
Abstract
The purpose of this study was to evaluate the combination effect of resveratrol and methoxyamine on radiosensitivity of iododeoxyuridine in spheroid culture of U87MG glioblastoma cell line using colony formation and alkaline comet assays. Spheroids on day-20 with 350 µm diameters were treated with 20 µM resveratrol and/or 6 mM methoxyamine and/or 1 µM iododeoxyuridine for one volume doubling time (67 h), and then irradiated with 2 Gy gamma-radiation (60Co) in different groups. After treatment, viability of the cells, colony forming ability and DNA damages were obtained by blue dye exclusion, colony formation and alkaline comet assay, respectively. Our results showed that methoxyamine and resveratrol could significantly reduce colony number and induce the DNA damages of glioblastoma spheroid cells treated with iododeoxyuridine in combination with gamma-rays. Therefore, methoxyamine as base excision repair inhibitor and resveratrol as hypoxia inducible factor 1-alpha inhibitor in combination with iododeoxyuridine as radiosensitizer enhanced the radiosensitization of glioblastoma spheroid cells.
Introduction
Gliomas are the most primary prevalent and aggressive form of intracranial tumors affecting adults 45–60 years of age.1,2 Despite advances in the combination of conventional surgery, radiotherapy and chemotherapy, median survival is poor in the range of 14 months. 3 Therefore, developing novel therapeutic strategies are obligatory for this disease. So there remains a need for more efficacious and less cytotoxic treatments. However, the dose essential to control these tumors, effectively far exceeds the tolerance of the surrounded normal tissues. 4 These unsatisfactory clinical results have motivated the search for methods to sensitize tumor cells to ionizing radiation as a choice. Therefore, efforts of many researchers have been focused on radiosensitizers, which lower the radiation dose-response threshold for cancer cells without enhancing the radio sensitivity of normal cells. One strategy to improve results uses halogenated pyrimidines analogues, iododeoxyuridine (IUdR), which is among the most potent non-hypoxic radiation sensitizers known. 5 These thymidine analogues are incorporated into DNA of cycling cells, so an increased rate of proliferation should result in increased radio sensitization of this subpopulation, and the degree of radio sensitization directly reflected the extent of thymidine replacement in the replicating DNA. 6 The absorption of IUdR decreases in spheroid culture. 7
Hypoxia, which activates the hypoxia inducible factor-1 alpha (HIF1-α) pathway, is a common feature in malignant gliomas and reduces the sensitivity of tumor cells to radiation (Rad) and chemotherapeutic drugs. 8 Thus, inhibition of HIF1-α may sensitize resistant glioma cells to the cytotoxic effect of Rad and chemotherapeutic agents. Research shows that resveratrol (trans-3, 4′, 5-trihydroxystilbene) (Res) inhibits HIF1-α protein expression in the hypoxia condition. 9 Res, a polyphenolic phytoalexin, considerably inhibited hypoxia-induced HIF1-α protein accumulation in cancer cells but did not change HIF1-α mRNA levels. 10 Thus hypoxia cells move out of G0 and uptake the analogues (IUdR), thus sensitized to Rad. This combined idea may be a novel plan to enhance the efficacy of anticancer therapy in human cancers.
Effectiveness of chemotherapy or Rad treatment is influenced by DNA repair. 11 Recent studies have shown that IUdR incorporated into DNA can be recognized by base excision repair (BER) proteins. 11 Enhancement of halogenated pyrimidine-induced radiosensitization may be performed by combining their use with repair inhibitors. 12 Many chemical compounds that specifically pick out pivotal steps in diverse DNA repair pathways are now being investigated, and some of them are already in various stages of clinical trials. Furthermore, combinations of anticancer agents or radiotherapy with these recently developed inhibitors of DNA repair pathways have considerable therapeutic potential. MX is a small molecule that binds directly to the apyrimidinic (AP) sites and blocking the BER pathway. 13 MX is known to synergistically enhance the therapeutic efficacy of DNA-damaging agents. 14 The purpose of this study is to investigate the merged effect of Res and MX on the radiosensitivity of IUdR in spheroid culture of U87MG glioblastoma with 350 µm diameter. The multicellular tumor spheroid represents an in vitro—in vivo transition model which exhibits important in vivo solid tumor correlates. These include intimate cell–cell contact, individual hypoxic cell populations and cycle times that range from comparable to exponential monolayer rates through an essentially non-dividing state. In brief, they combine the relevance of organized tissues with the accuracy of in vitro methodology. 15
Materials and methods
Cell line
Human glioblastoma cell line, U87MG, was purchased from Pasteur Institute of Iran. This cell line was cultured in minimum essential medium (MEM) (PAA, PAA Laboratories Ply Ltd, Australia) supplemented with 10% fetal bovine serum (FBS) (PAA), 100 U/mL of penicillin and 100 mg/mL of streptomycin (PAA).
Monolayer culture and doubling time calculation
Cells were cultured as monolayer at a density of 104 cells/cm2 in T-25 tissue culture flasks (SPL Life Sciences Co, Ltd, Korea). Cultures were incubated at 37℃ in a humidified atmosphere of 5% CO2. Cultures were propagated, and cells were harvested by trypsinizing cultures with 0.25% Trypsin (sigma, St Louis, MO, USA) (w/v) and 1 mM ethylene diamine tetraacetic acid (EDTA) (Sigma) in phosphate buffer saline (PBS) (Sigma).
Three passages after cells thawing, the cells were cultured at a density of 2 × 104 per well in multiwell plates (24 wells/plate) (SPL). At 24 h intervals, the cells from triplicate wells were removed by 1 mM EDTA/.25% Trypsin (w/v) treatment and counted in a hemocytometer. An average of nine counts was used to define each point. Doubling time was calculated using the slope of the logarithmic phase of growth curve.
Spheroid culture
Spheroids were established using the liquid overlay technique. Viable cells (5 × 105) were seeded into 100 mm Petri dishes (SPL) covered with a thin layer of 1% agar (Merck, Darmstadt, Germany) containing 10 mL of MEM supplemented with 10% FBS. The plates were incubated at 37℃ in a humidified atmosphere of 5% CO2. Half of the culture medium was recovered with new culture medium twice per week. 16
Irradiation procedure
For gamma radiation, the medium was replaced with fresh medium, and the spheroids were irradiated using 60Co source (Theratron-780 c, MDS Nordion) at a dose rate of 1.45 cGy/s for 2 Gy. For Rad treatment, culture flasks were put under collimator of equipment at 80.5 cm distance between the head of device and the floor of flasks, and the field size and the period of irradiation were 20 × 15 cm2 and 2.30 min, respectively.
Spheroid treatment
On day 20, spheroids with 350 µm diameter were treated with IUdR and/or Res and/or MX and/or gamma radiation. IUdR and/or Res and/or MX treatment were done for 1 volume doubling time (VDT) (67 h) at 37℃ in a humidified atmosphere and 5% CO2. As control, the spheroids of one plate were not treated. The treatments were carried out as stated in following groups:
Treated with Res (IC10:20 μM) for 67 h.
17
Treated with IUdR (1 μM) for 67 h. Treated with MX (6 mM) for 67 h. Irradiated with 60Co (2 Gy). Treated simultaneously with Res (20 μM) and IUdR (1 μM) for 67 h. Treated simultaneously with Res (20 μM) and IUdR (1 μM) and MX (6 mM) for 67 h. Treated with IUdR (1 μM) for 67 h and then irradiated with 60Co (2 Gy). Treated simultaneously with IUdR (1 μM) and Res (20 μM) for 67 h, then irradiated with 60Co (2 Gy). Treated simultaneously with IUdR (1 μM) and MX (6 mM) for 67 h, then irradiated with 60Co (2 Gy). Treated simultaneously with IUdR (1 μM) and Res (20 μM) and MX (6 mM) for 67 h, then irradiated with 60Co (2 Gy).
The viability of control and treated spheroids were determined using trypan blue dye exclusion assay. Then, the cell and DNA damages were evaluated using the colony formation and alkaline comet assay methods.
Trypan blue dye exclusion assay
The single cells from treated and control spheroid cultures were combined with trypan blue at a ratio of 9:1. After 2–3 minu, the combination was checked under an inverted microscope (Bell). The percentage of unstained cells out of the total number of cells was the viability of each cell group.
Colony formation assay
Treated and control single cells suspensions from spheroid culture were plated in 60 mm Petri dishes (SPL) with MEM containing 10% FBS for colony formation test. The cells were incubated at 37℃ in a humidified atmosphere of 5% CO2. After 10 days, cultures fixed with 2% formaldehyde in PBS, stained with crystal violet, then colonies which included a minimum of 50 cells were counted by an inverted phase microscope (Bell). In order to estimate the ability of cells to form colonies, various concentration of single cells from spheroid (500, 1000, 2000, 3000, 4000, 5000, 6000 cells) were seeded into 60 mm plates with 5 ml MEM supplemented with 10% FBS. Plating efficiency (PE) was calculated, using the following equation
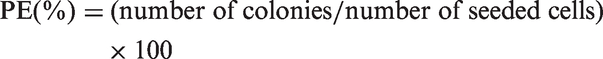
Alkaline comet assay
DNA damages induced by different treatments were determined using alkaline comet assay, previously optimized by our group. 18 Comet tail moment was determined by measuring the fluorescence intensity using the comet score software.
Statistical analysis
Data were given as mean values ± SEM with ‘n’ denoting the number of examination. Statistical analysis was performed using One-way analysis of variance (ANOVA) followed by Tukey’s test as the post hoc analysis using SPSS version16. The value of P < 0.05 was considered to be significant.
Results
Cell characteristics
Monolayer culture
Monolayer culture of the U87MG glioblastoma cell line was done on tissue culture flasks. The population doubling time calculated from the logarithmic phase of growth curve in the monolayer culture was approximately 25.61 ± 0.23 h.
Spheroid culture
Spheroids were cultured using the liquid overlay technique. The VDT (the period of time required for a spheroid to double in volume) calculated from the spheroid growth curve was approximately 67 ± 0.91 h which was applied as the drug treatment time consequently.
14
Figure 1 shows the phase contrast micrograph of 350 µm spheroids, 20 days after culture initiation.
Phase contrast micrograph of U87MG glioblastoma spheroids with 350 µm diameters on day 20 after culture initiation. Magnification 20×
Viability assay
Immediately after cell treatment with Res, IUdR, MX and Rad, cells were counted and viability was determined using the trypan blue dye exclusion assay. As can be seen in Figure 2, Res, IUdR, MX, Rad and combination of them did not have any effect on the viability of U87MG spheroid cells (P > 0.05).
Effect of resveratrol (Res), iododeoxyuridine (IUdR), methoxyamine (MX) and gamma radiation (Rad) and combination of them on the viability of U87MG spheroid cells with 350 µm in diameter, using trypan blue dye exclusion assay. The values are the mean ± SEM of three experiments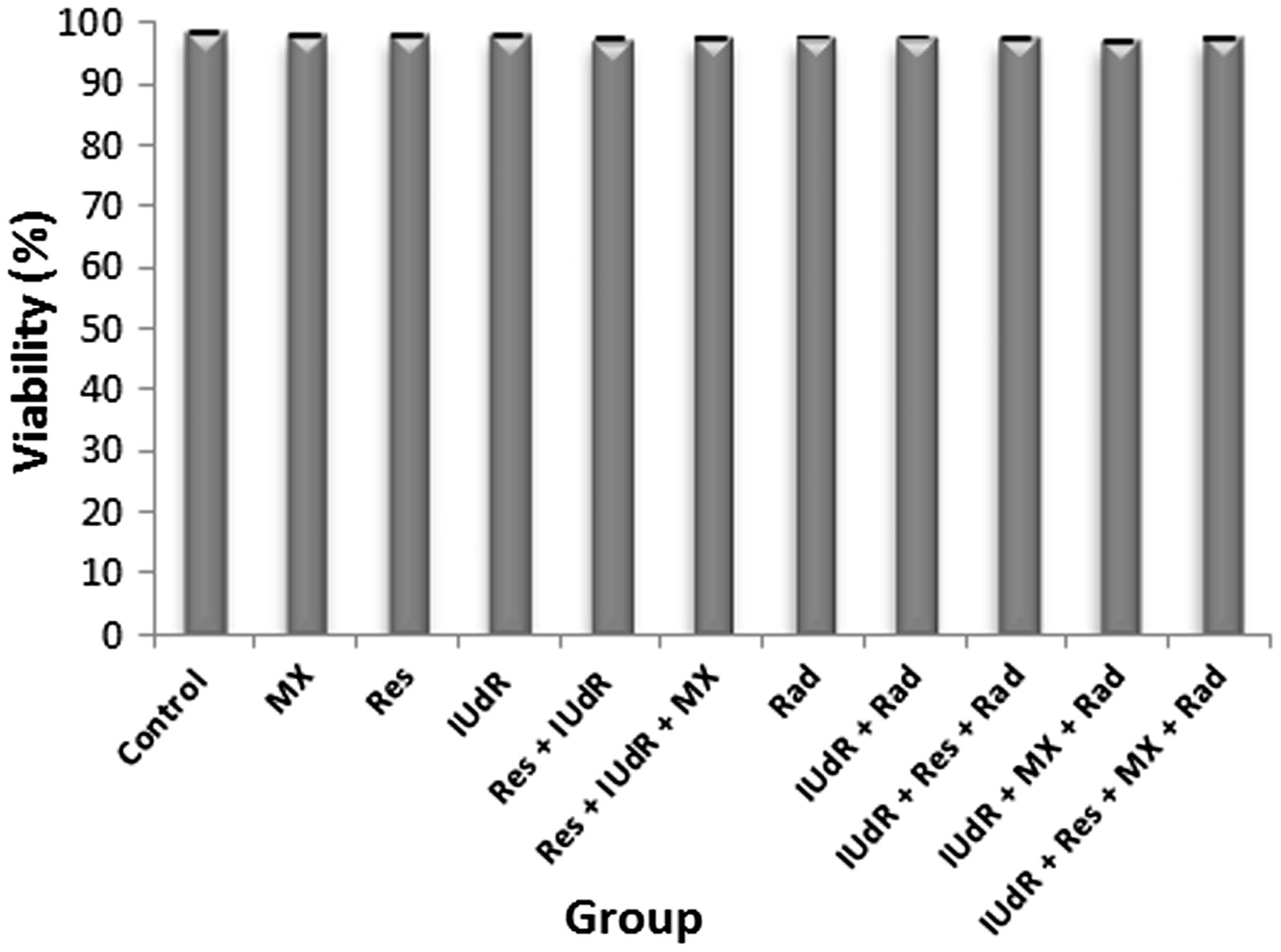
Effects of Res, IUdR, MX and gamma-ray radiation on colony forming ability
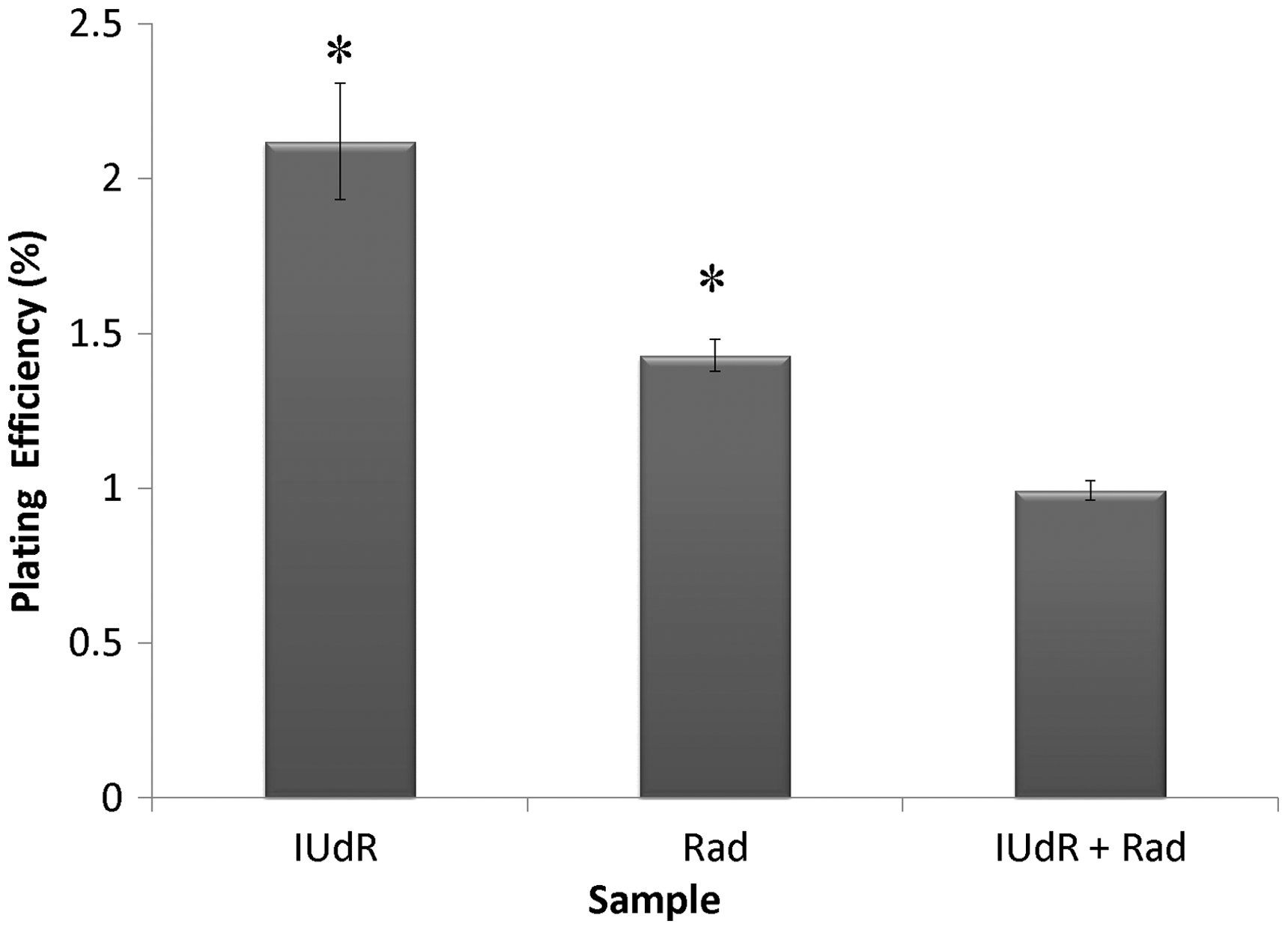
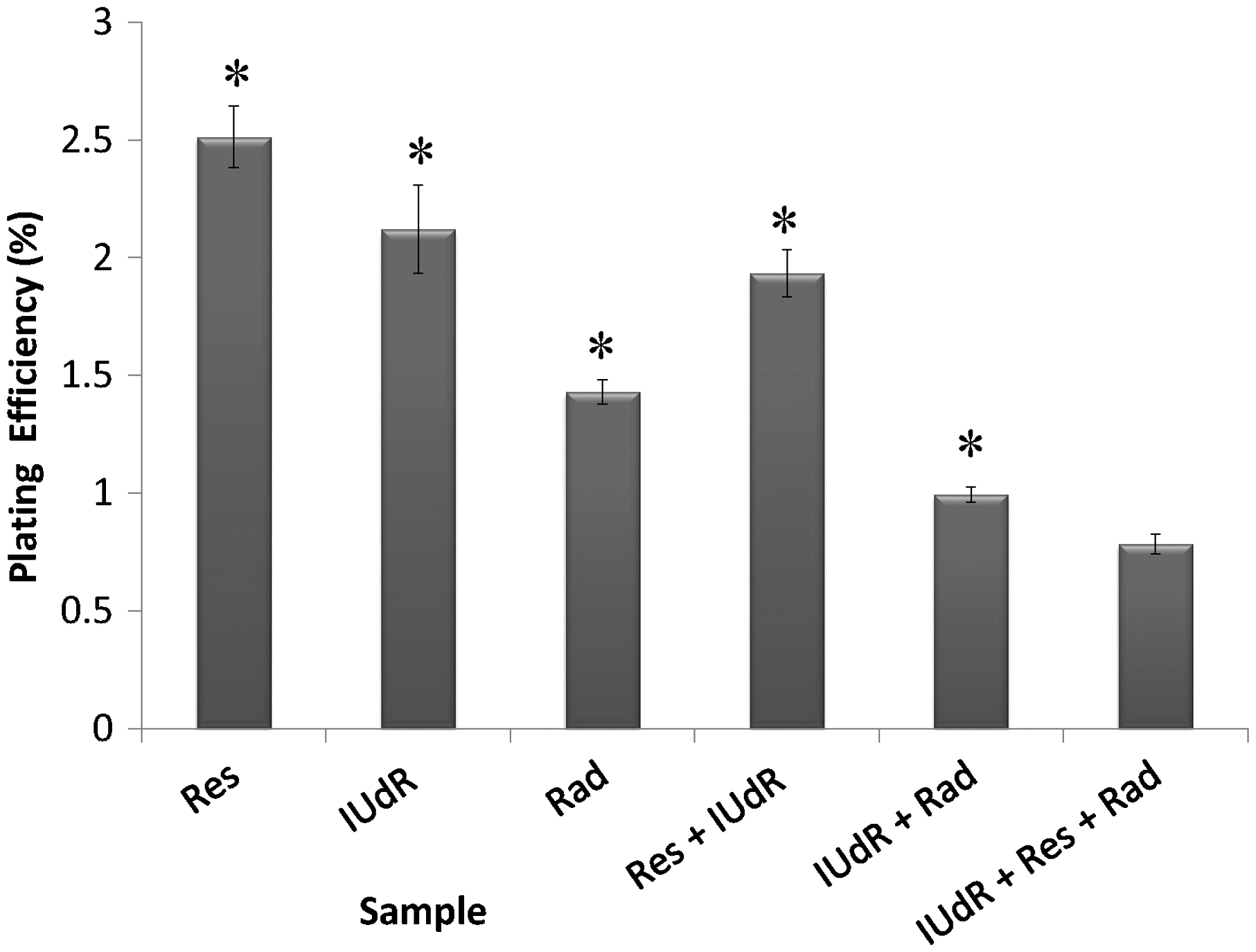
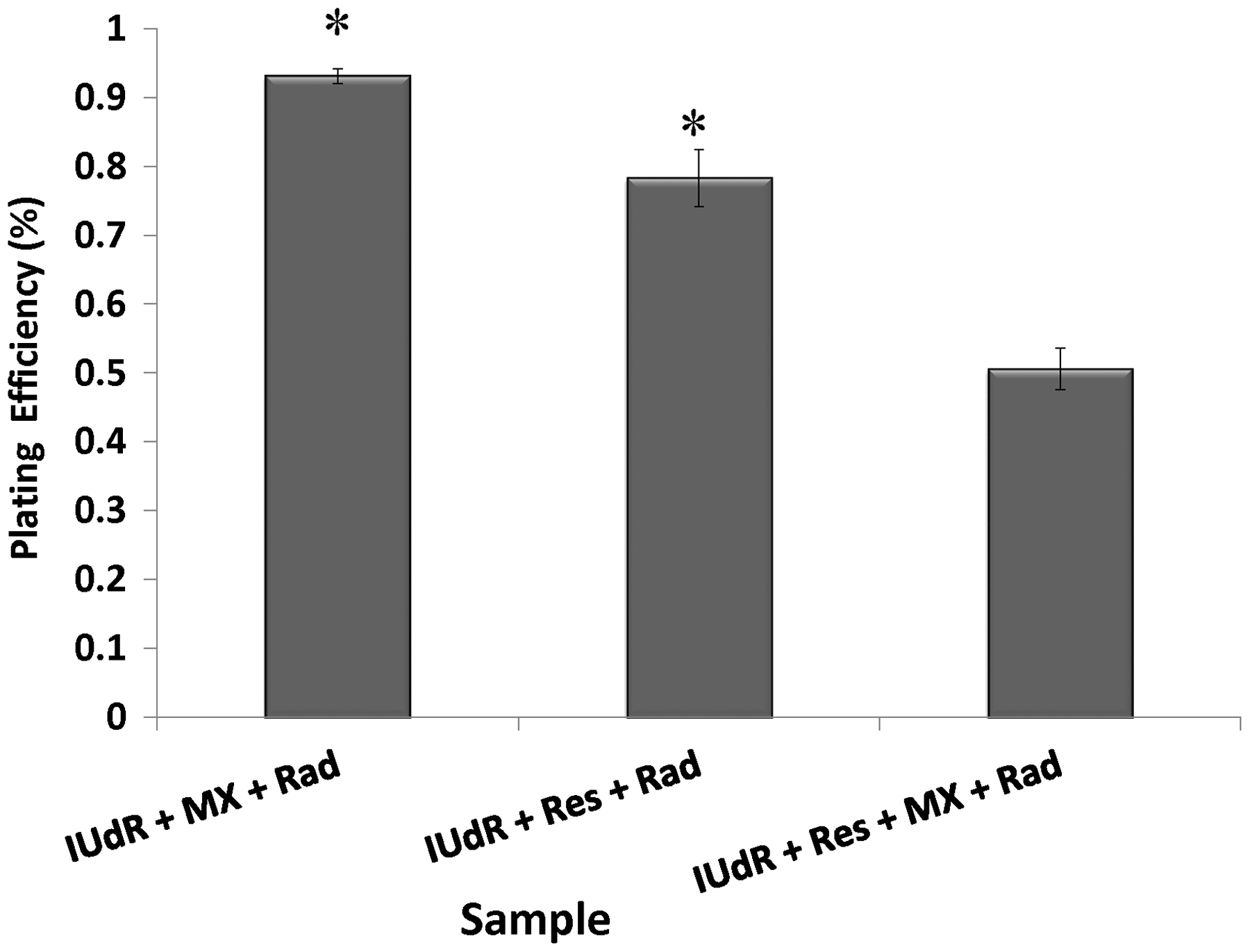
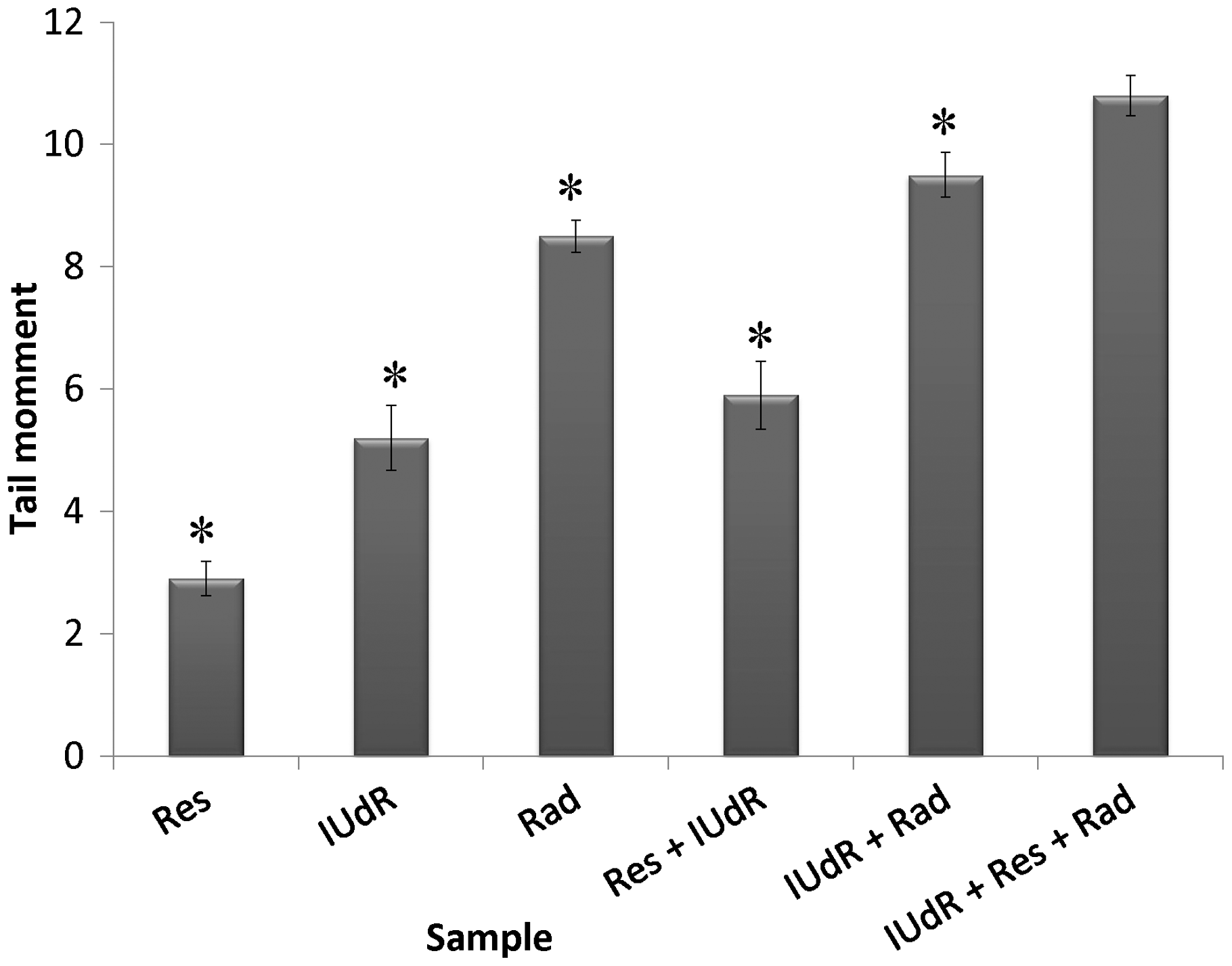
The cell response to drugs and Rad in terms of colony formation was studied by applying 20 µM Res, 1 µM IUdR, 6 mM MX and 2 Gy of gamma-ray radiation on the basis of 10 groups that described in the section of materials and methods. Figure 3 shows the effect of 20 µM Res, 1 µM IUdR, 6 mM MX and 2 Gy of gamma-ray radiation on the PE of U87MG glioblastoma cell line in spheroid culture. As can be seen, 20 µM Res and 6 mM MX alone had no significant effect on the PE of spheroid cells in comparison with control (P > 0.05). In contrast, 1 µM IUdR and 2 Gy of gamma-ray radiation separately reduced the PE of spheroid cells (P < 0.01). Figure 4 shows the role of IUdR in the radiosenesitivity of U87MG glioblastoma cell line. As can be seen, reduction of PE due to the combination treatment of IUdR and gamma-ray radiation is significantly more than each one alone (P < 0.01). In the next step, the role of Res as HIF inhibitor was studied on the radiosensitivity of IUdR. Figure 5 shows the effect of Res, IUdR and gamma radiation alone or in combination with each other on U87MG glioblastoma spheroids. As can be seen, the combination treatment of IUdR + Res + gamma radiation significantly reduced the colony formation ability of spheroid cells in comparison with each treatment alone or the combination of IUdR + Rad or IUdR + Res (P < 0.001). Finally, the combination effects of Res as HIF inhibitor and MX as BER inhibitor on the radiosensitivity of IUdR in U87MG glioblastoma cell line were evaluated. Figure 6 shows that combination treatment of IUdR + MX + Res + gamma radiation significantly reduced the colony formation ability of spheroid cells in comparison with IUdR + MX + gamma radiation or IUdR + Res + gamma radiation (P < 0.01).
Effect of 20 µM resveratrol (Res), 1 µM Iododeoxyuridine (IUdR), 6 mM methoxyamine (MX) and 2 Gy of gamma-ray radiation (Rad) on the plating efficiency of U87MG glioblastoma cell line in spheroid culture with 350 µm in diameter. The values are the mean ± SEM of three experiments Effect of 2 Gy of gamma-ray radiation alone or in combination with 1 µM IUdR on the plating efficiency of U87MG glioblastoma cell line in spheroid culture with 350 µm in diameter. (*P < 0.01 vs. IUdR + Rad).The values are the mean ± SEM of three experiments Effect of 1 µM IUdR and/or 20 µM resveratrol (Res) and/or 2 Gy of gamma-ray radiation on the plating efficiency of U87MG glioblastoma cell line in spheroid culture with 350 µm in diameter. (*P < 0.001 vs. IUdR + Res + Rad).The values are the mean ± SEM of three experiments Effect of 1 µM IUdR and/or 20 µM resveratrol (Res) and/or 6 mM methoxyamine (MX) and/or 2 Gy of gamma-ray radiation on the plating efficiency of U87MG glioblastoma cell line in spheroid culture with 350 µm in diameter. (*P < 0.01 vs. IUdR + Res + MX + Rad).The values are the mean ± SEM of three experiments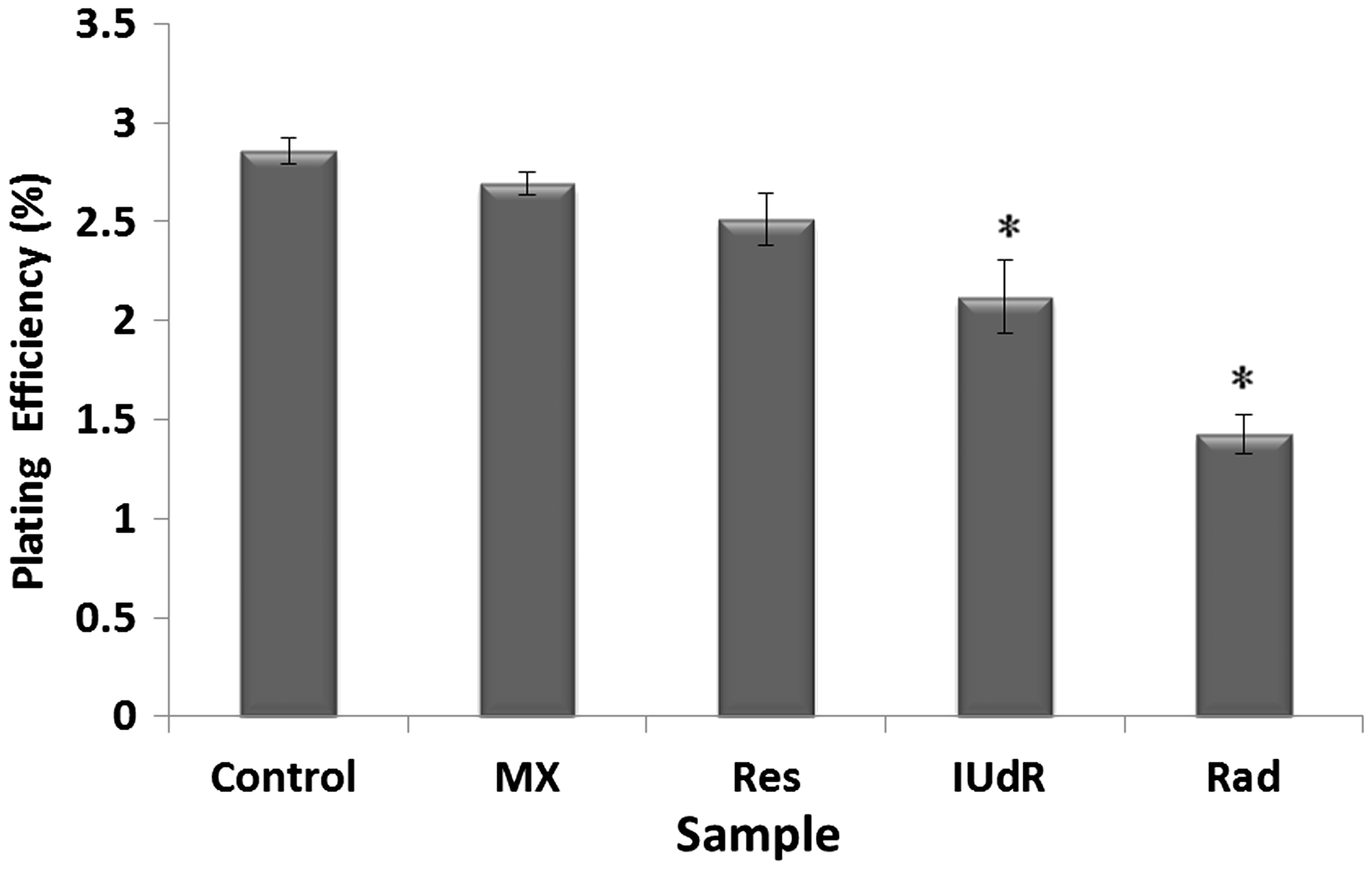
Effects of Res, IUdR, MX and gamma-ray radiation on DNA damages
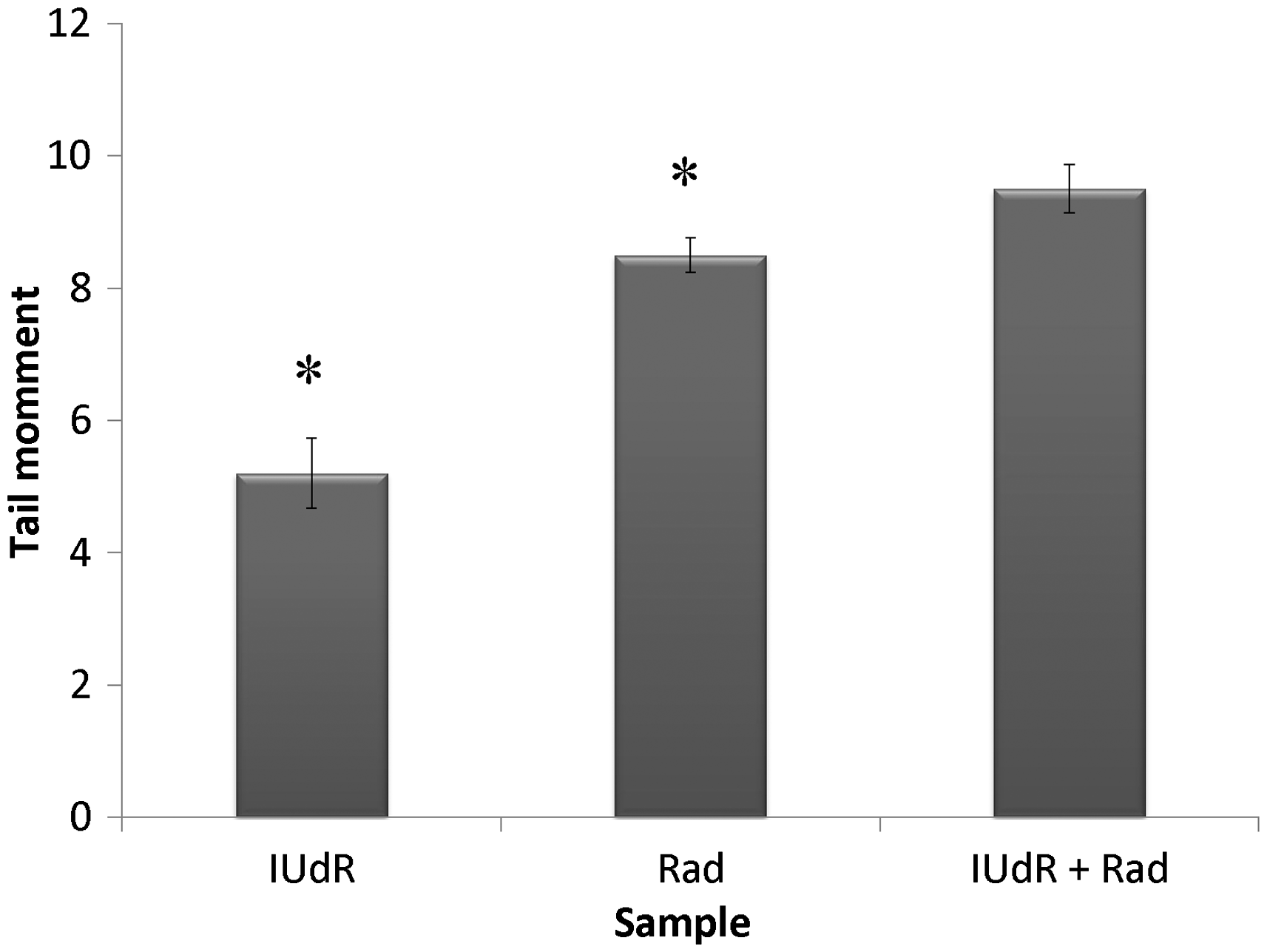
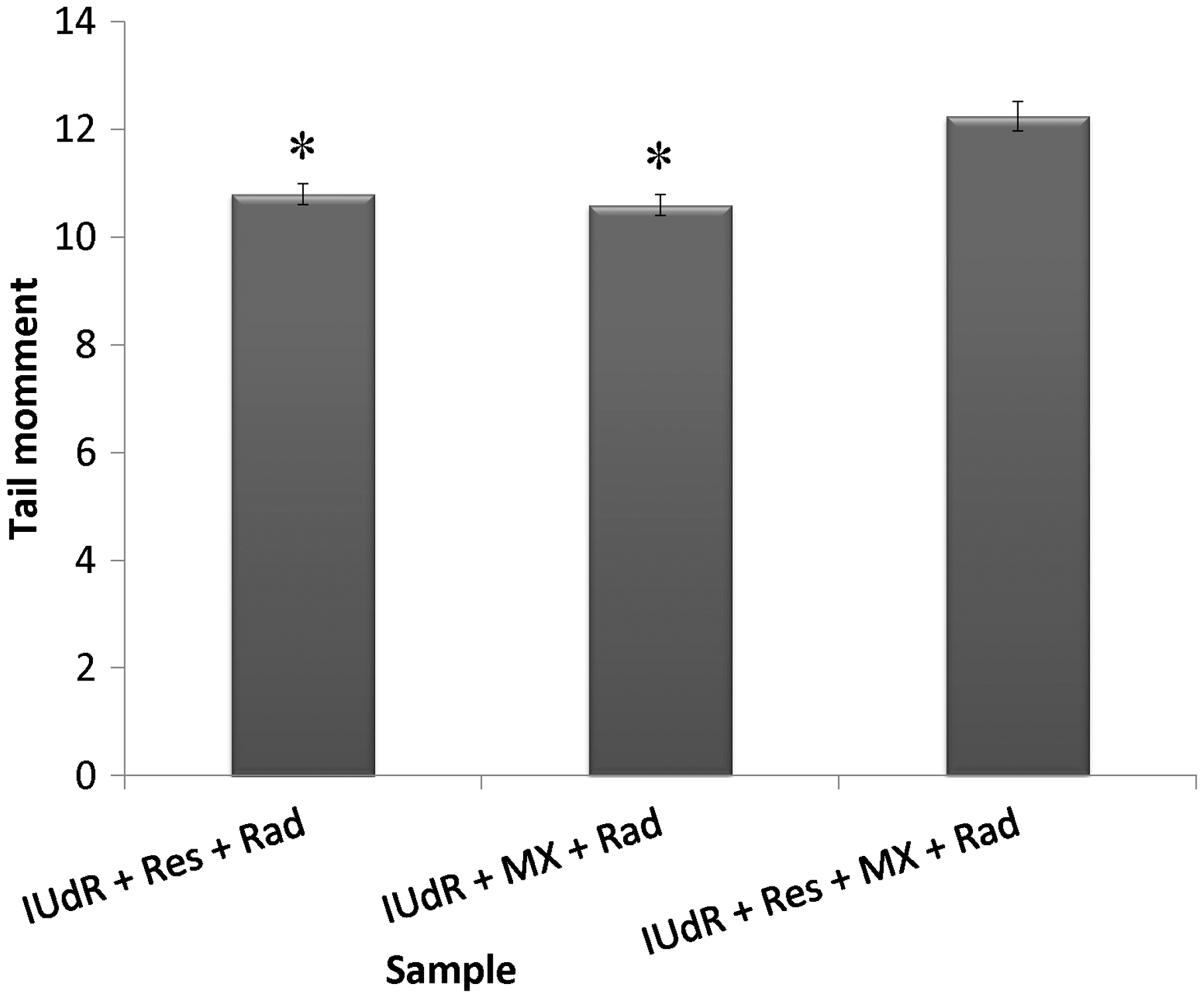
The level of DNA damages due to drugs and Rad treatment in terms of tail moment was studied by applying 20 µM Res, 1 µM IUdR, 6 mM MX and 2 Gy of gamma-ray radiation on the basis of 10 groups that described in the section of materials and methods. Figure 7 shows the effect of 20 µM Res, 1 µM IUdR, 6 mM MX and 2 Gy of gamma-ray radiation on the DNA damages induced in spheroid culture of U87MG glioblastoma cell line. As can be seen, 20 µM Res and 6 mM MX alone had no significant effect on the level of DNA damages in comparison with control (P > 0.05). In contrast, 1 µM IUdR and 2 Gy of gamma-ray radiation separately increased DNA damages induced in spheroid cells (P < 0.01). Figure 8 shows the role of IUdR in the radiosenesitivity of U87MG glioblastoma cell line using alkaline comet assay. As can be seen, induction of tail moment due to the combination treatment of IUdR and gamma-ray radiation is significantly more than each one alone. Then, the role of Res as HIF inhibitor was studied on the radiosensitivity of IUdR using alkaline comet assay. Figure 9 shows the effect of Res, IUdR and gamma radiation alone or in combination with each other on DNA damages induced in U87MG glioblastoma spheroids. As can be seen, the combination treatment of IUdR + Res + gamma radiation significantly induced DNA damages of spheroid cells in comparison with each treatment alone or the combination of IUdR + Rad or IUdR + Res (P < 0.001). Finally, the combination effects of Res as HIF inhibitor and MX as BER inhibitor on the radiosensitivity of IUdR in U87MG glioblastoma cell line were evaluated using alkaline comet assay. Figure 10 shows that combination treatment of IUdR + MX + Res + gamma radiation significantly induced the DNA damages of spheroid cells in comparison with IUdR + MX + gamma radiation or IUdR + Res + gamma radiation (P < 0.01).
Effect of 20 µM resveratrol (Res), 1 µM Iododeoxyuridine (IUdR), 6 mM methoxyamine (MX) and 2 Gy of gamma-ray radiation (Rad) on the level of DNA damages in spheroid culture of U87MG glioblastoma cell line with 350 µm in diameter. The values are the mean ± SEM of three experiments Effect of 2 Gy of gamma-ray radiation alone or in combination with 1 µM IUdR on the level of DNA damages in spheroid culture of U87MG glioblastoma cell line with 350 µm in diameter. (*P < 0.01 vs. IUdR + Rad).The values are the mean ± SEM of three experiments Effect of 1 µM IUdR and/or 20 µM resveratrol (Res) and/or 2 Gy of gamma-ray radiation on the level of DNA damages in spheroid culture of U87MG glioblastoma cell line with 350 µm in diameter. (*P < 0.001 vs. IUdR + Res + Rad).The values are the mean ± SEM of three experiments Effect of 1 µM IUdR and/or 20 µM resveratrol (Res) and/or 6 mM methoxyamine (MX) and/or 2 Gy of gamma-ray radiation on the level of DNA damages in spheroid culture of U87MG glioblastoma cell line with 350 µm in diameter. (*P < 0.01 vs. IUdR + Res + MX + Rad).The values are the mean ± SEM of three experiments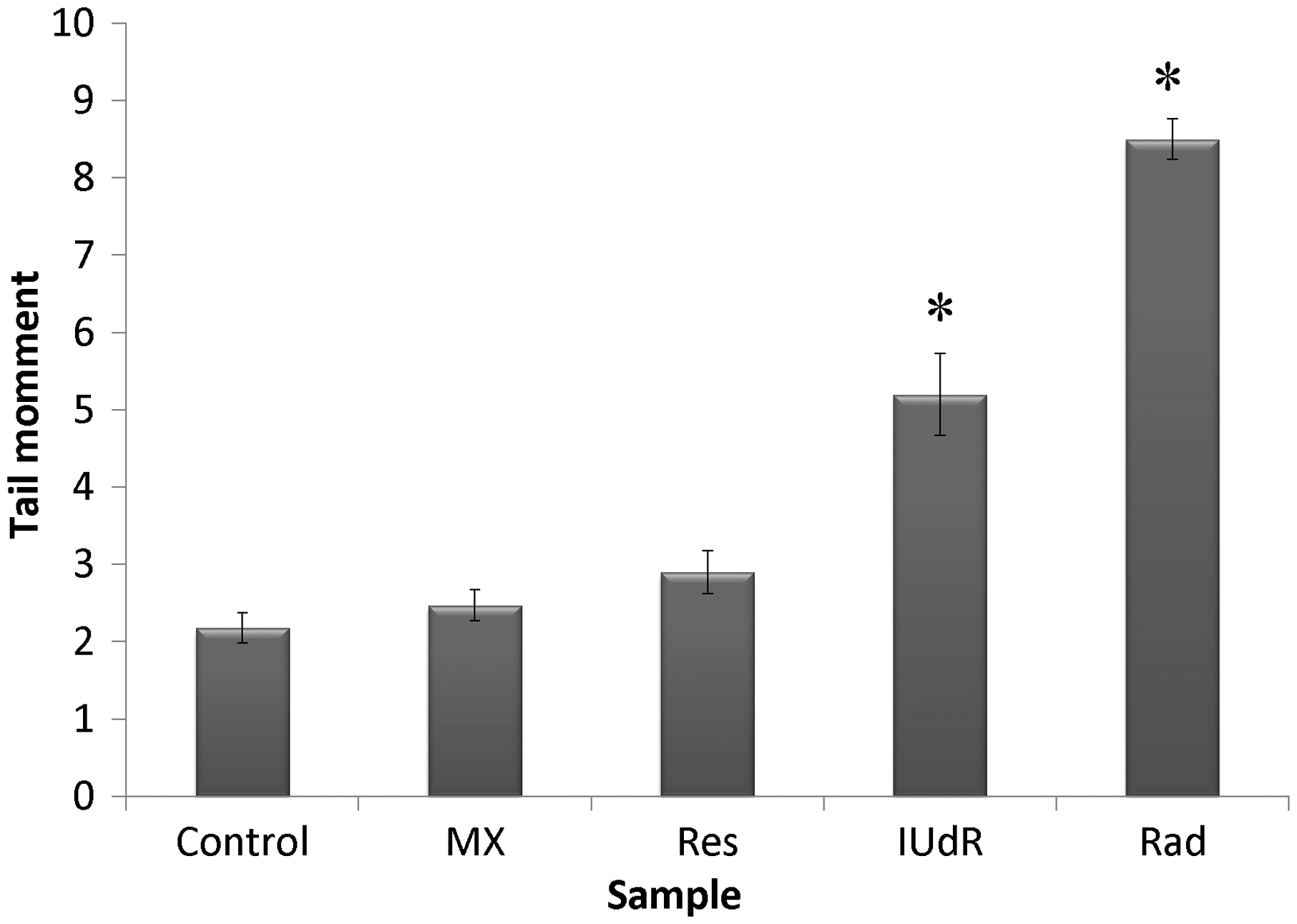
Discussion
Glioblastoma multiform (GBM) is a main type of human inherent and most radioresistant brain tumor. 19 Recently, integrate treatments using chemical/biological agents and radiotherapy have been engaged to either heighten tumor radiosensitivity or lessen ionizing radiation side-effects because the irradiation effective enough to control the tumors far exceeds the endurance of proximate normal brain tissues. 20 Eyvazzadeh et al. 21 showed that the percentage of double strand break and single strand break in DNA of U87MG blioblastoma cell line treated with 2 Gy of 6 MV X-ray radiation is 24% and 12%, respectively. IUdR is a thymidine analogue, known as a potential radiosensitizer for human cancers therapy. 22 In vitro and in vivo experimental works confirmed that when IUdR is incorporated into DNA instead of thymidine through replication and sensitizes the cells to ionizing radiation, the extent of radiosensitization corresponds directly with the level of halogenated pyrimidine-DNA absorption. 23 Although the cellular and molecular mechanisms of IUdR radiosensitization are not clearly elucidated, it is presumed that IUdR sensitizes cells through enhancing the formation of IR-induced double-strand break. 4 Two alternative, but not mutually exclusive, hypotheses have been formulated to interpret the radiosensitizing action of halogenated pyrimidines in living cells. 23 According to the first hypothesis, sensitization is caused by an increase in the amount of Rad damage induced in the DNA per unit absorbed dose. This increase in the induction of Rad damage is at least partly attributed to a reaction of the halogenated deoxyuridine with Rad-induced hydrated electrons forming highly reactive uracil radicals and halide ions that can abstract a hydrogen atom from the neighboring deoxyribose leading thus to a strand break. 24 According to the second hypothesis, the observed radiosensitization by halogenated pyrimidines is caused by an effect on cellular repair, or, by enhanced fixation of Rad induced PLD. 25 Williams et al. 6 showed the radiosensitivity of U251 glioma monolayer cells has been increased with 10 µM IUdR plus gamma radiation (2.5 and 5 and 10 Gy), with increasing Rad dose. Also, Neshasteh-Riz et al. 14 revealed that 1 µM IUdR and 2 Gy gamma 60Co inhibited the proliferation capacity of the U87MG glioblastoma spheroid cells (100 and 300 µm) compared with Rad alone. We also showed that 1 µM IUdR increased the cytogenetic effect of 2 Gy gamma-ray radiations using colony formation and comet assays (Figures 4 and 8).
Rad treatment requires free radicals from oxygen to demolish target cells, and to kill the same proportion of cells in hypoxic areas compared with oxic cells, approximately 2 to 3 times Rad dose is required. 26 Also IUdR absorption is significantly reduced when the tumor size is increased, and the cells in the median layers suffer from hypoxia due to oxygen deficiency. 27 A consequential component of the hypoxic response is the activation of the HIF1 transcription factor. 28 Under hypoxia conditions, HIF1-α is transferred from cytoplasm to nucleus and by attaching to HIF1-β, forms the HIF1 complex. 8 It activates more than 60 genes with various functions, leading to an increase in O2 delivery. Also in this condition, hypoxia induces cell cycle arrest in the G0 phase and decrease cells in S phase. 29 The implication of HIF1 in tumor resistance to therapies led us to hypothesize that inhibition of HIF1 may sensitize hypoxic cells to the cytotoxic effect of Rad by IUdR absorption. If hypoxic cells exit from G0 to S phase, can absorb IUdR and become sensitive to Rad. To achieve this situation in the in vitro cell culture, we used 350 µm spheroids to ensure the existence of hypoxia cells. 30
Res (trans-3,4',5-trihydroxystilbene) is a naturally occurring polyphenolic compound highly enriched in grapes, peanuts, red wine and a wide variety of food sources. 31 Recent evidence suggests that Res in combination with drugs, ionizing radiation or cytokines can be effectively used for the sensitization to apoptosis. 32 It appears that Res can sensitize the cells to various cytotoxic agents such as cyclosporin, paclitaxel, 5-fluorouracil and IUdR. Recent data have shown that Res inhibits activation of HIF1-α in the hypoxia conditions. 10 Zhang et al. 10 demonstrated the different doses of Res (5, 10, 50, 100 µM) decreased HIF1 and VEGF mRNA in SCC-9 human tongue squamous cell carcinoma and HepG2 hepatoma monolayer cells in hypoxia condition. Res remarkably inhibits HIF-1 a protein expression via regulating both protein translation and HIF1-α protein degradation. 17 After inhibition the HIF1-α, cells progress from G0 to cell cycle and could absorb IUdR in S phase. So the radiosensitivity of cells increased. Firouzi et al. 17 showed treatment of 350 µm U87MG glioblastoma spheroids with 20 µM Res can increase radiosensitivity of 1 µM IUdR. Panayiotidis et al. 33 reported that 25 µM Res decreased percentage of G0/G1 lung cancer cells (A549), and number of them has increased in S phase. In this study, we used this effect of Res to increase the IUdR absorption in hypoxia cells (Figures 5 and 9).
Efficacy of chemotherapy or Rad treatment is intimately dependent on DNA repair valence. 34 Robust repair of therapeutically induced DNA damage can provide significant resistance whereas tumor-specific defects in DNA repair or inhibition of specific DNA repair proteins can provide therapeutic advantage. Also, one mechanism of resistance to IUdR is attributed to efficient repair of IUdR-induced DNA lesions by the BER pathway. 14 BER is one of the major mechanisms conferring resistance to DNA damage-induced cytotoxicity. 35 A damaged base is excised by a damage-specific DNA glycosylase, leading to the formation of a potentially cytotoxic apurinic or AP site intermediate; the site (AP site) where this occurs is then processed by an AP endonuclease (APE1), via the so called short-patch BER pathway. 36 Two different approaches have been developed to suppress the BER pathway by preventing the AP site from being processed: chemical modification of AP sites to make them unsuitable for APE1 binding, and direct inhibition of APE1. 37 In this regard, the preferred option is the inhibition of BER after repair initiation, promoting the accumulation of cytotoxic BER intermediates such as AP sites and SSBs by inhibiting AP site repair with MX. MX is a small molecule that binds directly to the AP sites even more efficiently than the binding by APE1. 38 MX-bound AP sites cannot be processed by APE1, thus blocking the BER pathway and leading to the accumulation of the cytotoxic AP sites. 38 MX is known to synergistically improve the therapeutic efficacy of a wide range of DNA-damaging agents. Therefore, pharmacological agents that can manipulate the DNA repair system have the potential for enhancing the cytotoxic effects of chemotherapeutic agents and IR in cancer cells. Taverna et al. 11 showed that 6 mM MX plus 10 µM IUdR increase the Rad sensitivity in monolayer culture of CHO, EM9 and AA8 cell lines. This study confirmed our results that 6 mM MX increased the cytotoxic and genotoxic effect of 1 µM IUdR (Figures 6 and 10).
Khoei et al. 16 showed 2ME2 can inhibit HIF1-α activity and increased IUdR radiosensitivity and cytotoxic damages. They also showed that Res like 2ME2 has been acted as HIF1-α inhibitor but is safer because of its herbal nature and it can increase the cytotoxic damages of IUdR + Rad. They observed a significant decrease in PE percentage in combination treatment of Res + IUdR + Rad, and our results also indicated that the cytotoxicity of Res plus IUdR and Rad was mainly significant. 17
Finally, in this study, the combination effect of MX as BER inhibitor and Res as HIF inhibitor on the radiosentivity of IUdR was evaluated. Figures 6 and 10 show that the combination of IUdR + Rad + Res + MX was efficient to highly reduce clonogenic ability and induce DNA damages of U87MG cells in comparison with IUdR + MX + Rad or IUdR + Res + Rad. It seems Res could inhibit HIF-1α expression and induce to progress the hypoxia spheroid cells in the cell cycle to S phase. In this condition, these cells could uptake IUdR and sensitize to Rad. Finally, MX could increase all effects with inhibition of BER.
Conclusion
Our data suggest a new strategy for anticancer therapy for glioblastoma (GBM) because combination of MX and Res with IUdR can decrease colony formation ability and increase DNA damages of gamma-ray radiation in 350 µm spheroids. So the cytotoxicity effect of Rad and therapeutic ratio increases.
Footnotes
Authors’ contributions
All authors participated in the completion of the manuscript. MS conducted most of the experiments and wrote the manuscript, SK supplied critical reagents and contributed to the design and review of the study, and AM and FF contributed to the radiation experiments. The authors alone are responsible for the content and writing of the paper.
Acknowledgements
This work was supported by grants no. 20204 from the Razi Drug Research Centre of Iran University of Medical Sciences.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.