
Abstract
The present study was aimed to investigate whether a decrease of nitric oxide (NO) level is beneficial for sponateous resumptiom of meiosis in diplotene-arrested oocytes cultured in vitro. For this purpose, diplotene-arrested oocytes were collected from ovary of immature female rats after a single subcutaneous injection of 20 IU pregnant mare’s serum gonadotropins (PMSG) for 48 h. In vitro effects of S-nitroso-l-acetyl penicillamine (SNAP; an NO donor) and aminoguanidine (AG; an inducible NOS [iNOS] inhibitor), intracellular NO, cyclic guanosine monophosphate (cGMP), Cdc25B, Thr-14/Tyr-15 and Thr-161 phosphorylated cyclin-dependent kinase-1 (CDK1), and cyclin B1 levels were analyzed. The SNAP inhibited spontaneous meiotic resumption form diplotene arrest in a concentration-dependent manner, while AG-induced meiotic resumption form diplotene in 0.1 mmol/L 3-isobutyl-1-methylxanthine (IBMX)-treated oocytes in a concentration-dependent manner. The intracellular NO as well as cGMP levels were decreased significantly during spontaneous meiotic resumption from diplotene arrest. The reduction of Cdc25B expression level was associated with the accumulation of Thr-14/Tyr-15 phosphorylated CDK1 level. However, Thr-161 phosphorylated CDK1 as well as cyclin B1 levels were reduced significantly during meiotic resumption from diplotene arrest. Taken together, these data suggest that the inhibition of iNOS expression leads to a decrease of NO and cGMP levels thereby decreasing Cdc25B level. The reduced CDC25 B level leads to accumulation of Thr-14/Tyr-15 phosphorylated CDK1 level. As a result, Thr-161 phosphorylated CDK1 as well as cyclin B1 levels are decreased leading to maturation-promoting factor (MPF) inactivation. The inactive MPF finally induced meiotic resumption from diplotene stage in rat oocytes cultured in vitro.
Keywords
Introduction
Nitric oxides (NOs) act as a major signaling molecule and modulate various aspects of cell functions, including meiotic cell cycle of mammalian oocytes. 1 The NO is produced through three nitric oxide synthase (NOS) isoforms2,3 by oocytes as well as encircling granulosa cells and maintain meiotic arrest at diplotene stage in follicular oocytes.4,5 It is well known that the mammalian ovary expresses eNOS, iNOS, and nNOS4,6–9 but the inducible NOS (iNOS) is the major source of NO production in mouse and rat ovary10,11 and in mouse12–14 rat. 6
The role of NO during meiotic resumption from diplotene arrest in various mammalian species remains controversial. Few studies suggest that the increased level of NO induces meiotic resumption from diplotene arrest in mouse,7,15 rat, 16 porcine,9,17,18 and murine oocytes. 13 On the other hand, reduced level of NO is associated with meiotic resumption from diplotene arrest19,20 since NO donor maintained meiotic arrest, while iNOS inhibitor such as aminoguanidine (AG; an iNOS inhibitor) induced resumption from diplotene arrest in rat oocytes cultured in vitro. Further, lower concentration of a NO donor such as S-nitroso-l-acetyl penicillamine (SNAP; an NO donor, 0.01 mmol/L) induces meiotic resumption and higher concentrations (0.1 and 0.5 mmol/L) inhibit meiotic resumption from diplotene arrest in bovine oocytes cultured in vitro. 21
The NO produced through iNOS-mediated pathway is involved during various aspects of reproductive functions. NOS activity and NO presence are necessary for the correct course of meiotic maturation in the oocyte.7,13,18,22–25 The presence of NOS inhibitors in culture medium inhibits the meiotic resumption in mouse, 23 goat, 26 and cattle oocytes in vitro.21,25,27 NO is important for meiotic maturation of porcine oocytes.9,17,28 SNAP, a NO donor inhibits meiotic resumption in cattle,24,25 canine, 29 and porcine oocytes in vitro. 18 The high level of NO maintains meiotic arrest at diplotene stage, hence we hypothesized that the reduced level of intraoocyte NO may induce meiotic resumption from diplotene arrest possibly by modulating cyclic guanosine monophosphate (cGMP)-mediated pathway or directly by affecting Cdc25B activation which is required for maturation-promoting factor (MPF) activation. 30
Changes in the levels of signaling molecules regulate MPF activity and thereby meiotic cell cycle progression from diplotene arrest.6,19,20,31–33 The MPF is a complex of an enzymatic subunit p34cdc2 also known as cyclin-dependent kinase 1 (CDK1) and regulatory subunit cyclin B1.31,33,34 Growing body of evidences suggests that MPF inactivation does not solely dependent on cyclin B1 degradation. 35 The phosphorylation at Tyr-15/Thr-14 and dephosphorylation at Thr-161 residues of CDK1 make MPF inactive,36,37 while dephosphorylation at Tyr-15 and phosphorylation at Thr-161 residues activate MPF.33,38 However, a possible mechanism by which decreased NO modulates MPF activity during spontaneous resumption of meiosis remains unclear. Therefore, present study was aimed to analyze in vitro effects of NO donor, iNOS inhibitor, changes in NO, cGMP, Cdc25B phosphorylation status of CDK1, and cyclin B1 levels during meiotic resumption from diplotene arrest in rat oocytes cultured in vitro.
Materials and methods
Chemicals and culture media
All chemicals used in the present study were purchased from Sigma Chemical Co. (St. Louis, MO) unless stated otherwise. The M2 culture medium (M5910; Sigma) has widely been used to handle mammalian oocytes and embryos under in vitro culture conditions. Hence, in the present study we used M2 media (AL142) purchased from HiMedia Laboratories, Mumbai, India. This medium was HEPES buffered and contained lactic acid and sodium bicarbonate. The osmolarity of the liquid medium was 280 ± 10 mOsm and pH 7.0 ± 0.20, as per company manual data sheet.
Animals
Sexually immature female albino rats (Rattus norvegicus) of Charles Foster (CF) strain (22–24 day-old, 45 ± 5 g body weight) were housed in air-conditioned, light controlled rooms, with food and water available ad libitum. All procedures were in conformation to the stipulations of the University Animal Ethical Committee of Banaras Hindu University, Varanasi and in keeping with the Guidelines for the Care and Use of Laboratory Animals (NIH Publication).
Collection of diplotene stage oocytes
Immature female rats (22–24 days old; 45 ± 5 g body weight) were separated from existing colony and maintained in normal husbandry conditions with food and water ad libitum. To obtain diplotene stage oocytes, rats were given a single subcutaneous injection of 20 IU pregnant mare’s serum gonadotropin (PMSG) in 100 µL of sterile normal saline to promote growth and development of antral follicles in ovary. After 48 h of PMSG injection, rats were killed by cervical dislocation, ovaries were removed and transferred to a 35 mm petridish containing 2 mL of pre-warmed sterile M2 medium. Antral follicles (>0.8 mm diameter) were punctured with a sterile 26-gauge needle attached to a 1 mL syringe. Cumulus oocyte complexes (COCs) were isolated in pre-warmed medium containing 0.1 mmol/L of 3-isobutyl-1-methylxanthine (IBMX) to inhibit spontaneous meiotic resumption and then denuded using 0.01% (w/v) hyaluronidase in medium followed by repeated pipetting through a narrow-bore pipette. The 0.1 mmol/L IBMX (a reversible meiotic inhibitor) has been used to inhibit spontaneous resumption of meiosis in rat oocytes during collection and handling of COCs in vitro. 39 The denuded diplotene-arrested oocytes were washed quickly with fresh plain M2 medium to remove IBMX as well as hyaluronidase from culture medium and the used for in vitro studies. The average time for isolation and preparation of culture for denuded oocytes was 6 ± 2 min for all in vitro studies.
Effects of SNAP and AG on spontaneous resumption of meiosis
To increase intracellular NO level in the oocyte cultured in vitro, we used SNAP in the present study. The submillimolar concentration of SNAP has recently been used to inhibit spontaneous meiotic resumption from diplotene arrest in bovine oocytes cultured in vitro. 40 Hence to analyze the role of NO, we used SNAP as a NO donor and AG as iNOS inhibitor in the present study. A group of 20–22 oocytes were cultured separately in culture medium with or without various concentrations of SNAP (0.0, 0.1, 0.2, 0.3, and 0.4 mmol/L) and AG (0.0, 2.5, 5.0, 10.0, 20.0, and 40.0 µmol/L) in a CO2 incubator (Model; Galaxy 170 R, New Brunswick, Eppendorf AG, Hamburg, Germany) at 37℃, 95% humidity, and 5% CO2 for 3 h. At the end of incubation period, oocytes were removed, washed three times with plain M2 medium and transferred on to a grooved slide and then examined for morphological changes using a phase-contrast microscope (Nikon, Eclipse; E600, Tokyo, Japan) at 400X magnification.
Detection of iNOS expressions
The iNOS expression was detected in oocytes using anti-iNOS polyclonal antibody purchased from Santa Curz Biotechnology, MN, USA. For this purpose, a group of 20–22 oocytes from each group were prefixed with 3.7% buffered formaldehyde. Oocytes were permeabilized with triton X-100 (0.01% in phosphate-buffered saline [PBS]) for 10 min at 37℃ and then washed three times with pre-warmed PBS. The nonspecific sites were blocked using blocking buffer (2.5% PBS–bovine serum albumin [BSA] solution) at 37℃ for 30 min and then incubated with 100 µL of diluted (1:500 dilution in blocking buffer) iNOS polyclonal rabbit antibody (Santa Cruz Biotechnology Inc.) at 37℃ for 2 h and then after slides were washed five times with pre-warmed PBS, then exposed to 100 µL of fluorescein isothiocyanate (FITC)-labeled secondary antibody (1:1000 dilutions in blocking buffer) for 1 h at 37℃ in humidified chamber. Following incubation, slides were washed five times with pre-warmed PBS, mounted with VECTASHIELD fluorescence mounting media (Vector laboratories, US) for preventing photo bleaching and then observed under fluorescence microscope (Model, Ni-U, Nikon, Eclipse) at 520 nm at 400X magnification.
Quantitative analysis of NO concentration
The intracellular NO concentration was analyzed using NO assay kit purchased from R&D Systems (MN). Oocyte lysates were prepared and the total nitrite levels were analyzed following company manual protocol as published earlier. 19 In brief, all reagents, working standards, and samples were brought to room temperature before use. The 50 µL of reaction diluents was added to the blank well and 50 µL of nitrate standards or samples to remaining wells. The 25 µL of NADH and then 25 µL of diluted nitrate reductase were added to all wells. The plate was incubated at 37℃ for 30 min. At the end of incubation, 50 µL of Griess reagent I and then 50 µL of Griess reagent II were added. The content was mixed well by tapping the side of the plate and then incubated for 10 min at room temperature. At the end of the incubation period, the optical density (OD) was determined using a microplate reader (Micro Scan MS5608A, ECIL, Hyderabad, India) set at 540 nm with wavelength correction at 690 nm.
Quantitative analysis of cGMP concentration
The intracellular cGMP concentration was analyzed using cGMP assay kit purchased from R&D Systems (MN) as per company manual protocol. In brief, oocyte lysates were collected from different groups (100 oocytes from each group) were transferred to a microcentrifuge tube containing 100 µL of hypotonic lysis buffer (5 mmol/L Tris, 20 mmol/L EDTA, 0.5% Triton X-100, pH 8) for 1 h on ice for lysis. The lysates were centrifuged at 10,000 x g at 4℃ for 15 min and clear supernatant was used for the quantitative estimation of cGMP concentration by colorimetric assay as per company manual protocol. Reagents, samples, and standards were prepared according to instruction manual. In brief, 150 µL of calibrator diluent was added to nonspecific binding (NSB) wells and 100 µL of calibrator diluent was added to zero standard wells. The 100 µL of standard, control, or samples were added to the remaining wells. The 50 µL of cGMP conjugate was added to each well. Further, 50 µL of primary antibody was added to each well excluding NSB wells and then incubated for 3 h at room temperature. The plate was aspirated and washed four times with wash buffer and then 200 µL substrate solution was added to each well and then incubated for 30 min at room temperature. At the end of incubation period, 50 µL stop solution was added to each well and reading was taken using microplate reader set at 450 nm within 10 min. Three independent samples were run in triplicate and to avoid inter-assay and intra-assay variation was found to be 2.1%.
Detection of Cdc25B expression
The Cdc25 B level was detected in oocytes using anti-Cdc25B polyclonal antibody purchased from Santa Curz Biotechnology. For this purpose, a group 20–22 oocytes either arrested at diplotene stage and resumed meiosis from diplotene arrest were prefixed with 3.7% buffered formaldehyde. Oocytes were permeabilized with triton X-100 (0.01% in PBS) for 10 min at 37℃ and then washed three times with pre-warmed PBS. The anti-Cdc25B ([H-85] polyclonal antibody raised against amino acids 1-85 representing full length Cdc25B) were purchased from Santa Cruz Biotechnology. The nonspecific sites were blocked using blocking buffer (2.5% PBS–BSA solution) at 37℃ for 30 min and then exposed to 100 µL of their respective primary antibodies (1:500 dilution in blocking buffer) at 37℃ for 2 h. After five washes with pre-warmed PBS, slides were exposed to 100 µL of FITC-labeled secondary antibody (1:1000 dilutions in blocking buffer) for 1 h at 37℃ in humidified chamber. After 1 h of incubation, slides were washed five times with pre-warmed PBS, mounted with VECTASHIELD fluorescence mounting media for preventing photo bleaching and then observed under fluorescence microscope at 520 nm at 400X magnification.
Detection of pThr-14/Tyr-15, pTh-161, and total phosphorylation status of CDK1
The total as well as specific phosphorylation status at Thr-14/Tyr-15 and Th-161 were detected using anti-pThr-14/Tyr-15, anti-pThr-161, and anti-PSTAIR polyclonal antibody purchased from Santa Curz Biotechnology. For this purpose, a group of 20–22 oocytes from each group was prefixed with 3.7% buffered formaldehyde. Oocytes were permeabilized with triton X-100 (0.01% in PBS) for 10 min at 37℃ and then washed three times with pre-warmed PBS. The p-Cdc2 p34 (Thr-161) polyclonal antibody raised against a short amino acid sequence containing phosphorylated Thr-161 of Cdc2 p-34 and phosphorylated Tyr-15 of Cdc2 p-34 purchased from Santa Cruz Biotechnology, Inc. The nonspecific sites were blocked using blocking buffer (2.5% PBS–BSA solution) at 37℃ for 30 min and then exposed to 100 µL of their respective primary antibody (1:500 dilution in blocking buffer) at 37℃ for 2 h. After five washes with pre-warmed PBS, slides were exposed to 100 µL of FITC-labeled secondary antibody (1:1000 dilutions in blocking buffer) for 1 h at 37℃ in humidified chamber. After 1 h of incubation, slides were washed five times with pre-warmed PBS, mounted with VECTASHIELD fluorescence mounting media for preventing photo bleaching and then observed under fluorescence microscope at 520 nm at 400X magnification.
Detection of cyclin B1 expression
The cyclin B1 level was analyzed using anti-cyclin B1 polyclonal antibody purchased from Santa Curz Biotechnology. For this purpose, 20–22 oocytes from each group were prefixed with 3.7% buffered formaldehyde. Oocytes were permeabilized with triton X-100 (0.01% in PBS) for 10 min at 37℃ and then washed three times with pre-warmed PBS. The anti-cyclin B1 (H-433) polyclonal antibody raised against amino acids 1-433 representing full length cyclin B1 were purchased from Santa Cruz Biotechnology, Inc. The nonspecific sites were blocked using blocking buffer (2.5% PBS–BSA solution) at 37℃ for 30 min and then exposed to 100 µL of their respective primary antibodies (1:500 dilution in blocking buffer) at 37℃ for 2 h. After five washes with pre-warmed PBS, slides were exposed to 100 µL of FITC-labeled secondary antibody (1:1000 dilutions in blocking buffer) for 1 h at 37℃ in humidified chamber. After 1 h of incubation, slides were washed five times with pre-warmed PBS, mounted with VECTASHIELD fluorescence mounting media for preventing photo bleaching and then observed under fluorescence microscope at 520 nm at 400X magnification.
Statistical analyses
Data are expressed as mean ± standard error of mean (SEM) of three independent experiments. All percentage data were subjected to arcsine square-root transformation before statistical analysis. Data are analyzed either by Student’s t-test or one-way analysis of variance (ANOVA) followed by post hoc multiple test, i.e. Bonferroni test using SPSS software, Version 17.0 (SPSS, Inc. Chicago, IL). A probability of P < 0.05 was considered as statistically significant.
Results
SNAP inhibits spontaneous meiotic resumption
As shown in Figure 1, a NO donor such as SNAP inhibited spontaneous resumption of meiosis from diplotene arrest in concentration-dependent manner (one-way ANOVA: F = 51.59, P < 0.001). The maximum inhibition of spontaneous resumption of meiosis from diplotene arrest was noticed if the oocytes were exposed to 0.4 mmol/L SNAP (97.5%), however, majority of oocytes underwent spontaneous resumption from meiosis after 3 h of in vitro culture in plain medium (18.83%).
SNAP inhibited spontaneous resumption of meiosis in diplotene-arrested oocytes in a concentration-dependent manner in vitro. Diplotene-arrested oocytes were exposed to various concentrations of SNAP for 3 h in vitro. Data are mean ± SEM of three independent experiments and analyzed by one-way ANOVA followed by Bonferroni post hoc test. *P < 0.001. (A color version of this figure is available in the online journal.)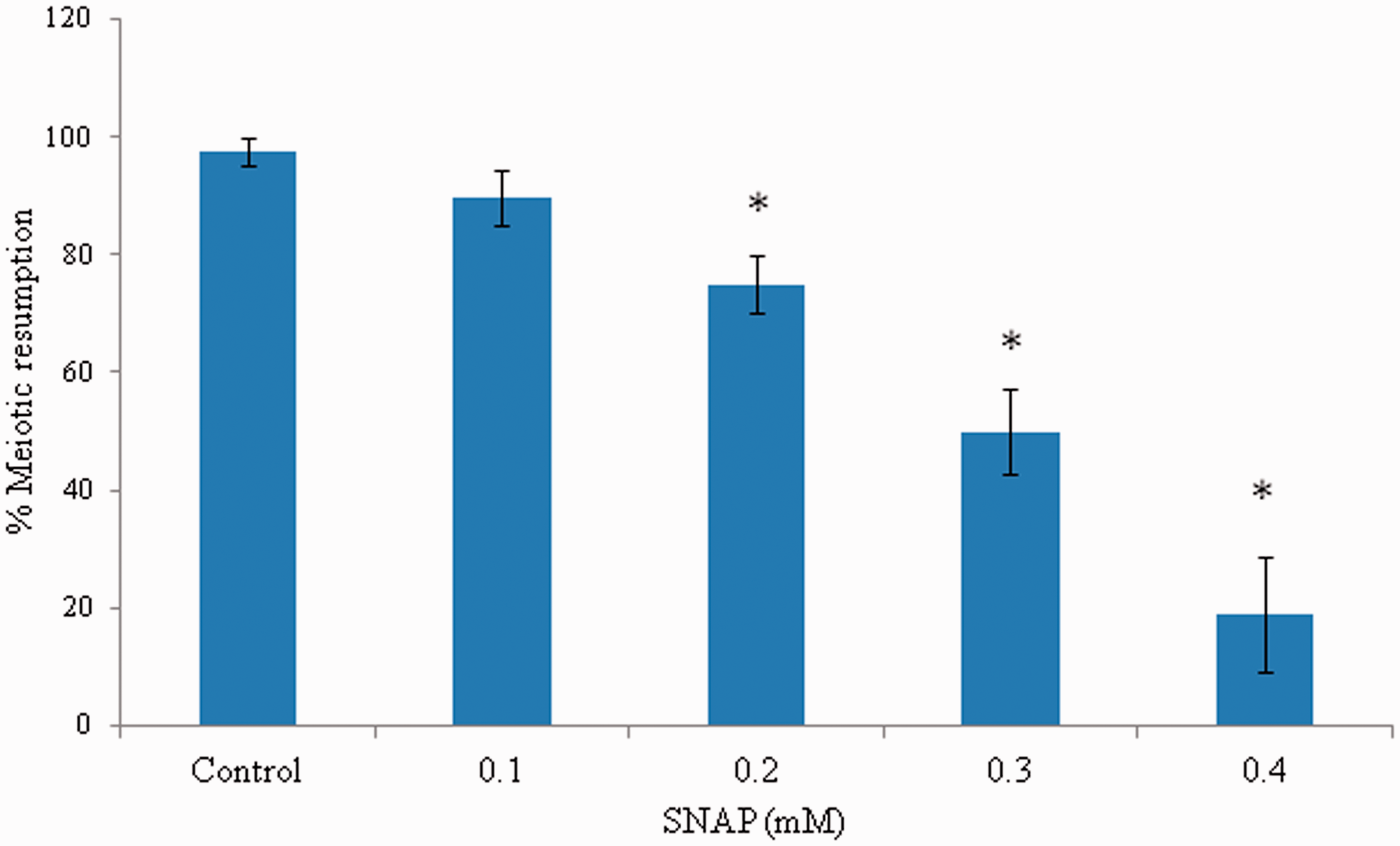
AG induces meiotic resumption
As shown in Figure 2, addition of 1 mmol/L IBMX in M2 medium at the time of collection of prevented meiotic resumption in majority of oocytes and only (30.0 ± 3.52%) oocytes underwent spontaneous resumption of meiosis in vitro. However, AG induced spontaneous resumption of meiosis in a concentration-dependent manner. The maximum spontaneous resumption of meiosis was noticed if the oocytes were exposed 40.0 µmol/L of AG for 3 h in vitro. Three independent experiments were conducted to confirm these results.
AG induced resumption of meiosis from diplotene arrest in IBMX-treated oocytes cultured in vitro. Diplotene-arrested oocytes were collected in medium containing 0.1 mmol/L IBMX to inhibit spontaneous resumption of meiosis at the time of collection. Oocytes were then washed three times with fresh medium and then exposed to various concentrations of AG for 3 h in vitro. Data are mean ± SEM of three independent experiments and analyzed by Student’s t-test. *P < 0.001, significantly decreased as compared to diplotene-arrested oocytes. (A color version of this figure is available in the online journal.)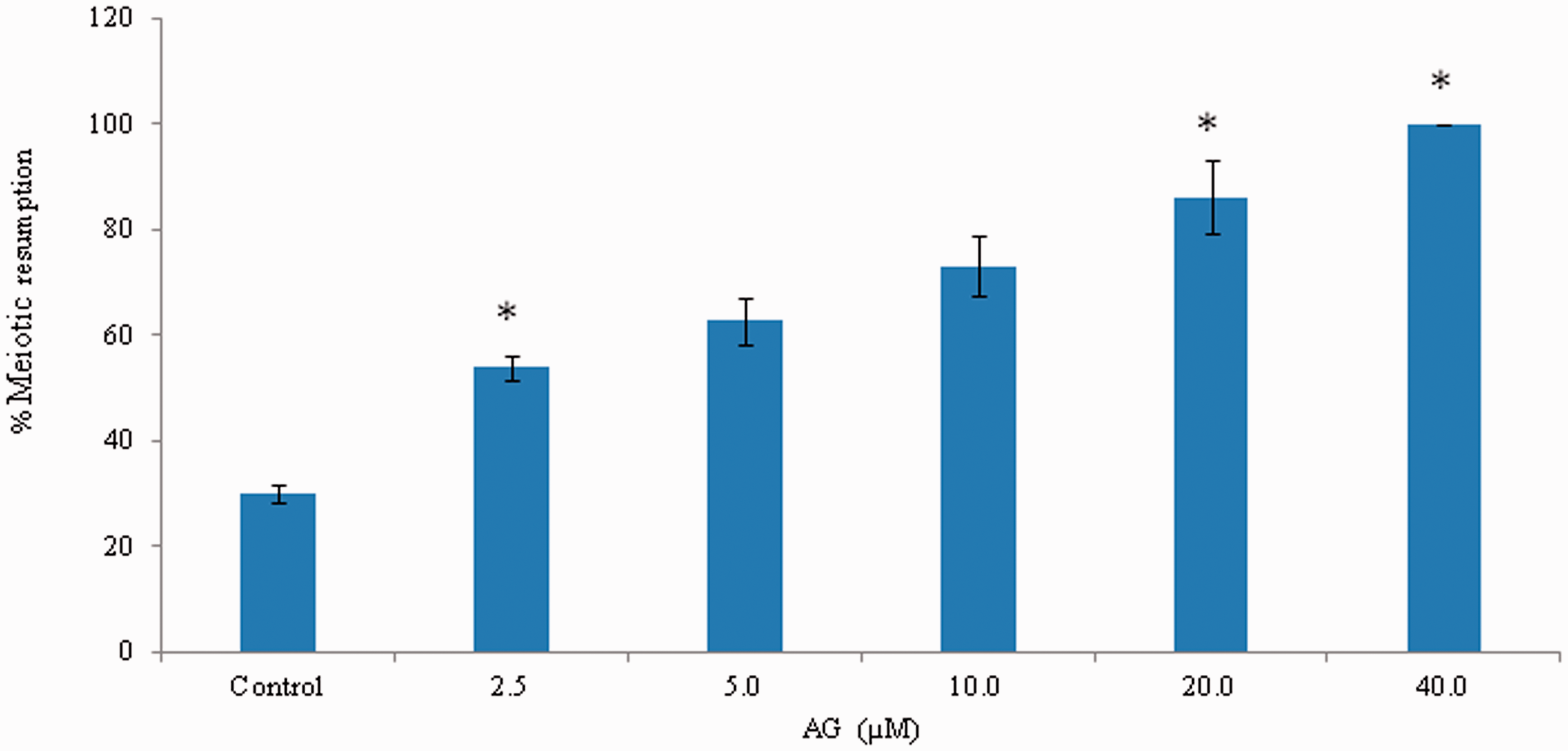
Decrease of iNOS expression and NO level results spontaneous meiotic resumption
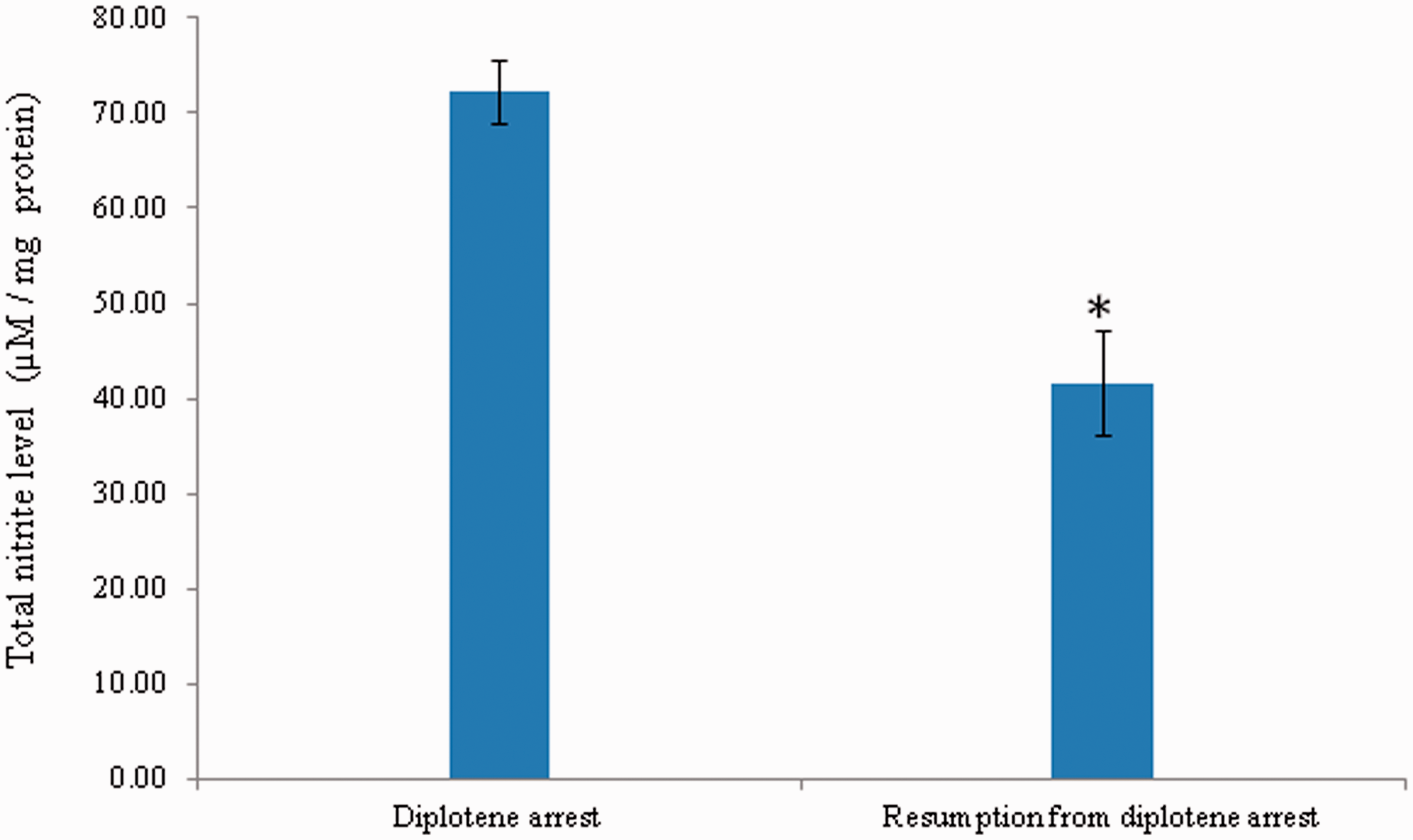
As shown in Figure 3, a significant reduction in the level of iNOS expression was observed in oocytes that underwent resumption of meiosis after 3 h of in vitro culture in response to AG treatment (Figure 3b) as compared to diplotene-arrested oocytes (Figure 3a). The corrected total cell fluorescence (CTCF) analysis using Image J software further confirms above observations (Figure 3c). The intraoocyte NO concentration was significantly (p < 0.05) decreased in oocytes that underwent spontaneous resumption of meiosis from diplotene arrest (41.65 ± 5.53 µmol/mg protein) as compared to diplotene-arrested oocytes (72.18 ± 0.01 µmol/mg protein) (Figure 4). Three independent samples were run in triplicate and to avoid inter-assay and intra-assay variation was found to be 2.4%.
Representative photographs showing the iNOS expression in oocytes cultured in vitro. The AG treatment significantly reduced iNOS fluorescence intensity (b; arrow) as compared to diplotene-arrested oocytes (a; arrow). Encircling cumulus cells showing iNOS fluorescence (red arrows). The CTCF analysis of three independent groups of oocytes further support above observations (c). Data are mean ± SEM of three independent experiments and analyzed by Student’s t-test. *P < 0.001, significantly decreased as compared to diplotene-arrested oocytes. (Bar = 20 µmol/L). (A color version of this figure is available in the online journal.) A significant reduction of intraoocyte NO level mediates spontaneous resumption of meiosis in diplotene-arrested oocytes cultured in vitro. Data are mean ± SEM of three independent experiments and analyzed by one-way ANOVA followed by Bonferroni post hoc test. *P < 0.05. (A color version of this figure is available in the online journal.)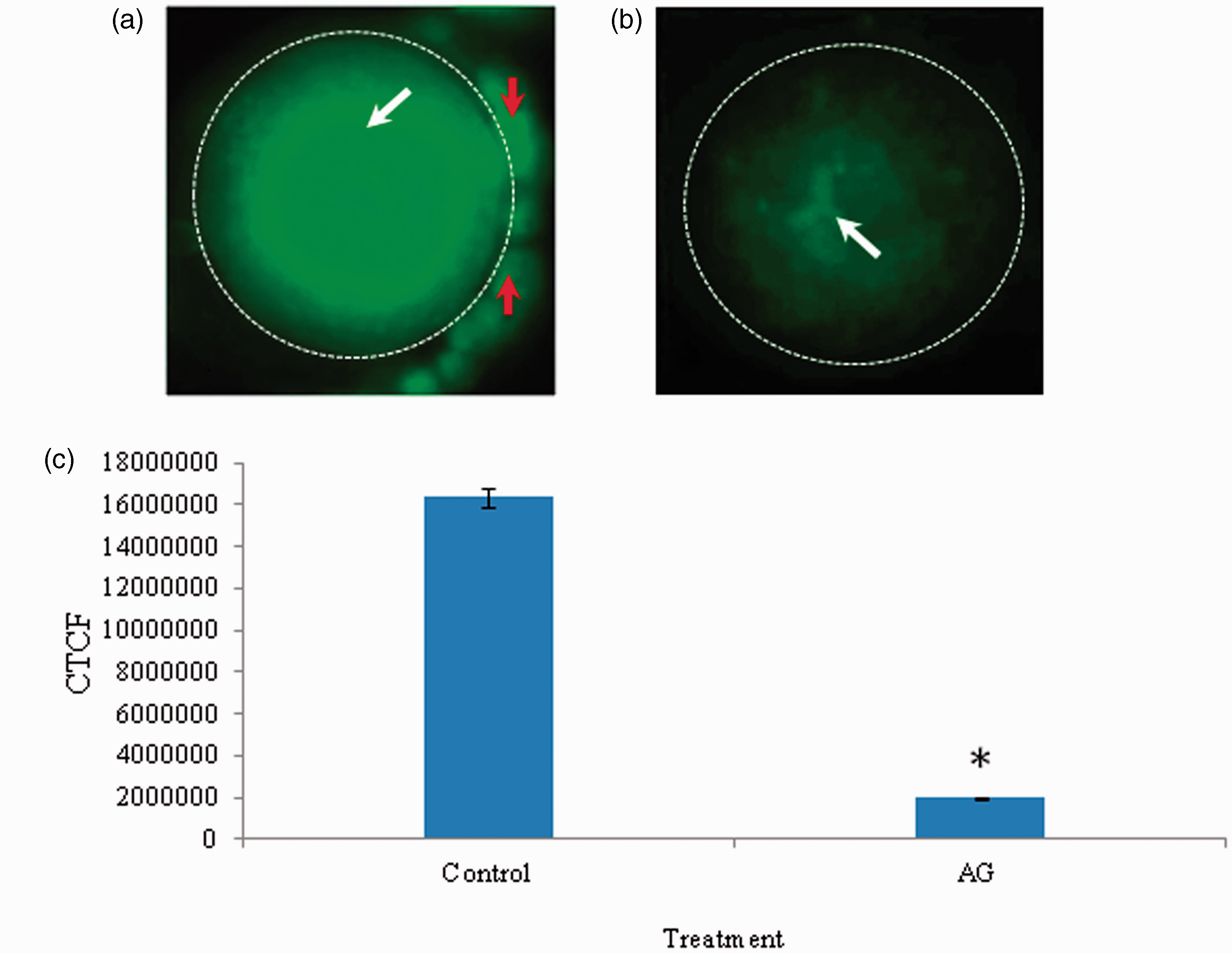
Decrease of intraoocyte cGMP level results spontaneous meiotic resumption
As shown in Figure 5, intraoocyte cGMP concentration was significantly (P < 0.05) decreased in oocytes that underwent spontaneous resumption of meiosis from diplotene arrest (0.28 ± 0.002 pmol/L/oocyte) as compared to diplotene-arrested oocytes (0.16 ± 0.038 pmol/L/oocyte). Three independent samples were run in triplicate to avoid inter-assay intra-assay variation and intra-assay variation was found to be 1.9%.
A significant decrease of intraoocyte cGMP is associated with spontaneous resumption of meiosis in diplotene-arrested oocytes cultured in vitro. Data are mean ± SEM of three independent experiments. *P < 0.001, significantly decreased as compared to diplotene-arrested oocytes. (A color version of this figure is available in the online journal.)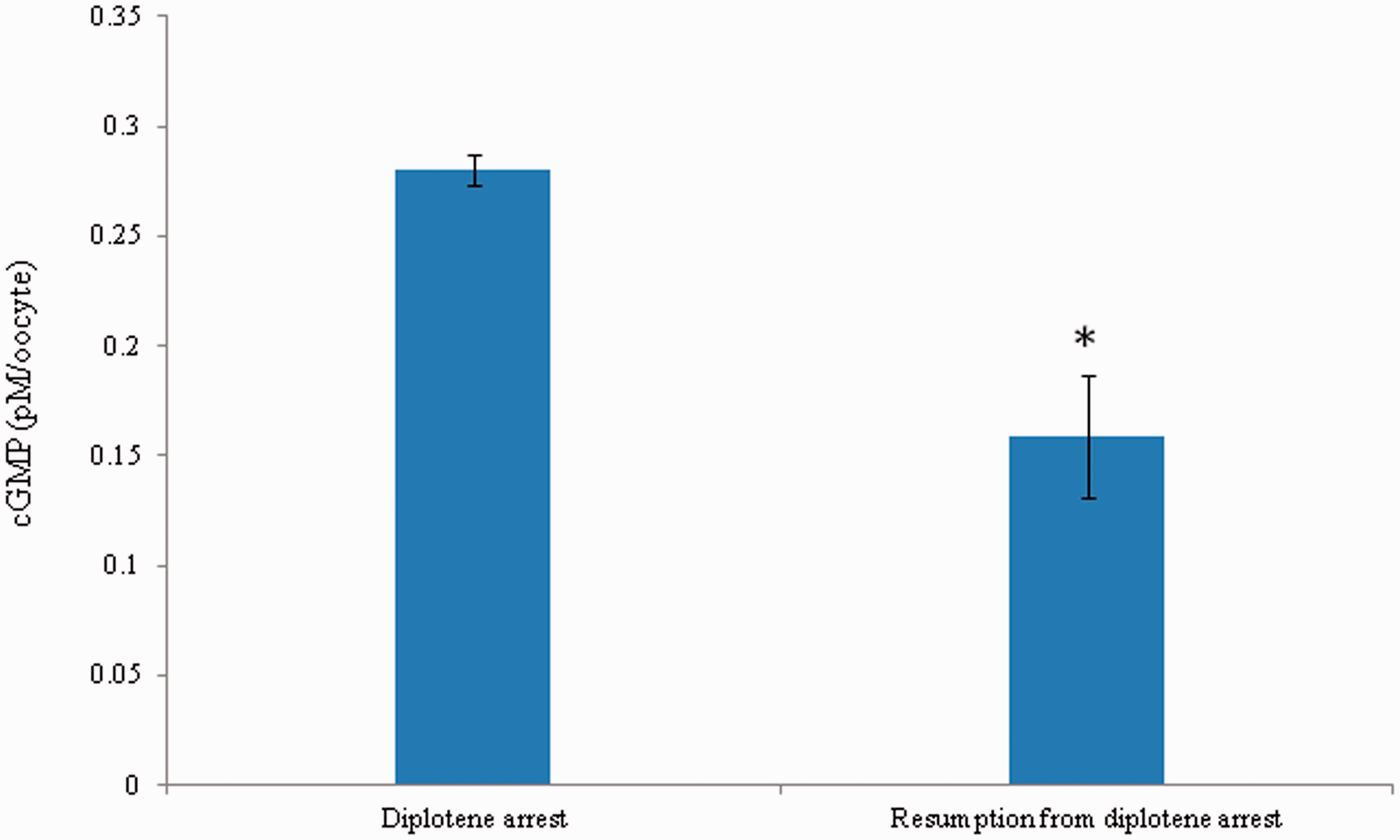
Reduction of Cdc25B level results spontaneous meiotic resumption
As shown in Figure 6, a significant reduction in the level of Cdc25B level was observed in oocytes that underwent resumption from diplotene arrest after 3 h of in vitro culture (Figure 6a) as compared to diplotene-arrested oocytes (Figure 6b). The CTCF analysis using Image J software further confirms above observations (Figure 6c). Three independent experiments were conducted to confirm these results.
Representative photographs showing changes in level of Cdc25B in oocytes. A significant reduction of Cdc25B fluorescence intensity (b; arrow) is associated with spontaneous resumption of meiosis as compared to diplotene-arrested oocytes (a). The CTCF analysis of three independent groups of oocytes further support above observations (c). Data are mean ± SEM of three independent experiments and analyzed by Student’s t-test. *P < 0.001, significantly decreased as compared to diplotene-arrested oocytes. (Bar = 20 µmol/L). (A color version of this figure is available in the online journal.)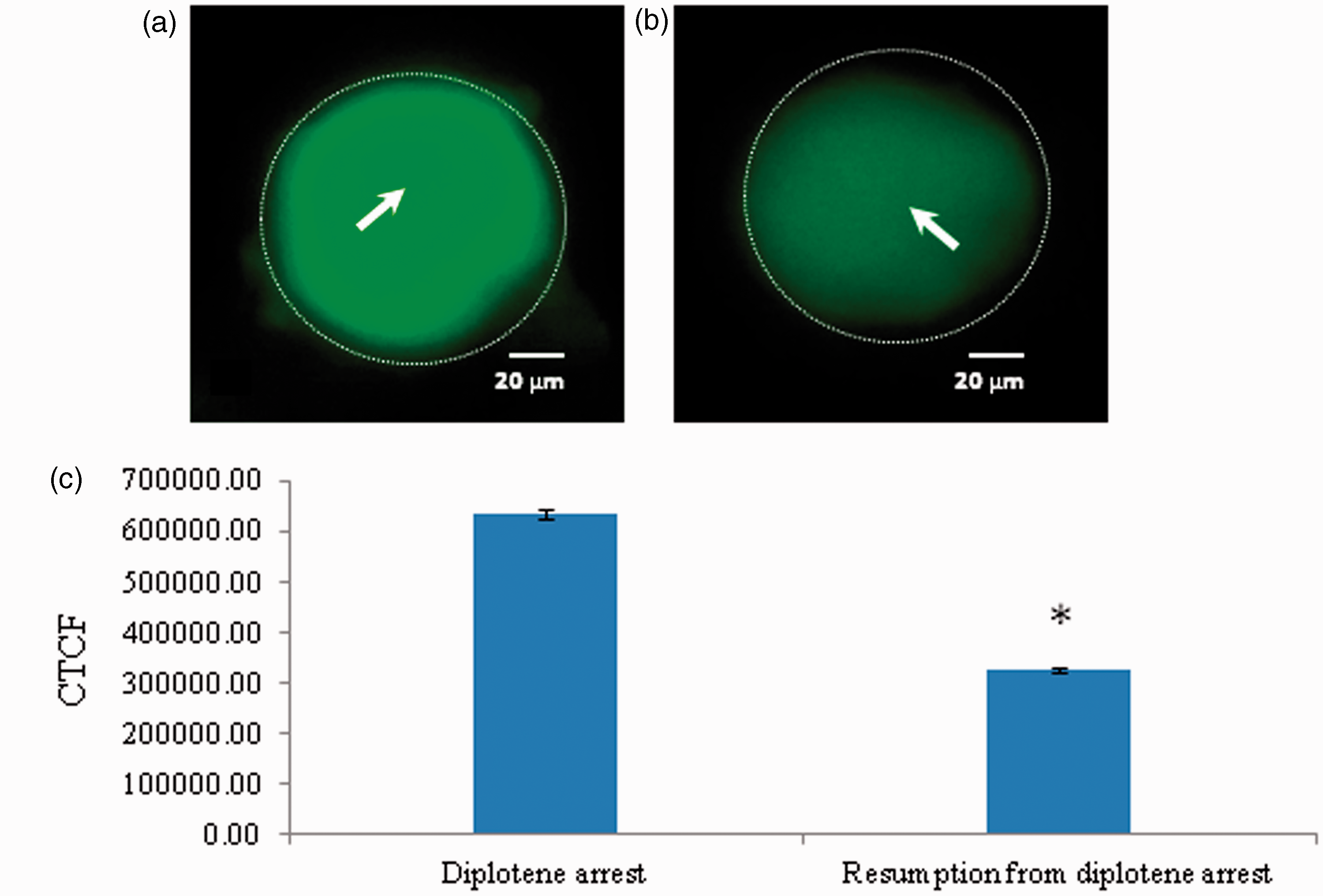
An accumulation of pThr-14/Tyr15 results spontaneous meiotic resumption
As shown in Figure 7, a significant increase of Thr-14/Tyr-15 phosphorylated CDK1 level was observed in oocytes that underwent spontaneous resumption of meiosis after 3 h of in vitro culture (Figure 7b) as compared to diplotene stage oocytes (Figure 7a). The CTCF analysis using Image J software further confirms above observations (Figure 7c). On the other hand, a significant decrease of Thr-161 phosphorylated CDK1 was observed in oocytes that underwent spontaneous resumption of meiosis after 3 h of in vitro culture (Figure 8b) as compared to diplotene stage oocytes (Figure 8a). The CTCF analysis using Image J software further confirms above observations (Figure 8c). However, total phosphorylation of CDK1 remained unchanged during spontaneous resumption from diplotene arrest (Figure 9b) when compared to diplotene stage oocytes (Figure 9a). This result is further supported by the CTCF analysis (Figure 9c).
Representative photographs showing changes in level of Thr-14/Tyr-15 phosphorylated CDK1 in oocytes. An increased level of Thr-14/Tyr-15 phosphorylated CDK1 fluorescence intensity (b; arrow) is associated with spontaneous resumption of meiosis as compared to diplotene-arrested oocytes a; arrow). The CTCF analysis of three independent groups of oocytes further support above observations (c). Data are mean ± SEM of three independent experiments and analyzed by Student’s t-test. *P < 0.001, significantly increased as compared to diplotene-arrested oocytes. (Bar = 20 µmol/L). (A color version of this figure is available in the online journal.) Representative photographs showing changes in level of Thr-161 phosphorylated CDK1 in oocytes. Reduction of Thr-161 phosphorylated CDK1 fluorescence intensity (b; arrow) is associated with spontaneous resumption of meiosis from diplotene arrest as compared to control (a). The CTCF analysis of three independent groups of oocytes further support above observations (c). Data are mean ± SEM of three independent experiments and analyzed by Student’s t-test. *P < 0.001, significantly decreased as compares to diplotene-arrested oocytes. (Bar = 20 µmol/L). (A color version of this figure is available in the online journal.) Representative photographs showing the general phosphorylation status of CDK. The general phosphorylation status of CDK1 remains unchanged in diplotene-arrested (a; arrow) as well as in oocytes that underwent spontaneous resumption of meiosis from diplotene arrest (b). The CTCF analysis of three independent groups of oocytes further support above observations (c). Data are mean ± SEM of three independent experiments and analyzed by Student’s t-test. (Bar = 20 µmol/L). (A color version of this figure is available in the online journal.)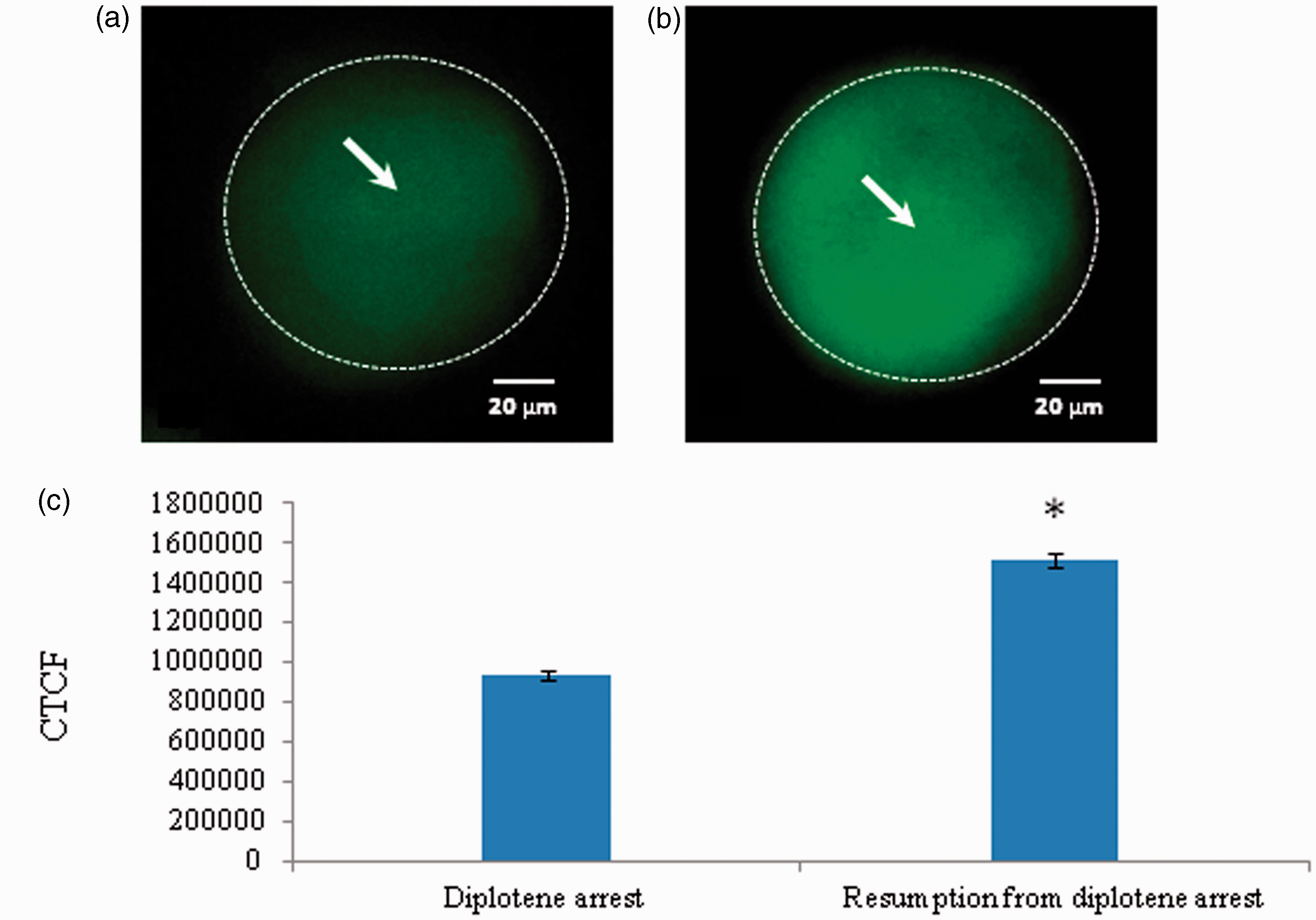
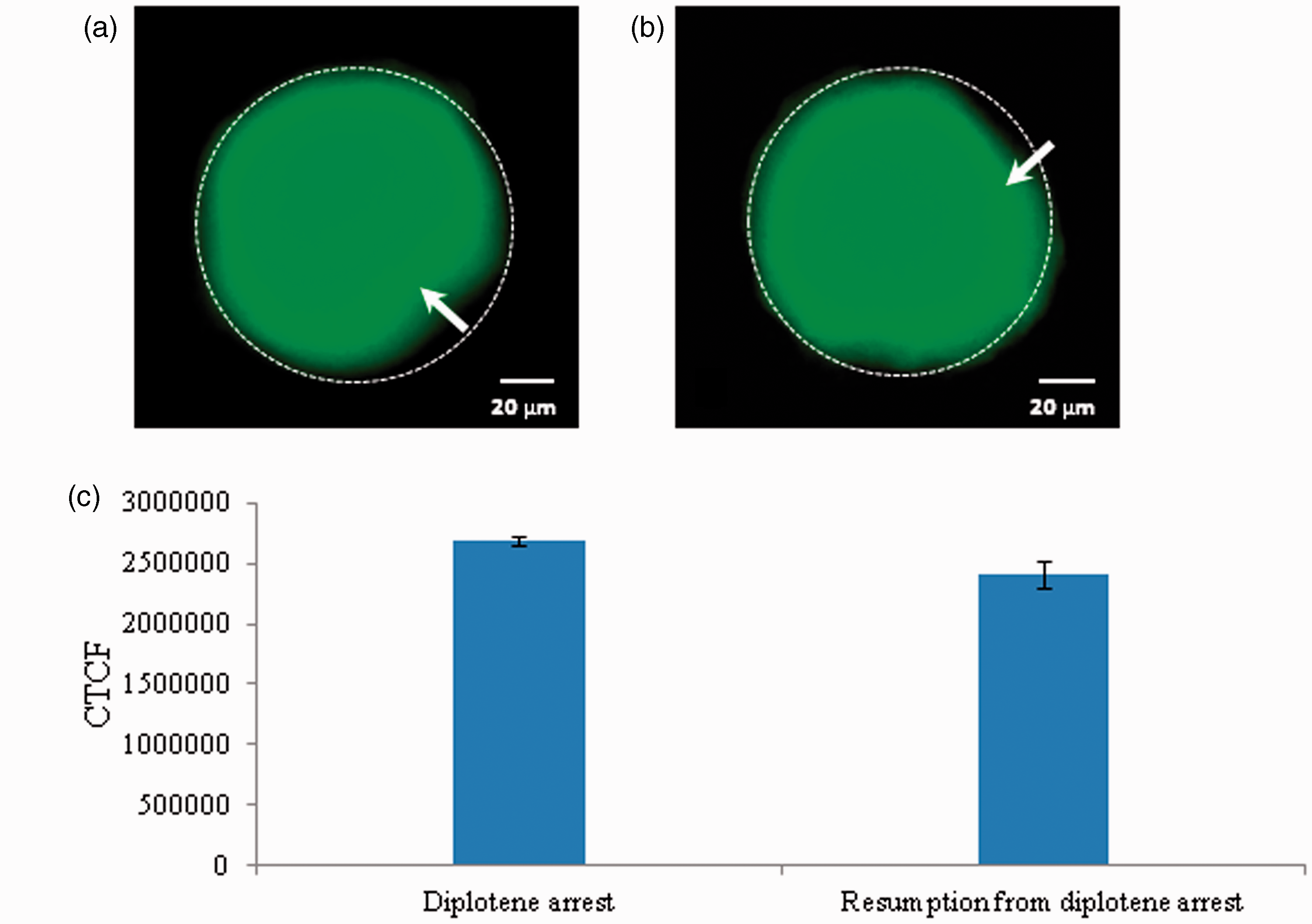
Reduced level of cyclin B1 results in spontaneous meiotic resumption
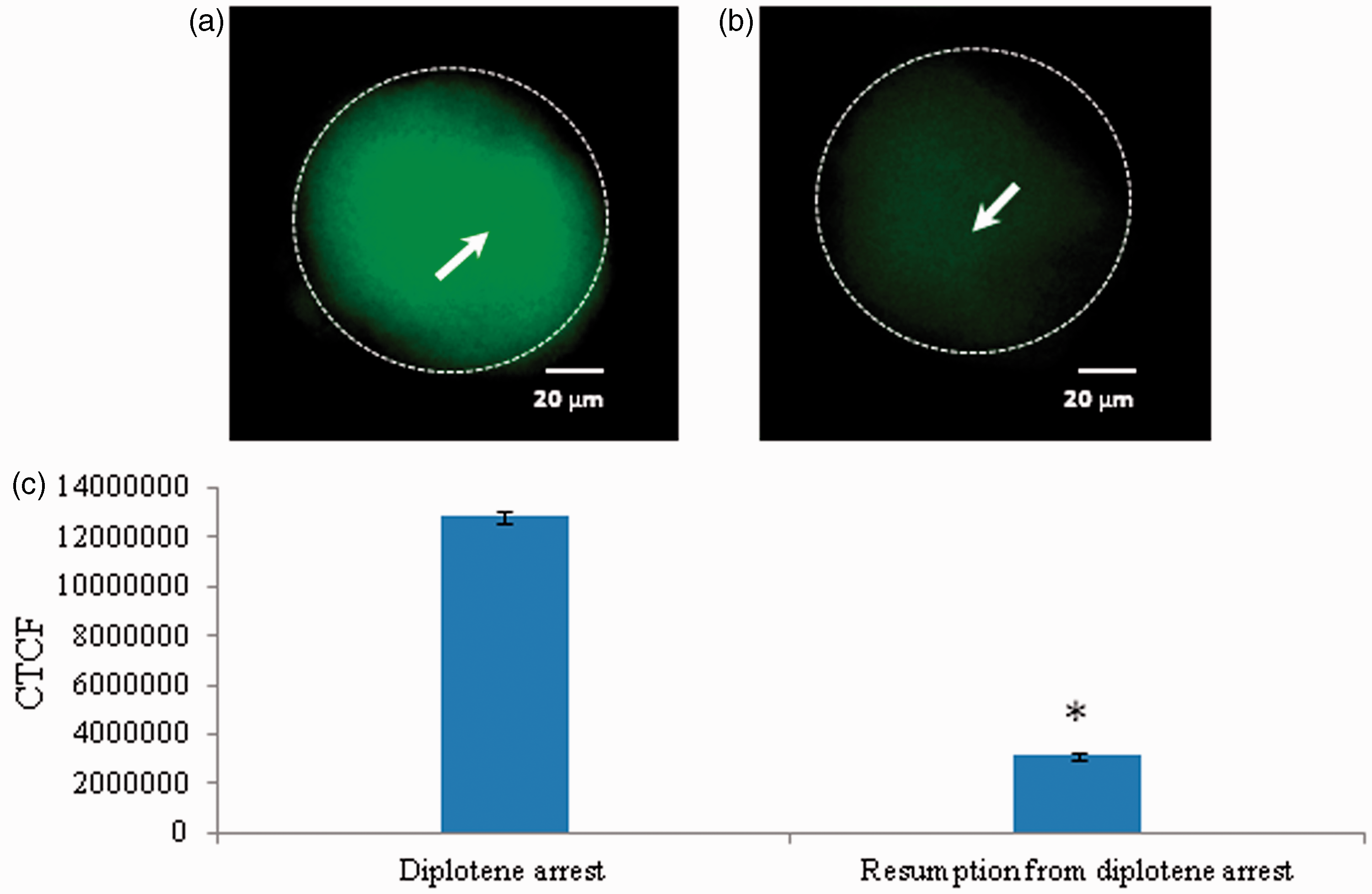
As shown in Figure 10, a significant reduction of cyclin B1 level was associated with spontaneous meiotic resumption from diplotene arrest (Figure 10b) as compared to diplotene-arrested oocytes (Figure 10a). The CTCF analysis using image J software further confirms above observations (Figure 10c).
Representative photographs showing changes in cyclin B1 level in oocytes. A significant decrease of cyclin B1 fluorescence intensity (b; arrow) is associated with spontaneous resumption of meiosis from diplotene arrest as compared to control (a). The CTCF analysis of three independent groups of oocytes further support above observations (c). Data are mean ± SEM of three independent experiments and analyzed by Student’s t-test. *P < 0.001, significantly decreased as compared to diplotene-arrested oocytes. (Bar = 20 µmol/L). (A color version of this figure is available in the online journal.)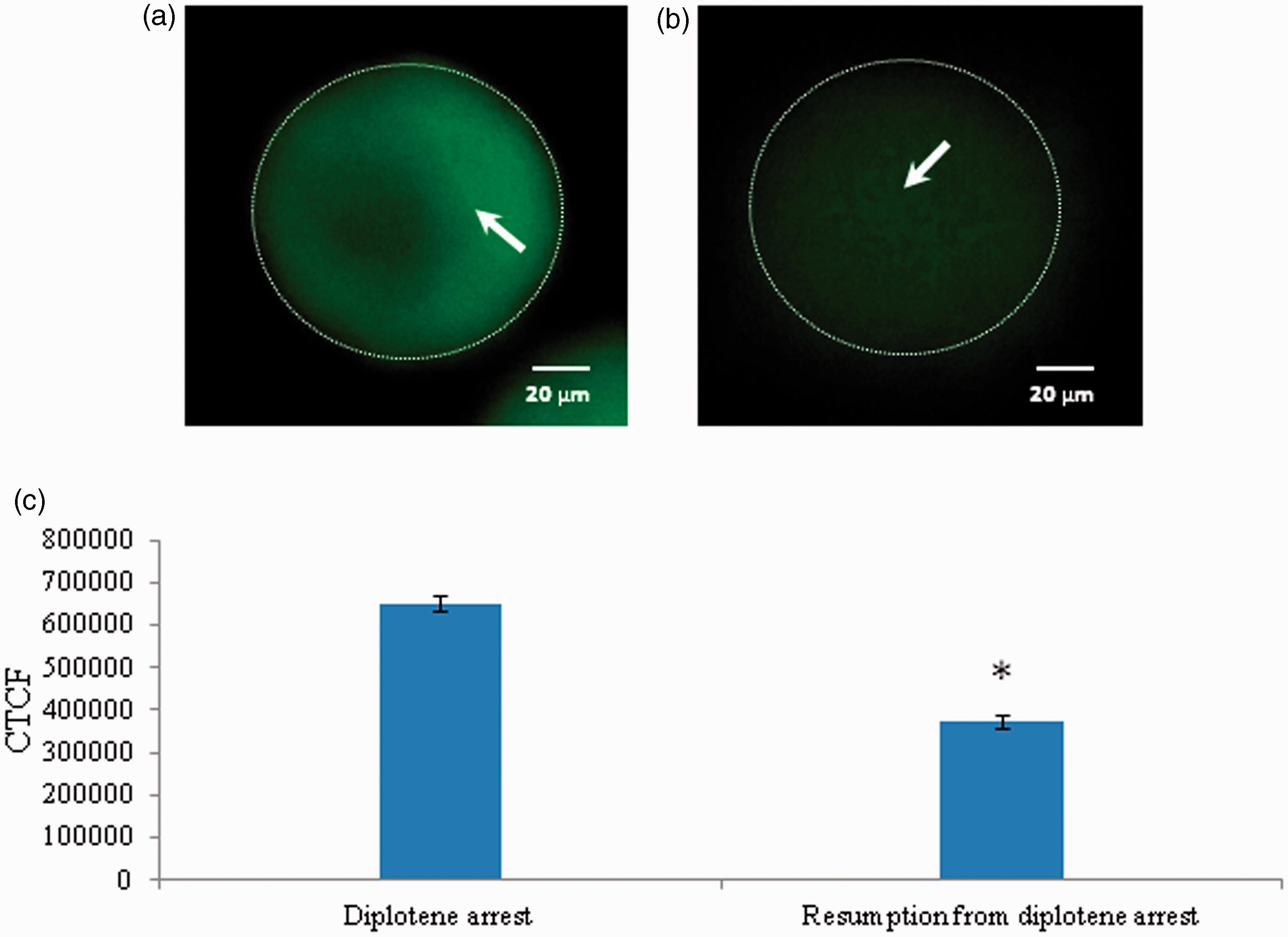
Discussion
The NO is one of the major signaling molecules that play an important role in ovarian physiology but its role of NO in the regulation of meiotic cell cycle remains controversial. Few studies suggest that high level of NO induces meiotic resumption from diplotene arrest in mouse,7,15 rat, 16 porcine,9,17,18 and murine oocytes. 13 On the other hand, NO inhibits meiotic resumption from diplotene arrest in rat oocytes 41 and a reduced level of NO triggers meiotic resumption from diplotene arrest in rat oocytes cultured in vitro. 19 Data of the present study revealed that a NO donor SNAP inhibited spontaneous resumption of meiosis in a concentration-dependent manner. The quantitative analysis further revealed that a high level of intraoocyte NO is required for the maintenance of diplotene stage, while a significant decrease of NO level was associated with meiotic resumption from diplotene arrest. These data corroborate with previous observations that increase of NO inhibits spontaneous meiotic resumption and decrease in its level induces meiotic resumption in rat oocytes cultured in vitro.4,19
The NO is produced by encircling granulosa cells and transferred to the oocyte via gap junctions, 42 however, oocyte may also generate NO sufficient to maintain meiotic arrest at diplotene arrest inside the follicular microenvironment. 42 Although, NO produced by three NOS isoforms in mammalian ovary,4,6–9 NO produced through iNOS-mediated pathway plays a major role in meiotic cell cycle regulation of diplotene-arrested oocyte. 4 Our results suggest that a specific iNOS inhibitor AG induced meiotic resumption from diplotene arrest in a concentration-dependent manner in oocytes pre-treated with IBMX. However, AG significantly inhibited iNOS expression in oocytes that underwent meiotic resumption from diplotene arrest. The CTCF analysis of fluorescence intensity further confirmed our results. These results further confirms our previous observations that a decrease of iNOS expression and thereby NO level is associated with meiotic resumption from diplotene arrest in rat oocytes. 19
The mechanism by which reduced level of NO mediates spontaneous meiotic resumption in diplotene-arrested oocytes remains poorly understood. There are two possibilities by which NO could mediate spontaneous meiotic resumption. The reduced level of NO may alter either cGMP or Cdc25B level or both and thereby MPF activity during spontaneous meiotic resumption from diplotene arrest. Our results suggest that the maintenance of meiotic arrest in oocytes at diplotene stage is associated with high level of cGMP, while reduction in its level was associated with spontaneous resumption of meiosis from diplotene arrest. Further, Cdc25B expression level was significantly reduced in oocytes that underwent spontaneous resumption of meiosis from diplotene arrest. The high level of intraoocyte cGMP maintains meiotic arrest at diplotene stage in mouse oocyte,20,43 while decrease of intraoocyte cGMP as well as Cdc25B levels triggers meiotic resumption from diplotene arrest.4,20,23,30,44,45 Taken together, these data suggest that the decrease of both cGMP as well as Cdc25B levels during spontaneous resumption of meiosis from diplotene arrest possibly by modulating MPF activity.
Cdc25B is involved in activation of CDK1–cyclin B and Cdc25 phosphatases that promote CDC2–cyclin B complex assembly 30 and demonstrated that in human transformed cells, Cdc25B, phosphatases have an effect on timing and efficiency of cyclin–kinase complex formation. An over expression of Cdc25B promotes earlier assembly and activation of Cdc25B-cyclin B complexes, whereas repression of these phosphatases has a reverse effect, leading to a substantial decrease in amounts of cyclin B-bound CDK1 in mitosis. 46 Taken together, our data suggest that the MPF complex assembly and maintenance is assured by a high level of intraoocyte Cdc25B.
The Cdc25B dephosphorylates Tyr-15 residue of CDK1 and cyclin-dependent kinase activating kinase phosphorylates Thr-161 residue of CDK1 and makes MPF active.47–49 The inhibition of Cdc25B induces Thr-14/Tyr-15 phosphorylation of CDK1 and leads to MPF inactivation.33,50–52 Based on these findings, we propose that the reduced level of Cdc25B might have induced accumulation of pThr-14/Tyr-15 but not pThr-161 of CDK1 and thereby spontaneous resumption of meiosis from diplotene arrest. Our results suggest that a significant increase in the Thr-14/Tyr-15 phosphorylated CDK1 level was in oocytes that underwent spontaneous resumption of meiosis from diplotene arrest. On the other hand, Thr-161 phosphorylated CDK1 level was significantly reduced during spontaneous resumption of meiosis. The total phosphorylation status of CDK1 remained unchanged. Further, cyclin B1 level was significantly reduced in oocytes that underwent spontaneous resumption of meiosis. These data suggest that reduced Cdc25B level induced accumulation of pThr-14/Tyr-15 but not pThr-161 of CDK1 level and induced spontaneous resumption of meiosis in rat oocytes cultured in vitro.
Conclusions
Data presented in the present study suggest that a decrease iNOS expression and thereby NO level leads to the reduction of intraoocyte cGMP as well as Cdc25B levels. The reduced Cdc25B level was associated with an accumulation of pThr-14/Tyr-15 CDK1 and reduced pThr-161 of CDK1 as well as cyclin B1 levels leading to MPF inactivation during spontaneous meiotic resumption from diplotene arrest in rat oocytes cultured in vitro.
Footnotes
Author contributions
All authors participated in the writing, review and editing of this manuscript.
ACKNOWLEDGMENT
This study was financially supported by Department of Science and Technology, Ministry of Science and Technology, Govt. of India.