
Abstract
We cultivated human bladder smooth muscle cells (HBSMCs) under pressures of 0 or 200 cm H2O pressure for 24 h, before using microarray technology to extract and analyze the different expressions of miRNAs and mRNAs in the two groups. We also predicted the target mRNA of the miDNA and performed functional forecasting. Changes in miRNA were identified by quantitative real-time polymerase chain reaction (qRT-PCR) after overexpressing miRNA by transfection. We used flow cytometry to examine HBSMC proliferation, and we used qRT-PCR and Western blot analyses to quantify the expression and activation of mRNAs and proteins. There were nine upregulated and four downregulated miRNAs involved in cell proliferation, including miR 4323, which was identified by qRT-PCR (p = 0.027). In addition, miR 4323 was shown to inhibit LYN (p = 0.031), decrease lyn kinase (p = 0.037), and promotes the phosphorylation of extracellular regulated protein kinases 1 and 2 (Erk1/2) (p = 0.004). Moreover, overexpression of miR 4323 activated the proliferation pathway regulated by Erk1/2. Then, miR 4323 was shown to stimulate the proliferation of HBSMCs, with the proliferation index improving from 30.84 ± 4.57 to 52.13 ± 3.41 (p = 0.001). In summary, when the miRNA miR 4323 was overexpressed under cyclic hydrodynamic pressure, LYN is decreased and the Erk1/2 signaling pathway is activated; in addition, miR 4323 is involved in HBSMC proliferation when under hydrodynamic pressure.
Keywords
Introduction
There is a growing body of evidence that cyclic hydrodynamic pressure (HP) can profoundly modulate the morphology and functionality of human bladder smooth muscle cells (HBSMCs), including their migration, proliferation, and apoptosis. 1 In our previous studies, we showed that cyclic HP seemed to promote the proliferation of HBSMCs,2,3 and that some other factors appeared to be involved in this process. These factors included gap-junction protein connexin 43 (Cx43), serum-glucocorticoid regulated kinase 1 (SGK1), p27, various integrins, mitogen-activated protein kinase (MAPK), and phosphoinositide 3-kinase/SGK1.2,4–6 However, it remains difficult to establish an integrated signaling pathway.
Recently, an increasing number of studies have focused on the role of miRNA in important biological processes.7–10 MiRNAs are a large class of evolutionarily conserved, small, noncoding RNAs, typically 20–26 nucleotides in length, which primarily regulate gene expression at the post-transcriptional level by interacting with the 3′-untranslated regions (3′-UTRs) of specific target mRNAs in a sequence-specific manner.9,10 Recently, some reviewers have demonstrated that some miRNAs are differently expressed in human bladder cells under certain conditions, such as when bladder cancer is present.7,8 Moreover, they found that some miRNAs can regulate the proliferation of human bladder cells. 7 However, there are no reports of the profile and function of these differently expressed miRNAs in HBSMCs when under cyclic HP conditions. Therefore, we aimed to investigate the changes in miRNA expression regulated by this stimulation and to explore the role of miRNAs in the promotion of HBSMC proliferation by cyclic HP.
Methods
Primary cell cultivation
HBSMCs (cat. no. 4310; ScienCell, Carlsbad, CA, USA) were cultured in 35-mm culture dishes utilizing Dulbecco’s modified Eagle’s medium (low glucose; Hyclone Laboratories, Inc., Logan, UT, USA) with 10% fetal bovine serum (Hyclone), without antibiotics, and maintained at 37℃ in a humidified atmosphere of 5% CO2 and 95% air.
Stimulation by cyclic HP
HBSMCs were seeded onto scaffolds and then subjected to cyclic HP that simulated the bladder cycle for up to 24 h. In the HP group, the bladder cycle was 2 h in length, increasing from 0 to 10 cm H2O during the first 1.75 h, then rapidly up to 200 cm H2O, which was maintained for 0.25 h, before being reduced back to 0 cm H2O. The control group was maintained at 0 cm H2O throughout.
Microarray analysis of miRNA and mRNA
Total RNA was extracted using a TRIzol reagent (Invitrogen, Carlsbad, CA, USA), and microarray analysis was performed by Kangcheng Bio-tech Inc. (Shanghai, China).
The miRNA microarray
Total RNA was labeled using the miRCURY™ Hy3™/Hy5™ Power labeling kit. Then, Hy3™-labeled samples were hybridized on a miRCURY™ LNA Array (v.18.0) (Exiqon) according to the manufacturer’s instructions. Microarrays were scanned using the Axon GenePix 4000B microarray scanner (Axon Instruments, Foster City, CA). Scanned images were then imported into GenePix Pro 6.0 software (Axon Instruments) for grid alignment and data extraction.
The mRNA microarray
For microarray analysis, an Agilent Array platform (Agilent Technologies) was employed. Sample preparation and microarray hybridization were performed according to the manufacturer’s standard protocols, but with minor modifications. The Agilent Feature Extraction software (version 11.0.1.1) was used to analyze the acquired images. Quantile normalization and data processing were performed using the GeneSpring GX v11.5 software package (Agilent Technologies).
Analysis of microarray data
All fold-changes in miRNA and mRNA were measured and identified through fold-change filtering (changes ≥2-fold, compared between two groups), as previously reported. 11 The potential target mRNAs (identified by fold-change filtering) of the differently expressed miRNAs were examined with the aid of three target prediction databases (miRBase, miRnada, and TargetScan). Gene ontology analysis and Kyoto encyclopedia of genes and genomes pathway analysis of gene function were then used to further explore the biology of the predicted target genes regarding molecular function and involvement in known proliferation regulatory pathways.
Quantitative real-time polymerase chain reaction
Total RNA was extracted using TRIzol before performing quantitative real-time polymerase chain reaction (qRT-PCR) with a Bio-Rad iQ5 machine (Hercules, CA, USA). The PCR product quality was monitored by post-PCR melt-curve analysis.
For miRNA
cDNA was synthesized using an All-in-One™ miRNA qRT-PCR Detection Kit (GeneCopoeia, Foster City, CA, USA), with the PCR conditions were programmed as per the manufacturer’s instructions. The following primer sequences were synthesized by GeneCopoeia, with snRNAU6 (Homo sapiens) used as an internal control.
For mRNA
cDNA was synthesized using an iScript™ cDNA Synthesis Kit (Bio-Rad, Richmond, CA, USA), with the PCR conditions programmed as per the manufacturer’s instructions for the SYBR® Premix Ex Taq II (TAKARA, Dalian, Liaoning, China). The LYN primer sequence was 5′-TTGAAGACTCAACCAGTTCCAGA-3′, with the housekeeping gene GAPDH used as the internal control.
Transfection with miRNA constructs
In the miR 4323 group, cells were transfected with 100 nM mimics miR 4323 (GenePharma, Shanghai, China) using a Lipofectamine 3000 transfection reagent (Invitrogen) according to the manufacturer’s instructions. Cells in the control group were transfected with an equal concentration of a non-targeting control mimic sequence (GenePharma). The transfection efficiency was tested by flow cytometry and reached 80–90%.
Flow cytometry
After transfection for 48 h, cells were harvested and fixed in 75% cold ethanol overnight at 4℃. After centrifugation, the sediment was then dyed with propidium iodide. Cell cycle distribution was determined by flow cytometry using an EPICS® Elite ESP flow cytometer (Beckman Coulter, Miami, FL, USA).
Luciferase assay
Human LYN cDNA containing putative and mutant target sites for miR 4323 was chemically synthesized and inserted into a PsiCHECK-2Vector (Promega, Madison, WI, USA). Human embryonic kidney 293 cells (293T) were then transfected in 48-well plates with wild-type (psiCHECK-LYN) or mutant (psiCHECK-LYN-mut) reporter vectors along with mimics miR 4323 using Lipofectamine 3000 (Invitrogen) according to the manufacturer’s instructions. Luciferase activity was measured 48 h after the final transfection utilizing a Dual-Luciferase Assay kit (Promega).
Western blot analysis
After transfection, HBSMCs were lysed in a radioimmunoprecipitation assay buffer (Beyotime Institute of Technology, Shanghai, China). Proteins (30 µg) were separated by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to polyvinylidene fluoride membranes (Millipore®), which were incubated with antibodies to LYN, extracellular regulated protein kinases 1 and 2 (Erk1/2), phosphorylated Erk1/2 (p-Erk1/2), and β-actin (i.e. anti-lyn, anti-Erk1/2, anti-p-Erk1/2, and anti-β-actin antibodies; Abcam, Cambridge, MA, USA) at ratios of 1:500.
Statistical analysis 3000
The Statistical Product and Service Solutions 22.0 was used for data analysis. Data are expressed as means ± standard deviation and were analyzed by unpaired Student’s t-tests or analysis of variance, as appropriate. Values of p ≤ 0.05 were considered statistically significant.
Results
Hydrostatic pressure regulates RNA expression
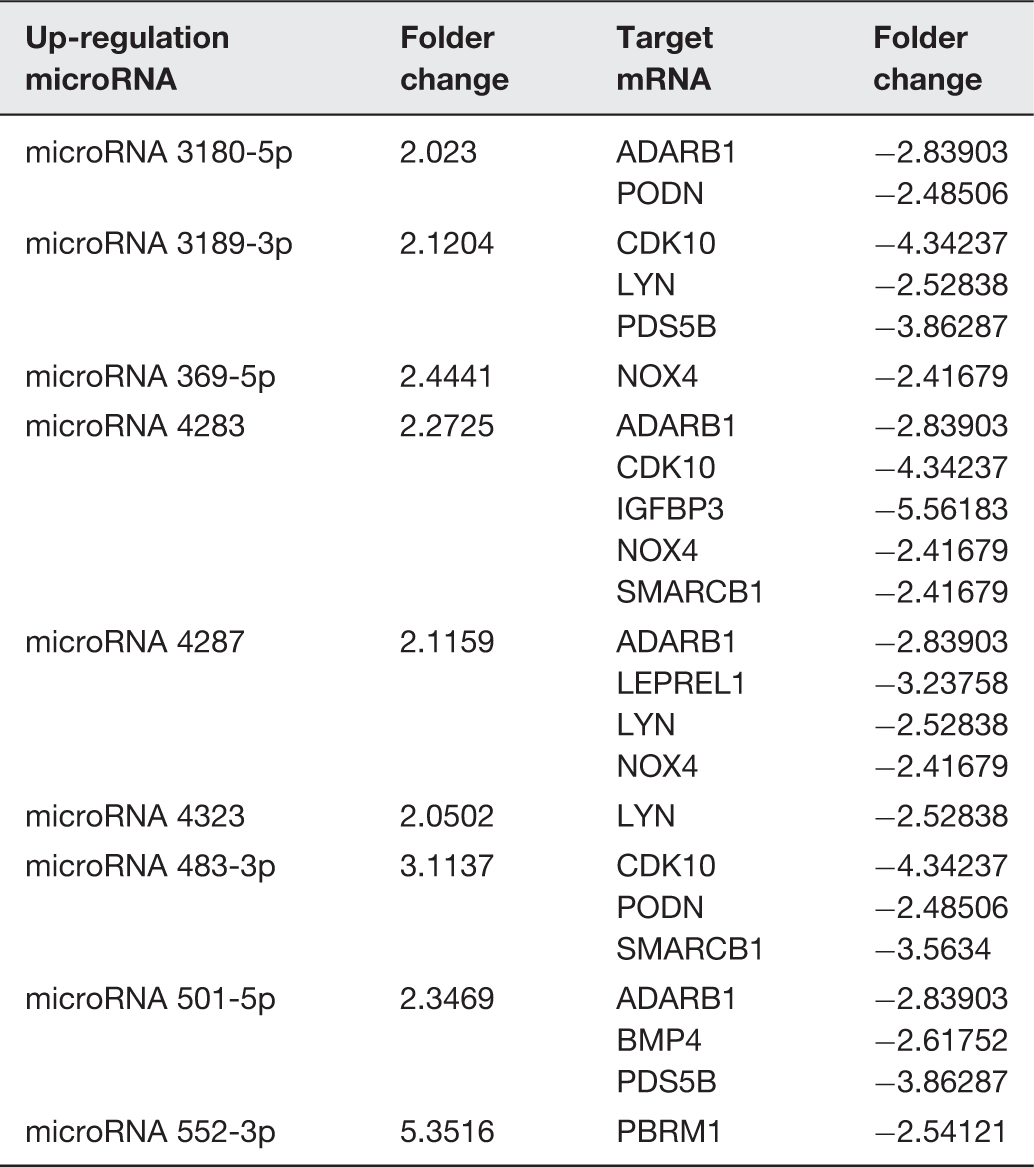
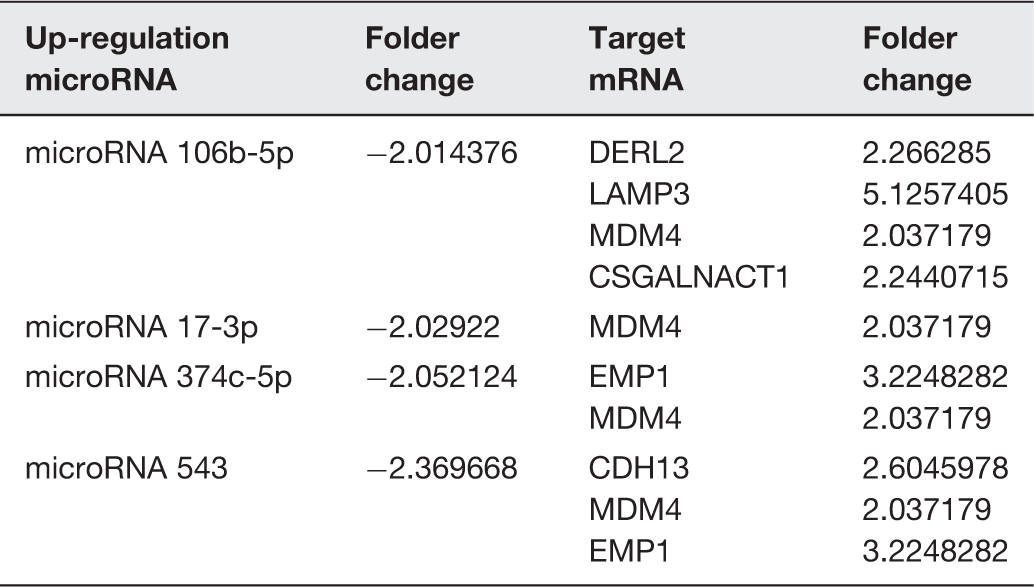
We used miRNA microarray assays to profile miRNA expression at baseline and after 24 h of exposure to hydrostatic pressure in HBSMCs. The differential expressions of specific miRNAs and mRNAs between the HP and control groups revealed upregulation of 103 miRNAs and 191 mRNAs in the HP group. Simultaneously, 60 miRNAs and 167 mRNAs were downregulated in the HP group (Figure 1). Target forecasting and functional analysis revealed nine upregulated miRNAs and four downregulated miRNAs related to cell proliferation. The upregulated miRNAs included miR 3180-5p, miR 3189-3p, miR 369-5p, miR 4283, miR 4287, miR 4323, miR 483-3p, miR 501-5p, and miR 552-3p (Table 1), while the downregulated miRNAs included miR 106b-5p, miR 17-3p, miR 374c-5p, and miR 543 (Table 2).
Hierarchical clustering of the microRNA and mRNA. The heat map diagram shows the result of the two-way hierarchical clustering of miRNAs and samples. Each row represents a miRNA, and each column represents a sample. The miRNA-clustering tree is shown on the left, and the sample-clustering tree appears at the top. The color scale shown at the top illustrates the relative expression level of a miRNA in the certain slide: red color represents a high relative expression level; green color represents a low relative expression levels. To analysis the different expression of microRNAs have with reference of Stem analysis, all of the changed microRNAs have the continue change trends in the Stem analysis: (a) up regulate microRNA, (b) down regulate microRNA, (c) increased mRNA and (d) decreased mRNA. (A color version of this figure is available in the online journal.) The up-regulation microRNA, target mRNA, and the folder change of RNAs The down-regulation microRNA, target mRNA, and the folder change of RNAs
Validation of the microarray findings by qRT-PCR
The 13 identified miRNAs were validated by qRT-PCR. The results confirmed that the expressions of miR 3180-5p and miR 4323 were significantly enhanced (p = 0.043 and p = 0.027, respectively; Figure 2), while that of miR 543 was significantly inhibited (p = 0.048; Figure 2). In addition, miRNAda target prediction indicated that LYN, a target of miR 4323, was involved in the Erk1/2 signaling pathway.
12
Furthermore, cyclic HP was able to regulate the Erk1/2 signaling pathway, as shown in our previous study.
2
In a parallel result, the expression of LYN was diminished by pressure stimulation (p = 0.032; Figure 2). Therefore, we tentatively propose that miR 4323 may be involved in HBSMC proliferation through the Erk1/2 pathway.
Different expressions of microRNAs and mRNAs were validated by PCR in HBSMCs between hydrostatic pressure (HP) and control static group. The black column means the HBSMCs were treated with 200 cm H2O, and the white column means the HBSMCs were treated with 0 cm H2O. The Y-axis means the relative fold expression, and the X-axis means the different mRNA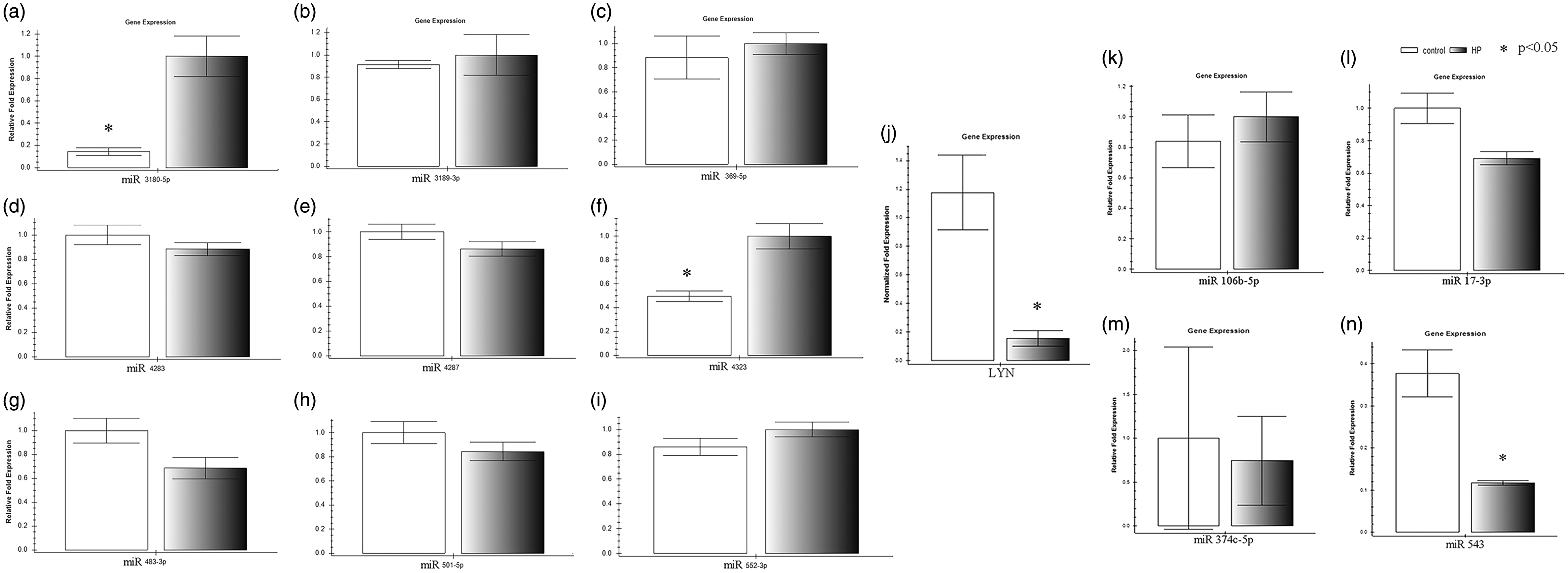
HBSMC proliferation was stimulated by miR 4323
Previously, we found that the cell proliferation index increased from 25.8% ± 7.1% under static conditions to 39.7% ± 4.2% under HP conditions,2,3 and hypothesized that miR 4323 was involved in this process. To confirm this, cells were treated with mimics miR 4323, and after transfection, miR 4323 expression was amplified by several orders of magnitude compared with the control group. So, we performed flow cytometry and calculated the cell proliferation index to demonstrate the cell proliferation rate, employing the formula: proliferation index (%) = [(S + G2/M)/(G0/G1 + S + G2/M)] × 100%. As shown in Figure 3, the proliferation of HBSMCs was enhanced by miR 4323 overexpression, and the proliferation index improved from 30.84 ± 4.57 in the control group to 52.13 ± 3.41 in the miR 4323 group (p = 0.001). These results indicate that miRNA 4323 was involved in the mechanism by which cyclic HP improved HBSMC proliferation.
The proliferations of HBSMCs after miR 4323 overexpressed. (a, b) The Y-axis means the cell numbers, and the X-axis means the DNA content. Different peak means different phase of cell cycle (G 0-1, S, and G 2). (c) The Y-axis means the proliferation index, and the X-axis means the different treatments. The yellow column means treated with NC, and the blue column means treated with mimics miR 4323. (A color version of this figure is available in the online journal.)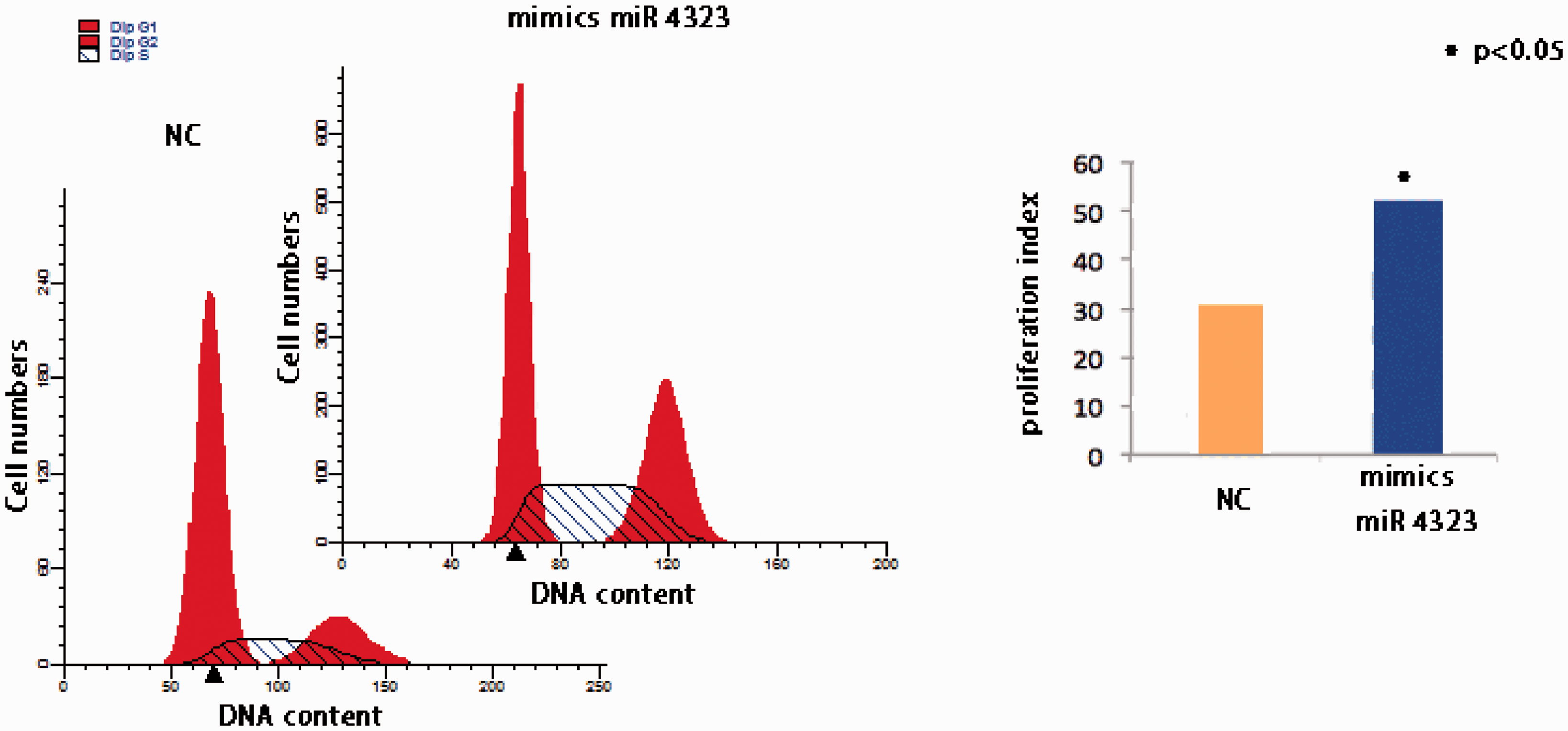
LYN was suppressed after transfection
Because miR 4323 appeared able to regulate HBSMC proliferation, we explored the possible mechanism of this process. The databases of computationally predicted human miRNA targets, miRNAda and TargetScan, indicated that LYN could be a potential target of miR 4323. To determine whether miR 4323 could regulate LYN expression, we performed PCR and Western blotting. After miR 4323 was over expressed by transfection with mimics into HBSMCs, expressions of LYN and lyn kinase (the functional protein of LYN) were downregulated and validated by qRT-PCR and Western blot.
Compared with the control group, LYN was significantly downregulated (p = 0.031), miR 4323 was upregulated (Figure 4(a)), and the level of lyn kinase was markedly suppressed (p = 0.037; Figure 4(b)) in the mimic-treated cells. These results show that miR 4323 inhibited LYN.
The miR 4323 directly regulates the LYN. (a, b) The expression of target mRNAs and function proteins belongs to miR 4323, which compared between the NC group and mimics miR 4323 group; (c) The luciferase activity in wild-type or mutant-type psiCHECK-LYN reporter vectors. 293T cells were co-transfected by psiCHECK-LYN (wild or mutant type) with mimics (mimics 4323 or non-targeting control mimic sequence also known as NC); the black column means that the 293T cells were co-transfected by NC, and the red column means the 293T cells were co-transfected by mimics 4323. (A color version of this figure is available in the online journal.)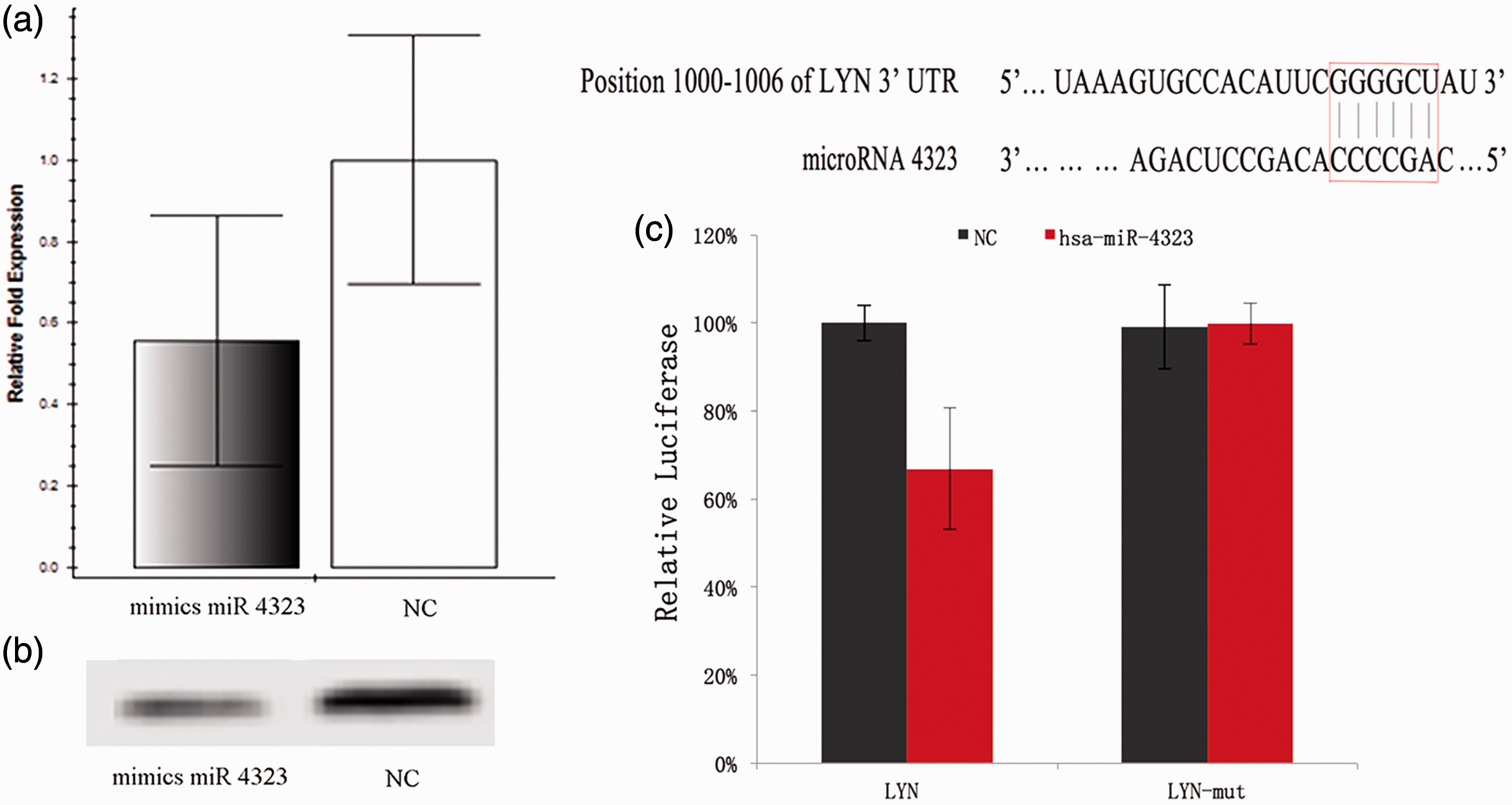
In addition, to observe the direct effect of miR 4323 on the 3′-UTR of LYN, we performed luciferase reporter assays with wild-type or mutant psiCHECK-LYN reporter vectors. We then co-transfected these vectors into 293T cells with either mimics miR 4323 or non-targeting control mimic sequences. As shown in Figure 4(c), the luciferase activities of 293T cells that co-transfected with wild-type psiCHECK-LYN and mimics miR 4323 were significantly lower than that of the control group (p = 0.016). Moreover, mutation of the binding site suppressed the effect of miR 4323 (p = 0.917). These results demonstrated that miR 4323 promoted HBSMC proliferation by regulating LYN.
The Erk1/2 signaling pathway was activated when lyn kinase decreased
Because previous research has confirmed that miR 4323 can regulate LYN, we examined whether miR 4323 was involved in the Erk1/2 signaling pathway by its action on LYN. To detect the change in phosphorylation of Erk1/2 induced by lyn kinase, we used post-transfection protein samples to perform Western blotting with anti-Erk1/2 and anti-p-Erk1/2. As shown in Figure 5, increased p-Erk1/2 may be associated with diminished lyn kinase expression (p = 0.004). Simultaneously, Erk1/2 expression did not differ between the two groups (p = 0.119). These results suggest that Erk1/2 activity was enhanced by the overexpression of miR 4323, and that this pathway regulated the proliferation of HBSMCs, as induced by hydrostatic pressure.
The expression of down stream proteins (erk1/2 singaling pathway)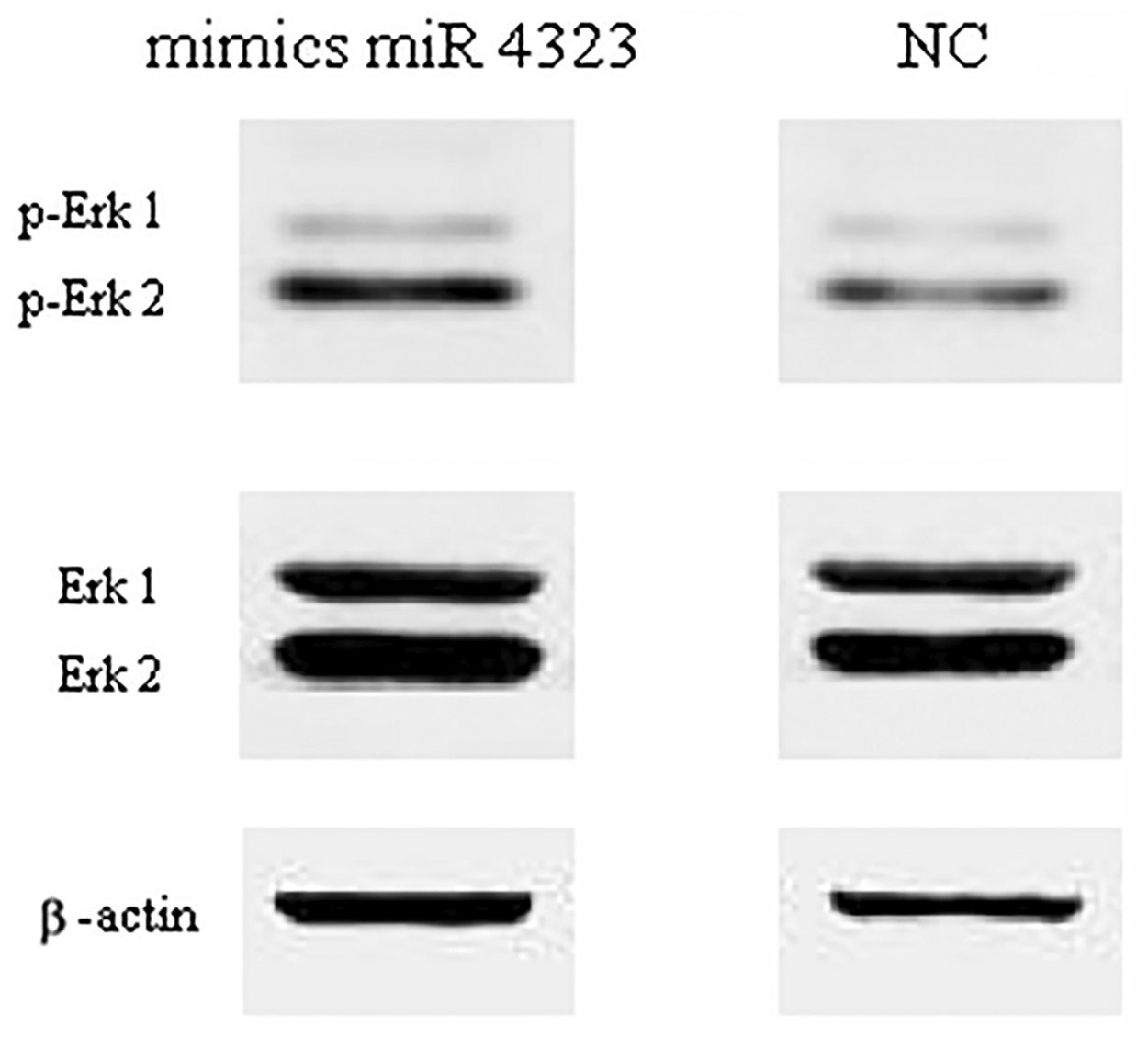
Discussion
In this study, miR 4323 was shown to promote HBSMC proliferation by targeting LYN and activating the Erk1/2 signaling pathway. The Erk1/2 pathway, also known as the MAPK pathway, is activated by numerous cytokines and is critical to the regulation of cellular proliferation and differentiation. 13 In addition, LYN is one of eight members of the src kinase family (SRC, YES, FGR, FYN, LCK, LYN, BLK, and HCK).14,15 Depending on the cellular milieu, this family regulates a variety of cellular functions, including cell proliferation, differentiation, and migration.14,15 Moreover, some authors have demonstrated that src kinases may induce cell proliferation 12 by limiting the duration and intensity of MAPK activation and by negatively regulating thrombopoietin expression, the latter of which is an essential hematopoietic cytokine that regulates megakaryocytopoiesis via activation of its cognate receptor, Mpl.16–18
According to a report by Brian et al., inhibition of lyn kinase resulted in enhanced thrombopoietin-induced proliferation in both hematopoietic cell lines and normal bone marrow cells, and that this physiologic effect may have been due to greater Erk1/2 phosphorylation and activity. 12 In our study, miR 4323 decreased lyn kinase expression and improved the Erk1/2 phosphorylation, it appeared to be involved in the process of HBSMC proliferation by the action of cyclic HP via the Erk1/2 pathway.
Other authors have shown that LYN can play both positive and negative roles in cellular processes, including proliferation, depending on the cell type and stimulus. 19 Similarly, Erk1/2 activation can regulate distinct aspects of cell cycle progression, including G1/S and G2/M transitions.20–22 In the present study, when HBSMCs were under the influence of cyclic HP, LYN acted as a negative regulator of proliferation, and increased the percentage of cells in S + G2/M, as shown by the flow cytometry results.
It is important to note that there are limitations to our study. Notably, this study was based on the overexpression of miR 4323 through transfection. This process can enhance the overexpression of miRNA for thousands times; but when HBSMCs were under cyclic HP conditions, it was only upregulated for two times. In our previous studies, similar to its effect on transfection, cyclic HP can also activate the Erk1/2 signaling pathway and can significantly increase cell proliferation.2–5 On the other word, we simulated the effect of cyclic HP through the transfection.
Further study is needed concerning the factors involved in miR 4323 regulation of HBSMC proliferation. As some authors have reported, long noncoding RNAs (lncRNAs) can inhibit or stimulate miRNA and regulate certain physiological processes.23,24 Although the repertoire of lncRNAs remains to be elucidated, a growing body of evidence suggests that they are important regulators of a diverse array of cellular function. 25 Thus, in future research, we need to describe the expression profile and functions of lncRNAs in HBSMCs.
Conclusion
In summary, miR 4323 expression was upregulated, and LYN was downregulated under cyclic HP conditions. After enhancing miR 4323 expression by the use of mimics, LYN expression was inhibited and lyn kinase expression decreased. In addition, the activation (phosphorylation) of Erk1/2 was promoted when LYN was downregulated by the overexpression of miR 4323, and the proliferation of HBSMCs was induced. Thus, our results indicate that miR 4323 is critical to the regulation of HBSMC proliferation by cyclic HP, and that the Erk1/2 signaling pathway is involved.
Footnotes
Authors’ contributions
Guarantor of integrity of entire study: YS, DL, YZ
Study concepts: YS, DL, YZ
Study design: YS, KW, YZ
Literature research: YS, DL, KW
Experimental studies: YS, YZ
Data analysis/interpretation: YS, KW
Statistical analysis: YS, KW
Manuscript preparation: YS, DL
Manuscript definition of intellectual content: YS, DL
Manuscript editing: YS, DL
Manuscript revision/review: KW
Manuscript final version approval: YS, DL, KW
YS and DL contributed equally to this work and should share the co-first author.
Acknowledgment
This work was supported by the National Science Foundation for Young Scholars of China (Grant No. 81300579, 81100494), the National Natural Science Foundation of China (Grant No. 81470927, 31370951, and 31170907), 135 Project of West China Hospital of Sichuan University, Technology Department of Chengdu City (No. 2014-HM01-00120-SF), Outstanding Youth Foundation of Sichuan University (2014SCU04B21), and Ministry of Organization of Sichuan (Grant No. JH2015017).
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.