
Abstract
Electromagnetic field at extremely low frequencies plays a significant role in the physiological function of human tissues and systems. It is shown that electromagnetic field inhibits production of reactive oxygen species which are involved in heart injury triggered by oxidative stress. We hypothesize that low frequency electromagnetic field protects function of cardiac cells from ischemia–reperfusion injury. Human cardiac myocytes, endothelial cells, and cardiac fibroblast underwent ischemia–reperfusion conditions in the presence or in the absence of low frequency electromagnetic field. LDH and MMP-2 activities (as markers of cell injury), and cell metabolic activity (by fluorescein diacetate staining) were measured to determine the protective role of low frequency electromagnetic field. Our data showed that short courses of low frequency electromagnetic field protect cardiac cells from cellular damage and preserve their metabolic activity during ischemia–reperfusion. This study demonstrates the possibility to use of low frequency electromagnetic field as strategy for the prevention or therapy of ischemia–reperfusion injury.
Impact statement
In our study, we showed that LF-EMF may be protective for heart during ischemia–reperfusion (I/R). Following is the short description of the main findings: (a) the response to the I/R injury was different for endothelial cells, fibroblasts, and cardiomyocytes; (b) I/R decreases MMP-2 activity in cardiac myocytes and fibroblasts; (c) I/R increases MMP-2 activity in endothelial cells; (d) LF-EMF reverses these changes; (e) LF-EMF protects cells from injury and preserves their metabolic activity.
Introduction
Generation of reactive oxygen species (ROS) in the area of hypoxia and reoxygenation is a fundamental process underlying heart triggered by myocardial infarction (MI) or ischemia/reperfusion (I/R). Since low level of ROS regulates physiological cardiovascular function, an increased production of superoxide (O2−), hydrogen peroxide (H2O2), and peroxynitrite (ONOO−) affects function of numerous cells and tissues. Disruption of the oxidant-antioxidant balance leads to decreased contractility of cardiomyocytes, 1 apoptosis of endothelial and vascular smooth muscle cells, 2 and remodeling of the failing myocardium. 3 In the heart, toxic oxygen radicals can be generated by endothelium, fibroblasts, and cardiomyocytes leading to peroxidation of membrane lipids, denaturation of proteins and enzymes, damage of nucleic acids and cells. 4
It is known that increased activity of matrix metalloproteinase (MMP) may be responsible for the degradation of extracellular 5 and intracellular cardiac proteins6–8 or ventricular remodeling, 9 as a consequence of ischemia and reperfusion. Moreover, an increased activity of gelatinases in arteries plays a role in arterial tissue remodeling after injury due to activation of vascular smooth muscle and neointima formation. 10 Because of the role of MMPs in pathological processes, MMP-2 and MMP-9 are considered as a good markers of cell injury. This is why numerous preventive and therapeutic strategies targeting MMPs were undertaken to restrain remodeling.11–13
Cardiac muscle is composed of several types of cells which play different roles in the heart contractile function. This is why several distinct mechanisms of heart injury should be targeted to prevent heart contractility. Besides, since MMP implicated in the disease process are also essential for physiological functions, 14 an inhibition of broad spectrum of MMP with nonselective pharmacological inhibitors may be a source of adverse side-effects.
Taking into account that ROS is one of the most emphasized factors of cells’ injury during the lack of oxygen supply or following reoxygenation, magnetotherapy has been considered in a term of prevention. Low frequency electromagnetic field (LF-EMF) has been indicated as a regulator of biological functions including reduction/induction of inflammatory process,15,16 regulation of cell differentiation,15,17 and gene expression. 17 We previously showed that LF-EMF protects mechanical function of hearts subjected to ischemia and reperfusion. 18 We also showed that LF-EMF reduced heart injury by maintaining the integrity of the endothelium. 18
In the current study, an influence of short-term exposure of main cardiac muscle cells undergoing in vitro chemical ischemia and reperfusion (I/R) to LF-EMF was tested. The main aim was to find the contribution of each cells type in the mechanism of heart injury. We also hypothesized that electromagnetic field protects the metabolic function of different types of heart cells; hence, it may be used as a therapeutic agent in the prevention or treatment of heart injury during ischemia and reperfusion.
Materials and methods
Cell culturing
The primary cells of human cardiac myocytes (HCM, cat. no. 6200), human aortic endothelial cells (HAECs, cat. no. 6100), and human cardiac fibroblasts (HCFs, cat. no. 6300), all from ScienCell Research Laboratories (Carlsbad, CA, USA) were cultured in Dulbecco's Modified Eagle's Medium containing: cardiac myocyte growth supplement (cat. no. SC6252), endothelial cell growth supplement (cat. no. SC1052) or fibroblast growth supplement-2 (cat. no. SC2382), respectively, supplemented with 5% of fetal bovine serum, 100 U/mL of penicillin, 100 µg/mL of streptomycin (all from Sigma-Aldrich, St. Louis, MO, USA). Each cell line was cultured at 37°C in a water-saturated, 5% CO2 atmosphere. Cells were passaged at 80% confluence by harvesting with 0.25% trypsin-EDTA (Sigma-Aldrich).
Protocol for in vitro chemical ischemia and reperfusion
HCM, HAECs, and HCFs underwent in vitro chemical ischemia–reperfusion in flat petri dishes 35 × 10 mm (Greiner, Kremsmünster, Austria). In brief, cells were submitted to 9 min in vitro chemical ischemia (I) and 20 min aerobic reperfusion (R) after 15 min of aerobic stabilization. Cells were subjected to aerobic stabilization, and reperfusion in HEPES buffer (5.5 mmol/L HEPES, 63.7 mmol/L CaCl2, 5 mmol/L KCl, 2.1 mmol/L MgCl2, 5.5 mmol/L glucose, 10 mmol/L taurine) containing additional 55 µmol/L CaCl2 and 0.75 mg/mL BSA and ischemia was obtained with chemical ischemia buffer consisting of HEPES buffer containing 4.4 mmol/L 2-deoxyglucose and 4.0 mmol/L sodium cyanide (an inhibitor of electron transport chain). The optimal duration of in vitro chemical ischemia, 9 min, was established in preliminary study on cardiomyocytes and confirmed in endothelial cells and fibroblasts (data not shown). After 15 min of aerobic conditions, the cells were centrifuged (1 min 1500 × g) and the pellet was resuspended in the chemical ischemia buffer for 9 min. Then the buffer was removed (centrifugation 1 min 1500 × g) and the cells pellet was resuspended in the fresh portion of HEPES buffer containing additional 55 µmol/L CaCl2 and 0.75 mg/mL BSA. After reperfusion (R), the cells were centrifuged at 1500 × g for 5 min and the cells pellet was homogenized before further analysis. The aerobic control group was exposed to atmospheric conditions for 44 min, and the chemical ischemia group with LF-EMF underwent the same experimental protocol but with the presence of LF-EMF during incubation steps (please see the protocol shown in Figure 1(a).
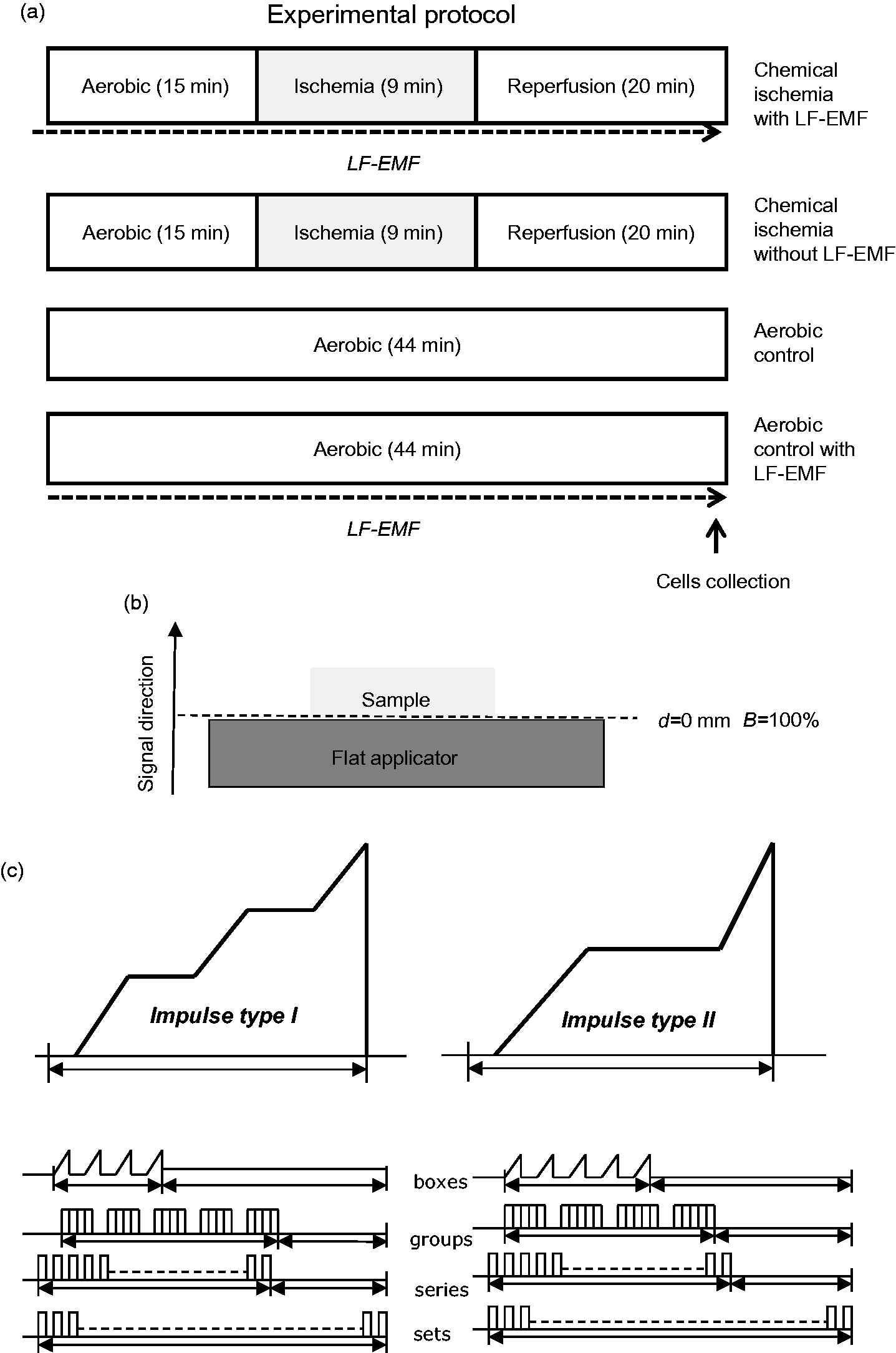
Experimental protocol for in vitro chemical ischemia and aerobic control with or without LF-EMF (a). Diagram showing the estimated values of electromagnetic field (B) in dependence form the distance (d) between the applicator head and the sample (b). The distance from the inducing flat applicator to the cells (d) was 0 mm, hence the estimated value of the electromagnetic field (B) induction was 100%. The structure of magnetostimulation signal in Viofor JPS system (c). Isolated cells were subjected to LF-EMF for all time of experiment. LF-EMF: low frequency electromagnetic field.
LF-EMF was generated by a flat applicator (FA) connected to a Viofor JPS control unit (Classic, Med & Life, Komorów, Poland) with the frequency 50–60 Hz and maximum application current at the intensity of 12 when magnetostimulation was 2A. The distance from the inducing FA to the cells in the flat petri dishes (Greiner, Kremsmünster, Austria) placed centrally on the applicator (d) was 0 mm, hence the estimated value of the electromagnetic induction (B) in the relation to the distance was 100% (4560 µT) (induction at value “12”) (Figure 1(b)). Viofor JPS control unit with connected FA generates an impulse magnetic field consisting of two types of impulses, which in the rising part have linear acceleration lines with varying inclination. The impulses are grouped hierarchically and form a spectrum with the following characteristic frequencies: basic pulses in packets: 180–195 Hz, packets-12.5–29 Hz, groups of 2.8–7.6 Hz, series: 0.08–0.3 Hz (Figure 1(c)). This signal forms a multi-peak frequency spectrum from 0.08 Hz to 195 Hz and harmonics.
Cells homogenization
After 44 min of the experimental protocol, cells were degraded by three cycles of freezing (in liquid nitrogen) and thawing (at 37°C) and homogenized mechanically (three times for 10 s) on ice in 50 mmol/L Tris-HCl (pH 7.4), containing 3.1 mmol/L sucrose, 1 mmol/L DTT, 10 µg/mL leupeptin, 10 µg/mL soybean trypsin inhibitor, 2 µg/mL aprotinin, and 0.1% Triton X-100. Homogenates were centrifuged at 10,000 × g, for 5 min at 4°C, and the supernatant was immediately transferred into a fresh tube and stored at −80°C for further biochemical analysis.
Determination of intracellular MMP-2 activity
MMP-2 activity was assessed by gelatin zymography, previously described by Heussen and Dowdle, 19 with modifications. Prior to electrophoresis, protein content in homogenates was analyzed using Bradford Protein Assay (Bio-Rad Laboratories, Munich, Germany) and bovine serum albumin (heat shock fraction, ≥ 98%, Sigma-Aldrich) was used as a protein standard.
Briefly, supernatants of cells homogenates were adjusted to contain the same protein concentration with distilled water and mixed with a proper amount of 4× Laemmli Sample Buffer (Bio-Rad Laboratories, Hercules, CA, USA). Samples containing 20 µg of protein were loaded in each lane of 8% polyacrylamide gel copolymerized with gelatin (2 mg/mL) and 0.1% SDS (denaturing but non-reducing conditions). After electrophoresis, gels were washed in 2.5% Triton X-100 (3 × 20 min). Subsequently, gels were placed in the incubation buffer (50 mmol/L Tris–HCl pH 7.5; 5 mmol/L CaCl2; 200 mmol/L NaCl; 0.05% NaN3) and incubated at 37°C for 18 h. After incubation, gels were stained with the staining solution (0.5% Coomassie Brilliant Blue R-250; 30% methanol; 10% acetic acid) for 2 h and destained with the destaining solution (30% methanol; 10% acetic acid) until bands were clearly visible. The activity of MMP-2 was visualized as bright bands against a dark background. Gels were scanned using GS-800 Calibrated Densitometer (model PowerLook 2100 XL-USB) with Quantity One v. 4.6.9 software (Bio-Rad Laboratories) and the relative MMP activity was determined and expressed in arbitrary units (AUs) as activity per microgram of total protein in a sample.
Assessment of LDH activity
Lactate dehydrogenase activity assay kit (Sigma-Aldrich) was used to determine the activity of LDH in tested cells, following the manufacturer’s instructions. Briefly, lactate dehydrogenase interconverts pyruvate and lactate with the reduction of NAD to NADH, which is detected with a colorimetric assay at 450 nm. Lactate dehydrogenase served as a marker of cell damage.
Determination of metabolic activity of human cardiomyocytes, endothelial cells, and fibroblasts
Fluorescein diacetate (FDA) was used to estimate the metabolic activity of cells subjected to in vitro chemical ischemia and reperfusion. FDA is accumulated in cells if they are not injured and if appropriate esterases are present in an active form. The damage of cell membrane or enzymatic apparatus leads to a rapid diffusion of esterified fluorescein from cell to liquid medium, not allowing cell visualization. To determine the metabolic activity of cells, the simultaneous double-staining procedure with FDA (Sigma-Aldrich) and DAPI (4′,6-diamidino-2-phenylindole, Sigma-Aldrich) was used. Briefly, the cells were cultured on to 80% confluence, washed three times with PBS and stained with 1 µmol/L FDA for 15 min, 37°C in a dark and humid chamber. DAPI was used to stain cells’ nuclei. Nikon Eclipse E600 fluorescence microscope (Nikon, Tokyo, Minato, Japan) was used to estimate the bright green fluorescent metabolically active cells.
Statistical analysis
Data were extrapolated from six separate experiments. Analysis was based on Statistica software v. 12 (Statsoft Polska, Warsaw, Poland). The test for comparison of two means (normal distribution) or Mann–Whitney test was used in this study as appropriate. P < 0.05 indicated statistical significance. Data are presented as the mean ± S.E.M.
Results
Effect of LF-EMF on cell injury
In order to determine the effect of I/R and LF-EMF on severity of cell damage, the intracellular activity of LDH was measured. Results showed that endothelial cells and fibroblasts were affected by I/R, whereas myocytes were not (Figure 2).
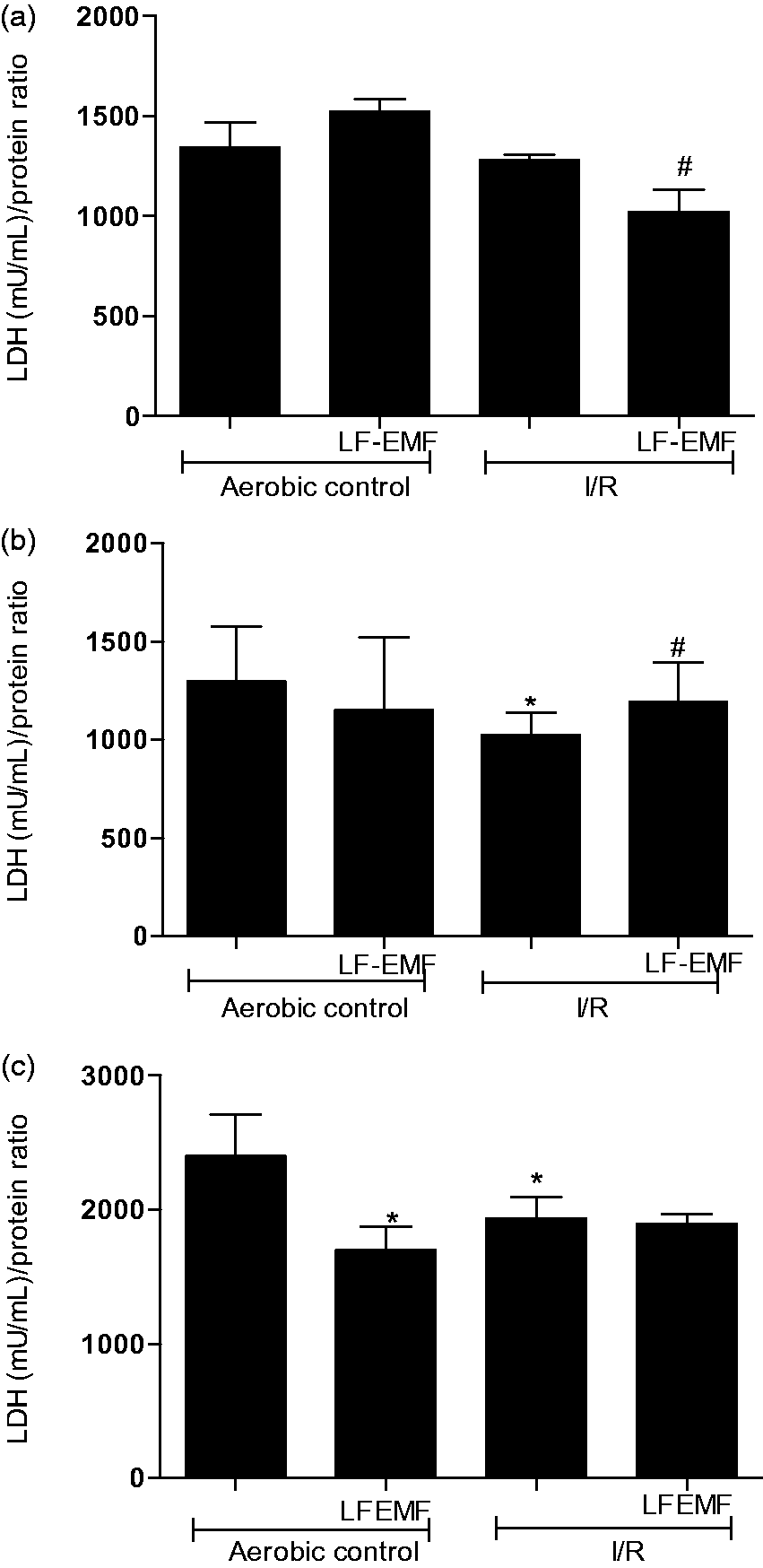
Effect of LF-EMF on injury of myocytes (a), endothelial cells (b), and fibroblasts (c) by measurement of LDH in cells homogenates. n = 6 per group; *P < 0.5 vs. Aero; #P < 0.5 vs. I/R. I/R: in vitro chemical ischemia-reoxygenation; LF-EMF: low frequency electromagnetic field; mU/mL: milli international enzyme activity unit per milliliter.
The activity of LDH in endothelial cells and fibroblasts subjected to 9 min in vitro chemical ischemia and 20 min of reperfusion was significantly lower than in aerobic control (Figure 2(b) and (c)). Exposure of these cells to LF-EMF during I/R reduced the damage of endothelial cells (Figure 2(b)), but not fibroblasts (Figure 2(c)). In contrary, LF-EMF application led to further damage of cardiomyocytes subjected to I/R (Figure 2(a)).
Effect of LF-EMF on MMP-2 activity in cardiac cells subjected to I/R injury
Cardiac myocytes, endothelial cells, and fibroblasts were subjected to I/R in the presence or absence of LF-EMF. I/R led to a significant decrease of MMP-2 activity in myocytes (more than 30%) and fibroblasts (more than 40%) in comparison to cells undergoing aerobic conditions (Figure 3(a) and (c)). In contrast, the activity of endothelial MMP-2 was increased approximately by 20% (Figure 3(b)). Level of MMP-2 activity in cells exposed to LF-EMF was similar to aerobic control level (Figure 3).
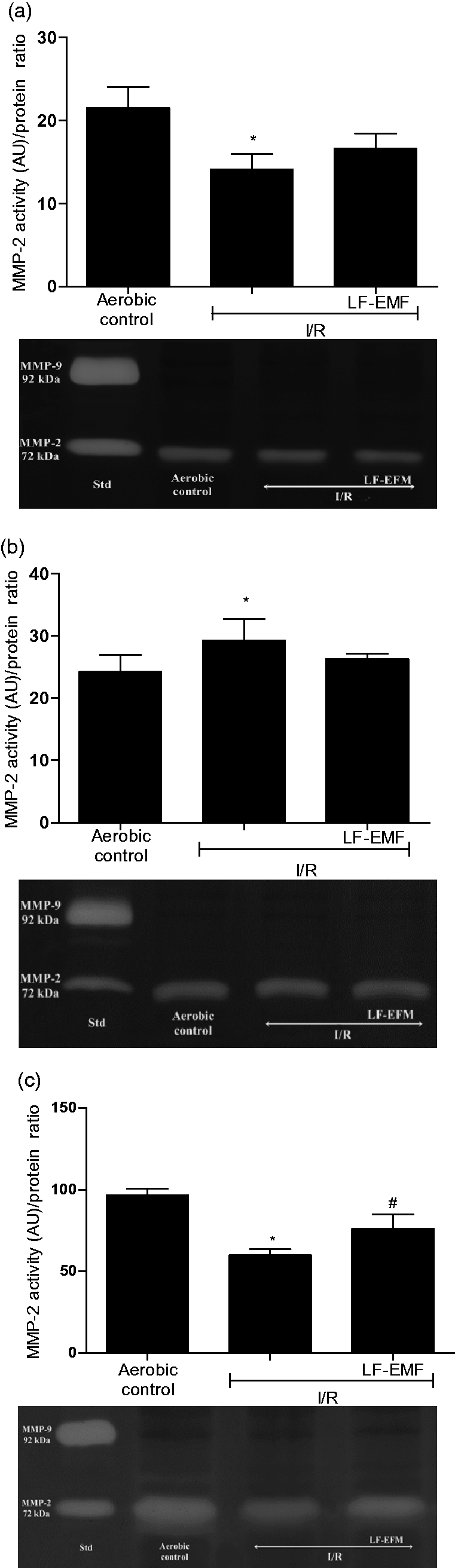
An influence of LF-EMF on MMP-2 activity in cardiomyocytes (a), endothelial cells (b), and cardiac fibroblasts (c) subjected to I/R. n = 6 per group; *P < 0.5 vs. Aero; #P < 0.5 vs. I/R. I/R: in vitro chemical ischemia-reperfusion; LF-EMF: low frequency electromagnetic field; MMP-2: matrix metalloproteinase 2. Figures and zymograms.
Effect of LF-EMF on metabolic activity of cardiac cells subjected to I/R injury
I/R led to decreased metabolic activity in cardiac myocytes, endothelial cells, and fibroblasts (Figure 4(a) to (l)). Exposure to LF-EMF protected metabolic activity of cells in comparison to highly reduced metabolic activity of cardiac cells subjected to I/R without LF-EMF (Figure 4(a) to (l)).
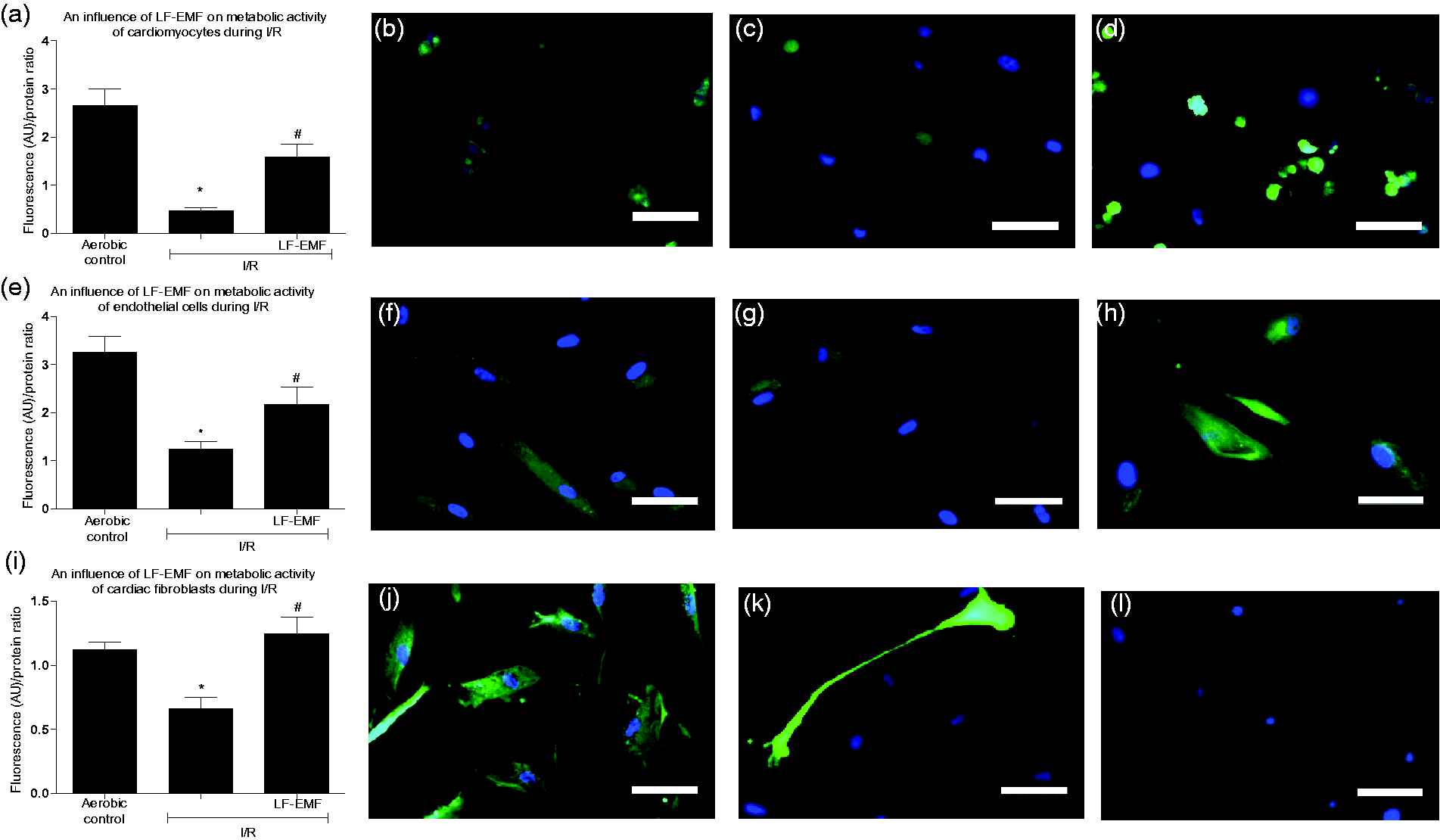
An influence of LF-EMF on metabolic activity of cardiomyocytes (a), endothelial cells (e), and cardiac fibroblasts (i) subjected to I/R. Panels b-d (cardiomyocytes), f-h (endothelial cells), j-l (cardiac fibroblasts) show representative images of appropriate cells group. Graph bars show the average of total fluorescence of cells in each experimet. N = 6 per group; *p<0.5 vs Aero; #p<0.5 vs I/R. I/R: in vitro chemical ischemia-reoxygenation; LF-EMF: low frequency electromagnetic field; MMP-2: matrix metalloproteinase 2. Scale bar = 100 µm. (A color version of this figure is available in the online journal)
Discussion
Naturally occurring LF-EMF in the range of extremely low frequencies (ELFs; 8–60 Hz)20,21 is normally involved in the interactions and physiological function of the nervous system, bones, heart tissue, and skeletal muscles.15,22,23 In this study, we used a signal which forms a multi-peak frequency spectrum with maximum of 195 Hz, which allows to classify it into low frequency EMF. There are several studies reporting contradictory observations regarding the relationship between LF-EMF and physiological as well as pathological processes. Raggi et al. 24 showed a significant reduction of blood malondialdehyde levels, which is a free radical marker, following LF-EMF exposure. Ma et al. 25 demonstrated acute exposure to LF-EMF protects isolated cardiomyocytes from I/R induced cell death by mediating ROS production and maintaining the NO/ONOO− balance. For this reason, we hypothesized that cardiac cells exposure to LF-EMFs can have beneficial effects and important cardioprotective actions. Continuing the hypothesis that electromagnetic field may be used as a therapeutic agent26,27 and following the study of Ma et al., 25 who demonstrated the inhibition of ROS generation by LF-EMF, we showed the cardioprotective role of LF-EMF in ex vivo model of cardiac I/R. 18 Cardiomyocytes are highly specialized, high-oxygen-requiring cells with large number of mitochondria. They occupy nearly 75% of the cardiac mass, but make only about one-third of the total cell number in the heart. The remaining cells in heart tissue are fibroblasts, endothelial cells, smooth muscle cells, epicardium cells, and others. 28 It is widely accepted that endothelial–fibroblast–cardiac myocyte interactions play a key role in regulation of heart function, even if the precise mechanisms of this interactions have not been completely understood.29,30 Capillaries are located next to cardiomyocytes and the crosstalk between myocytes and endothelial cells is involved in cardiac development, maturation, and function. 31 In this study, we showed in the I/R model that LF-EMF protects the metabolic function of main types of cells forming the structure of heart muscle, suggesting the potential use of LF-EMF as strategy for the prevention or therapy of I/R injury. The controversial data indicating the antioxidant capacity of LF-EMF in cardiac tissue 25 and its reduced antioxidant activity in the nervous tissue 32 prompted us to examine the potential protective role of LF-EMF in main types of cardiac cells separately. We examined primary lines of myocytes, endothelial cells, and fibroblast to show which cells, as the most sensitive to I/R injury, can be affected by LF-EMF.
As expected, the response to the I/R injury was different for endothelial cells, fibroblasts, and cardiomyocytes. The LDH activity (the marker of cell injury) was significantly lower in endothelial and fibroblast cells in comparison to cells perfused aerobically, likely due to an increased permeability of cell membrane and increased protein release. Moreover, LF-EMF protected only endothelial cells from I/R injury. Taking into account that LF-EMF increases antioxidant capacity of cardiomyocytes subjected to I/R injury, 25 and that the reduced production of oxygen radicals might decrease mitochondrial permeability, it could be suggested that LF-EMF is involved in restoration of proper function of sarcoplasmic reticulum and mitochondrion. 31 As a consequence, the inhibition of calcium overload and lipid peroxidation as well as inhibition of oxidative damage of DNA could have protected the cells from I/R injury. Interestingly, cardiomyocytes as cells containing a large amount of mitochondria (about 35% of cell volume) being the principal source of cellular ROS, 33 were more resistant to I/R injury than endothelial cells and fibroblasts. This may arise from paradoxical function of mitochondria as a central upstream regulator of cellular response to hypoxia. Klimova and Chandel 34 showed that mitochondrial ROS is essential for stabilization of hypoxia inducible transcription factors (HIFs), which allow transcriptional regulation of genes involved in cell survival. Moreover, LF-EMF affected cardiomyocytes in a different manner than endothelial cells. It intensified cardiomyocyte injury during ischemia and reperfusion, showing the diverse influence of LF-EMF on different cell types. This observation stays in agreement with the thesis of Golbach et al. 35 who suggested a different sensitivity of different cell types to LF-EMF. Taking into account that LF-EMF may lead to increase of intracellular calcium 36 and cardiomyocytes are particularly dependent from Ca2+ content, enhanced calcium load in myocytes together with ROS-induced dysfunction of sarcoplasmic reticulum may induce hypercontracture of myocytes and heart death. This may explain an increased cell damage of cardiomyocytes subjected to LF-EMF during I/R.
One of the well-known hypotheses about mechanisms of I/R injury of the heart is an increase in the activity of MMPs. 37 MMP-2 belonging to a group of gelatinases is abundantly expressed in normal cardiomyocytes and fibroblasts,38,39 and degrades extracellular 40 and intracellular matrix proteins. 38 Despite that function of MMP is essential for the proper function of many tissues and organs, 41 ROS-dependent enhanced activity of MMP-2 is related with an increased degradation of contractile proteins and heart failure.38,42 Testing the gelatinolytic activity of different cells types in the current study, we showed that 9 min in vitro chemical ischemia and 20 min reperfusion led to significantly decreased activity of MMP 2 in both cardiomyocytes and fibroblasts in comparison to cells maintained in aerobic conditions. In our previous study, we showed an increased activity of MMP-2 in the different model of ischemia and reperfusion. 43 We are aware that fibroblasts account for up to two-thirds of the total cell number in the normal human heart and remodeling of heart tissue induced by I/R is accompanied by decreased number of functional myocytes, hypertrophy of surviving myocytes and consequently by proliferation and migration of cardiac myofibroblasts. 44 The transformation of normal cardiac fibroblasts into the myofibroblast phenotype cells, which show an increased expression and activation of MMP, may be the main source of increased activity of MMP-2 in tested hearts. In the current study, we used the homogenous, established lines of HCM and fibroblasts isolated from human heart. Data showed that I/R decreased MMP-2 activity in these cells. This stays consistent with significantly decreased metabolic activity of myocytes and fibroblasts suggesting their inability to produce energy and consequently their metabolic insufficiency leading to death. Interestingly, endothelial cells undergoing chemical ischemia and reperfusion showed an increased activity of MMP-2 in comparison to endothelium maintained in aerobic condition. Endothelial cells play a crucial role in response to I/R due to angiogenesis. During generation of new vasculature, components of extracellular matrix surrounding the endothelium are intensively degraded. The study by Ben-Yosef et al. 45 showed that remodeling of extracellular matrix during post hypoxia and reoxygenation angiogenesis is a highly dynamic process depending from time of hypoxia and reoxygenation. MMP-2 activity was changing from decreased activity after 6 h of hypoxia to increased activity after 24 h of hypoxia. Our study showed that 9 min of chemical ischemia and 20 min of reperfusion led to an increased activity of MMP-2 in endothelial cells and its decreased activity in cardiomyocytes and fibroblasts suggesting a different role of MMP-2 in these cells. The variety of studies showing the distinct responses by different cell types to hypoxic conditions in recent years suggests that hypoxia/ischemia–reperfusion/reoxygenation injury is a complex and diverse issue dependent on cells types. However, irrespective of the direction of MMP-2 activity changes in different cells types, the use of LF-EMF during stabilization I/R led to reversion of these changes to the level of aerobic control. Both MMP-2 activity and metabolic activity of cardiomyocytes, fibroblasts, and endothelial cells (except MMP-2 activity in the latter ones) were significantly increased due to LF-EMF in comparison to I/R control.
In conclusion, this study demonstrates that short courses of LF-EMF protect cells of cardiac tissue from cellular damage and preserve their metabolic activity during in vitro chemical ischemia and reperfusion. We think that LF-EMF could be good addition to a pharmacological treatment to reach better heart protection from I/R injury. Taking into account the possibility of using LF-EMF in clinical practice, further studies are required to recognize the molecular mechanisms of LF-EMF’s cardioprotection.
Footnotes
Authors’ contributions
All authors participated in the design, interpretation of the studies and analysis of the data and review of the manuscript; DB, MW, IBL, AKZ, ASW, JA conducted the experiments; IBL, MW, GS designed the experimental part of the project; MW, DB supplied critical reagents, MW, IBL, GS provided the clinical expertise; IBL, GS, AKZ analyzed the data; IBL and GS wrote the manuscript.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Polish National Science Centre [grant number: UMO-2016/23/B/NZ3/03151]