
Abstract
MicroRNA-122 (miR-122) is liver specific and plays an important role in physiology as well as diseases including hepatocellular carcinoma (HCC). Downregulation of miR-122 in HCC modulates apoptosis. Similarly, the putative targets of miR-122, the forkhead box (FOX) family genes also play an important role in the regulation of apoptosis. Hence, an interplay between miR-122 and FOX family genes has been explored in this study. Initially, an augmentation of apoptosis was noticed in HepG2 cells after transfection with miR-122. Further, the predicted miR-122 targets, the FOX family genes (FOXM1b, FOXP1, and FOXO4) were selected via in silico analysis based on their role in apoptosis. We checked the expression of all these genes at transcript level after the transfection of miR-122 and found that the relative expression of FOXP1 and FOXM1b was significantly downregulated (p < 0.005) and that of FOXO4 was upregulated (p < 0.005). Thus, the finding indicates deregulation of these FOX genes as a result of miR-122 augmentation might be involved in the modulation of apoptosis.
Impact Statement
Here, we have investigated the crosstalk between microRNA-122 (miR-122) and selective FOX family genes in HepG2 cells. miR-122 is a prominent miRNA in liver and has been reported to be downregulated in hepatocellular carcinoma (HCC). It has been speculated that diminished level of miR-122 during HCC might be one of the reasons for tumor progression. However, the exact molecular interactions are not clear yet. This study unravels one of the molecular mechanisms of miR-122 through which it might impact the tumorigenesis of HCC.
Introduction
MicroRNA-122 (miR-122) is abundantly present in adult liver, representing 70% of the entire hepatic miRNAs. It is vital for normal liver development, function, and plays an essential role in various liver diseases including hepatocellular carcinoma (HCC). Downregulated miR-122 has been found in HCC tissue as well as several HCC cell lines and aberrant expression of the same is involved in the modulation of apoptosis, a prime player in tumorigenesis. 1 However, the mechanisms of apoptosis induced by miR-122 and their interaction with target genes are not well established in HepG2 cells.
Forkhead box (FOX) genes are a super family of evolutionarily conserved transcriptional regulators and are known to regulate diverse biological processes. Their role in tumorigenesis is well established in various cancers including HCC. 2 An aggressive malignant phenotype of HCC showed correlation with increased expression of FOXP1. 3 Further, Kalinichenko et al. 4 observed the antimetastatic potential of FOXM1b in HCC. Besides, several studies have also highlighted the miRNA–FOX gene interaction in various cancers. FOXC1 expression was found to be inversely correlated to miR-204 in endometrial carcinoma. 5 Additionally, FOXG1, FOXQ1, and FOXE1 were proved to be negative regulators of miR-422 a in HCC. 6 In this study, we have investigated the consequence of miR-122 interaction with FOX family genes involved in the modulation of apoptosis in HepG2 cells.
Materials and methods
The human hepatoblastoma cell line HepG2 was purchased from National Centre for Cell Science, Pune, India. Cells were maintained in minimum essential medium supplemented with 10% fetal bovine serum (Sigma, USA), 100 U/mL penicillin, and 100 µg/mL streptomycin (Hi-media, India) at 37℃ in a humidified atmosphere of 5% CO2 conditions. The miR-122 expression vector miRNASelect™ pEGP-mmu-miR-122 (Cell biolabs, USA), containing the precursor sequence of miR-122 was used for its stable expression along with Green Fluorescent Protein (GFP)-encoding gene. Cells were transfected with miR-122 expression vector using Lipofectamine LTX and Plus reagent (Invitrogen, USA) according to manufacturers’ instructions. Cells transfected with vector without miR-122 construct (pIRES, Clontech, USA) were used as control.
After transfection, expression of miR-122 vector was confirmed by fluorescence microscope (Olympus, CKX41, Japan) and quantitatively by flow cytometry using FACS Canto flow cytometer (CA, USA). Total RNA was isolated from the miR-122 vector-transfected cells at 48 h post-transfection using Trizol reagent (Invitrogen, USA). The miRNAs were polyadenylated and reverse transcribed using the miRNA first strand cDNA synthesis kit (Stratagene, USA) according to manufacturers’ instructions. cDNA template was used for real-time quantification of miR-122. 7 The U6 snRNA was taken as an internal control. The relative expression of miRNA-122 in transfected cells was determined in terms of their fold change by using the formula (2−ΔΔCT). The quantitative Real Time Polymerase Chain Reaction (qRT-PCR) data were expressed as the mean ± SD from each experiment performed in triplicates.
Analysis of apoptosis induced by miR-122 transfection was examined by Acridine orange (AO) and Ethidium bromide (EtBr) 8 staining followed by flow cytometry and fluorescence microscopy, respectively. Further, apoptosis was also analyzed at different time intervals by using Alexa Fluor 488 annexin V/Dead Cell Apoptosis Kit (Invitrogen). 9
In silico analysis was performed for the search of predicted targets of miR-122 using five databases—miRanda (www.microrna.org), miRDB (www.mirdb.org), TargetScan (httpwww.targetscan.org), PicTar (http://pictar.mdc-berlin.de/), miRWALK (http://www.umm.uni-heidelberg.de/apps/zmf/mirwalk/).
Total RNA was isolated from the cells at 24, 48, and 72 h post-transfection with miR-122. Expression of FOXM1B, FOXP1, and FOXO4 was quantified by real-time PCR with reaction conditions 95℃ for 10 s, 60℃ for 15 s, and 72℃ for 20 s. Each PCR reaction was performed in triplicates with GAPDH as an endogenous control. Fold difference in mRNA expression was calculated in transfected and non-transfected cells using 2−ΔΔCT method. 10
The statistical calculations were performed using Statistical Package for the Social Sciences (SPSS) for windows, version 16 (SPSS Inc., Chicago, USA). The statistical significance between transfected and control groups was analyzed using the Student’s t-test. The difference was considered significant if the p value is less than 0.05.
Results
HepG2 cells were transfected with miR-122 vector and analyzed by fluorescence microscopy, flow cytometry as well as qRT-PCR for miR-122 expression. miR-122 construct containing the GFP-encoding gene was detected under fluorescence microscope showing green fluorescence compared to control (Figure 1(a)). FACS analysis revealed 5.1, 11.2, and 28.6% population of HepG2 cells showed the GFP expression after 24, 48, and 72 h transfection (Figure 1(b)). Further, miR-122 expression was found to be 20-fold higher in transfected cells by qRT-PCR analysis (Figure 1(c)). These results depict the successful transfection and expression of miR-122 in HepG2 cells.
miR-122 transfection into HepG2 cells. (a) Fluorescence microscopy in control (i), and miR-122 transfected cells at 48 h (ii). (b) Flow cytometry analysis in control (i), and miR-122-transfected HepG2 cells at 24 h (ii), 48 h (iii), 72 h (iv). (c) miR-122 quantification in control, mock control (vector without miR-122), and miR-122-transfected cells by quantitative Real Time Polymerase Chain Reaction (qRT-PCR) at 48 h (***p < 0.001). (A color version of this figure is available in the online journal.)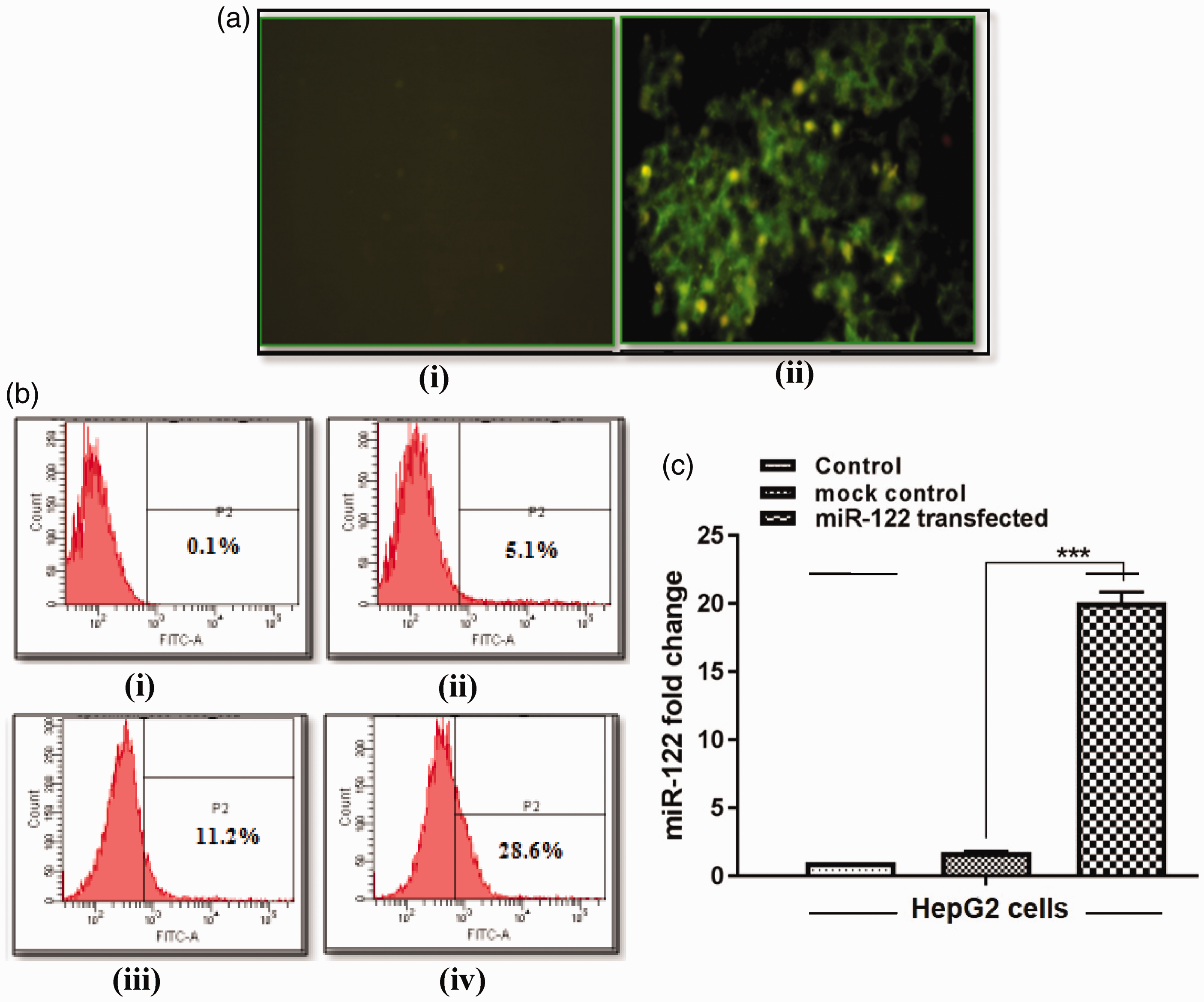
To characterize apoptosis due to miR-122 induction, HepG2 cells were stained with AO (green fluorescence)/EtBr (red fluorescence) and examined by fluorescence microscope at 72 h. Three types of cell populations were observed, which included viable cells (V) with bright green appearance, apoptotic cells (A) yellow/orange, and necrotic cells (N) red (Figure 2(a)). Apoptotic index (A.I.) was found to be significantly higher (A.I. = 65%) in miR-122-transfected HepG2 cells as compared to the control (A.I. = 22%) suggesting that miR-122 may induce apoptotic cell death (Figure 2(b)).
Apoptosis analysis of miR-122-transfected HepG2 cells. (a) AO/EtBr staining of control (i), and miR-122-transfected cells (ii), at 72 h post-transfection. A: apoptotic cells; N: necrotic cells; V: viable cells. (b) Apoptotic index (%) calculation at 72 h post-transfection (A.I. = apoptotic cells × 100/total number of cells) (*p < 0.05). (c) Annexin/PI staining in control (i), and miR-122-transfected cells (ii), at 24, 48, and 72 h. A.I.: apoptotic index; AO: acridine orange; EtBr: ethidium bromide; PI: Propidium Iodide. (A color version of this figure is available in the online journal.)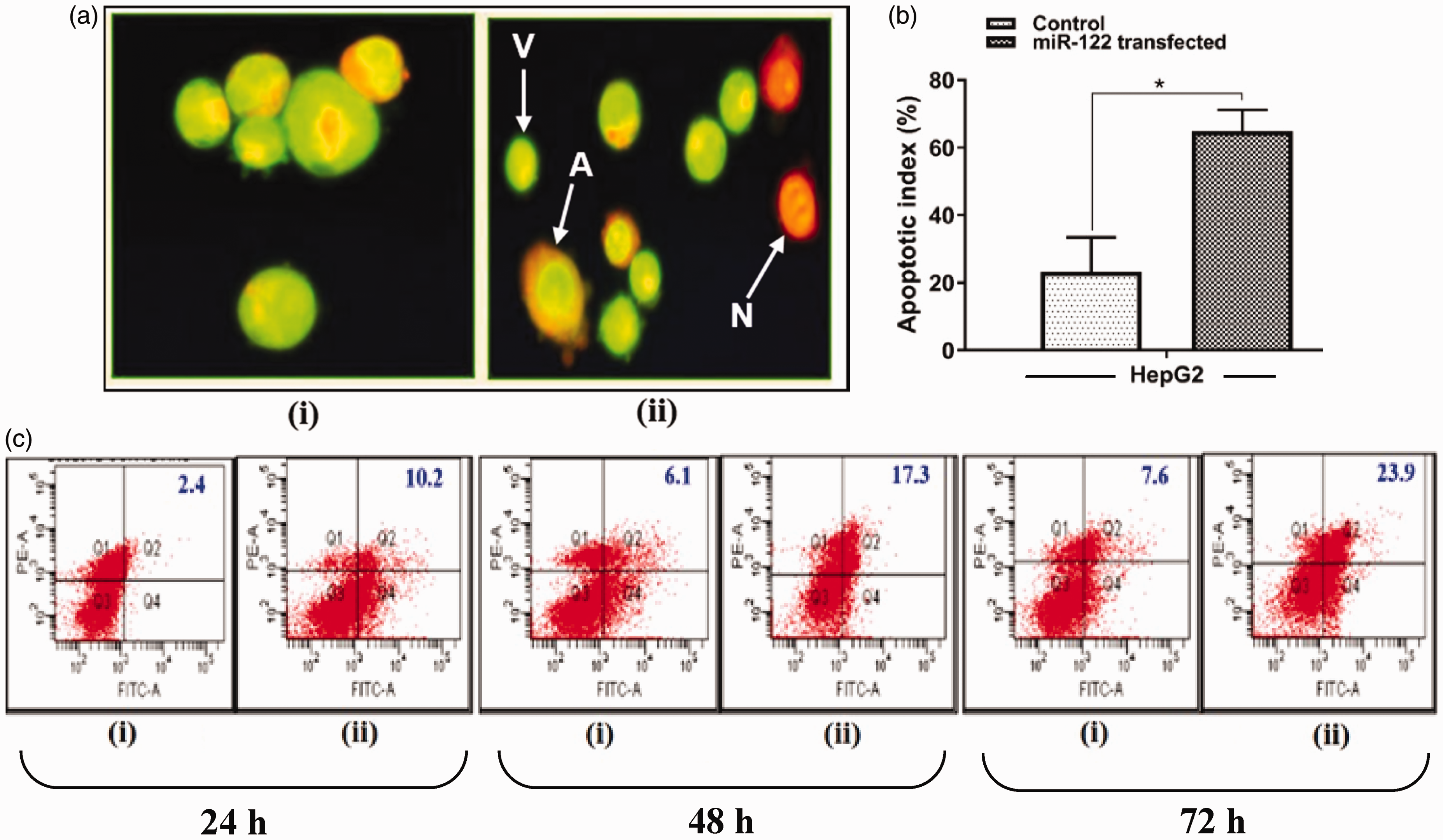
To confirm the phenomenon of apoptotic cell death, miR-122-transfected HepG2 cells were further stained with annexin V at different time intervals. The populations of apoptotic cells were found to be increased linearly (10.2, 17.3, and 23.9%) as compared to respective control (2.4, 6.1, and 7.6%) at 24, 48, and 72 h (Figure 2(c)). Overall, apoptotic rate was found to be higher in transfected cells as compared to non-transfected control.
Several algorithms were used to search for putative targets of miR-122. FOX family members were markedly enriched to have putative miR-122-binding elements in their 3 terminal untranslated regions (3′-UTRs); in particular, FOXP1, FOXO4, and FOXM1b had high fidelity scores using five databases.
Expression of these target genes was quantified in miR-122-transfected cells at 24, 48, and 72 h by qRT-PCR. HepG2 cells showed insignificant change (p > 0.05) in the expression of FOXM1b after 24 h of transfection; however, a significant downregulation was observed after 48 h (2.1-fold) and 72 h (1.5-fold) (p < 0.05) (Figure 3(a)). Similarly, significant downregulation in the expression of FOXP1 was observed after 24 h (2.43-fold), 48 h (1.97-fold), and 72 h (1.62-fold) of transfection (p < 0.05) (Figure 3(b)). In contrast, FOXO4 showed a significant upregulation after 24 h (2.58-fold), 48 h (1.41-fold), and 72 h (1.26-fold) of transfection (p < 0.05) (Figure 3(c)).
Fox family gene expression analysis in miR-122-transfected HepG2 cells at 24, 48, and 72 h by qRT-PCR. (a) FOXM1b, (b) FOXP1, and (c) FOXO4. (A color version of this figure is available in the online journal.)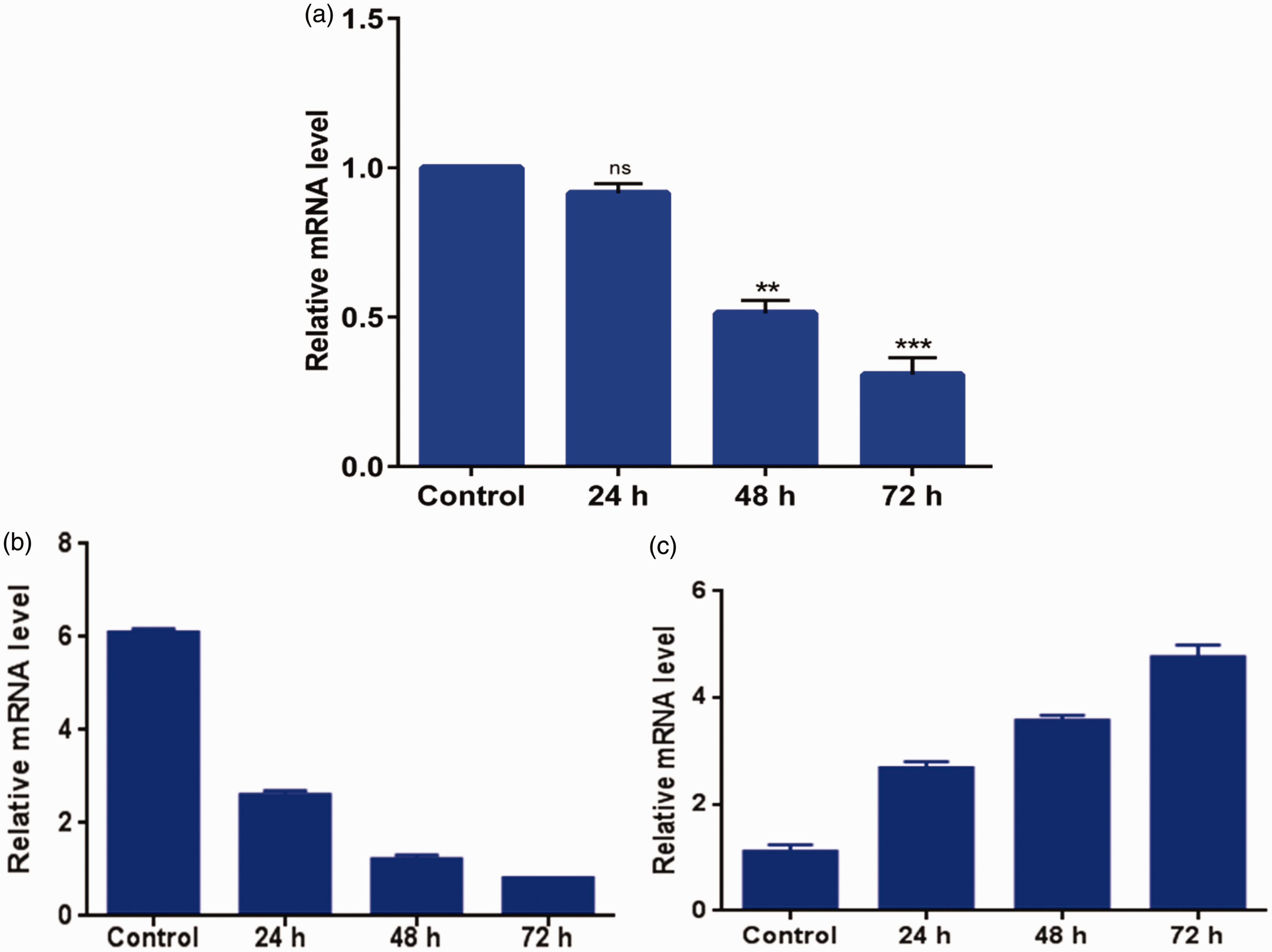
Discussion
Biological rationale behind reduced expression of miR-122 in HCC is not clear. However, it has been speculated that diminished level of miR-122 during HCC might be one of the reasons for tumor progression by altering the expression of its target genes.Our study has shed light on how miR-122 modulation regulates apoptosis via targeting Fox family genes in hepatoblastma cell line HepG2.
This study for the first time unraveled the crosstalk between miR-122 and Fox family genes. Here we report that augmentation of miR-122 leads to downregulation of FOXM1b as well as FOXP1 and upregulation of FOXO4. Among the predicted targets of miR-122, FOXM1b is a proliferation-specific transcription factor ubiquitously expressed in embryonic tissues, cultured cells, and malignant tumors. 4 It is considered as a crucial proto-oncogene in HCC progression and is also involved in apoptosis. 11 Further, FOXP1 acts as both tumor suppressor and oncogene in a tissue-specific manner. It has been described as an oncogene in HCC. 4 Additionally, FOXP1 was found to promote inhibition of caspase-dependent apoptosis in B cell lymphoma. 12 Moreover, FOXO4 is known to act as a tumor suppressor in various cancers; however, its role in HCC has not been reported. 13 Urbich et al. 14 reported that FOXO4 induces apoptosis via upregulation of Bim in the mitochondria-dependent pathway in clear cell renal cell carcinoma. Van Boxtel et al. 15 reported that FOXO activation in FOXP1-knockdown cells resulted in increased apoptosis. Even though both FOXO4 and FOXP1 are direct targets of miR-122, it is also possible that decreased expression of FOXP1 might further aid in the activation of FOXO4 as observed in our study resulting in increased apoptosis.
In conclusion, our results demonstrated that augmentation of miR-122 induces apoptosis by modulating Fox family target genes at various levels to exert its antitumor effects in HCC. This induction might involve intricate interactions with Fox family genes which needs further investigation.
Authors’ contributions
This work is part of PhD thesis work of SuK. AB and ShK performed FOX expression analysis and participated in writing the manuscript. SG participated in interpretation of studies and review of manuscript. AC participated in design, interpretation, and analysis of results and review of manuscript.
Footnotes
Acknowledgements
We acknowledge the Indian Council of Medical Research (ICMR), New Delhi, for providing Research Fellowship to Mr Subodh Kumar, Mrs Ankita Batra and Mrs Shruthi Kanthaje.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.