
Abstract
Kallmann syndrome, a form of idiopathic hypogonadotropic hypogonadism, is characterized by developmental abnormalities of the reproductive system and abnormal olfaction. Despite association of certain genes with idiopathic hypogonadotropic hypogonadism, the genetic inheritance and expression are complex and incompletely known. In the present study, seven Kallmann syndrome pedigrees in an ethnic Han Chinese population were screened for genetic mutations. The exons and intron–exon boundaries of 19 idiopathic hypogonadotropic hypogonadism (idiopathic hypogonadotropic hypogonadism)-related genes in seven Chinese Kallmann syndrome pedigrees were sequenced. Detected mutations were also tested in 70 sporadic Kallmann syndrome cases and 200 Chinese healthy controls. In pedigrees 1, 2, and 7, the secondary sex characteristics were poorly developed and the patients’ sense of smell was severely or completely lost. We detected a genetic mutation in five of the seven pedigrees: homozygous KAL1 p.R191ter (pedigree 1); homozygous KAL1 p.C13ter (pedigree 2; a novel mutation); heterozygous FGFR1 p.R250W (pedigree 3); and homozygous PROKR2 p.Y113H (pedigrees 4 and 5). No genetic change of the assayed genes was detected in pedigrees 6 and 7. Among the 70 sporadic cases, we detected one homozygous and one heterozygous PROKR2 p.Y113H mutation. This mutation was also detected heterozygously in 2/200 normal controls and its pathogenicity is likely questionable. The genetics and genotype–phenotype relationships in Kallmann syndrome are complicated. Classical monogenic inheritance does not explain the full range of genetic inheritance of Kallmann syndrome patients. Because of stochastic nature of genetic mutations, exome analyses of Kallmann syndrome patients may provide novel insights.
Keywords
Introduction
Idiopathic hypogonadotropic hypogonadism (IHH) is a congenital condition characterized by a lack of or incomplete pubertal development in individuals 18 years or older because of varying degrees of GnRH secretory defects. 1 Among IHH cases, nearly half of the patients have varying degrees of olfactory abnormalities (olfactory bulb, olfactory tract absence or hypoplasia) and are referred to as Kallmann syndrome (KS). 2 The remainder are termed normal-olfactory IHH (nIHH).
GnRH neurons originate outside the central nervous system (CNS) in the medial-olfactory placode and thereafter migrate along the olfactory epithelium to the olfactory bulb, and thence to the olfactory tract.3,4 Ultimately, the GnRH neurons dissociate from their olfactory guiding fibers to reach the hypothalamus. 5 These complex development defects are tightly regulated by specific spatiotemporal expression patterns of growth factors, adhesion molecules and/or diffusible attractants, and repellents. 6
The etiology of KS is either spontaneous or hereditary. Its incidence is estimated as 1 in 8000 males and 1 in 40,000 females. 7 The majority are sporadic cases, but approximately one-third have a family history and genetic mutations can be detected in about 30% of all KS individuals. 8 The severity of the KS clinical phenotype is heterogeneous in its presentation. Some patients present with atypical KS, such as reversible KS 9 (patients experience a degree of spontaneous pubertal development after intermittent exogenous hormone replacement therapy) or adult-onset KS (patients have normal pubertal development, but KS symptoms occur during adulthood). 10 In addition, particular gene mutations are often associated with specific non-olfactory, non-reproductive system clinical manifestations, such as renal hypoplasia or cleft lip/palate.
The heterogeneous presentations of KS-related hypogonadotropic hypogonadism likely result from varied contributions of the genes.11–21 For example, the same mutation in fibroblast growth factor receptor (FGFR) can cause manifestations of disease, as well as a cohort of patients can undergo reversal of phenotype with the trophic hormone replacement. Even two or more gene–gene interactions can actually manifest the disease process. Furthermore, in the same gene involved, diseases may be manifested with or without olfactory involvement (KS vs. nIHH). In addition, disease heterogeneity is manifested in ethnic groups, like for example, nIHH was first identified in an isolated Bedouin population. 22 Thus, the rationale may be expanded that it merits testing whether the reported genes or its variations can be identified across different ethnic groups across the globe because of the stochastic nature of genotype–phenotype presentations in KS.
KS and nIHH manifestations in an ethnic Han Chinese population have only been scantily reported.23,24 We have our own large case series of independent pedigrees, as well as isolated presentations, in which we examined the reported disease mutations in the current study. To explore these relationships between KS gene mutations and the clinical phenotype, we evaluated the clinical phenotypes in seven KS pedigrees and screened the patients for mutations in the known IHH-related genes. We also screened for the detected mutations in 70 sporadic KS cases and 200 healthy controls.
Materials and methods
Ethical statement
All participating subjects signed an informed consent form, and the research was approved by the Institutional Review Board of Chinese PLA General Hospital. All experiments were performed in accordance with the Declaration of Helsinki.
Patients
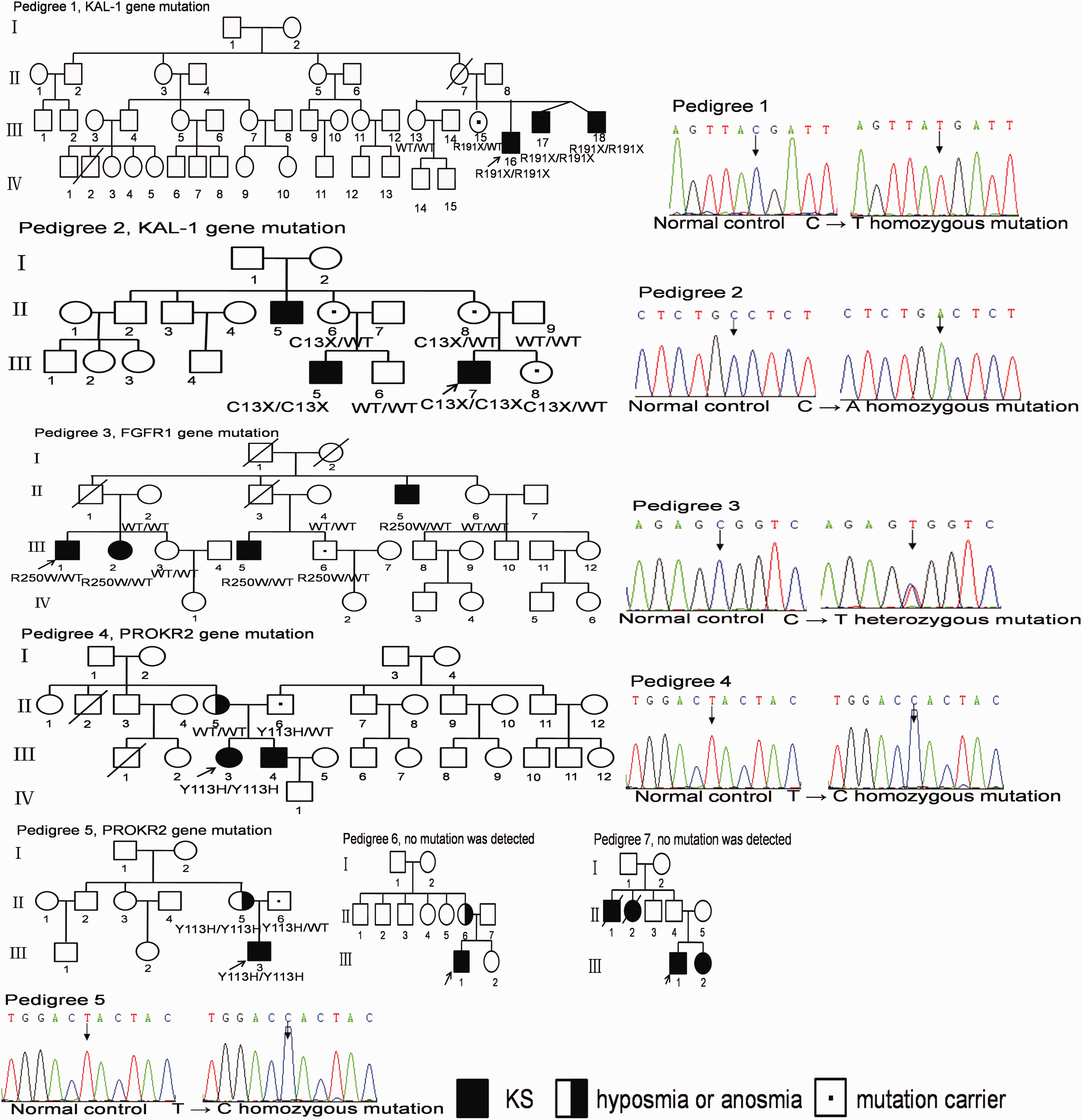
We diagnosed seven Chinese KS pedigrees, containing 19 patients in total, 17 of whom agreed to genetic testing (Figure 1). We also collected 70 sporadic KS patients and 200 Chinese healthy community volunteers (n = 400 chromosomes) for use as the controls. Patient diagnostic criteria were based on the following: (1) age greater 18 years or older with absent or incomplete pubertal development; (2) male serum testosterone (T) ≤ 100 ng/dL, female serum estradiol (E2) ≤ 20 pg/mL; (3) normal adrenal axis and growth hormone axis (assessed by insulin hypoglycemia stimulation test); (4) normal levels of serum thyroid hormone, prolactin, and ferritin; (5) hypothalamic–pituitary region magnetic resonance imaging (MRI) normal; (6) reported features of anosmia or hyposmia. Control populations included subjects with normal sexual development, without any endocrine or systemic diseases.
Genes examined in the current study
Currently, several genes have been causally related to pathogenesis of IHH. The genes that have been examined in KS include: KAL1, FGFR1, FGF8, PROKR2, PROK2, CHD7, NELF, WDR11, HS6ST1, KISS1R, KISS1, TAC3, TACR3, LEPR, LEP, PCSK1, GNRHR, GNRH1, SEMA3A, and NDN7. These genes are essential for the correct differentiation, migration, upstream signal regulation, and action of embryonic GnRH neurons. Among these genes, those that are clearly related to KS are KAL1, FGFR1, PROKR2, PROK2, CHD7, and FGF8. CHD7 has been related to CHARGE syndrome and NELF, 25 HS6ST1, 26 and WDR11 27 may also be related to KS. The observed patterns of inheritance include all three classical Mendelian models: X-linked (OMIM#308700; KAL1); autosomal dominant (OMIM#147950; FGFR1, FGF8, and CHD7), and autosomal recessive (OMIM#244200; PROKR2 and PROK2). In addition, some cases of two gene mutations have been reported, with either both being KS-related genes, (e.g. FGFR1 + PROKR2) or one being a KS-related gene and the other an IHH-related gene (e.g. PROKR2 + GNRHR). 28
Genetic mutational analysis
We obtained 5 mL blood samples from all subjects and extracted genomic DNA using a BloodGen Midi Kit (CWBIO, Beijing, China). Primers were designed to amplify the exons and exon–intron boundaries of 19 of the reported 20 IHH-related genes listed (supplementary Table 1). We did not analyze CHD7. Genomic DNA was PCR amplified in 50 µL reactions using ExTaq polymerase (Takara, Dalian, China) with the following cycling conditions: predenaturation at 95℃ for 5 min; 30 cycles of denaturation at 95℃ for 30 s, annealing at 55–65℃ for 30 s, and extension at 72℃ for 1 min; followed by a final extension at 72℃ for 10 min. The PCR products were directly sequenced in both directions using the original PCR primers with a BigDye v. 3.1 kit on an ABI 3100 (Applied Biosystems, Foster City, CA). Sequences were analyzed using CodonCode Aligner (http://www.codoncode.com/index.htm) and DNASTAR (http://www.dnastar.com/). If a mutation was found, we tested for the presence of this mutation in all other patients and controls, including unaffected family members.
Nonsense changes resulting in either a truncated protein, frameshift insertion, or deletion were categorized as definite mutations. Nucleotide changes that met the following criteria were also classified as disease-causing mutations, vide earlier studies: 29 (1) absent from single-nucleotide polymorphism (dbSNP) (http://www.ncbi.nlm.nih.gov/projects/SNP/) and expressed sequence tag databases; (2) absent in ≥200 ethnically matched subjects; (3) evolutionarily conserved across species; and (4) segregated appropriately in the family.
Clinical assessment of patients
Patients’ clinical characteristics and phenotypic features like appearance, presence of pubic hair, penile length measurement, presence or absence of testicles, development of gynecomastia, and BMI assessment were initially documented. Blood samples were collected for hormone status (FSH, LH, and testosterone) assessment by routine biochemical methods.
Briefly, pubic hair was staged according to the standard Tanner staging (P1–P6). Testicle volume was measured with a Prader orchidometer. Male patients were divided into groups with or without breast development. The reference value of the male’s luteinizing hormone (LH) was 1.5–9.3 IU/L; follicle stimulating hormone (FSH) was 1.4–18.1 IU/L; testosterone was 8.4–28.7 nmol/L. In the GnRH stimulating test, the normal male LH reflection peak was more than five times of the basic value and the peak value is ideally greater than 9 IU/L, which occurred at 30–60 min during the test. In the human chorionic gonadotropin (hCG) stimulating testosterone test, the testosterone peak of normal males occurred at 48–72 h and the peak value increased at least two times of the control. The patients were not treated with any medication before the above examination.
For the olfactory tests, the olfactory sensation scoring scale was: < 1.0, normal; 1.1–2.5, slightly damaged; 2.6–4.0, moderately damaged; 4.1–5.5, heavily damaged; > 5.5, complete loss. Other anomalies examined included non-reproductive system and non-olfactory system symptoms, such as craniofacial malformations (cleft lip/palate, high-arched palate, eye symptoms), deafness, abnormal fingers, tooth agenesis, unilateral renal hypoplasia, nervous system defects (oculomotor nerve abnormalities, movement associated with both hands), sleep disorders, and mood disorders. Patients’ MRI were obtained by standard imaging.
Results
Variants detected in the pedigrees
The clinical assessment of the KS pedigrees

Individuals III-2 in Pedigree 3, II-6 in Pedigree 6, and III-2 in Pedigree 7 were female. The proband in each family was: Pedigree 1: III-17; Pedigree 2: III-7; Pedigree 3: III-1; Pedigree 4: III-3; Pedigree 5: III-3. Y: yes; N: No; ND: not detected; NT: not touched; T: testosterone; * indicates LH peak value in GnRH stimulation test at 30 min; † indicates LH peak value in GnRH stimulation test at 60 min; ‡ indicates FSH peak value in GnRH stimulation test at 60 min; § indicates LH peak value in GnRH stimulation test at 120 min.
The gene analysis of the KS pedigrees
Pedigree 1
There were three male patients in this pedigree (III-17 and III-18 were twins) and their clinical symptoms were severe. They did not have unilateral renal hypoplasia or synkinesia. A homozygous KAL1 nonsense mutation c.C571T (p.R191ter) was detected in the proband (III-17) and both other patients (III-16, III-18). Individual III-15 was a heterozygous carrier of this X-linked mutation and had a normal phenotype. Individual III-13 had no genetic abnormality.
Pedigree 2
There were three male patients in this pedigree. Only the proband had severe clinical symptoms. There was no unilateral renal hypoplasia or synkinesia. A homozygous KAL1 nonsense mutation c.C39A (p.C13ter) was detected in the proband (III-17) and the two other patients (III-5, III-15). Individuals II-6, II-8, and III-8 with a normal phenotype were found to be heterozygous carriers of this X-linked mutation. Individuals II-9 and III-6 were genetically normal. In both pedigrees 1 and 2, the differential mutation involving KAL1 gene produced consistency in the phenotypic manifestation of the hypogonadotropic hypogonadism.
Pedigree 3
There were three male patients and one female patient in this pedigree. The reproductive system, olfactory phenotypes, and olfactory bulb MRI findings of the female were milder than those of the males. There was no cleft lip/palate, abnormal tooth development, or hearing impairment. A heterozygous FGFR1 missense mutation c.C748T (p.R250W) was detected in all three patients. Individual III-6 also carried the mutation but had a normal phenotype. Imperfect penetrance and heterozygosity of the mutational status may potentially explain the milder or apparently normal phenotype in this individual. Individual III-3 was phenotypically and genetically normal.
Pedigree 4
There were one male patient and two female patients in this pedigree. The patients in this family declined clinical examination, but II-5 had self-reported hyposmia. Individual III-4 received sequential therapy with hCG and human menopausal gonadotropin, and after one year he had spontaneous pubertal development. He now has one child and is considered to have reversible KS. Individuals III-1 and III-2 did not have KS symptoms, but both had vision loss since birth. There were no sleep disorders, mood disorders, obesity or other non-reproductive, non-olfactory symptoms. A homozygous PROKR2 missense mutation c.T337C (p.Y113H) was detected in the proband (III-3). Individual II-5 was genetically normal and individual III-4 refused genetic testing. However, individual II-6, with a normal phenotype, was heterozygous for this mutation.
Pedigree 5
There were two patients in this pedigree. The proband had typical KS symptoms; the mother had self-reported dysosmia, and her MRI findings of the olfactory bulb and olfactory groove showed dysplasia. A homozygous PROKR2 missense mutation c.T337C (p.Y113H) was detected in both. Individual II-6, with a normal phenotype, was a heterozygous carrier of this mutation. In the current study, we have not examined the genotypic contributions of the parents of the individual II-6, but propose to pursue this test in future studies.
Pedigree 6
There were two patients in this pedigree. The proband had classical KS symptoms; the proband’s mother had a severe olfactory deficit based on a test of olfaction, and MRI demonstrated that the olfactory bulb and olfactory tract were not developed. Interestingly, no mutation was found in the proband in any of the 19 genes tested.
Pedigree 7
The proband (III-1) and his sister (III-2) had typical KS symptoms. Individuals II-1 and II-2 had perished; they had no diagnosis, but the other family members reported that they had abnormal olfaction, were unmarried, and had no children. No mutation was found in the proband in any of the 19 genes tested.
In addition to the above-mentioned mutations, we also detected several SNPs that were present in dbSNP. SNPs in LEPR and KISS1 (LEPR: c.A326G [p.K109R; rs1137100] and c.A668G [p.Q223R; r1137101]; KISS1: c.C242G [p.P81R; rs4889]) were found in all seven pedigrees; an SNP in PCSK1 (c.G239A [p.R80Q; rs1799904]) was found in pedigrees 1 and 3. According to SIFT (http://sift.jcvi.org/), all four of these SNPs are potentially harmful. However, the three LEPR/KISS1 SNPs were present in the 1000 genomes database (http://browser.1000genomes.org/index.html) and can therefore be considered non-pathogenic.
Mutations detected in the sporadic KS patients and controls
We screened for the four mutations described above in the 70 sporadic KS patients with no reported or documented antecedent family history and 200 control subjects. The only gene identified in sporadic KS was PROKR2 p.Y113H, for which we detected one homozygous mutation and one heterozygous mutation. Among the normal controls, we detected a heterozygous PROKR2 p.Y113H variant in 2/200 of the current case series. The PCSK1 SNP p.R80Q was detected in 8/200 normal control subjects, which suggests that this is likely non-pathogenic, despite its absence in the 1000 genomes database.
Discussion
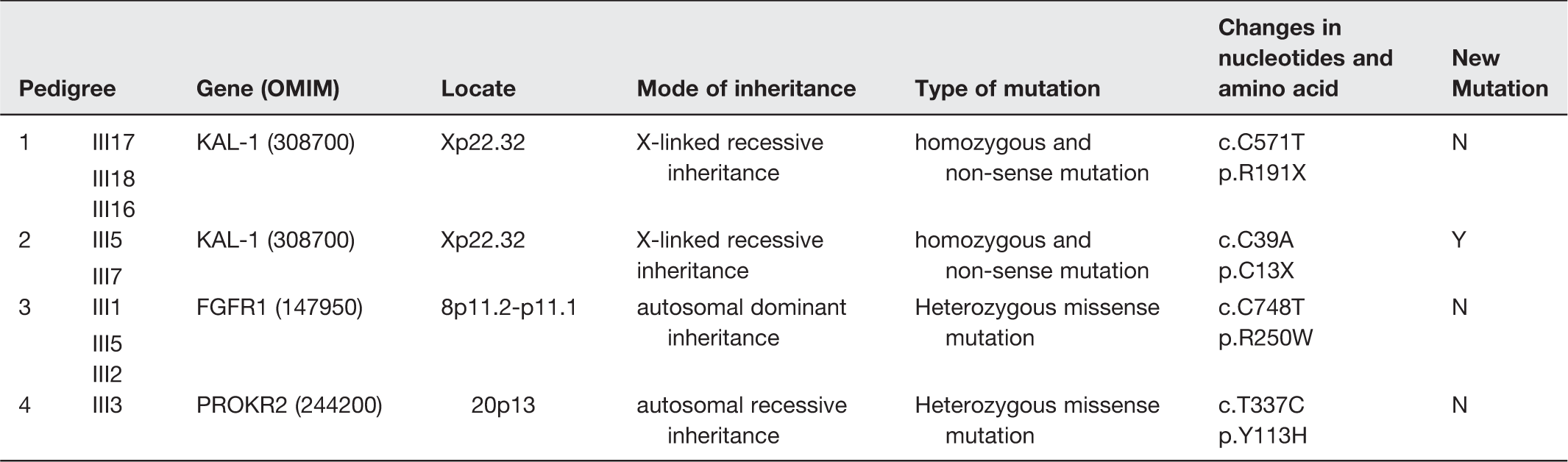
It has been previously reported that genetic abnormalities can be detected in about 30% of KS patients. 8 Here, we detected a genetic mutation of the currently reported repertoire of involved genes in five of the seven pedigrees in this study. For individual genes, it has been reported that KAL1, FGFR1, and PROK2/PROKR2 mutations account for 10–14%, 30 10%, 31 and 9% 32 of all KS, respectively.
The present study is a report of the largest cohort of Han Chinese ethnic patients (an ethnic group that comprises more than 90% of the population of mainland China) with inherited KS. Interestingly, of the four mutations detected in the pedigrees, only the PROKR2 p.Y113H mutation was detected among the 70 sporadic KS patients (1.4% heterozygous and 1.4% homozygous), and this was also the only mutation detected among the normal controls (1% heterozygous). The sporadic cases and normal controls were screened only for the four mutations detected rather than for the 19 candidate genes; so it would be erroneous to assume that the mutation rate in our KS pedigrees is higher than that in the sporadic cases.
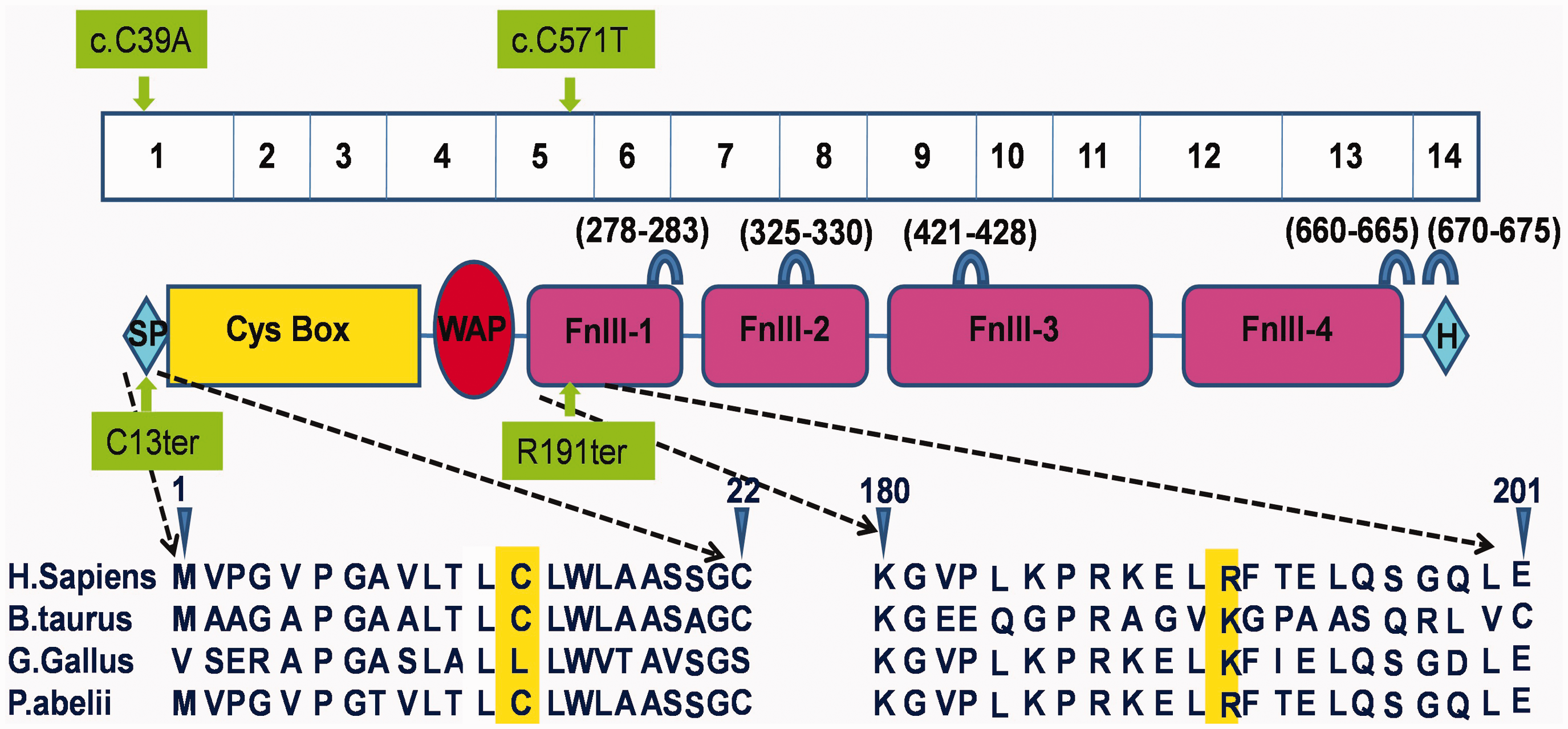
All our detected mutations have been described before,8,33,34 except KAL1 p.C13ter (pedigree 2). KAL1 encodes the extracellular matrix protein KS 1 sequence (also known as anosmin-1) (Figure 2). In this protein, the Whey acidic protein (WAP) domain and the first fibronectin III domain are conserved across species and interact directly with FGFR1.
35
Furthermore, anosmin-1 has five heparan sulfate (HS)-binding sites with very strong HS-recruiting capability, which are essential for its function. We speculate that in pedigree 2, the anosmin-1 protein would be truncated after the nonsense mutation, resulting in the absence of all these functional domains.
There are several possible reasons why we did not detect a mutation in pedigrees 6 or 7: the patients may have had an intronic mutation or intron copy number variation. 36 It is also likely that the mutation in these families is in a novel KS-related gene. Epigenetic variation such as gene methylation might also play a pathogenic role. Further analysis of these patients in these pedigrees is necessary.
The genotype–phenotype relationships among the different mutations are not fully understood. For instance, patients in pedigrees 1 and 2, carrying a KAL1 mutation, had severe symptoms, but pedigree 3 patients with an FGFR1 mutation had variable clinical symptoms. The FGFR1 gene, encoding the fibroblast growth factor receptor 1 protein, is a member of the receptor tyrosine kinase superfamily. 8 The combination of fibroblast growth factor (FGF) with FGFR1 results in receptor dimerization, tyrosine kinase domain phosphorylation, and the activation of a downstream pathway. HS strengthens the weak FGFR–FGF interaction, 37 and the HS is recruited by anosmin-1. 38 The resulting anosmin-1/FGFR1/FGF/HS complex can activate the FGFR1 pathway.33,39 This pathway would be ablated in pedigrees 1 and 2, who carry a truncating anosmin-1 mutation. This could explain the relatively severe symptoms.
In contrast, the FGFR1 missense mutation in pedigree 3 might affect the binding of ligand and anosmin-1 to some extent, but would not completely ablate the pathway, thus resulting in relatively mild symptoms. The milder symptoms in the female patients compared with the male patients in this family may be explained by an increased dosage of the X-chromosome-encoded anosmin-1. 29 The incomplete penetrance observed for individual III-6 in this family might be an example of maturity-onset KS and could be explained by delayed dominance (a phenomenon in which a dominant virulence gene in the heterozygous state only exerts its function after a certain age is reached); follow-up studies of this individual could confirm this.
The pathogenicity of the PROKR2 p.Y113H heterozygous mutation is questionable. We detected this mutation heterozygously in 1% of our normal controls. Moreover, mice with gene knockout of Prokr2 show a phenotype only in the homozygous state. 40 However, one sporadic patient had a heterozygous mutation, and this phenomenon has been reported previously. 41 Patient III-4 in pedigree 4 was a reversible KS case and patients III-3 and III-4 responded well to treatment. Patient II-5 showed a phenotype but was genetically normal. It is possible that this individual carried a microdeletion that spans the mutation site and would not have been detected by our methods. We deduced that individual III-3 inherited one chromosome with a microdeletion (from II-5) and one with a mutation (from II-6; because II-6, with a normal phenotype, was a carrier of this mutation), so her genotype was detected as homozygous. Quantitative PCR is needed to confirm this speculation.
We detected four SNPs in LEPR, KISS1, and PCSK1. All were present in the 1000 Genomes database or were found among normal controls and are thus probably non-pathogenic. However, functional tests are needed to exclude their involvement.
It is clear that the genetic origins of KS/IHH are complex, and it is widely accepted that the most reasonable explanation for the various genotype–phenotype relationships is digenic or oligogenic inheritance, 42 together with complex interactions between the gene products. 29 Moreover, these interactions may be affected by environmental factors, modifier genes, and epigenetics. 43 In this study, we examined all genes (except CHD7) currently related to KS and nIHH but digenic or oligogenic inheritance mode was not found. Recent studies have shown that CHD7 mutation is only present in KS patients with CHARGE syndrome phenotype characteristics. If the patient was diagnosed with hypogonadism and anosmia, attention merits to be paid to the presence of clinical characteristics of CHARGE syndrome. 12 CHD7 needs to be detected only in case of deafness, ear morphological abnormalities, and/or hypoplasia or aplasia of the semicircular canals of the inner ear. Our patients had no clinical characteristics of CHARGE syndrome after evaluation, so CHD7 was not examined. Therefore, we speculate that the other genetic factors involved in these pedigrees and sporadic cases are as-yet-undiscovered KS/nIHH-related genes or mutations within introns of the 19 known genes.
It has been reported that particular genetic mutations are often associated with a specific non-reproductive system, non-olfactory phenotype; for example, KAL1 mutation is associated with unilateral renal hypoplasia and synkinesia; 37 FGFR1 mutation is often accompanied by cleft lip/palate, 44 dental hypoplasia, 44 and abnormal hearing; PROKR2 mutation is often associated with mood disorders, 45 obesity, 46 and abnormal circadian endocrine rhythms. 47 However, we did not find any of these abnormalities in our seven pedigrees by questioning, renal ultrasound, and pure-tone audiometry examination of some patients. It is possible that these associations vary by ethnicity and are uncommon in the Han Chinese population.
In conclusion, we detected diverse genetic mutations in five of our seven pedigrees, including the novel nonsense mutation KAL1 p.C13ter. There were complicated genotype–phenotype relationships in the families, which might be due to skewed X-inactivation, delayed dominance, and microdeletion. Moreover, some relationships could not be explained by monogenic inheritance, and they might result from multiple or novel KS/nIHH gene inheritance, with modification by environmental and/or epigenetic factors. Gene-chip exome mutation analysis should be used to identify new candidate genes that may be important in pedigrees 6 and 7 in which we found no mutations. In addition, further genetic (intron analysis), gene methylation, and other epigenetic analyses, and then in vitro and in vivo functional studies, are needed.
Footnotes
Authors’ contributions
YMM participated in the design of the study, performed data analysis, and helped to revise the manuscript. YQW carried out Data collection, Genetic mutational analysis, performed data analysis and Statistical analysis, and helped to draft the manuscript. WJG, ZHL, JTD performed Genetic mutational analysis and data analysis. JMB, XLW, APW, TPH, DLZ, JKY, JD, LJY, GQY, NJ, KC, LZ, GN carried out Data collection, Genetic mutational analysis. All authors read and approved the final manuscript.
Acknowledgements
We thank all the patients and their family members for their participation in this study. We also thank the staff of the Peking University Third Hospital, Nanjing Gulou Hospital, Shanghai Ruijin Hospital, and Beijing Tongren Hospital for providing some of the KS pedigrees and the sporadic KS cases. Gene amplification and sequence analysis were conducted at the Joy Orient Translational Medicine Research Center Co. Ltd. Beijing, China. This study is supported by Construction of Technical Platform for New Drug Development (fund number: 2011ZX09307-001-08).
Conflict of interest
None declared.