effects on EFb proliferation (P < 0.05), whereas 10 ng/mL TNF-α treatment alone had no significant effect on EFb proliferation (Figure 5(a) and (c)). In contrast, the PLVX-TRADD-EGFP lentivirus effectively inhibited HSFb proliferation with or without 10 ng/mL TNF-α (P < 0.05), whereas TNF-α treatment alone and PLVX-EGFP lentivirus had no significant effect on HSFb proliferation (Figure 5(b) and (d)).
Effect of the PLVX-TRADD-EGFP lentivirus and TNF-α on proliferation of EFb and HSFb. Cells were counted and inoculated into five 96-well plates. One 96-well plate was analyzed at each of the following time points: 0 h, 24 h, 48 h, 72 h, and 96 h. OD values of the six groups of EFb and HSFb (blank control group, blank control group + TNF-α, experimental control group, experimental control group + TNF-α, experimental group, and experimental group + TNF-α) were measured on each plate at 490 nm. To generate a growth curve for each group, time was plotted on the horizontal axis, and the average OD value was plotted on the vertical axis. (a) and (c): effect of the PLVX-TRADD-EGFP lentivirus and TNF-α treatment on EFb proliferation. (b) and (d): effect of the PLVX-TRADD-EGFP lentivirus and TNF-α treatment on HSFb proliferation. aP < 0.05, Compared with the blank control group at the same time point. bP < 0.05, Compared with the experimental control group at the same time point. cP < 0.05, Compared with the experimental group at the same time point. dP < 0.05, Compared with the blank control + TNF-α group. eP < 0.05, Compared with the experimental control + TNF-α group at the same time point. (A color version of this figure is available in the online journal.)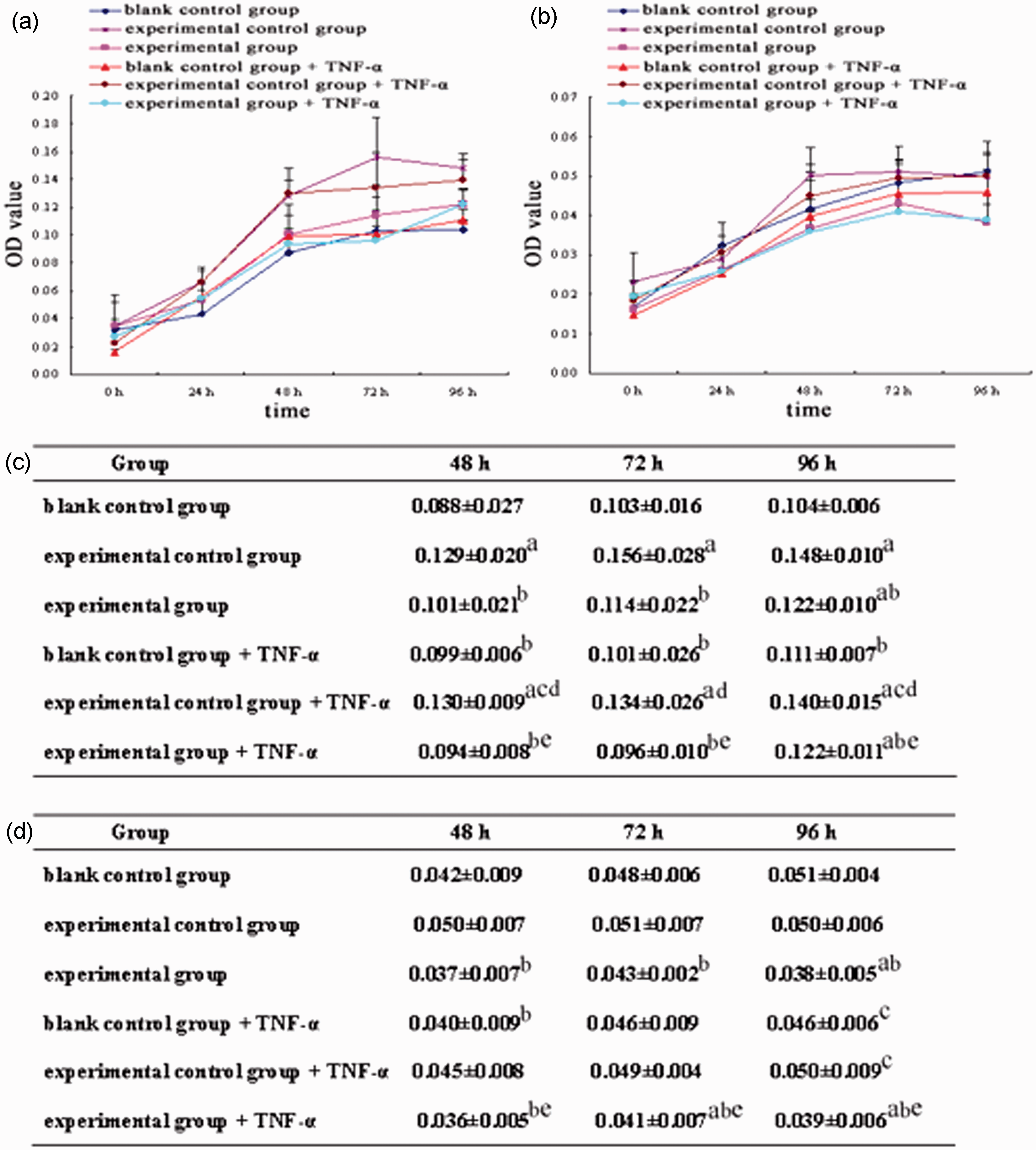
Flow cytometry evaluation of cell cycle and apoptosis
The PLVX-TRADD-EGFP lentivirus and PLVX-EGFP lentivirus alone, treatment with 10 ng/mL TNF-α alone, and the PLVX-TRADD-EGFP lentivirus and PLVX-EGFP lentivirus combined with 10 ng/mL TNF-α independently did not have a significant effect on EFb cell cycle or apoptosis (Figure 6(a)). The PLVX-TRADD-EGFP lentivirus caused HSFb to exhibit G2/M phase arrest, and treatment with 10 ng/mL TNF-α promoted this phase arrest. In addition, the PLVX-TRADD-EGFP lentivirus combined with 10 ng/mL TNF-α treatment induced the occurrence of a sub-G1 apoptosis peak in HSFb, suggesting that HSFb cell division was inhibited, and apoptosis had occurred. PLVX-EGFP lentivirus, both alone and treated with 10 ng/mL TNF-α, had no significant effect on HSFb (Figure 6(b)).
Effect of the PLVX-TRADD-EGFP lentivirus and TNF-α on cell cycle and apoptosis of EFb and HSFb. (a): effect of the PLVX-TRADD-EGFP lentivirus and TNF-α on EFb cell cycle and apoptosis, (b): effect of the PLVX-TRADD-EGFP lentivirus and TNF-α on HSFb cell cycle and apoptosis. (A color version of this figure is available in the online journal.)
ELISA detection of collagen synthesis
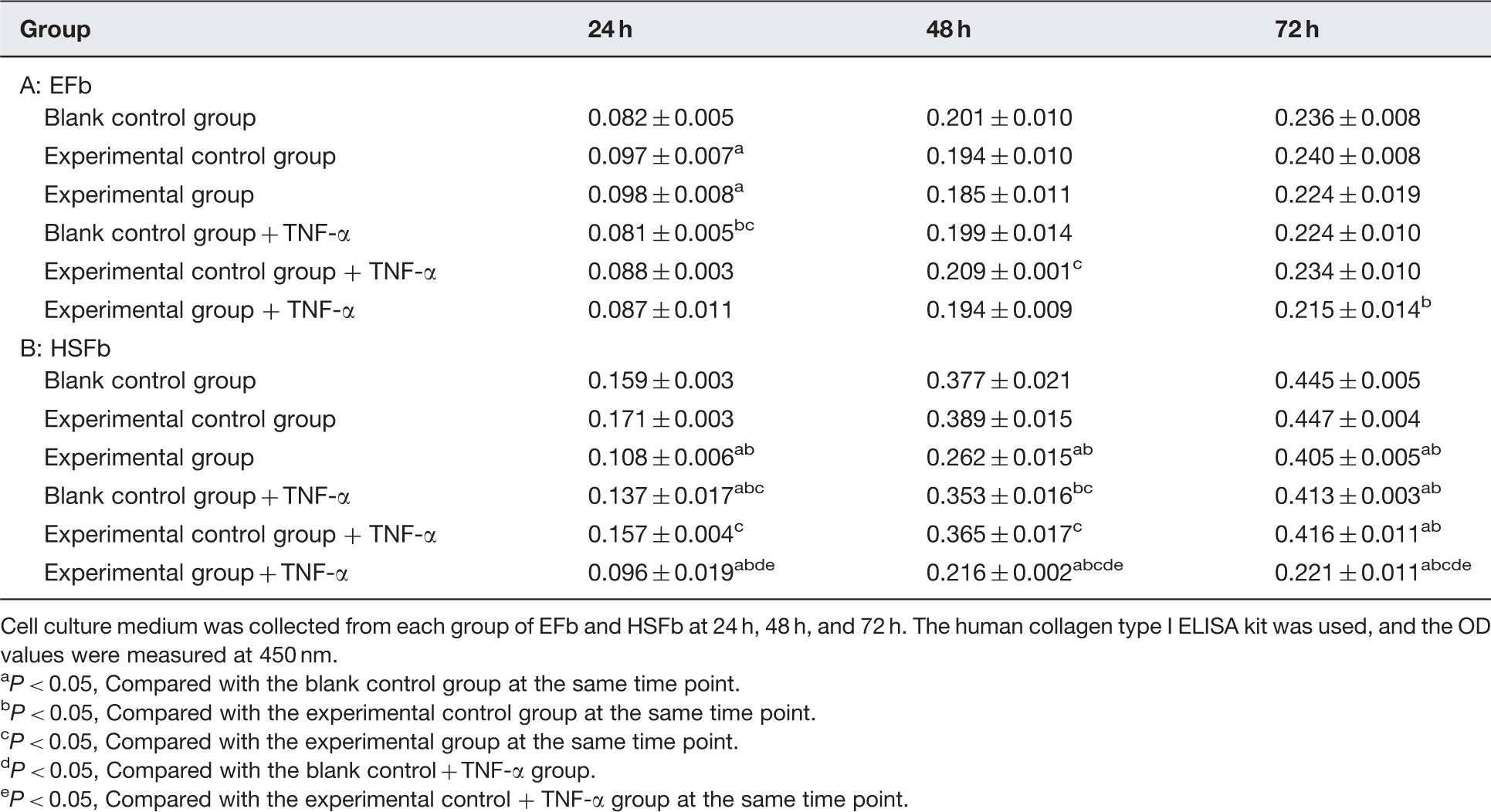
The EFb experimental control group and experimental group secreted increased levels of type I collagen at 24 h compared with the blank control and blank control + TNF-α groups (P < 0.05) (Table 1, A). The HSFb blank control group + TNF-α secreted less type I collagen at 24 h and 72 h compared with the blank control group (P < 0.05), and the experimental and experimental + TNF-α groups secreted less type I collagen at 24 h, 48 h, and 72 h (P < 0.05). The experimental group + TNF-α exhibited a significant reduction in type I collagen secretion (Table 1, B). Adjusted type I collagen expression levels of EFb and HSFb (
Cell culture medium was collected from each group of EFb and HSFb at 24 h, 48 h, and 72 h. The human collagen type I ELISA kit was used, and the OD values were measured at 450 nm. P < 0.05, Compared with the blank control group at the same time point. P < 0.05, Compared with the experimental control group at the same time point. P < 0.05, Compared with the experimental group at the same time point. P < 0.05, Compared with the blank control + TNF-α group. P < 0.05, Compared with the experimental control + TNF-α group at the same time point.
Statistical analysis
The SPSS13.0 data processing software was used for the statistical analysis of the data. The data were expressed as the means ± standard deviation (
Discussion
The binding of TRADD to TNF receptor 1 (TNFR1) is an important step in the TNF/TNFR1-mediated apoptosis signal transduction pathway. TRADD is an essential messenger molecule in the Toll-like receptor-mediated apoptosis signaling pathway, which is dependent on TNFR1 and Toll-IL-1-resistance domain-containing adaptor-inducing IFN-β (TRIF) in the fibroblast cell membrane. 17 Similar to another study, 18 the present work confirmed that the expression level of TRADD was significantly reduced in HSFb. Low levels of TRADD expression may result in an excessive proliferation of HSFb. Castagnoli et al. 19 and Peruccio et al. 20 used immunohistochemistry to demonstrate that the proportion of TNF-α-positive infiltrating cells in HS tissue was significantly decreased compared with normal scar tissue (8% in HS and 35.4% in normal scar tissue). We hypothesize that increased HSFb TRADD expression combined with TNF-α treatment may inhibit HSFb proliferation and reduce scar formation. Lentiviral vectors are ideal gene transfer vectors for in vivo gene therapy due to their highly efficient gene transfer, long-term stability of gene expression, and biological safety.21,22 Therefore, we chose a lentiviral vector to deliver the exogenous tradd gene for efficient integration into the genomes of HSFb and EFb, thus achieving long-term and effective expression of the TRADD fusion protein in these cells. After adding 10 ng/mL TNF-α, we performed an MTT assay and found that the PLVX-TRADD-EGFP lentivirus, both alone and in combination with 10 ng/mL TNF-α, inhibited HSFb, but not EFb, proliferation. Furthermore, 10 ng/mL TNF-α treatment alone had no significant effect on HSFb or EFb proliferation. Flow cytometry revealed that 10 ng/mL TNF-α promoted the HSFb G2/M phase arrest caused by the PLVX-TRADD-EGFP lentivirus, thus inhibiting cell division in HSFb. In addition, the PLVX-TRADD-EGFP lentivirus combined with 10 ng/mL TNF-α effectively promoted HSFb apoptosis. However, apoptosis and the cell cycle were not significantly affected in EFb. ELISA analysis revealed that the PLVX-TRADD-EGFP lentivirus, either alone or in combination with 10 ng/mL TNF-α, inhibited the secretion of type I collagen in HSFb, with the latter showing a significant effect. However, type I collagen secretion by EFb was not inhibited.
The above results show that TRADD is expressed at a low level in HSFb, thus leading to the suppression of TRADD-mediated cell apoptosis via the TNF-α/TNFR1 signaling pathway. This low TRADD expression is a principal reason for the excessive proliferation of HSFb. After transduction of the PLVX-TRADD-EGFP lentivirus, TRADD expression in HSFb recovered to normal levels. The overexpression lentivirus combined with 10 ng/mL TNF-α treatment increased HSFb apoptosis and inhibited HSFb proliferation and collagen secretion, thereby reducing HS formation. In contrast, this combination had no inhibitory effects on cell proliferation, apoptosis, or collagen secretion in EFb, with normal TRADD expression. Micheau and Tschopp 23 and Peter 24 found that the intracellular death domain (DD) of TNFR1, once bound to TNF-α, could interact with TRADD, producing two induction and regulatory mechanisms: (1) TRADD binds to the Fas-associated death domain and caspase-8 and then activates caspase-3, inducing apoptosis; (2) TRADD binds to TNF receptor-associated factors or recruits receptor-interacting protein to activate NF-κB, which inhibits apoptosis and promotes inflammation. Boldin et al. 25 demonstrated that these two TRADD-mediated signaling pathways of apoptosis and NF-κB activation are independent. In the present study, overexpression of the TRADD fusion protein selectively inhibited HSFb proliferation, and this process may be a mechanism to balance the two pathways, the details of which currently remain unclear. TNF-α has dual effects on fibroblast proliferation: at low concentrations, it stimulates fibroblast proliferation, while it inhibits fibroblast proliferation at high concentrations. Fibroblasts are heterogeneous; even under the same culture conditions, fibroblasts from different tissues may exhibit varying responses to TNF-α. This is due to differences in surface receptor number and density and in their capacity for collagen synthesis. In our study, 10 ng/mL TNF-α had no significant effect on the proliferation of HSFb or EFb but effectively inhibited collagen synthesis in HSFb. Therefore, on the basis of the selective inhibition of HSFb proliferation by the PLVX-TRADD-EGFP lentivirus, stimulation with 10 ng/mL TNF-α further inhibited the synthesis of type I collagen in HSFb, further preventing HS formation.
In this study, we investigated the effects of TNF-α and TRADD, a messenger molecule in the TNF-α/TNFR1-mediated apoptosis signaling pathway, on HSFb function, and we laid the experimental foundation for the clinical application of a PLVX-TRADD-EGFP lentivirus in antiHS treatment.
Footnotes
Author contributions
All authors participated in the study design, data interpretation and analysis, and review of the manuscript. GP: study concept and design, obtained funding. YY: acquisition of data, analysis and interpretation of data, drafting of the manuscript. KX: technical support. YL: acquisition of data. JD: statistical analysis. HW: material support.
ACKNOWLEDGEMENTS
This work was supported by the National Natural Science Foundation of China (81070384).