
Abstract
Phospholipid membranes are necessary for the compartmentalization of chemistries within biological cells and for initiation and propagation of cell signaling. The morphological and chemical complexities of cellular membranes represent a challenge for dissecting the biochemical processes occurring at these interfaces. Therefore, investigations of the biological events occurring at the membrane require suitable models to reproduce the intricacy of this surface. Solid-supported lipid bilayers (SLBs) are simplified physical replicas of biological membranes that allow for bottom-up reconstruction of the molecular mechanisms occurring at cellular interfaces. In this brief review, we introduce how the properties of SLBs can be tuned to mimic biological membranes, highlighting the engineering approaches for creating spatially resolved patterns of lipid bilayers and supported membranes with curved geometries. Additionally, we present how SLBs have been employed to reconstitute molecular mechanisms involved in intercellular signaling and more recently, membrane trafficking.
Impact statement
Artificial membranes with complex topography aid the understanding of biological processes where membrane geometry plays a key regulatory role. In this review, we highlight how emerging material and engineering technologies have been employed to create minimal models of cell signaling pathways, in vitro. These artificial systems allow life scientists to answer ever more challenging questions with regards to mechanisms in cellular biology. In vitro reconstitution of biology is an area that draws on the expertise and collaboration between biophysicists, material scientists and biologists and has recently generated a number of high impact results, some of which are also discussed in this review.
Keywords
Introduction
Phospholipid membranes allow for compartmentalization of chemical signals within a biological cell. They separate the extracellular milieu from the cytoplasmic interior, defining a cell’s boundaries and delimit all the organelles within its cytosol. They also protect the genome from the cytosolic environment and attack from nucleases. Biological membranes are dynamic structures that permit bidirectional transport of cargo, cellular communication and signaling. Therefore, they are more than just a diffusion barrier and participate in cellular signaling, serving as scaffolds and surfaces for protein complexes. 1
The phospholipid bilayer has been known as a functional two-dimensional fluid since the 1970s 2 with the first bilayer structure based on amphiphilic phospholipids presented in 1925. 3 Today, our understanding of molecular processes at the biological membrane has been built upon studies performed on models of the cell membrane. These include structures such as liposomes, giant unilamellar vesicles (GUVs), monolayers and polymer-supported lipid bilayers4,5 (Figure 1).
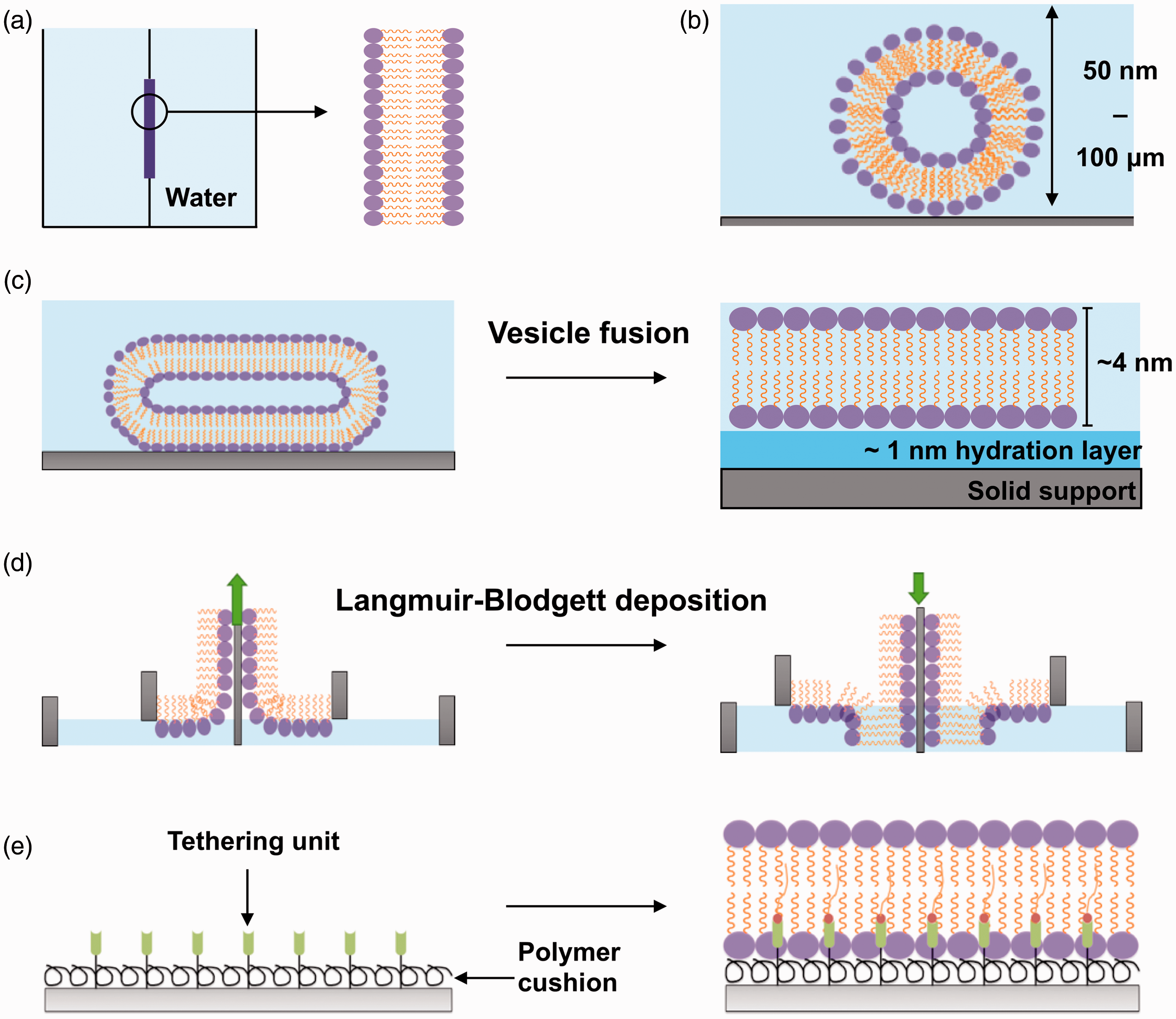
In vitro approaches used to model the biological membrane. (a) Black lipid membranes are formed when dried down lipids are hydrated with an aqueous medium over an aperture between two aqueous phases. (b) Liposomes (50 nm) or giant uni-lamellar vesicles (100 µm) are formed when dried down lipids are hydrated in an aqueous medium. (c) SLB formation by the vesicle fusion method. Liposomes are incubated onto a hydrophilic surface where they fuse, deform, and spontaneously rupture to form a uniform, continuous, fluid lipid bilayer. (d) SLB formation with the Langmuir-Blodgett technique. Lipid monolayers are formed at the air–water interface when lipids are added to an aqueous medium. This monolayer is transferred onto a hydrophilic solid support twice to form a lipid bilayer. The green arrow indicates the direction the substrate (grey) is moving to pick up another monolayer of lipids. (e) Polymer-supported/tethered lipid bilayers are formed on top of a polymer cushion hydrogel and also tethering units such as nickel NTA for anchoring of proteins. (A color version of this figure is available in the online journal.)
The planar lipid bilayers or solid-supported lipid bilayers (SLBs) are model systems constructed via self-assembly of lipids on a solid support. They can be engineered to exhibit similar fluid properties of crowded natural cell membranes, displaying lateral diffusion coefficients between 1 μm2 s−1 (similar to cell membranes) or slightly faster diffusion rates up to 3–4 μm2 s−1, depending on the physical properties of the support interface. 6 The lateral diffusion of lipids in SLBs is also influenced by the liquid-crystalline properties of the bilayer that change with lipid composition. 7 Lipid diffusion is confined to the plane of the solid support where the fluidity of lipid leaflets is sustained by a ∼1 nm layer of water parting the bilayer from the substrate. 8
SLBs have been utilized to investigate membrane organization, 9 protein self-assembly, 10 membrane remodeling11,12 and deduce mechanisms for spatial and temporal initiation of cell signaling at the plasma membrane.13–15 The major impact from investigating biochemical processes occurring at the membrane may be in the field of drug development, given that around 50% of drugs target transmembrane proteins. 16
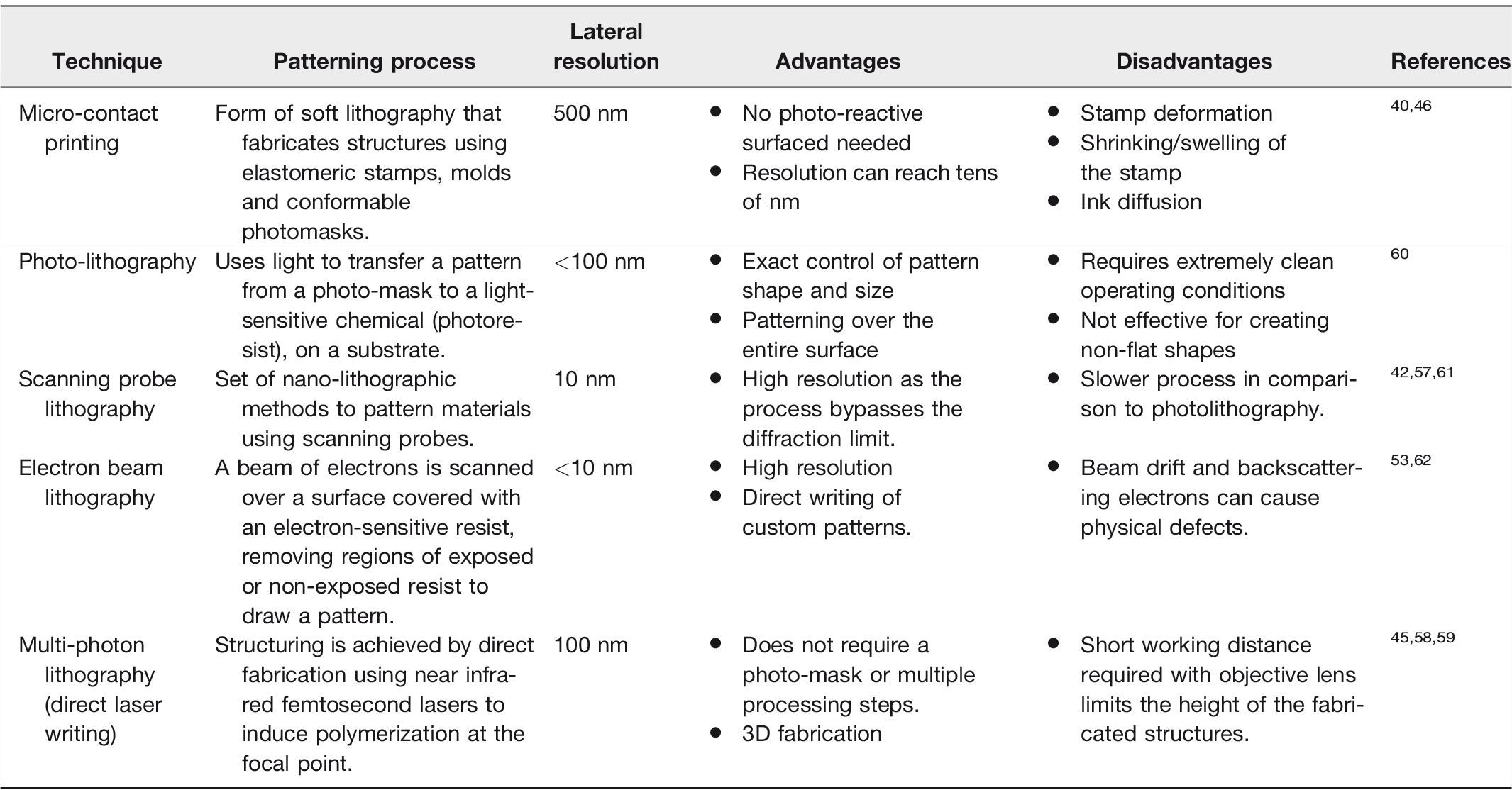
Importantly, SLBs can be designed to be more than just a flat lipid bilayer. Solid support and membrane bilayer morphology can be easily altered to accurately mimic a system of interest. Specifically, the spatial organization and mechanical deformation of supported membranes can be manipulated by patterning the underlying substrate with modern micro and nano-fabrication techniques (Table 1).
Outline of the techniques for patterning SLBs, their resolutions, and disadvantages/advantages.
In this mini review, we introduce the technologies to engineer spatially resolved flat lipid bilayers and membranes containing patterns of curvature. We discuss how these artificial systems facilitate the study of the biological pathways controlled by the interplay between cellular membranes and membrane-interacting proteins.
Towards building a realistic model of the cell membrane
Cellular membranes exhibit heterogeneity in lipid composition and display a variety of shapes and morphologies, which are necessary for specialization of function. For example, asymmetry in cholesterol lateral composition at the plasma membrane plays a role in the compartmentalization of protein complexes and in activating and regulating cell signaling via membrane trafficking.17,18 The function of membrane-bound organelles is linked to their complex morphologies. An exquisite example is the continuous membrane network of the endoplasmic reticulum (ER), with its closely packed membrane sheets. The ER membrane architecture maximizes the surface areas for protein synthesis, whereas the tubules in the peripheral are used for contacting other membrane organelles. 19 Some organelles change morphology progressively to acquire diverse functions, such as endosomes ‘maturing’ into multivesicular bodies during cargo trafficking to the lysosomes. 20
The earliest model system of a cell membrane was the black lipid membrane, developed by Mueller in the 1960s, 21 but it was not very versatile (Figure 1(a)). Today, liposomes and GUVs (Figure 1(b)) offer the possibility to study the role of lipid composition and membrane mechanics in the context of protein–membrane interactions. 4 Generally, GUVs are more appropriate for studying how a wide range of membrane physical parameters affect the interaction between membranes and other molecules. Nonetheless, SLBs offer specific advantages for quantifying kinetic processes at the molecular level since they are amenable to coupling with surface-sensitive analysis techniques, such as surface plasmon resonance and quartz microbalance. Low- and high-resolution imaging techniques, such as atomic force microscopy (AFM), fluorescence recovery after photobleaching, and total internal reflection microscopy (TIRF) permit image acquisition of membrane topography, quantification of membrane fluidity, and localization of protein assemblies at the supported membrane. 22
There is a choice of methods available to produce SLBs, depending on the extent of surface coverage required and the type of solid support used. The direct vesicle fusion 23 method is performed in aqueous buffers and involves liposome interaction, deformation, and spontaneous rupture into a single lipid bilayer, coating uniformly the solid substrate, usually silica or mica (Figure 1(c)). It is the most straightforward and commonly used approach to form SLBs, although it is less suitable to form membranes containing a high proportion of cholesterol, which impacts negatively on the efficiency of vesicle rupture. 24 In the solvent-assisted lipid bilayer method, the bilayer is formed following gradual exchange of a water-miscible organic solvent with an aqueous buffer solution, over lipids deposited onto the solid support. 25 The method is suitable to form bilayers onto a wide range of surfaces (e.g. silicon dioxide and gold 26 ), since it does not require an interaction between lipid vesicles and the surface. In the Langmuir-Blodgett technique,27,28 a lipid monolayer formed at an air–water interface is transferred to a hydrophilic surface. This technique can be used to produce asymmetric lipid bilayers, which mimic natural cell membranes more closely and are not accessible with the vesicle fusion method (Figure 1(d)). Recently, spin coating has also been established as a facile method to deposit membranes onto a variety of solid supports.29,30
A major difference between reconstituted flat lipid bilayers and biological membranes is the lack of the bulk aqueous phase of the cytosol underneath the lipid leaflet, which confers fluidity to cellular membranes. The small hydration layer, spacing the membrane from the solid support, does not accurately mimic the properties of the cellular cytosol. Therefore, in SLBs, the solid support and the lipid bilayer are not fully decoupled, and the nature of the surface can have major effects on the properties of the membrane. For instance, stable lipid bilayers can form around highly curved substrates, such as 100 nm diameter silica nanoparticles, 31 by means of electrostatic interactions between the lipid vesicles and the silica support. Moreover, supported lipid bilayers can adapt to changes induced by substrate plasticity,32,33 a behavior that can be exploited to control the mechanical properties of the membrane.
In biological membranes, cytoskeletal scaffolds disengage the lipid leaflets from the bulk phase of the cytosol. Synthetic materials that exhibit mechanical properties similar to those of the cytoskeleton can be sandwiched between the bilayer and the solid support. For example, polymer brushes 34 or generic polymer nanofibers 35 can be used to recapitulate the function of actin networks and provide the reconstituted lipid bilayer with fluidity close to that observed in biological membranes. These polymer-supported SLBs can also be spatially patterned and incorporate proteins of interest36,37 (Figure 1(e)). Conversely, SLBs have also been used also to study the assembly of cytoskeletal proteins, such as the polymerization of an actin scaffold triggered from vesicles containing ponticulin, an initiator of actin self-assembly. 38
Engineering of spatially patterned bilayers to investigate cell signaling
SLBs can be engineered as spatially organized patterns of lipid bilayers with specific compositions. Spatial patterning can be achieved with methods where either the membrane is locally confined by the substrate topography (e.g. microcontact printing)39–41 or parts of the membrane are removed after lipid bilayer deposition (e.g. photolithographic techniques).42–45 Therefore, solid supports can be engineered to introduce ad hoc morphological features to study the biological process of interest. An overview of methods for patterning a solid support is presented in Table 1 and summarized below.
Microcontact printing is a fairly versatile approach to form patterned membranes. It is based on a form of soft lithography as it utilizes soft elastomers, typically polydimethylsiloxane (PDMS). 39 The PDMS surface is patterned and used as a contact stamp to transfer a thin layer of material onto a substrate. Its main limitations include stamp deformation or stretching and compressing of the stamp, introducing unwanted features into the pattern. During transfer, the ink can laterally spread into unwanted regions not replicating the desired pattern. 46 Polymer “lift-off” techniques are a form of microcontact printing where a thin layer of a sacrificial material (aluminum or a polymer) is patterned onto glass. Once an SLB has formed, the material is etched away removing lipids in the patterned regions47–49 (Figure 2(a)).
Currently, the techniques that can fabricate membrane arrays with the highest resolution are electron beam 53 and scanning probe lithography, 42 which can reproduce features down to 10 nm in size (Table 1). Scanning probe lithography uses an AFM tip to pattern substrates, meaning that features with size beyond the diffraction limit are accessible via this technique. Other types of scanning probe lithography either involve removal of material from the surface of the substrate (thermal scanning probe lithography) 43 or use diffusion at the tip of the scanning probe to coat patterns onto substrates, such as alkanethiol arrays on gold substrates (dip-pen lithography).44,54 Patterned bilayers can be formed by removal of lipids from a membrane using UV/ozone photolithography55,56 or using atomic force microscopy. 57 Multi-photon lithography exploits the chemical changes occurring in substrates that can simultaneously absorb two or more photons, such as hydrogels, natural polymers, and even proteins. 58 The critical size of the features generated with this technique can be as small as 100 nm, since the chemical transformations occur in a region with a volume of few attoliters. 59
Patterned SLBs provide an advanced platform for the investigation of complex biological processes at the molecular level. For instance, they can be used to study the molecular mechanisms of endocytosis, 63 cell–cell interactions, 64 and communication.65,66 Lipid membranes can be spatially organized with micrometer size diffusion barriers and employed to study molecular events occurring within confined geometries, such as the molecular details of T-cell activation. 67 Similarly, patterned arrays of lipid bilayers 68 can be used as sensor arrays for cell adhesion.13,69 Physical barriers for lipid diffusion can be made using masks created from materials that prevent membrane formation (e.g. metals, some polymers). These barriers are usually taller than the average height of a lipid bilayer (∼4 nm), but recently, it has been shown that graphene 70 offers the possibility to create diffusion barriers as thin as one-atom.
The role of spatial organization of transmembrane receptors in intercellular signaling has been extensively studied using patterned SLBs. 71 At the plasma membrane, internal and external inputs can alter the spatial organization of cell surface receptors. For instance, intercellular signaling requires close contact between transmembrane receptors to allow direct communication between neighboring cells. Reproducing, in vitro, an intercellular junction is a complex task. However, the combination of solid-state nanolithography with supported lipid bilayers allow to study the relationship between spatial organization and transmembrane receptor activity. 72 For example, Greene et al. 73 used electron beam lithography to create physical barriers to lipid bilayer lateral mobility creating membrane corrals containing the tyrosine kinase receptor Ephrin A1 (EphA1). This system was used to guide breast cancer cells to the EphA1-patterned membranes via the interaction with the cognate receptor Ephrin A2 (EphA2), thereby creating a semi-synthetic junction. This spatial setup of Ephrin receptors revealed the relevant molecules involved in the endocytosis of EphA2, an important step in the regulation of the EphA2 receptor signaling pathway. 73
Similar experimental approaches have been applied to the investigation of events involved in the mechanical regulation of cell surface proteins. For instance, patterned SLBs have been employed to study the relationship between the organization of epidermal growth factor receptor (EGFR) and its phosphorylation levels 74 and for the understanding of Ras signaling at a single-molecule level. 75
Engineering curved membranes to understand the mechanisms of membrane remodeling
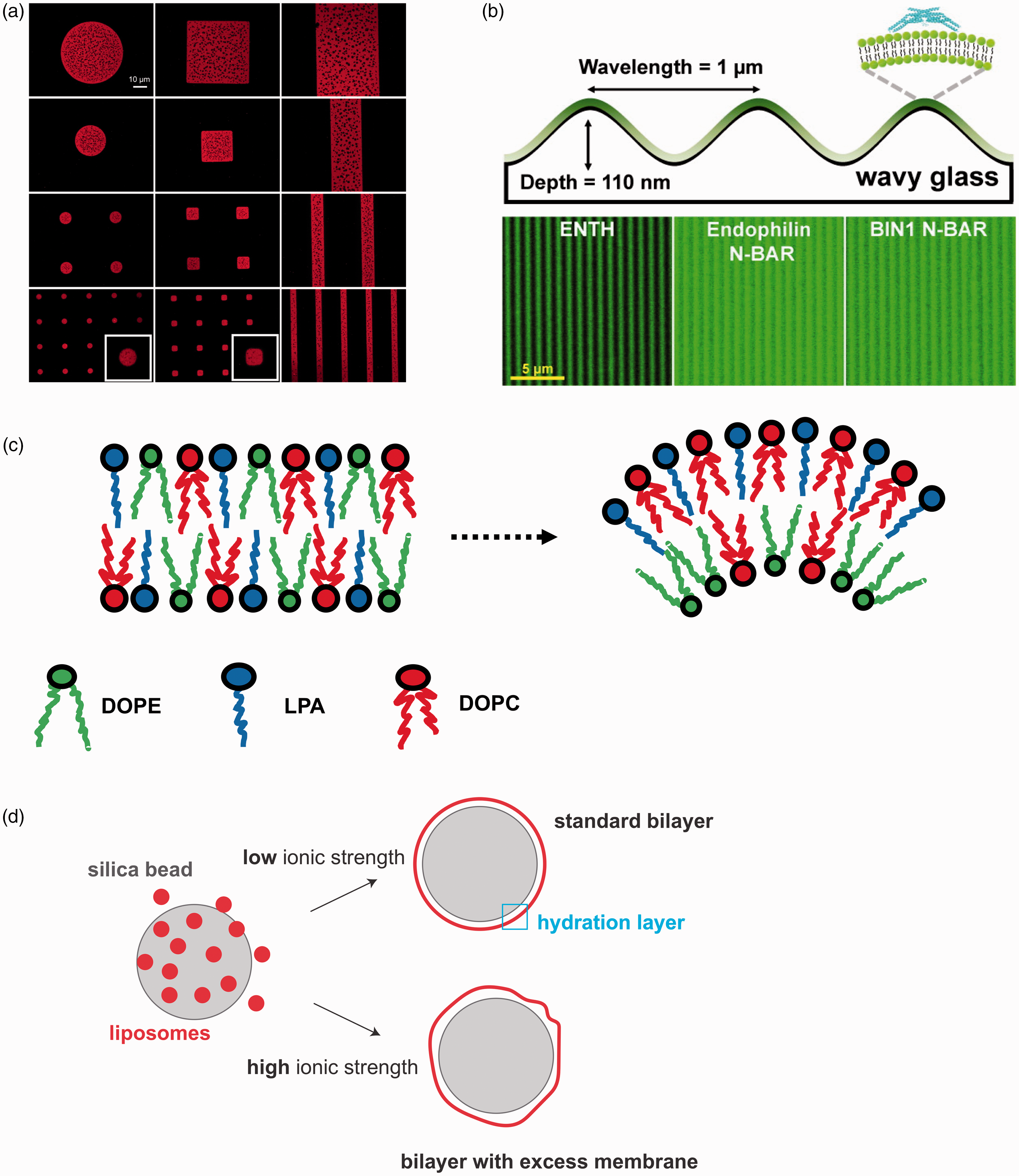
Membrane geometry and cellular signaling are intimately connected, 76 particularly in biological processes where macromolecular assembly at the membrane is regulated by curvature.1,77,78 Membrane shape and lipid composition are features of membrane organelles, which are not only evolutionarily conserved but are also finely controlled to maintain spatial segregation of molecules during membrane trafficking.79,80 Over a specific threshold (∼0.8 µm−1), curvature appears to regulate the spatial organization of lipid phases composed of cholesterol 81 and the segregation of lipids such as hexadecanoic acid 82 or cardiolipin. 83 Importantly, local membrane curvature also controls the segregation of curvature-sensing proteins 50 (Figure 2(b)). For instance, lipidated proteins appear to be selectively recruited by regions of specific membrane curvature. 84 The correlation between substrate curvature and lipid organization underlines the existence of feedback mechanisms that regulate the interaction between the membrane and protein assemblies. 85 Therefore, methods to introduce regions of curvature in model membranes are highly desirable to study events where lipid sorting and membrane deformation are crucial mechanistic steps for recruiting protein complexes, 86 such as membrane trafficking.
Curved supported membranes are formed via direct fusion of lipid vesicles onto the pre-patterned solid support. Localized regions of membrane curvature can be introduced by means of grooves, edges, or other non-flat features at the surface of the SLB solid support. For example, patterns of curvature can be produced by manufacturing cylindrical concavities with a focused-ion beam, on a chromium-coated coverslip onto which the lipid bilayers are assembled. 87 Alternatively, nanopits 88 and nanoparticles 89 can be used to guide the formation of regions of local curvature in lipid bilayers. It is worth noting that there is a physical limit to the curvature achievable for a phospholipid membrane on a solid support. This was exemplified by the study of Roiter et al., 90 which explored the behavior of membranes deposited on surfaces decorated with nanoparticles. They observed that lipid bilayers form continuously on silica nanoparticles smaller than 1.2 nm or larger than 22 nm, while pore formation in the membrane around the particle was likely within this size range.
Non-flat membranes facilitate the investigation of processes where an asymmetric segregation of lipids occurs laterally or between the leaflets of the membrane at very high curvatures (e.g. tens of nm). 91 Coupling between lipid structures and curvature preference is a consequence of the interactions between different lipid shapes, which prefer to arrange as locally curved bilayers.51,81 Lipid sorting can be a key signal to drive transmembrane protein clustering 92 and cell signaling.93,94 For instance, lysophosphatidic acid displays an inverted cone shape that confers its tendency to populate regions of membranes with positive curvature51,95,96 (Figure 2(c)).
Complex membrane topographies containing curves and ridges 97 can enable the investigation of how protein complexes remodel phospholipid membranes. Membrane remodeling proteins display curved structures, which are used for sensing curvature and for inducing further membrane bending.1,86,98 Special types of SLBs have been recently developed to specifically follow membrane remodeling reactions. The SUPported bilayers with Excess membrane Reservoir52,99 are assembled on micron-scale silica beads in such a manner that a loosely fitted membrane is deposited on the solid support (Figure 2(d)). The excess membrane allows for membrane deformation to occur and be monitored by fluorescence and electron microscopy. These supported membranes have been successfully used to reveal the physical basis of membrane remodeling promoted by protein crowding 11 or performed by endocytic100–103 and autophagic proteins. 104 Similarly, the supported membrane tubes (SMrT) 105 mimic the membrane tethers pulled by proteins106,107 and can be used to study the impact of local curvature on protein–membrane interactions.
A unique membrane remodeling protein complex has recently received attention in the membrane biophysics community, the Endosomal Sorting Complex Required for Transport (ESCRT). ESCRT function is vital for key cellular processes, such as multivesicular body biogenesis, cytokinesis, neuron pruning, plasma and nuclear membrane repair, and viral budding108,109 (Figure 3(a)). In all these processes, the topology of membrane deformation by ESCRT is the same: the budding occurs away from the cytosol, 110 in contrast, for example, to bud formation by dynamin in endocytosis.103,111
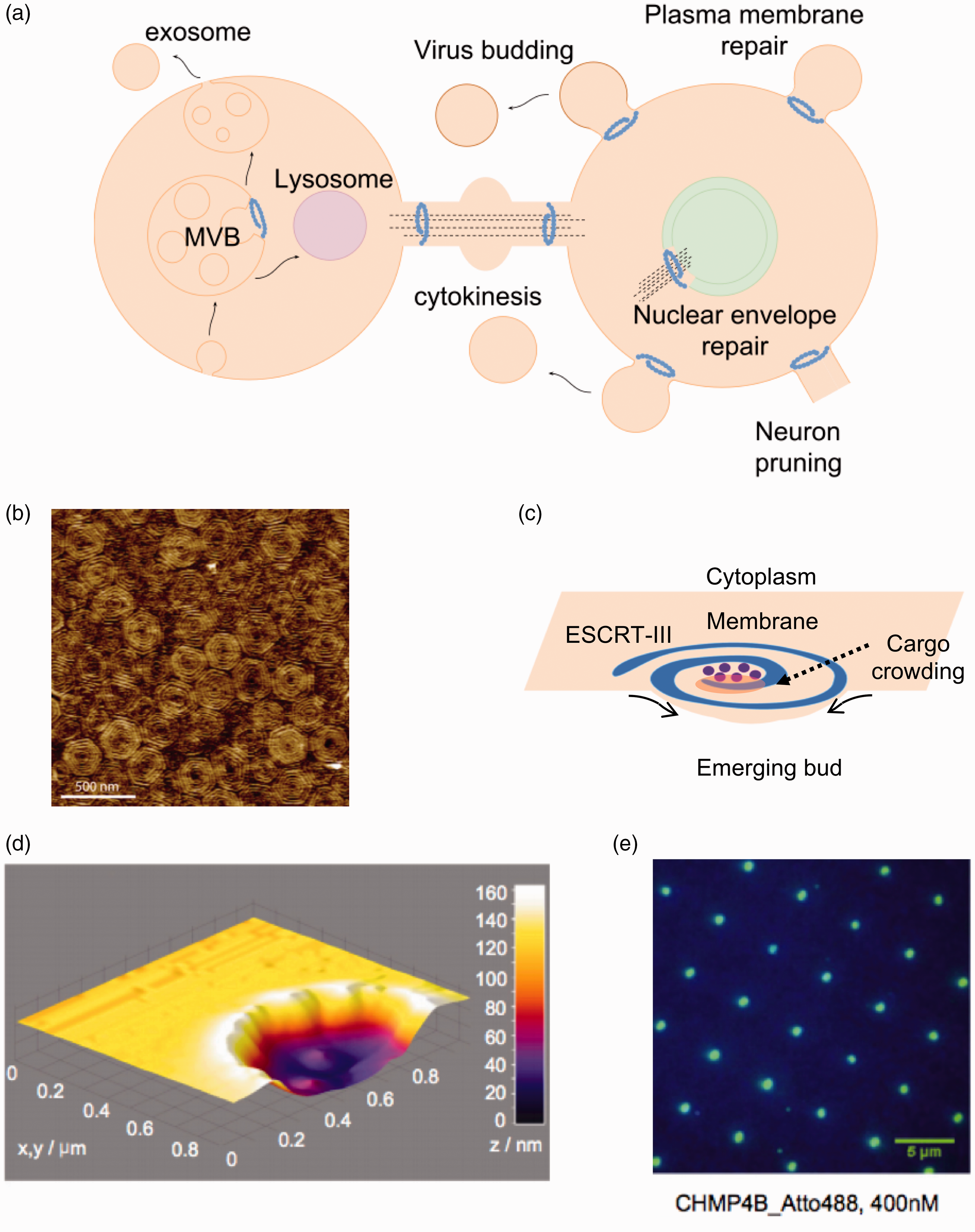
Membrane remodeling activity by the ESCRT complex. (a) Brief overview of some of the cellular functions of the ESCRT complex in mammalian cells. In all processes, ESCRT-III remodel membranes away from the cytosol. The core remodeling machinery (ESCRT-III) is indicated as a blue spiral. ESCRT-III organizes as circular filaments to stabilize the neck of membranes buds. (b) Atomic force microscopy topographic image of the center of a Snf7 patch assembled on GUV membrane. Image reprinted from Chiaruttini et al. 115 with permission from Elsevier. (c) The spiral structure acts as a spring collecting elastic energy which upon release buckles the membrane. (d) 3D surface plot of the atomic force microscopy profile showing lipid bilayer formation on top of the 100-nm-deep invaginated template for negative curvature, which approximates the shape of a nascent HIV-1 bud produced by focused ion beam. Image reproduced from Lee et al. 87 with permission from Proceedings of the National Academy of Sciences USA. (e) Total internal reflection fluorescence (TIRF) image after 20-min incubation with 400 nM of the human ESCRT-III core subunit CHMP4B-Atto488 (DiD fluorescein used as a blue marker for lipids). Image reproduced from Lee et al. 87 with permission from Proceedings of the National Academy of Sciences USA. (A color version of this figure is available in the online journal.)
Given its unique activity, the biophysics and biochemistry of budding and scission reactions by the ESCRT complex have been the focus of intense investigations. The assembly of ESCRT proteins into an active complex has been reproduced using GUVs112,113 and SLBs, which allowed to dissect the biophysical basis of membrane remodeling. The main scission machinery of ESCRT, namely ESCRT-III, has been observed to assemble into concentric circle-like structures on lipid bilayers114,115 (Figure 3(b)). These studies suggested a potential mechanism for membrane deformation that involves flat “spirals” of ESCRT-III acting like springs. ESCRT-III spirals undertake lateral compression on the membrane and incorporate elastic energy, which leads to membrane deformation upon energy release (Figure 3(c)). 115 In another study, SLBs were used to decipher how the growth of ESCRT-III circular structures is self-regulated and how ESCRT-mediated membrane remodeling is coupled to ATP hydrolysis. 116
ESCRT proteins assemble and stabilize the neck of a budding membrane; therefore, membrane curvature plays a role in protein recruitment and assembly at these locations. 108 Lee et al. 87 created a SLB with features resembling membrane bud necks, by assembling a lipid bilayer on a substrate with 100 nm deep concavities, creating regions of negative curvature (Figure 3(d)). TIRF microscopy was used to monitor the assembly of the human ESCRT-III complex at the membrane, which revealed a clustering of the proteins specifically at the invaginations of the membrane, demonstrating ESCRT activity at membrane bud necks 87 (Figure 3(e)).
Perspective and conclusions
We have presented a brief overview of the SLB technology and reviewed the fabrication of spatial and curved patterns for in vitro reconstitutions of biological processes at the membrane.
Flat synthetic membranes are a powerful tool to investigate how the biophysics of lipid–lipid interactions contributes to the assembly of complex biological structures.117–119 Advances in the engineering of structural features in solid supports is allowing the generation of ever more complex spatial or curvature patterned bilayers, thus creating more realistic mimics of cellular membranes. SLBs with rough surfaces facilitate the systematic dissection of the interplay between curvature and lipid sorting and crucially, how these physical properties are coupled and controlled in a complex biological process. Lipid phases can be coupled with nanometer-scale curvature patterns, 120 providing a strategy for organizing a controlled segregation of biomolecules and lipids.
A yet unmet challenge in the SLB field is the in vitro reconstruction of more complex biological membranes, such as the nuclear envelope. The nuclear envelope is a double membrane, 121 in which transmembrane proteins couple the inner membrane to chromatin and the outer membrane relays signals from the cytoskeleton. 122 Such an in vitro model would allow the study of the molecular basis of epigenetic mechanisms and nuclear envelope membrane repair in physiological and diseased contexts. 123 We are still far from being able to design a fit-for purpose model of the nuclear envelope although SLBs composed of multiple bilayers have been recently developed.48,124 Therefore, it appears that the field is ripe for moving towards increasing complexity with the destination of yet more complex models to interrogate biological mechanisms and/or reconstruct cell biology from the bottom-up.
Footnotes
Authors’ contributions
All authors contributed equally to the writing and editing of this article.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
BC and FC thank the EPSRC for a studentship to Sarina Chand under the DTA scheme (EP/L505055/1). BC and PB thank the EPSRC (grants EP/M027821/1 and EP/M027929/1) for funding.