
Abstract
Dead Sea and magnesium salt therapy are two of the oldest forms of treatment for skin disease and several other disorders, supported by a body of largely anecdotal evidence. In this paper we review possible pathways for penetration of magnesium ions through the epidermis to reach the circulation, in turn replenishing cellular magnesium levels. We also discuss mechanisms for intercellular movement of magnesium ions and possible mechanisms for the interaction between magnesium ions and inflammatory mediators. Upon addition of magnesium ions in vitro, the expression of inflammatory mediators such as tumour necrosis factor α (TNFα) and nuclear factor κβ (NFκβ) is down regulated. Dysregulation of these and other inflammatory mediators has been linked to several inflammatory disorders, including asthma, arthritis, atherosclerosis and neuroinflammation.
Keywords
Introduction
Dead Sea therapy is one of the oldest forms of treatment for skin disease and some chronic inflammatory diseases like arthritis and psoriasis. 1 Much of the research to date has attributed the clinical effects of Dead Sea therapy to its mineral composition; mostly to magnesium salts.2,3 Magnesium salts, such as magnesium sulphate (Epsom salts), have long been used as a spa product and as a therapeutic to manage clinical conditions. 4
The central question addressed by this review is, “What are the underlying mechanisms by which magnesium ions could play a role in the regulation of inflammatory responses in the skin and systemically?” Several systematic studies have been conducted in humans, over the last two decades, in an effort to understand the effect of magnesium ions (Mg2+) in healing skin disorders. These will be reviewed. There has been renewed interest over the past decade in understanding the role of magnesium salts in clinical medicine, nutrition and physiology. This review will discuss the known clinical effects of magnesium deficiency, and both summarise and suggest molecular mechanisms that could mediate the inflammation induced by magnesium deficiency. We first discuss transdermal absorption as a possible route of administration for prevention and treatment of magnesium deficiency and for controlling inflammation.
Transdermal absorption of magnesium
Transport of Mg2+ across skin is a critical precondition for the function of topical, therapeutic compounds in treating skin and inflammatory diseases. Transdermal absorption is a potentially important route of transport for components that are involved in biological processes. 5 Even though much research has been carried out in the area of cutaneous permeation and transdermal absorption,5–7 mechanisms that lead to permeation of Mg2+ ions through the skin are not clearly understood and need further research.
Past studies on magnesium and other metal ion permeation through human skin demonstrated that it is not readily absorbed under normal physiological conditions, when the skin is intact and healthy.8–10 However, there is a considerable body of anecdotal and research data that attributes to magnesium a role in skin barrier and epidermal recovery after damage.3,11–13 In the case of compromised stratum corneum (SC), the viable epidermis and nerve endings (in atopic dermatitis [AD]) are exposed to incoming particles and chemicals.14,15 There is no effective barrier to restrict the movement of magnesium ions to epidermal cells or nerve endings, thus permitting a role for Mg2+ in skin recovery and modulation of the immune or nervous systems.3,16 The permeability of the skin is modified in pathological conditions, with both macroscopic and microscopic lesions17,18 that would allow penetration of magnesium below the SC, and subsequent transport or diffusion by mechanisms to be discussed below. Thus it is necessary to consider both the normal skin as well as barrier compromised or diseased skin.
In normal skin, the SC forms the outermost layer, formed by continuous replacement from the newly differentiated daughter cells of keratinocyte stem cells, displacing outwards. 19 It functions as a physical barrier hindering, but not completely preventing, transdermal penetration through its cellular structure. 5 The radius of the hydrated magnesium ion relative to the radius of the dehydrated ion is greater compared to other ions such as calcium, potassium and sodium.10,20 Irrespective of oral or transdermal administration, this greater radius could sterically and energetically hinder transport across cellular membranes. A review article by Lansdown 8 reports that magnesium in the form of hydrous polysilicate (talc) is not readily absorbed by normal skin, however, commonly used therapeutic formulations of magnesium utilize other salts such as chloride, or sulphates. The absorption kinetics such as solubility and permeation coefficients of chlorides and sulphates are different to those of polysilicates.4,8,21,22 Moreover, Table 7 in the same review article presents a positive score for percutaneous absorption of magnesium ions (although the temperature conditions are not specified). The same article cites the ability of Mg2+ to bind to hair. 23 This provides the possibility of magnesium permeation by shunt diffusion. Shunt diffusion is the mechanism by which diffusion occurs through hair follicles, pilosebaceous units and sweat glands, 5 although these constitute a small proportion of skin surface area, with the density dependent on the location of the skin. In the case of bulk diffusion, water soluble molecules are able to enter through 10 Å pores created by protein subunits in the lipid of SC.5,6,24 These could provide an entrance for hydrated magnesium, the radius of which is 4.76 Å.25,26 Subsequently, the transport of Mg2+ into cells could be facilitated by transmembrane proteins such as SLC41A1 and transient receptor potential melastatin 7 (TRPM7).27,28 SLC41A2, a cell surface transmembrane protein with its N-terminus outside and C-terminus inside the cell membrane, is responsible for magnesium transport across the plasma membrane. 27 Immunohistochemistry on epidermal cells has demonstrated a plasma membrane localization of murine SLC41A2. 27 The N-terminus of this protein, accessible to extracellular components, is involved in transcellular movement of Mg2+, which is in turn required for homeostasis, cell growth and neuronal function. 27 Similarly, the human SLC41A1 functions as a Mg2+ transporter involved in magnesium homeostasis in epithelial cells. 27
Another important mechanism by which Mg2+ intracellular homeostasis in humans is facilitated, is via the protein TRPM7. 28 Knockout of TRPM7 in DT40 B cells (derived from an avian leucosis virus induced bursal lymphoma in a white leghorn chicken) resulted in lowered intracellular Mg2+ and inhibition of cellular proliferation. 29 Under stress (apoptotic stimuli), the TRPM7-knockdown fibroblast cell line (3T3-M7shRNA6) was more resistant to apoptosis and had a lower intracellular concentration of reactive oxygen species (ROS) compared to control cells. This suggests a role for Mg2+, mediated by the magnesium transporter TRPM7, in cell survival and regulation of cellular ROS concentration. 29
Another factor influencing percutaneous absorption of magnesium ions through skin, is the negative charge carried on the surface of tissues. 30 Accordingly, it is likely that the positively charged magnesium ions can be absorbed on the negatively charged SC, enhancing the retention time and bioavailability on the skin surface. 31 This coupled with bulk diffusion and the factors mentioned above could enhance magnesium ion penetration through normal human skin.
In normal human skin, factors such as temperature and humidity, osmolarity, dehydration and penetration enhancers, could lead to enhanced percutaneous absorption of magnesium. In certain therapies, increased temperature conditions can also provide improved skin permeability enabling penetration of mineral salts.5,32,33 A study conducted to measure the effect of heat on skin permeability showed a strong dependence of permeability on temperature. Short pulses of high temperature resulted in increases in calcein permeability in human cadaver skin. 34 In relation to Dead Sea therapy on normal human skin, the high salt concentration coupled with the hydrated state of the skin could together cause an osmotic effect, 35 leading to increased flux of ions through the skin due to a concentration gradient across the skin. However, in commercially available topical magnesium formulations it is likely that penetration enhancers would be necessary in order to enhance passage through the SC layer in normal skin. The role of these enhancers is to penetrate into the skin, reversibly decrease the barrier resistance of the SC and to create a water equilibrium between SC and viable epidermis. 36
Control of inflammation by magnesium: Possible mechanisms
It is well established that Mg2+ deficiency has a direct influence on inflammation.
37
However, the molecular mechanisms by which Mg2+ suppresses inflammation are unclear. A possible link could be activation by Mg2+ of the thiamine pyrophosphate (TPP)-dependent riboswitch, resulting in increased synthesis of thiazole from thiazole pyrophosphate
38
(see Figure. 1). The TPP-dependant riboswitch is the only known functional riboswitch mechanism in eukaryotes and it is known that TPP binding to thiA riboswitch is increased by Mg2+.38,39 The adenine rich sites on the riboswitch bind Mg2+ leading to structural changes favouring TPP binding.
38
The TPP-dependant thiA riboswitch up-regulates thiazole synthase, an enzyme required for catalysing the conversion of thiazole pyrophosphate to thiazole.40,41 Thiazole derivatives have a spectrum of anti-inflammatory and neuroprotective activities.42–46
Possible influence of Mg2+ in thiazole synthesis in turn resulting in anti-inflammatory properties. Thiamine pyrophosphate (TPP), an activated form of thiamine binds to thiA riboswitch in the presence of magnesium ions (Mg2+). An adenine rich site in the riboswitch binds Mg2+ increasing the affinity of TPP binding.
13
The riboswitch undergoes structural change then expresses thiazole synthase,
13
resulting in the formation of TPP intermediate and in turn forming thiazole derivatives. Thiazole derivatives are known to inhibit cyclooxygenase, hindering the formation of prostanoids, in turn producing anti-inflammatory effects.17–19 The thiA riboswitch is the only known eukaryotic riboswitch.
14
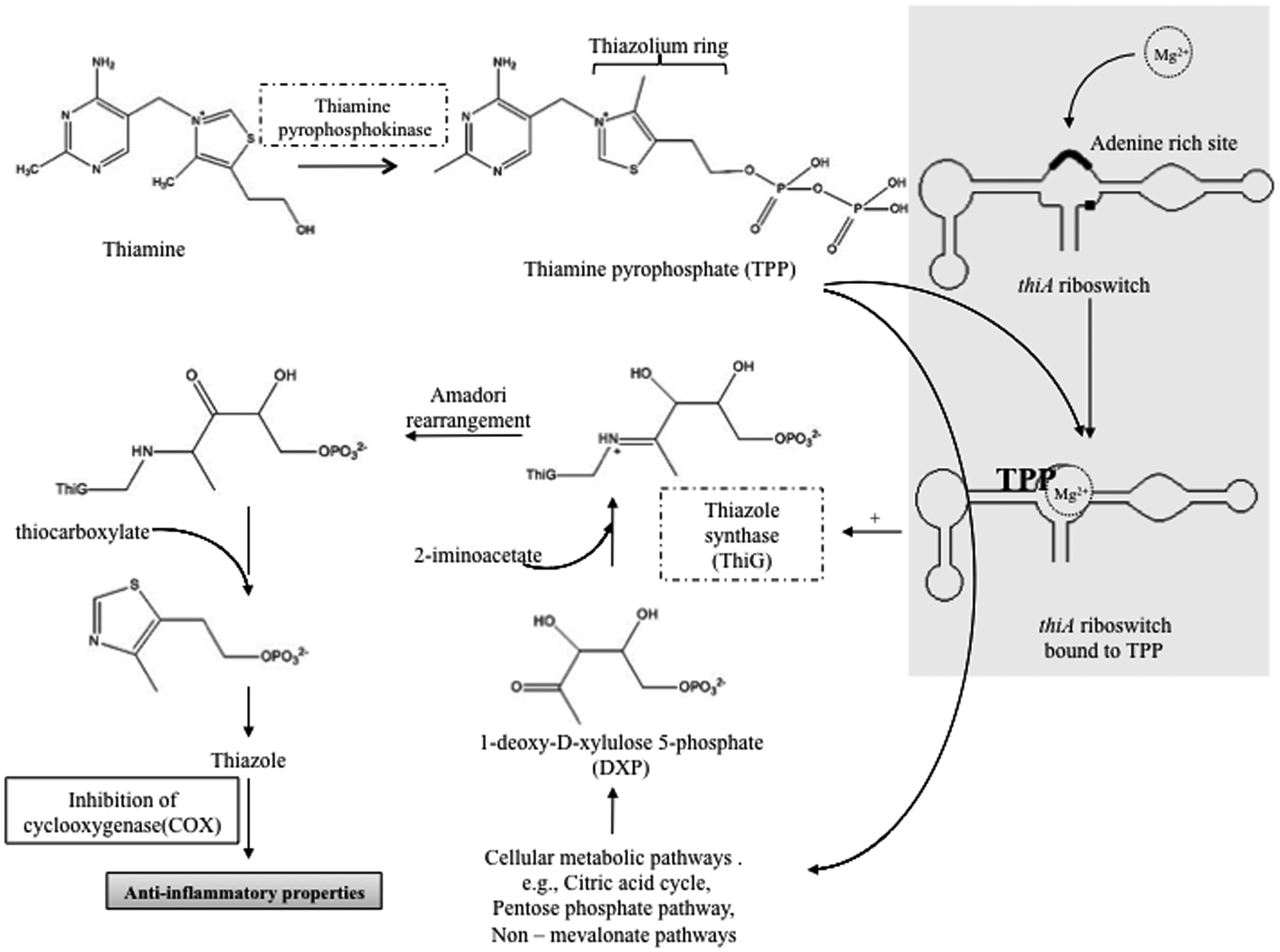
Molecular mechanisms and inflammatory pathways
Illnesses related to inflammation following a state of chronic or acute hypomagnesaemia are well documented, both in humans and experimental rat models.37,47–49 However, there is a paucity of information in the literature regarding the cascade of molecular events culminating in inflammation during hypomagnesaemia.
Here we review known molecular mechanisms and propose additional ones that could be responsible for inflammation resulting from Mg2+ deficiency, additionally suggesting how this might relate to inflammation localized in the skin. Firstly an examination of how the hypomagnesaemic state results in tumour necrosis factor α (TNFα) and nuclear factor κβ (NFκβ) activation will be considered, in view of the importance as inducers of transcription of pro-inflammatory genes. These changes pave the way for pro-inflammatory cytokine responses, followed by alteration of macrophage and neutrophil activity, including their participation in a pro-inflammatory positive feedback loop (see Figure 2).
General summarized pathway of inflammation following hypomagnesaemia. Evidence suggests that hypomagnesaemic conditions can exacerbate trauma and hypoxia (leading to increased oxidative damage to cells and tissues from free radicals,112,113 and stimulates the production of interleukin 1 (IL-1), IL-17, interferon γ (IFN-γ) and granulocyte macrophage colony-stimulating factor (GM-CSF). The presence of these soluble proteins allows for the stimulation of various immune cells to produce tumour necrosis factor α (TNFα) and its reciprocal receptor (for cell surface presentation) following the nuclear translocation of nuclear factor κβ (NFκβ) acting as a transcription factor. The secretion of TNFα from cells (after TNFα converting enzyme (TACE) converts tm TNFα to its soluble form) allows for autocrine and paracrine effects that upon TNFα binding facilitates amplification of inflammatory responses such as transcription, translation and secretion of more TNFα, IL-1, IL6 and IFN-γ. A positive feedback loop is established and the latter two cytokines cause inflammation. Normally prostaglandins, IL-10 and corticosteroids have an inhibitory effect on TNFα transcription; however this is promoted by Mg2+ and may therefore be disrupted in hypomagnesaemic conditions.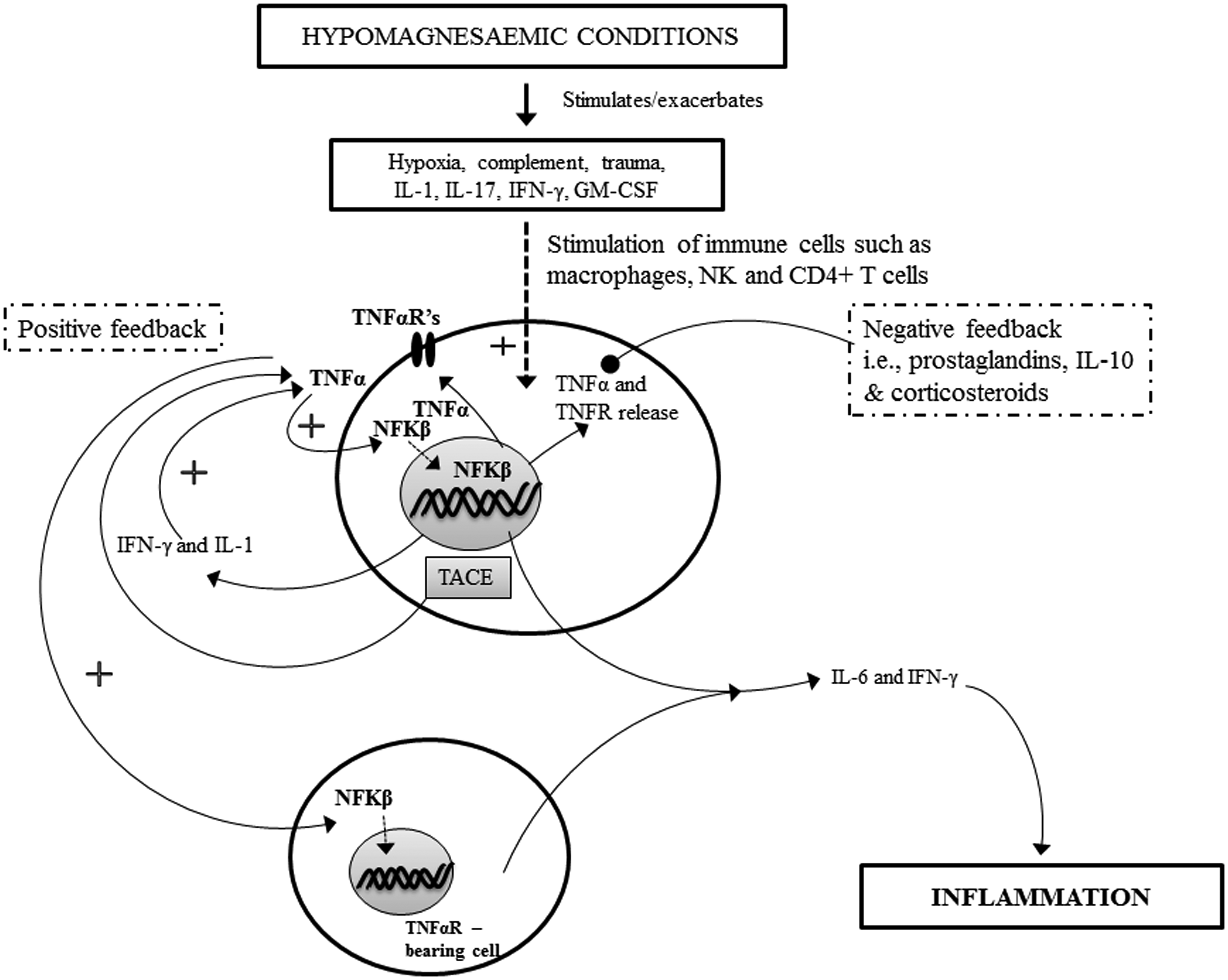
Magnesium in TNFα and NFκβ signalling
It has been demonstrated that hypomagnesaemia in rodents 37 and in people with metabolic syndrome 47 result in elevated serum concentrations of TNFα compared to healthy counterparts (in the human studies), or controls (in rodent studies). An elevation in TNFα and magnesium deficiency was also observed in obese human subjects when compared to healthy and moderately overweight individuals. 50 A recent study revealed that TNFα levels declined following in-vivo administration of MgSO4 to human subjects. 51 Additional work showed that the magnesium ion component, not the sulphate, was responsible for the immunomodulatory effect. 51 Other research has shown that magnesium deficiency in mice with a knockout of the gene encoding the TNFα receptor (TNFαR) caused less adverse effects on bone loss than in the wild-type controls fed the same diet. 48 These results suggest an inverse relationship between magnesium intake, TNFα concentration and TNF actions mediated by the TNFαR. The physiological significance of these observations becomes evident in the context of the known pro-inflammatory actions of TNFα. 52 TNFα is a regulatory cytokine produced by various cell types including macrophages, T-helper cells (CD4+ TH cells) and natural killer (NK) cells. 53 The predominant role of this cytokine is the systemic regulation of immune cells with beneficial outcomes such as augmented recruitment of defence mechanisms during infection, including fever induction. 52 However, in common with all endogenous immune mediators, balance is essential and a prolonged high serum concentration of TNFα results in prolonged inflammation and effective damage locally and systemically. 52 Systemic diseases such as systemic lupus erythematosis and local organ diseases including psoriasis and osteoarthritis are all associated with dysregulation (increased concentrations) of this cytokine.49,54,55
The biosynthesis of TNFα is increased by an array of stimuli including hypoxia, trauma, complement components 56 and various cytokines including interleukin 1 (IL-1), IL-17 interferon-γ (IFN-γ) and granulocyte macrophage colony-stimulating factor (GM-CSF). 52 It has been shown in vitro, using various cell culture models (including human and rat cell lines) that under hypomagnesaemic conditions in the culture medium, the concentrations of the aforementioned cytokines show an increase compared to concentrations in cell lines maintained under normomagnesaemic conditions.57–60 The increased concentration of IL-1 and IL-17 61 can result in establishment of positive feedback loops (which can occur locally or systemically depending on the disease in question; see reviews referenced62,63 for more details beyond the scope of this review), with TNFα facilitating the generation of IL-1 and IFN-γ, in turn driving further TNFα effects.52,64–66 Transcription of TNFα is governed by NFκβ, the inflammation-related transcriptional factor, 67 discussed in more detail below.
Negative feedback loops also operate, these are known to regulate TNFα levels by inhibition of transcription of TNFα mRNA. 52 This negative feedback arises when TNFα stimulates the production of molecules that inhibits TNFα transcription including prostaglandins, IL-10 and corticosteroids. Mg2+ is also known to promote prostaglandin synthesis, and so it can be speculated that in the hypomagnesaemic individual, the negative feedback loop is disrupted, with less feedback control over elevated TNF-α levels.52,64,68–70 It remains to be determined what levels of magnesium deficiency tip the balance towards uncontrolled positive feedback in humans, resulting in clinically observable effects. Nor is it known how prolonged such effects would be. There is a paucity of data regarding the effects of Mg2+ on IL-10 or corticosteroid levels, hence the interaction between Mg2+ and the endocrine system is ripe for investigation.
After translation, TNFα exists in a cell-surface bound precursor form, termed transmembrane TNFα (tmTNFα). This intermediate is converted to a soluble cytokine via TNFα-converting enzyme.71,72 Both tmTNFα and sTNFα are biologically active and capable of binding to their receptors, TNFR1 and TNFR2, triggering different downstream signalling events, which result in outcomes such as apoptosis, necroptosis, transcriptional factor AP-1 activation, or NFκβ activation and translocation to the nucleus. 73 Once binding has occurred between TNFα and its receptor (either TNFR1 or 2), a conformational change occurs in the latter, followed by the interaction between the intracellular domains of TNFR and other proteins including TRAF2, cIAP1 and cIAP2, forming a complex. This complex formation leads to the activation of the IКB kinase (IKK) complex. The IKB complex consists of two kinase subunits, which phosphorylate the NFκβ inhibitor protein IКBα, tagging it for the ubiquitin-proteosome pathway of degradation. This allows NFκβ to freely translocate to the nucleus where it coordinates the transcription of genes such as cIAP1/2 and TRAF2, which are important in the regulation of NFκβ and apoptotic pathways.74,75
Interplay and importance of Ca2+ and Mg2+ signalling
NFκβ activity is regulated by various secondary messengers including intracellular calcium ions
The critical role of Ca2+ in the NFκβ pathway is supported by the observation that Ca2+ chelators prevent the induction of NFκβ activity in vivo in murine models. 79 Other research has shown that when Mg2+ deficient rats were fed a Ca2+ deficient diet, the inflammatory effect was greatly reduced (as measured by reduced inflammation scores, prevention of leucocytosis and reduced splenomegaly) when compared to other hypomagnesaemic rats. 80 Once the active NFκβ crosses the nucleus it up-regulates the transcription of TNFα.81,82 It should be mentioned, however, that the evidence for NFκβ-mediated TNFα expression is mostly limited to murine models, and studies relating to humans are limited.83,84 However, in 2010 one study utilized mouse bone marrow-derived dendritic cells to demonstrate NFκβ-mediated positive expression of the TNFα gene. 81 Additionally it would be of interest to study whether a high Ca2+, low Mg2+ state up-regulates NFκβ activity and TNFα expression permitting TNFα synergism with STAT6 to switch B cells to IgE production. 85 This is important to establish the role of IgE in atopic and inflammatory conditions such as AD. 85
The antagonism between Mg2+ and Ca2+, and competition for binding sites on receptors, enables Mg2+ to overcome the toxic effects produced by excessive Ca2+ concentrations in cells of the immune system that are located in the brain (human microglial cells in tissue culture). 86 In the case of neuroinflammation, in vitro experimentation has shown that an influx of Ca2+ into microglia (brain resident macrophages) and THP-1 cells activates their associated purinergic receptors and subsequently inflammation. 87 Mg2+ is effective in ameliorating the neurotoxic effect produced by over-activation of human microglial cells that occurs as a result of elevated levels of inflammatory cytokines in the cells such as TNF-α, IL-6 and nitrite ions.86,88 These agents are released as a result of intracellular inflammatory pathway activation, via P38 MAPK and NFκβ. 86
Pro-inflammatory cytokine responses and resident microflora
In addition to stimulating TNFα transcription, NFκβ activates the transcription of IL-6 and IFN-γ after binding to the cognate promoter regions.81,89,90 These potent pro-inflammatory cytokines have been shown to be present in high levels during periods of Mg2+deficiency.91–93 Additionally, in one study of septic shock using ex vivo human whole-blood, it was shown that following addition of Mg2+, the baseline level of TNFα and IL-6 production fell. 94 This observation provides further evidence of the inverse relationship between these inflammatory mediators and magnesium concentration. 94 However, the concentration of Mg2+ was well above physiologically relevant levels. 94 More research in other conditions, using a similar experimental set up is needed to further elucidate links between Mg2+ levels in human blood and pro-inflammatory cytokine production. Further, a more recent study, using THP-1 cells isolated from human neonatal cord blood, found that treatment with Mg2+ (at levels known to be clinically effective in vivo) reduced the production of IL-1β, TNF-α and IL-8 cytokines and IL-6 in cord blood monocytes. 95
IL-6 exerts effects on a variety of cells including T and B lymphocytes, hepatocytes, hematopoietic progenitor cells and fibroblasts, with consequent systemic effects such as acute phase reactant protein production, immunoglobulin synthesis and naive CD4+ T cell differentiation into Th17 cells. IL-17 secreted from Th17 cells is responsible for autoimmune tissue injury. 96 In relation to skin pathology, it has been shown that the epidermis from psoriatic skin produces high levels of IL-6 in addition to over-expression of TNFR, due in turn to the higher levels of IFN-γ. 97 Many of the characteristic phenotypic features of keratinocytes from psoriatic skin, including growth activation and ICAM-1 up-regulation, are a result of the actions of IL-6, TNFα and IFN-γ.97,98 The histological features of psoriasis such as the presence and accumulation of inflammatory cells (including polymorphonuclear leukocytes) and epidermal hyperplasia have been attributed to elevated TNFα activity.99,100 Given the aforementioned studies 97 indicating a role for magnesium in modulating the production of pro-inflammatory cytokines such as IL-6 and TNFα, there may be a role for magnesium-containing compounds in the treatment of psoriasis. However, whether such treatment would be effective in either acute or chronic cases, differing levels of severity, or what the most effective dose and route of administration would be (i.e. topical or oral) remain to be shown.
Another interesting finding is that mice fed on a Mg2+ deficient diet for four days showed higher levels of IL-6 and TNFα mRNA in the liver and intestine, a drop in the levels of the mRNA of zonula occludens-1, occludin and proglucagon in the ileum (three factors controlling gut barrier integrity and function) and possessed reduced gut bifidobacteria levels when compared to controls. 101 Bifidobacterium strains have been shown to repress inflammation in a variety of situations including studies of ulcerative colitis and skin inflammation (i.e. acne).102–104 With respect to skin inflammation, Bifidobacterium strains appear to facilitate reduction in substance P, a molecule that increases TNFα expression. One study in human female volunteers demonstrated a reduction in sensitive skin and heightened resistance to physical and chemical insults to the skin (in contrast to a negative control cream) following topical application of B. longum spp. products. Additionally, in the same paper, the authors report a statistically significant reduction in markers of inflammation (including oedema, mast cell degranulation and TNFα release) following the application of a B. longum preparation on ex-vivo human skin explants. 104
However, in the above mentioned 101 mouse study, the animals on a Mg2+-deficient diet for 21 days demonstrated a potentially compensatory increase in caecal Bifidobacteria levels, restoration of intestinal barrier function and a waning of inflammation when compared to control mice. This is in contrast to mice on the same diet for four days only; in the latter case decreased levels of Bifidobacteria, and an increase in IL-6, TNFα and other markers of pathology were present. Drawing generalizable conclusions from this study is difficult given that it was a mouse-model study (not human) and measurements of bacteria and relevant mRNA levels were not continued after 21 days. If there was adaptation, were there any other long-term adverse effects on physiology? In the future it will also be important to work out the complex relationship between the gut bacteria and cytokine responses; is it bidirectional or unidirectional? What feedback loops exist? Moreover, the regulation is unlikely to be the same for all cytokines. These are all questions that need to be investigated.
Changes in macrophages and neutrophils
In rodent studies, a state of hypomagnesaemia has been associated with the activation of macrophages and neutrophils.105,106 It has been suggested that this is due to the increase in circulating pro-inflammatory mediators, including substance P, IL-6, TNFα and IFN-γ; however, given that macrophages and neutrophils are amongst the cells that produce these cytokines, it remains difficult to say whether the aforementioned cells are activated by these substances or if they are activated directly by the low levels of Mg2+.
107
Malpuech-Brugère and colleagues suggest that it is due to the latter, a drop in circulating Mg2+ concentration, which would lead to a significant increase in Ca2+ levels,37,108 in turn stimulating cellular proliferation. Whilst macrophages are also able to be activated independently of
Aside from enhanced protein expression of IL-6 and TNFα, activated macrophages also demonstrate increased expression of IL-1 proteins in Mg2+-deficient rats when compared to a control group. In the same study it was postulated that this contributed to cardiac lesions (IL-1 promotes expression of endothelin from heart endothelial cells which causes vasospasm). 106 Additionally, IL-1 is antagonistic to endothelial proliferation, suggesting another mechanism behind the lesions seen. 109 Furthermore, in the skin, IL-1 is implicated in wound repair and skin pathology, with IL-1 receptor (IL-1R) knockout mice demonstrating reduced cutaneous and deep tissue fibrosis and scarring and restoration of skin architecture. 110
Neutrophils are capable of releasing superoxide anions via their NADPH oxidase system and can contribute to tissue damage during Mg2+ deficiency, as demonstrated in a study which also demonstrated phagocytic activity of neutrophils in rats on a magnesium deficient diet. 105 The study also showed that the free radical production from neutrophils is inhibited when high Mg2+ levels are present in the extracellular space. 105 Mg2+ was also found to inhibit superoxide in cultured human neutrophils. 110 The latter experiment also revealed synergistic inhibition of superoxides with the addition of zinc ions to the magnesium solutions. 110 Neutrophil migration to the skin, a process that promotes keratinocyte apoptosis is TNFα dependant. These phenomena are key features of hyper-proliferative skin diseases such as psoriasis. 111
Clinical implications of magnesium deficiency
Magnesium is a micronutrient required for normal growth and development. Numerous clinical disorders have been associated with magnesium deficiency. Inflammation is a primary reaction brought about by magnesium deficiency, creating oxidative stress and subsequent immune stress. The clinical disorders could be a consequence of this stress response.91,112,113 Magnesium ions bind to macromolecules and cell membranes. Mg2+ is known to affect cellular functions, including the transport of potassium and Ca2+, modulation of signal transduction, cell proliferation and energy metabolism.
114
Early stages of Mg2+ deficiency can be characterized by a wide range of symptoms such as anorexia, vomiting, weakness, paraesthesia, muscular cramps, irritability and impaired cognitive functioning reflected by a decreased attention span. Mg2+ deficiency is related to poor dietary Mg2+ intake, often as a result of lifestyle changes, leading to the aforementioned health disorders.114–116 Mg2+ deficiency triggers inflammatory responses, including abnormal calcium homeostasis, activation of N-methyl-
The pathogenesis of asthma, a chronic inflammatory disorder involves activation of NFκβ and, expression of pro-inflammatory cytokines, chemokines and inflammatory mediators (IFN-
A characteristic property of Mg2+ is its antagonism of Ca2+ (see prior section ‘Interplay and importance of Ca2+ and Mg2+ signalling’). It competes with Ca2+ for entry into cells through voltage-gated channels and receptors and inhibits
The synovial fluid from patients with rheumatoid arthritis contains elevated levels of TNFα (an activator of NFκβ), which is important in the pathogenesis.118,121 Mg2+ deficiency can lead to lipid peroxidation and membrane oxidation, which in turn activates the NFκβ pathway.91,122 Activation of inflammatory responses due to Mg2+ deficiency causes chronic inflammation leading to different types of arthritis, depending on the site of NFκβ activation.
Studies in humans indicate that low Mg2+ intake and blood plasma concentration are linked with enhanced risk of atherosclerotic disease. 123 Atherosclerosis is currently classified as an inflammatory disease, having interactions between modified lipoproteins, macrophages, T lymphocytes and the components of arterial walls, 91 leading to the development of atherosclerotic lesions. Experimental results suggest regression of such lesions and suppression of atherogenesis in low-density lipid receptor deficient mice fed with Mg2+supplement.124,125 Infusion of Mg2+ at supra-physiological concentrations causes vasodilation of coronary arteries and systemic vasculature, antiarrhythmic effects and platelet inhibition. 126 Studies also show dietary administration of Mg2+ attenuates atherosclerotic lesions by lowering serum cholesterols and triglycerides in cholesterol fed animals. 91 On the basis of these studies, it appears that Mg2+ concentration regulates lipid metabolism and reduces atherosclerosis in animal models. 91
Even though the immune system and, in particular, the inflammatory response operates systemically, the inter-relationship between inflammation occurring in the nervous system and systemic inflammation needs to be better understood. It is widely accepted that several neurological disorders are characterised by an inflammatory component.127,128 There are several drugs undergoing test that are posited to act by reducing neurodegeneration, at least in part through inhibition of the inflammatory response of glial cells. 129 However, these drugs exert their effect throughout the body, resulting in global immunosuppression. 130 It would be ideal for such drugs to specifically target the glial cells and control inflammation in the brain without producing systemic immunosuppression.129,131 Studies conducted in mice using the compound 4,6-diphenyl-3-(4-(pyrimidin-2-yl)piperazin-1-yl) pyridazine (MW01-5-188WH), aimed at selective suppression of neuroinflammation, has yielded some positive outcomes without producing extra-neural inflammation. 129
Mg2+ administration could potentially be an effective treatment of neurodegenerative diseases via its antagonism of Ca2+ channels. This selectively suppresses neuroinflammation.86,91 If treatment for neurodegenerative diseases involved administration of Mg2+ locally to the brain, it could conceivably avoid the generalised stress on the immune system that is caused by non-targeted anti-inflammatory drugs. Experiments conducted on rat ischaemic and excitotoxic brain injury models shows the activity of Mg2+ as a neuroprotective agent.114,132,133 This is achieved by Mg2+ blockade of NMDA receptors and enhancement of regional cerebral blood flow to ischaemic areas of the brain. Mg2+ also inhibits entry of Ca2+ into cells through voltage-operated and receptor-operated channels.114,126
The nervous and immune systems interact bi-directionally. Mg2+ deficiency is known to induce a systemic stress response by activating neuroendocrine pathways, modifying production and activity of neuromediators such as acetylcholine, catecholamines and substance P. These have well established roles in the progression of both local and systemic inflammatory responses. 134 Administration of Mg2+ has been shown to block Ca2+ traffic through cell surface channels, acting as a broad inhibitor of neuroinflammation.86,135 Elevated systemic levels of Mg2+ have been shown to reduce damaging consequences of Ca2+ induced neuroinflammation in Parkinson's disease and Alzheimer's disease. 86
AD is a skin disease that is a result of interactions between skin, nervous system and immune cells. Nerve growth factor (NGF) is a neurotrophin, mainly produced in the basal keratinocytes and are present in elevated levels in plasma of AD patients. 136 In normal skin, less expression of NGF was found.137,138 These NGFs are known to be involved in the extension of C-fibres and promote increased density of nerve endings. In AD this results in exposure of a high density of unprotected nerves to external conditions, leading to the itch and scratch cycle that characterizes the disease.14,139 Semaphorin3A (Sema3A) is another factor involved in the development of AD. It is an axon guidance molecule that inhibits outgrowth of sensory neurons. It does so by binding to plexin–A1-4 and its co-receptor neuropilin-1 (NRP-1), subduing the effects caused by NGFs.140,141 Sema3A acts by suppressing nerve extension, and inhibition of histamine release from mast cells (existing treatments for AD rely on blocking the histamine receptors (H1-R) with anti-histamines or topical steroids). Further, Sem3A also binds to NRP-1. 142 NRP-1 is known to activate the NFκβ pathway and to initiate keratinocyte proliferation. 143
One study has shown that increased calcium ion concentration (0.45–0.75 mmol/L) in normal human epidermal keratinocytes augments the expression of Sema3A. 144 While calcium and magnesium have antagonist effects at the cellular level, it is possible that the skin barrier recovery after treatment with Dead Sea minerals (comprising calcium and magnesium salts) is due to the combined role of calcium in upregulating Sema3A, and the prevention by m magnesium of mast cell degranulation through other mechanisms (potentially via effects on TNFα or STAT-6145,146). However, a direct role for magnesium ions in regulation of Sema3A and NGFs remains to be established, and is an important area for further investigation, given the role of these factors in AD.
In relation to skin disease, a clinical study was conducted on 30 AD candidates, in which subjects were tested over six weeks for transepidermal water loss (TEWL), skin hydration, skin redness and skin roughness. 3 Upon treating one of their arms with a 5% Dead Sea salt solution at 38–42℃ and the other arm with tap water (38–42℃) as control, an improvement in TEWL, with reduction in AD symptoms in the Dead Sea salt treated group was shown. 3 Further work is needed, to confirm these studies and to measure the intracellular and molecular correlates of the structural changes in the skin.
Conclusion
Absorption of Mg2+ ions across the normal SC could occur under conditions of elevated temperature or changed hydration conditions (for example high salt concentrations). Absorption of magnesium will take place in cases of skin pathology or injury, where there is physical disruption of the SC. Subsequently, transmembrane proteins, such as SLC41A2 could assist intercellular transport of magnesium ions, leading to further penetration through the organ systems. 27 The action of magnesium ion as an anti-inflammatory agent could be via several pathways, such as activation of the TPP-dependant riboswitch. 38 Magnesium deficiency results in activation of TNFα and NFκβ, which can further facilitate pro-inflammatory cytokines.76,106 It would also be of interest to study the effect of Mg2+ on the synergism between TNF-α and STAT6, a mediator of IgE receptor mediated mast cell responses in late phase allergic responses and AD.145,146 Experimental data from humans and mice suggest an inverse relationship between magnesium intake and TNFα concentration, in addition to several other markers of inflammation.147–149 The hypomagnesaemic condition increases the influx of calcium into cells, resulting in elevated NFκβ activity.150,151 Evidence for activation of neutrophils and macrophages by calcium ions in mice has also been found. 150 Collectively, the inflammatory responses triggered by magnesium deficiency can result in clinical disorders. The interaction between Mg2+ and inflammatory mediators is ripe for investigation. For example, there is a paucity of data relating to the effects of Mg2+ on IL-10 or corticosteroid concentrations. More research is needed to further elucidate links between Mg2+ levels in human blood and pro-inflammatory cytokines. The effect of treatment with magnesium containing compounds in acute or chronic diseases with differing levels of severity, and the most effective doses and routes of administration in these cases, remain to be systematically determined. Although a role of magnesium deficiency in neurodegenerative disease is established, and may be mediated by interaction of magnesium ions with glial cells, 134 the interrelationship between inflammation in the nervous system and systemic inflammation needs to be better understood. Thus, there exist plausible mechanisms by which several metabolic and inflammatory conditions might potentially be alleviated through magnesium administration, either systemically or locally. These mechanisms are ripe for further investigation.
Footnotes
Author contributions
All authors participated in the writing, review and editing of this manuscript. NCC and CW contributed equally to the writing of this manuscript.
ACKNOWLEDGMENTS
This work was funded by grants from the University of Queensland, The National Health and Medical Research council of Australia, and Cancer Council Queensland.