
Abstract
Advances in depression research have targeted inflammation and oxidative stress to develop novel types of treatment. The JAK/STAT signaling pathway plays pivotal roles in immune and inflammatory responses. The present study was designed to investigate the effects of N-acetylcysteine, a putative precursor of the antioxidant glutathione, in an animal model of depression, with an emphasis on the JAK/STAT signaling pathway. Fluoxetine, a classical antidepressant drug was also under investigation. Male Wistar rats were subjected to forced swimming test and given N-acetylcysteine and fluoxetine immediately after the pre-test session, 5 h later and 1 h before the test session of the forced swimming test. N-acetylcysteine decreased immobility time (P < 0.05), serum corticosterone (P < 0.001), and hydrogen peroxide (P < 0.001), while restored glutathione concentration. Treatment of the rats with N-acetylcysteine produced significant (P < 0.001) down-regulation of STAT3 mRNA expression and protein phosphorylation. On the other hand, N-acetylcysteine significantly (P < 0.001) increased SOCS3 gene expression; however, SOCS3 protein was not changed. In conclusion, our study suggests that modulation of the JAK/STAT pathway might mediate the antidepressant-like effects of N-acetylcysteine. Therefore, depression research may target the JAK/STAT signaling pathway to provide a novel effective therapy.
Introduction
Major depressive disorder (MDD) is a common mental disease and one of the leading causes of disability worldwide. 1 The pathogenesis of depression is thought to occur according to the monoamine hypothesis, which suggests that there is a deficiency or imbalance in the monoamine neurotransmitters, including serotonin, dopamine, and norepinephrine. 2 In addition, alterations in neurogenesis and neuroendocrine functions could play roles in the pathogenesis of MDD.3,4 Recent developments in research investigating major depression have led to the hypothesis that oxidative stress and inflammatory processes are also involved in the pathogenesis of major depression and may contribute to dysfunction of the serotonergic and noradrenergic systems.5,6
Oxidative stress has been reported to play key pathologic roles in diverse disease states and may be a common mechanism underlying several major psychiatric disorders. 7 The brain is more vulnerable to the detrimental effects of reactive oxygen species (ROS) due to its high metabolic rate and low antioxidant levels, which could explain why oxidative stress is a central feature in most neurodegenerative diseases, including major depression.6,8 Multiple studies have demonstrated that depression is associated with increased levels of the redox products malondialdehyde, 8-iso-prostaglandin F2 9–12 and 8-hydroxy-2′-deoxyguanosine. 9 Overproduction of ROS generates an inflammatory response and increases the release of proinflammatory cytokines. In addition, a large body of evidence has shown that major depression is characterized by disturbances in inflammatory pathways, including the expression of cytokines such as interleukin-1β (IL-1β), IL-6, interferon-γ (IFN-γ), and tumor necrosis factor-α (TNF-α). 13 Therefore, antioxidants might represent efficient therapies with beneficial effects in treating depression.
The Janus kinase (JAK)/signal transducer and activator of transcription (STAT) signaling pathway is associated with inflammation, and its components are widely expressed in several areas of the brain such as the cerebral cortex and hippocampus.14,15 This pathway plays important roles in cell growth, survival, development, and differentiation and in the regulation of gene expression. 16 Dysregulation of the JAK/STAT pathway has been found to be a key factor in a variety of neurodegenerative diseases, which highlights the importance of determining how this pathway influences the fate and functions of brain cells. 17 The JAK/STAT signaling pathway acts downstream of the binding of various ligands, such as cytokines, growth factors, and ROS 18 and is inhibited by the action of suppressor of cytokine signaling (SOCS) proteins. 19 The observation that ROS such as hydrogen peroxide (H2O2) activate certain components of the JAK/STAT pathway (STAT1, STAT3 and JAK2) indicates that increases in ROS associated with depression could result in activation of the JAK/STAT pathway and that this effect might be inhibited by antioxidants such as glutathione (GSH), which is the most important low molecular weight antioxidant that is synthesized in cells.20–22
Accordingly, several clinical studies have targeted inflammation and oxidative stress to develop novel types of treatments for MDD to overcome the treatment resistance associated with most antidepressants. It has been demonstrated that patients who suffer from depression have significantly decreased concentrations of GSH in their blood. 23 Therefore, patients with major depression could benefit from GSH treatment. However, oral administration of GSH alone does not adequately restore GSH levels because this compound is rapidly hydrolyzed in the liver and intestines and because its ability to cross the blood–brain barrier is poor. 24 In contrast, the administration of N-acetylcysteine (NAC) results in a significant increase in the plasma level of cysteine, leading to an increase in the plasma level of GSH, which can protect against ROS.25,26 NAC has been examined in the context of diverse neuropsychiatric disorders, and add-on treatment with NAC was shown to exert significant effect on depressive symptoms by up-regulating the GSH pathway in a placebo-controlled randomized trial. 27 In addition, NAC showed antidepressant-like activity in several preclinical models, including the rat forced swimming test (FST), 28 mouse tail suspension test (TST), 29 and the FST in bulbectomized rats. 30 Unlike GSH, NAC has been shown to successfully penetrate the blood–brain barrier.31,32
NAC acts through different mechanisms to mediate its antidepressant-like effects, and it is likely that NAC exerts these effects through mechanisms beyond those of being a precursor of the antioxidant GSH, which modulates the glutamatergic, 33 dopaminergic, 34 and inflammatory pathways. 35 Therefore, the current study was designed to investigate the potential involvement of the JAK/STAT signaling pathway in the pathogenesis of depression and the modulatory effect of NAC.
Materials and methods
Experimental animals
Adult male Wistar rats weighing 150–175 g were supplied by the Animal Care Centre at the College of Pharmacy, King Saud University, Riyadh, Saudi Arabia. The rats were housed in standard polypropylene cages in controlled environment (25℃ ± 1℃) on a 12 h light/dark cycle and were provided free access to food and water. Rats were kept under observation for 1 week to acclimatize to the laboratory conditions prior to the experiment. All animal procedures were undertaken with the approval of Institutional Research Ethics Committee of the College of Pharmacy at King Saud University (Riyadh, Saudi Arabia).
Drugs and chemicals
NAC and fluoxetine were purchased from Sigma-Aldrich (St. Louis, MO, USA). H2O2 Amplex® Red assay kit was supplied by Invitrogen (USA) and corticosterone assay kit was purchased from MyBioSource (San Diego, CA, USA). All other chemicals were of analytical grade and obtained from either Sigma-Aldrich (St. Louis, MO, USA) or Cell Signaling Technology (Beverly, MA, USA).
Treatments and behavioral tests
The FST was performed as described by Porsolt et al.36,37 with slight modification. Rats were individually forced to swim in an open tank (25 × 30 cm, height 40 cm) filled with water at 24 ± 1℃ (the animals could not touch the bottom). On the first experimental day, the animals were gently placed in water for 15 min (pre-test session). The next day, they were once more placed gently in the water and observed for 5 min (test session). Immobility time of the rats was recorded in each 5 s during the test session. A rat was considered immobile when no additional activity (floating with only occasional alternate movements of paws and tail necessary to keep its head above the water) is observed. After each swim session, the rat was removed from the cylinder, dried with paper towels, placed in a resting cage for 20 min, and returned to its home cage. Water in the cylinder was changed between subjects.38,39
Forty-eight rats were included in the present study and were randomly allocated into four groups having 12 in each as follows:
NAC and fluoxetine were dissolved in sterile 0.9% NaCl. Drugs and vehicles were administered immediately after the pre-test session, 5 h later and 1 h before the test session of the FST.38,39
Open field test
To rule out any interference of locomotor activity in interpretation of the results obtained for the FST, ambulatory behavior was assessed in an open field test (OFT). 41 Briefly, the apparatus consists of a large open field (70 cm × 70 cm × 40 cm) divided into 16 equal squares drawn on the floor of the arena. The test room was dimly illuminated with a red bulb (25 W) located 130 cm above the center of the arena. A single rat was placed in the center of the floor and the number of squares crossed (with the four paws), the number of rears (posture sustained with hind-paws on the floor), and the time spent grooming (including washing or mouthing of forelimbs, hind paws, face, body and genitals) were counted for 5 min. The open field was cleaned with 90% ethanol after each rat was tested to prevent the possible cueing effects of odors left by the previous subject. 42 This test was performed 1 h after the last dose of drug was administered and before the test session of the FST (approximately 1 h between tests).
Blood and brain samples collection
Animals were sacrificed by decapitation following exposure to behavioral tests. Since anesthetic drugs affect the levels of corticosterone, 43 decapitation was done without anesthesia. Trunk blood was collected from the site, where the animal was decapitated in anticoagulant-free tubes. Blood samples were centrifuged at 1000 × g for 15 min to separate serum. Sera were then collected and kept at −20℃ for further analysis.
The whole brain was quickly removed, rinsed with cold saline, and dissected on an ice-cold plate. The prefrontal cortex and hippocampus were harvested, washed, weighed, frozen in liquid nitrogen and stored at −80℃. Frozen samples (10% w/v) were homogenized in chilled phosphate-buffered saline (PBS) using tissue homogenizer (Omni International Inc., Kennesaw, GA, USA), and the homogenates were centrifuged at 1500 × g for 10 min. The clear homogenates were collected and stored at −20℃ for subsequent assays. Other samples were kept frozen at −80℃ and used for western blotting and gene expression assays.
Biochemical assays
Measurement of H2O2 content
The H2O2 content in the tissue homogenates of the hippocampus and prefrontal cortex was determined using the Amplex Red® assay kit according to the manufacturer’s instructions. This assay is based on the reaction between H2O2 and N-acetyl-3,7-dihydroxyphenoxazine (Amplex Red), which is a colorless and non-fluorescent derivative of dihydroresorufin, in a 1:1 stoichiometry to produce the red-fluorescent oxidation product, resorufin. 44 Resorufin has excitation and emission maxima of approximately 571 nm and 585 nm, respectively. This reaction can be used to detect as little as 10 picomoles of H2O2.
Measurement of GSH content
GSH content was determined based on Ellman’s method. 45 Briefly, tissue homogenates were deprotonated with trichloroacetic acid (TCA) and incubated at room temperature for 5 min followed by centrifugation at 10,000 × g for 10 min at 4℃. The clear supernatant was then allowed to react with 6 mM 5,5′-dithiobis-(2-nitrobenzoic acid) (DTNB) reagent dissolved in absolute methanol. The volume was completed to 3 ml with 0.2 M Na2HPO4. The absorbance was measured at 412 nm against a blank reagent using Ultrospec 2100 Pro (Amersham, GE Healthcare Life Science, Piscataway, NJ, USA). Different concentrations of standard GSH were diluted for standard curve preparation. The concentration of GSH was expressed in nmol/100 mg tissue.
Measurement of serum corticosterone level
Serum levels of corticosterone were assayed using specific enzyme-linked immunosorbent assay kit (MyBioSource, San Diego, CA, USA) following the manufacturer’s instructions. The assay sample and buffer were incubated together with corticosterone-horseradish peroxidase (HRP) conjugate in pre-coated plate for 1 h. The wells were decanted, washed five times, and incubated with a substrate for HRP enzyme. A stop solution was added and intensity of the formed yellow color was measured spectrophotometrically at 450 nm in a microplate reader. A standard curve was plotted and coricosterone concentration in each sample was interpolated from this standard curve.
RNA isolation and quantitative polymerase chain reaction
Total RNA was isolated from approximately 50 mg of frozen hippocampus and prefrontal cortex using RiboPure™ RNA purification kit (Life Technologies, Eugene, Oregon, USA), and concentrations were quantified at 260 nm using NanoDrop 1000 spectrophotometer (Thermo Scientific, Wilmington, DE, USA). RNA samples with A260/A280 ratios ≥ 1.7 were selected. The RNA integrity was assessed using 1% agarose gel electrophoresis. Reverse transcription of RNA to complementary DNA (cDNA) was performed with 2 µg RNA using GeneAmp® RNA PCR core kit (Life Technologies, Eugene, Oregon, USA), in Applied Biosystems® 2720 thermal cycler.
Primer pairs used for qPCR
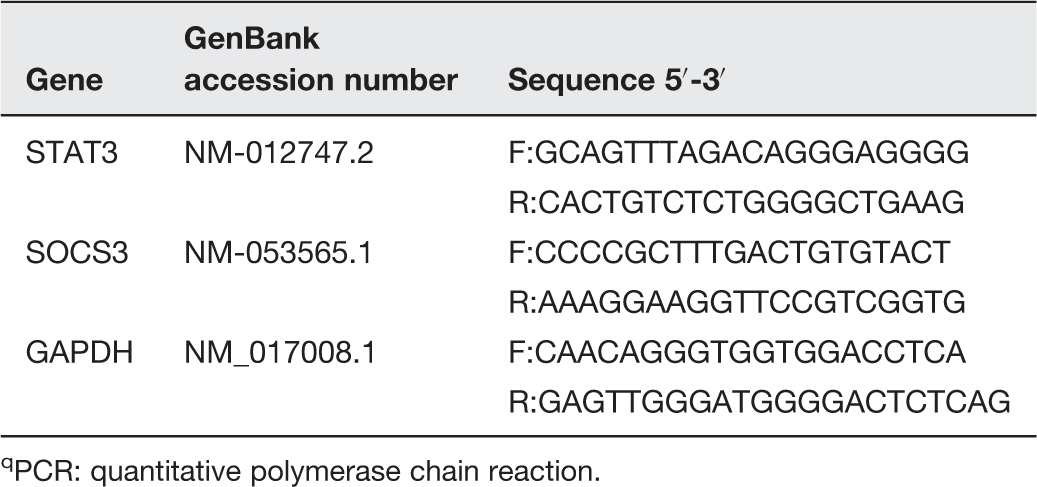
PCR: quantitative polymerase chain reaction.
Western blotting
Protein expression of SOCS3, STAT3, and GAPDH was determined in hippocampus and prefrontal cortex tissue homogenates using Western blotting analysis. Briefly, 100 mg of tissue was homogenized in radioimmunoprecipitation assay (RIPA) buffer supplemented with proteinase inhibitors. The homogenate sample was then centrifuged at 10,000 × g for 15 min at 4℃. Protein concentration in the supernatant was determined using Bradford protein assay. 47 Samples were mixed with Laemmli loading buffer, denatured at 99℃ for 5 min and separated by 12% SDS-PAGE. The separated proteins were transferred overnight at 4℃ to polyvinylidene difluoride (PVDF) membranes using wet transfer system (Bio Rad Trans-Blot® Cell, Hercules, CA, USA). Membranes were blocked using 5% (w/v) non-fat dry milk in tris buffered saline – tween 20 (TBST) for 1 h at room temperature. Membranes were incubated overnight at 4℃ with rabbit polyclonal anti-p-STAT3 (Tyr-705) diluted 1:1000, anti-STAT3 (1:1000 dilution), anti-SOCS3 (1:2000 dilution), and anti-GAPDH (1:1000 dilution). Membranes were then washed three times with TBST and incubated with HRP-conjugated anti-rabbit secondary antibody (1:2000 dilution) for 1 h at room temperature. All antibodies were supplied by Cell Signaling Technology (Beverly, MA, USA) and diluted in TBST. After washing, protein bands were visualized by Super Signal West Pico Chemiluminescent Substrate (Pierce, Rockford, IL, USA) using the ImageQuant™ LAS 4000 mini biomolecular imager (GE Healthcare Life Science, Piscataway, NJ, USA) and quantified using ImageJ software (NIH, USA). The pSTAT3 was normalized to STAT3 and GAPDH, while the SOCS3 protein level was determined after normalization to the GAPDH and presented as % of control.
Statistical analysis
Statistical analysis was performed using GraphPad Prism (GraphPad Software, San Diego, CA, USA), and all statistical comparisons were made by means of the one-way analysis of variance test followed by Tukey’s test post hoc analysis. Results were expressed as mean ± standard error of the mean (SEM) and a P-value < 0.05 was considered significant.
Results
Effect of NAC and fluoxetine on immobility time and locomotor activity
Data represented in Figure 1(a) show the effect of NAC and fluoxetine on immobility time in rats subjected to FST. Treatment with NAC significantly (P < 0.05) decreased immobility time when compared with the depressed control rats; the recorded data were 91.67 ± 23.01 and 190.8 ± 31.61 s for NAC and depressed group, respectively. Rats treated with fluoxetine showed a significant (P < 0.01) decrease in immobility time (44.17 ± 10.60 s) when compared with the corresponding control group. On the other hand, the effect of either NAC or fluoxetine on the locomotor activity measured in the OFT was non-significant (P > 0.05) when compared with the control rats (Figure 1(b)).
Effect of NAC and fluoxetine on (a) the immobility time in FST and (b) the number of crossings in OFT. Data are mean ± SEM (N = 12).*P < 0.05 and **P < 0.01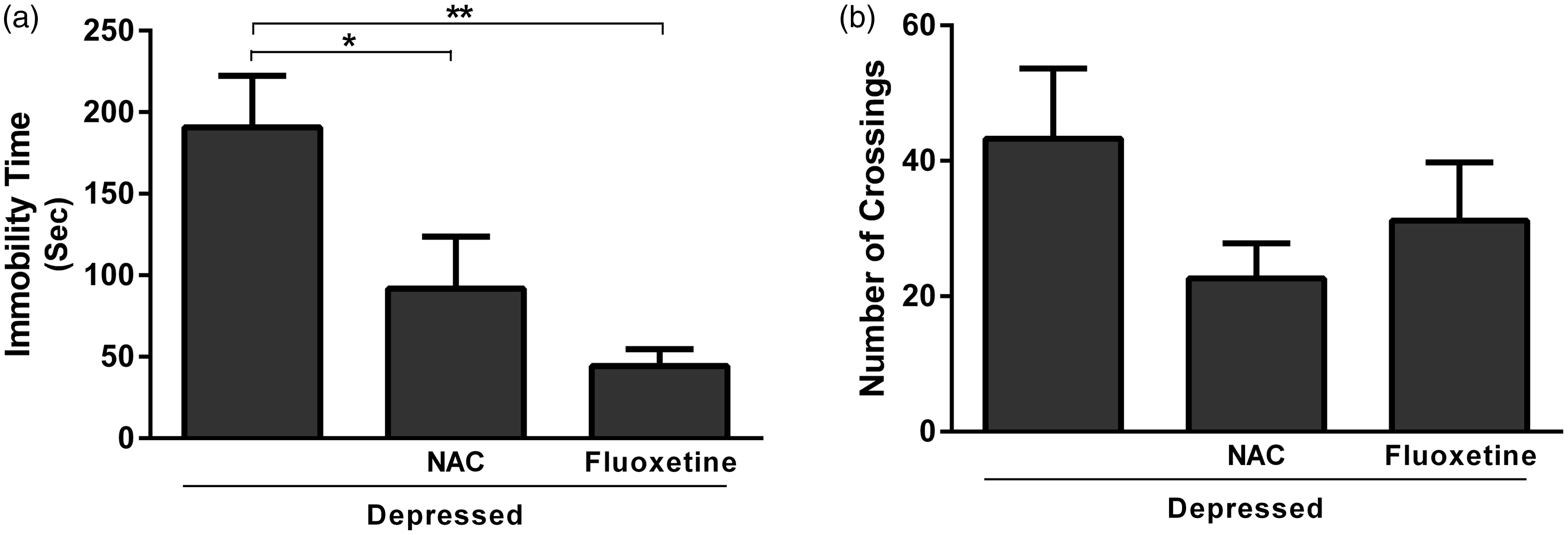
Effect of NAC and fluoxetine on H2O2 and GSH content
Rats exposed to FST showed a significant (P < 0.001) increase in hippocampus and prefrontal cortex H2O2 concentration (61.99 ± 1.30 nmol/100 mg) when compared with the normal control rats (48.01 ± 0.52 nmol/100 mg), as represented in Figure 2(a). The observed increase was significantly (P < 0.001) reversed after treatment with either NAC or fluoxetine; the recorded results were 36.58 ± 2.11 and 42.60 ± 1.02 nmol/100 mg, respectively.
Effect of NAC and fluoxetine on (a) hydrogen peroxide and (b) reduced glutathione in rats subjected to the FST. Data are mean ± SEM (N = 12).*P < 0.05, **P < 0.01 and ***P < 0.001.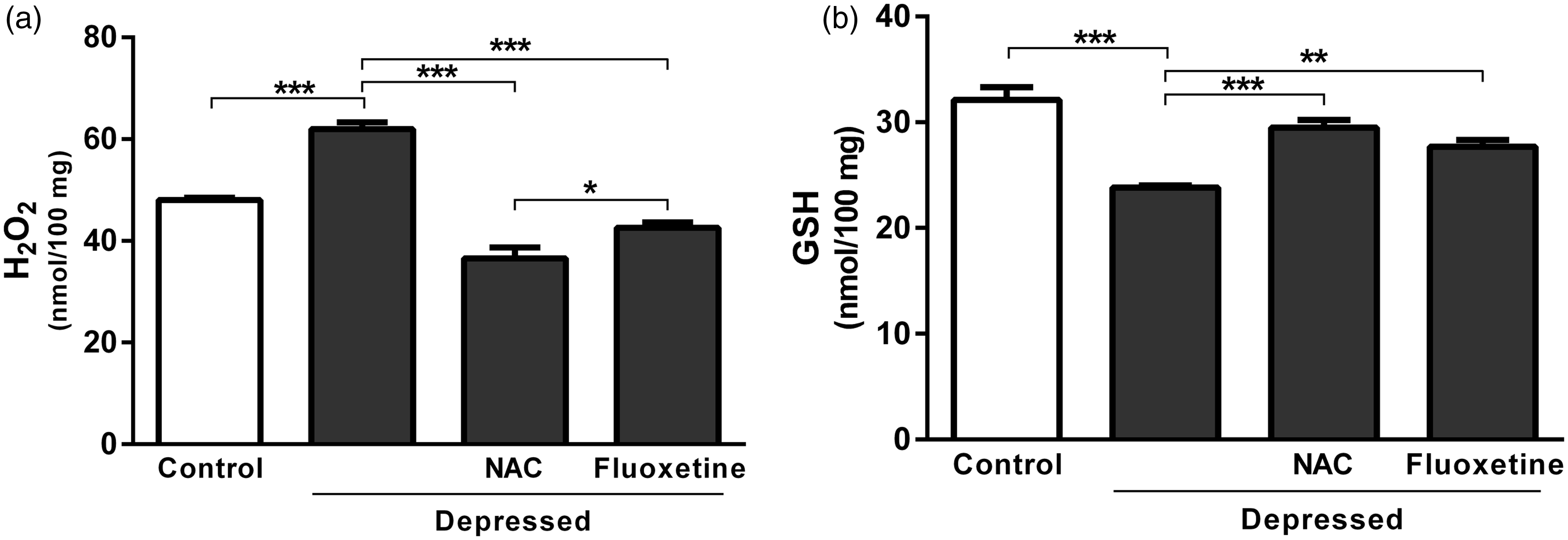
On the contrary, GSH content of FST-exposed rats was significantly (P < 0.001) declined when compared with the corresponding control rats. Treatment with NAC significantly (P < 0.001) restored GSH levels, as represented in Figure 2(b). Similarly, FST-induced decrease in GSH was markedly (P < 0.01) alleviated following treatment with fluoxetine. NAC was more potent than fluoxetine in ameliorating GSH levels in the brain of rats exposed to FST (Figure 2(b)).
Effect of NAC and fluoxetine on serum corticosterone levels
Data concerning the effect of acute treatment with either NAC or fluoxetine on serum levels of corticosterone are represented in Figure 3. The results revealed significant (P < 0.01) increase in serum corticosterone after exposure to FST. The recorded results were 98.17 ± 2.65 and 64.20 ± 8.95 ng/ml for the depressed and control rats, respectively. On the other hand, NAC-treated rats showed significantly (P < 0.001) decreased serum corticosterone level (68.07 ± 3.35 ng/ml) when compared with the non-treated depressed rats. Treatment of the FST-exposed rats with fluoxetine produced non-significant (P > 0.05) change in serum corticosterone level (Figure 3).
Effect of NAC and fluoxetine on serum corticosterone levels in rats subjected to the FST. Data are mean ± SEM (N = 12). **P < 0.01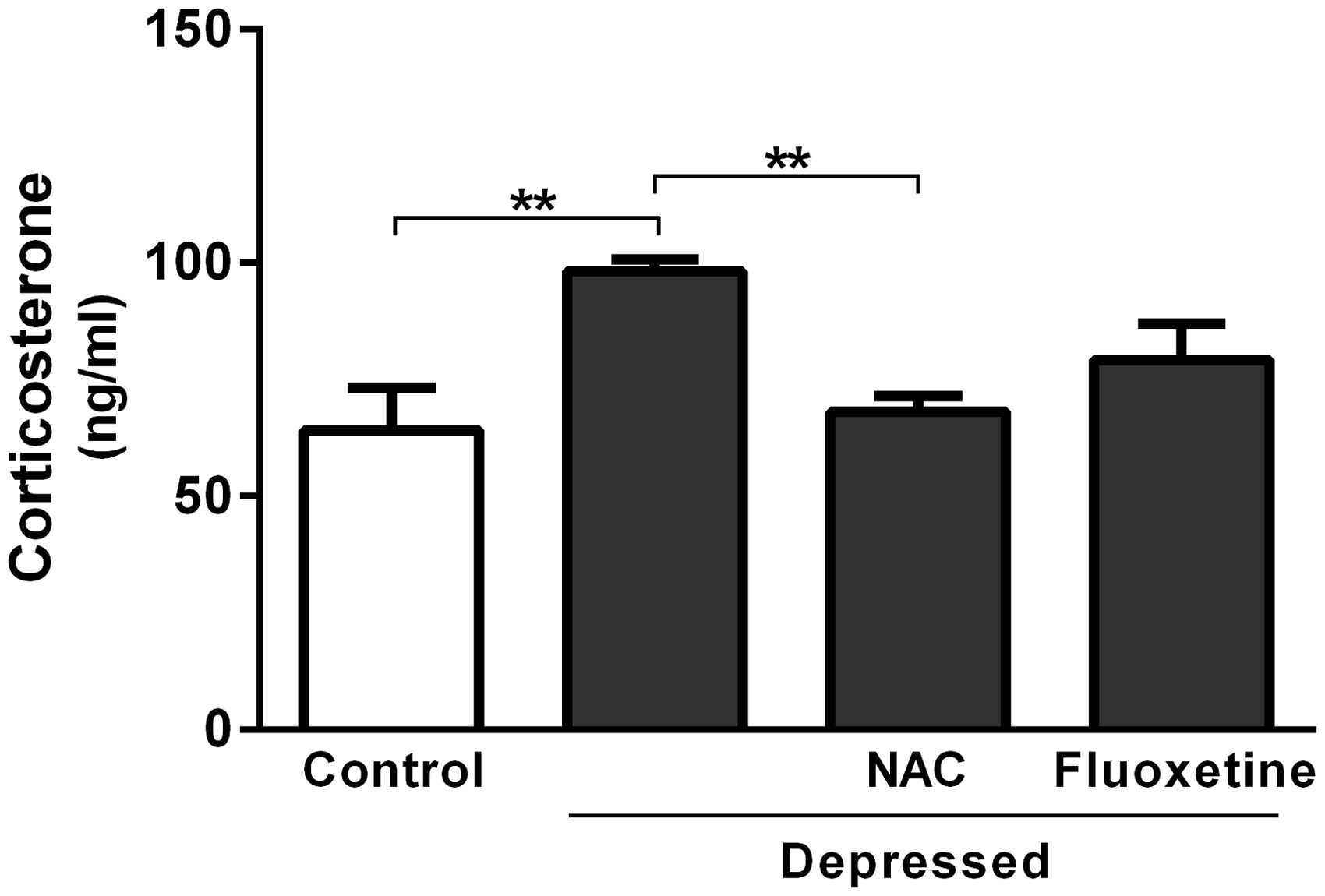
Effect of NAC and fluoxetine on SOCS3 and STAT3 gene expression
Gene expression levels were assessed in the hippocampus and prefrontal cortex of control and experimental rats using qRT-PCR (Figure 4). SOCS3 mRNA expression revealed non-significant (P > 0.05) difference in depressed rats when compared with the control group. Treatment of the depressed rats with NAC produced significant (P < 0.001) up-regulation of SOCS3 mRNA expression as depicted in Figure 4(a). Also, fluoxetine significantly (P < 0.01) up-regulated SOCS3 in the hippocampus and prefrontal cortex of rats exposed to FST.
Effect of NAC and fluoxetine on mRNA expression of (a) SOCS3 and (b) STAT3 in rats subjected to the FST. Data are mean ± SEM (N = 12). **P < 0.01 and ***P < 0.001.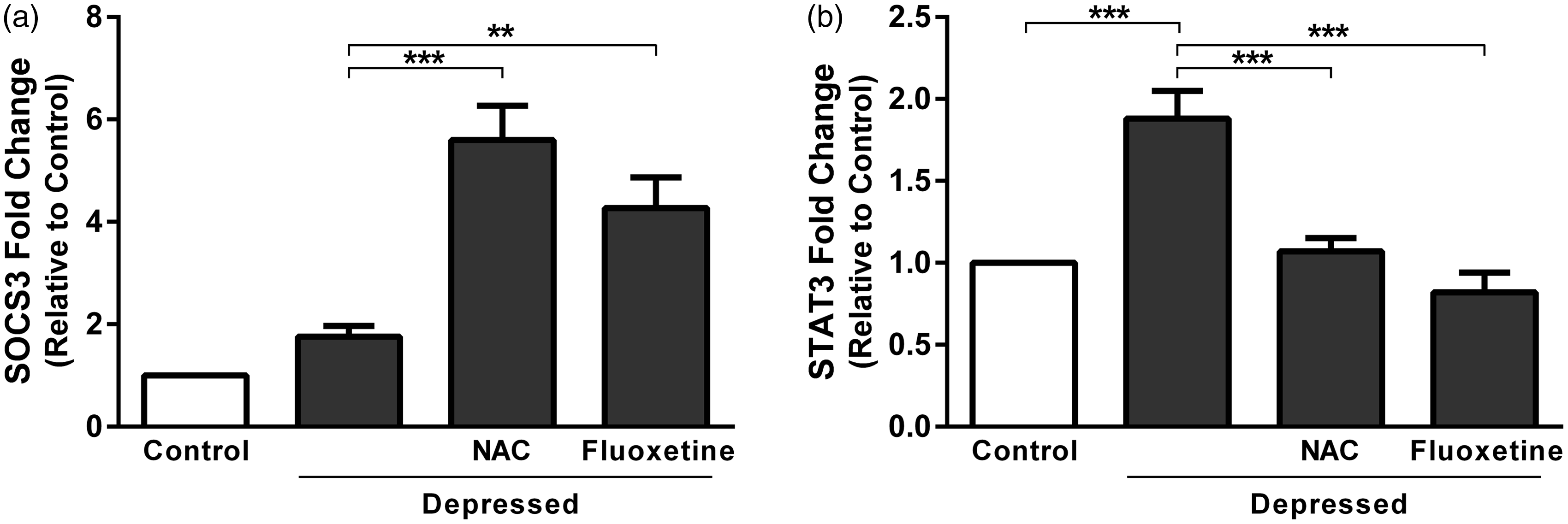
STAT3 was significantly (P < 0.001) up-regulated in hippocampus and prefrontal cortex of the depressed rats as compared to the corresponding control rats (Figure 4(b)). Treatment of the depressed rats with either NAC or fluoxetine produced significant (P < 0.001) decrease in STAT3 expression.
Effect of NAC and fluoxetine on SOCS3 protein expression and STAT3 phosphorylation
FST did not significantly affect SOCS3 protein expression levels when compared with non-depressed control rats as represented in Figure 5(a). Unlike gene expression data, treatment of the FST-exposed rats with either NAC or fluoxetine produced non-significant (P > 0.05) effect on SOC3 protein expression. On the other hand, FST-exposed rats exhibited significant (P < 0.01) increase in STAT3 phosphorylation when compared with the corresponding control rats (Figure 5(b)). Treatment with NAC significantly (P < 0.001) decreased STAT3 phosphorylation. In addition, fluoxetine supplementation produced a marked (P < 0.01) decline in phosphorylation of STAT3, as outlined in Figure 5(b).
Effect of NAC and fluoxetine on protein expression of (a) SOCS3 and (b) pSTAT3 in rats subjected to the FST. Data are mean ± SEM (N = 12). **P < 0.01 and ***P < 0.001.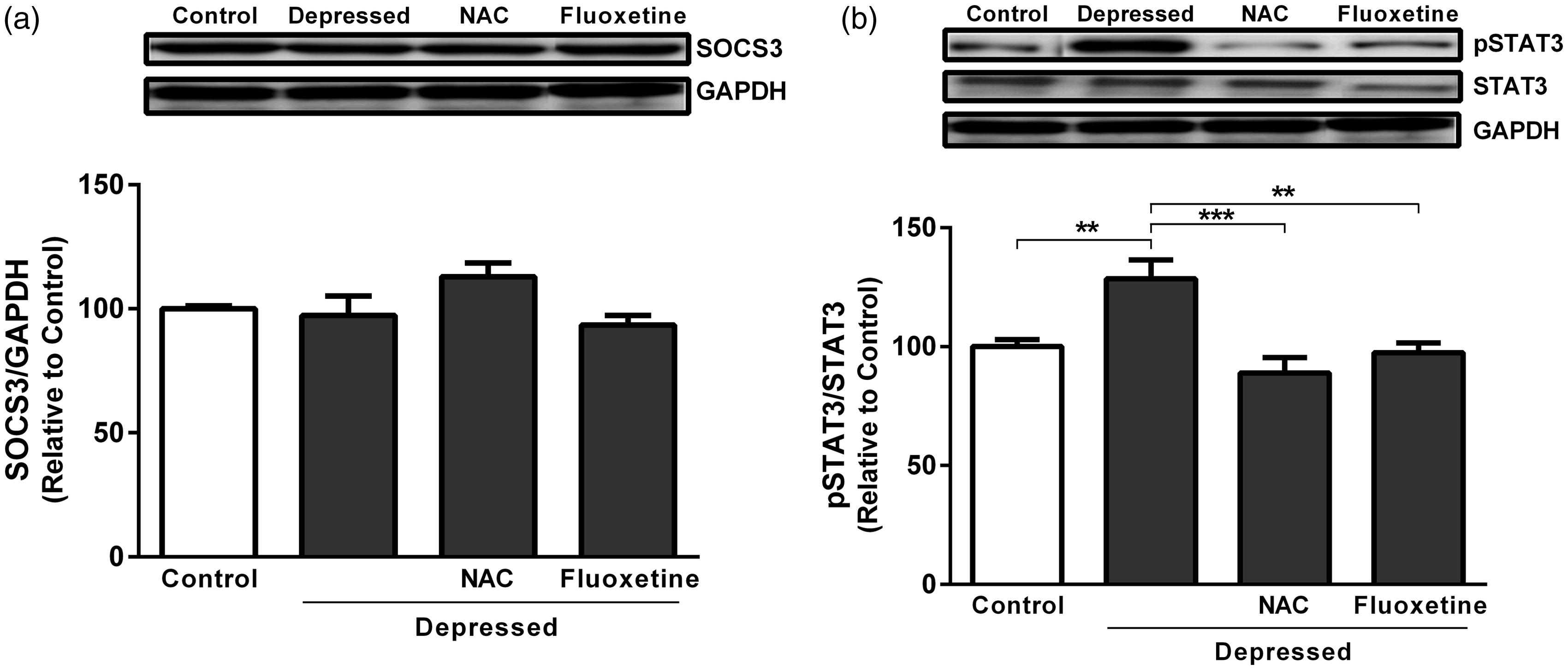
Discussion
NAC targets diverse aspects of nervous system functioning contribute to the pathophysiology of major depression including glutamatergic transmission, the antioxidant GSH, neurotrophins, apoptosis, mitochondrial function, and inflammatory pathways. 48 NAC exhibits a better tolerability profile compared to other treatments. 48 Recent evidence has suggested that NAC may exert therapeutic benefits in multiple neuropsychiatric disorders, including major depression. 49 Therefore, the precise analysis of how NAC works is crucial to both understanding the core biology of MDD and investigating promising novel therapies that act on relevant pathways.
Due to the complex pathophysiology and symptoms of depressive disorders in humans, the creation of animal models has been difficult so far. Additionally, investigation of the antidepressant-like activity of novel compounds is sensitive for predictive validity. The FST remains one of the most common antidepressant screening tools with good reliability and strong predictive validity. This test considered to be a suitable model to detect antidepressant activity due to its ability to detect the majority of antidepressants and to discriminate antidepressants from neuroleptics and anxiolytics. 50 For these reasons, a range of studies have used FST as a behavioral model for the evaluation of depressive-like behavior and the prediction of antidepressant drug efficacy in laboratory rodents.36,37 In this study, we used the FST as a reliable screening test to assess antidepressant effect of NAC. The present results demonstrated that NAC exerts an antidepressant effect that can be observed following its acute administration. NAC decreased the immobility time in the FST. This result supports the findings of Ferreira et al. 28 who demonstrated that the acute administration of NAC (15–150 mg/kg) induced antidepressant-like effects in adult male Wistar rats subjected to the FST. Also, a recent clinical study conducted by Berk et al. 27 suggested the antidepressant-like effects of NAC. As NAC and fluoxetine did not induce any significant change in exploratory activity in the OFT, these results demonstrate the antidepressant-like effects of NAC.
Strong evidence suggests a link between MDD and the hypothalamic–pituitary–adrenal (HPA) axis dysfunction. 51 Indeed, the current study showed marked increase in serum corticosterone concentrations occurred immediately after the 5 min FST session. In agreement with our results, swim stressors have been reported to elevate corticosterone plasma levels immediately after the FST.52,53 In contrast, Karandrea et al. 54 demonstrated that acute single stress episodes raised corticosterone levels more than repeated chronic stress did. Furthermore, medications that normalize the HPA axis system are hypothesized to play an important role in mediating antidepressant activity. 55 Interestingly, for the first time, the present data indicated that acute treatment with the antioxidant NAC significantly reduced serum corticosterone levels of depressed rats subjected to the FST compared to the depressed control rats. The mechanism of action that NAC utilizes to lower serum corticosterone levels is unclear and requires further investigation. However, to a lesser extent, NAC may contribute to the negative feedback mechanism of the corticotropin-releasing factor, which is under the control of monoamergic systems.
In contrast, acute treatment with fluoxetine did not attenuate corticosterone levels significantly in depressed rats. Fluoxetine is a classical antidepressant that was used in this study as a positive control. Its action is mediated by blocking serotonin (5-hydroxytryptamine, 5-HT) reuptake at the synapse, resulting in an elevation of extracellular 5-HT concentrations in the limbic regions of brain that can act through various postsynaptic 5-HT receptors.56,57 This behavioral profile elicited by fluoxetine is similar to that previously reported.38,58 The normalization of serum corticosterone levels and the negative feedback mechanism have been reported to be more effective after chronic fluoxetine treatment. 59 Additionally, Chen et al. 60 reported that the abnormalities in the HPA axis system induced in mice by swimming stress could be reversed by chronic, but not acute fluoxetine treatment. This might be attributed to the desensitization of hypothalamic post-synaptic receptors and/or the up-regulation of central corticosteroid receptors involved in the negative feedback mechanism. 61 On the contrary, Connor et al. 62 confirmed that chronic treatment with the selective serotonin reuptake inhibitor (SSRI) paroxetine exhibited no effect on corticosterone levels. The reason for this discrepancy is that fluoxetine may mediate HPA axis stimulation by increasing serotonergic activity. 63 In addition, many factors such as the duration of stress and treatment, as well as the time of blood collection, may influence the effect of antidepressant drugs on HPA axis activity.
Recently, less than two-thirds of patients who received treatment with the currently available SSRI antidepressant drugs achieve remission. 6 Therefore, exploration of new antidepressants with different mechanisms of action is essential to ameliorate treatment outcome. Many studies support a crucial role for oxidative stress in the pathophysiology of major depression.6,8 In the current study, FST generated an increase in H2O2 and a decrease in GSH levels in the prefrontal cortex and hippocampus of depressed rats. The involvement of endogenous ROS H2O2 and its scavenging antioxidant enzymes, catalase and GSH peroxidase together with the antioxidant GSH, has long been recognized in major depression.6,8 Accordingly, the disturbances in oxidative stress parameters observed in this experiment may reflect a decline in the activities of free radical scavenging enzymes. This oxidative damage likely further exacerbates lipid peroxidation and neuronal damage. 64 In addition, oxidative damage may interfere with critical GSH-related processes such as antioxidant defense, the detoxification of electrophilic xenobiotics, the storage and transport of cysteine, and the regulation of cell proliferation and signaling pathways. 65 Similar findings were reported for recent preclinical66,67 and clinical trials.68,69
Antioxidants are of great importance in preventing oxidative stress-related disorders such as major depression, and recently, they have been considered as promising therapeutic agents for treating such disorders. In the present study, treatment of rats subjected to the FST with NAC significantly reduced the oxidative damage caused by ROS and restored the depleted GSH levels in the prefrontal cortex and hippocampus. These results are in accordance with similar findings obtained from previous preclinical studies.30,66 Furthermore, several preclinical studies have suggested that NAC protects the brain from oxidative stress due to its antioxidant properties.30,66 NAC acts as an antioxidant by modulating the redox status of cells; the compound augments the synthesis of the endogenous antioxidant GSH, which is usually depleted as a result of increased oxidative stress. 70 Moreover, NAC acts as a direct scavenger of free radicals in the cells. 71 Both of these antioxidant activities of NAC may be related to protective effects against oxidative stress and the consequent reduction in immobility time induced by the FST.
Similarly, treatment with the classical antidepressant fluoxetine produced a significant decrease in H2O2 and an increase in GSH levels. This investigation may have provided core evidence that fluoxetine not only reduces the immobility time by increasing the serotonin concentration at the synapse but also modulates oxidative stress pathways. Lobato et al. 66 investigated the similar effects of acute and chronic fluoxetine treatment; however, these studies were performed in mice using the FST. Another study reported by Zafir et al. 72 confirmed the neuroprotective effect of chronic treatment with certain antidepressants (fluoxetine, imipramine and venlafaxine) in rats through the modulation of oxidative stress pathways using the FST, along with other animal models of depression. In one clinical study, the authors demonstrated that combined therapy with fluoxetine and acetylsalicylic acid resulted in an improvement in oxidative stress parameters in patients treated for depression. 73
Increased ROS and decreased antioxidant protection associated with MDD can provoke inflammatory responses. In addition, recent evidence suggests that inflammatory process, working in concert with ROS, may play a crucial role in the pathogenesis of depression.74,75 The JAK/STAT signaling pathway is an inflammatory pathway, and its components are widely expressed in several areas of the brain, such as the cerebral cortex and the hippocampus.14,16 Dysregulation of the JAK/STAT pathway has been found to be a key factor in a variety of neurodegenerative diseases, highlighting the importance of understanding how this pathway can influence the fate and function of brain cells. 17 In this study, FST induced an increase in STAT3 mRNA expression, as well the protein phosphorylation in the hippocampus and prefrontal cortex of depressed rats. In consequence of FST-evoked oxidative stress, the released H2O2 activates STAT3 signaling pathway. The phosphorylation of STAT proteins at a specific tyrosine residue via an SH2 domain-phosphotyrosine interaction is required for the dimerization of the STAT proteins and their translocation to the nucleus. 76 H2O2 has been demonstrated to stimulate tyrosine phosphorylation in some cells, as well as to inhibit tyrosine phosphatases. 77 Therefore, the mechanism of H2O2 activation of STAT3 proteins likely involves the activation of cellular tyrosine kinases such as JAK2 and tyrosine kinase 2 (TYK2) that phosphorylate the STAT3 proteins or inactivates a STAT-3 phosphatase. 78 Moreover, cytokines released by immune cells during inflammatory processes may activate the STAT3 transcription factor. 79
In the present work, we tested the hypothesis that NAC may induce anti-inflammatory effects by inhibiting the activation of the JAK/STAT pathway. We found that treatment with NAC significantly decreased STAT3 mRNA expression and STAT3 protein phosphorylation levels in depressed rats. NAC reduced the FST-induced ROS, inhibited the activation of STAT3, and suppressed STAT3 transcription factor expression. The underlying mechanisms that mediate NAC’s reduction of STAT3 expression in the rat hippocampus and prefrontal cortex may include the inhibition of STAT3 phosphorylation at Tyr705 by blocking the phosphorylation and thus the tyrosine kinase activity of the upstream proteins JAK1, JAK2, JAK 3, and Tyk2. 80 In addition, NAC is well known to function as GSH precursor and is a major regulator of the redox system in the cells, which may mediate its antioxidant properties. 81 The glutathionylation of STAT3 inhibits its activation through JAK proteins. 82 Moreover, NAC may modulate cytokines receptors that signal through the JAK/STAT pathway. 83
More interestingly, there was diminished expression of STAT3 mRNA and protein phosphorylation in fluoxetine-treated rats. These novel findings may be involved in the mechanisms through which fluoxetine is able to improve depressive symptoms. Chronic fluoxetine administration has been reported to inhibit ERK1/2 phosphorylation in the rat brain 84 and interferon alpha (IFN-α)-induced 5-HT uptake by interfering with ERK1/2. 85 Inhibition of the ERK1/2 pathway mimics the function of an antidepressant. 86 In addition, fluoxetine may inhibit glycogen synthase kinase-3β activity by increasing glycogen synthase kinase-3β phosphorylation at Ser9. 87
Suppressors of cytokine signaling (SOCS), key physiological regulators of inflammation, are considered to be the main negative feedback loop that suppresses the JAK/STAT signaling pathway. 19 SOCS3 can be markedly up-regulated by different cytokines and in a variety of neuroendocrine and neuroinflammatory states. 19 Our results indicated that no differences exist between the depressed and non-depressed control groups in the induction of SOCS3 mRNA expression. These results suggest that at least one other negative regulatory pathway is involved in the regulation of the STAT3 transcription factor. In contrast to our findings, Mingmalairak et al. 88 reported that the SOCS3 gene is up-regulated in non-depressed mice compared with depressed mice. Treatment with either NAC or fluoxetine evoked a remarkably significant increase in SOCS3 mRNA expression. SOCS3 regulates JAK/STAT signaling by inhibiting JAK kinase activity, facilitating the proteasomal degradation of JAK and STAT proteins, and competing with STATs for binding to cytokine receptors. 89 On the other hand, neither the FST nor any of the treatment agents affect the SOCS3 protein levels. Since mRNA is eventually translated into protein, this result possibly indicates that SOCS3 mRNA is not expressed at a sufficiently high level to result in a more correlated level of protein abundance. Other possible explanation could be due to post-translational modification.
In conclusion, our study provides new findings suggesting the possible anti-inflammatory mechanisms through which NAC is able to treat MDD. The inhibition of excessive STAT3 protein activation and SOCS3 gene induction contribute to the anti-inflammatory activity of NAC. In addition, this study provides new evidence of the involvement of the STAT3/SOCS3 signaling pathway in the pathogenesis of depression. Therefore, depression research may target the JAK/STAT signaling pathway to provide a novel effective therapy for treating MDD. However, additional preclinical studies and clinical trials are needed to better elucidate the effects of this therapeutic strategy.
Footnotes
Authors’ contributions
MMA and IHH carried out the animal experimentation and biochemical estimations, MMA and ND conducted the gene and protein analysis, Nouf MA, NMA, and SA conceived of the study, and participated in its design and coordination, AMM and IHH analyzed the data, AMM drafted the manuscript and all authors read and approved the final manuscript.
Acknowledgements
This research project was supported by a grant from the Research Center of the Female Scientific and Medical Colleges, Deanship of Scientific Research, King Saud University, Saudi Arabia.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.