
Abstract
Plant species with recognized estrogenic activity and widely used by Brazil’s female population to prevent the unpleasant symptoms of menopause were investigated in this work to demonstrate if constituents of taro-inhame (Colocasia esculenta), cumaru (Dipteryx odorata), and camapu (Physalis angulata) have the ability to mimic or interfere with the action of estrogens. Moreover, their potential use as natural sources of estrogen-like substances for hormone replacement therapy was evaluated. (a) In vivo pharmacological assays were conducted to determine the estrogenic effects of D. odorata isoflavone-rich extract, P. angulata physalin-rich extract, and C. esculenta flavonoid glycoside-rich fraction (FG) on endocrine glands and reproductive organs of female rats at three different stages of the life cycle. The protocols consisted of uterotrophic assays and cytological evaluation of vaginal smears to detect mucosa cell alterations correlated with changes in hormone levels in each phase of the estrous cycle of female rats. The results indicated that C. esculenta FG exhibited estrogenic activity in prepubescent, pubescent, and adult ovariectomized female rats, while D. odorata isoflavones only promoted a weight increase in the pituitary gland of prepubescent rats after prolonged treatment and P. angulata physalins induced a weight increment in the adrenal glands of ovariectomized rats. Additionally, C. esculenta exerted a significant effect on the opening of the vaginal canal in prepubescent rats and on vaginal epithelium. Prolonged treatment of ovariectomized rats with FG altered the proportion of different types of vaginal epithelial cells in these animals, suggesting an interference of FG with estrogen levels. Colocasia esculenta FG induced hypertrophy of the uterus and pituitary in ovariectomized rats similar to estradiol. To elucidate the mechanism of action of FG, its effects were compared to those of estradiol and of the selective estrogen receptor modulator raloxifene. The results suggest that the efficacy of C. esculenta FG is mediated by binding to selective estrogen receptors present in each organ and that raloxifene inhibits the mechanism of action of FG in the same way as it inhibits the effects of estradiol. The overall findings indicate that C. esculenta FG mimics the action of estrogens, with reduced harmful effects on specific tissues.
Impact statement
The results of this work provide preliminary evidence that ovariectomized rats display the vaginal smear cells typical of the estrous phase of mature rats after long-term ingestion of Colocasia esculenta flavonoid glycosides. Moreover, the treatment caused beneficial effects on endocrine and reproductive organs compared to estradiol. The animals did not exhibit significant body weight alterations among the groups, demonstrating the maintenance of standard metabolism and energy balance. The overall findings obtained with the rat model of menopause highlight the importance of dietary consumption of C. esculenta constituents as potential selective estrogen receptor modulators and suggest that they may prevent some of the metabolic disorders related to estrogen deficiency.
Introduction
Quality of life before, during, and after menopause has been pursued by women worldwide. Menopause is the period that marks the end of the menstrual cycle and is characterized by a reduction in ovarian estrogen secretion. The deficiency of this hormone leads to uncomfortable changes in metabolism, causing hot flashes, night sweats, mood changes, sexual dysfunction, urogenital atrophy, bone loss, predisposing to osteoporosis, lipid profile alterations that result in the accumulation of fats, cardiovascular diseases, and diabetes. 1
Since the mid-1970s, with the advancement of the pharmaceutical industry, the use of synthetic hormones that mimic the action of endogenous hormones has become the first-choice option to minimize menopause discomforts. However, some adverse effects associated with the consumption of these drugs have encouraged the search for drugs with fewer or no adverse effects such as medicinal plant-derived drugs or phytotherapics. Considering the many restrictions on the use of synthetic hormones, especially in women with a family history of breast or uterine cancer, 2 the safe use of this class of chemicals is a matter of concern to both patients and clinicians. Therefore, complementary and alternative therapies, especially the use of plants rich in phytoestrogens, are an increasingly common trend in contemporary society. 3
The development of ethical phytomedicines requires correlation between preconized therapeutic effects and chemical profile of tested phytocomplexes, besides elucidation of the mode of action of bioactives. For those reasons, it is important to develop phytochemical approaches to obtain fractions rich in the potential estrogen-like compounds to be tested instead of using whole plant material in the pharmacological assays.
The estrogenic effect of phytosterols β-sitosterol and stigmasterol was evaluated and confirmed to be functionally similar to 17 β-estradiol, 4 acting as estrogen agonists for estrogen receptor (ER) α and β, preferentially binds to ERβ.5,6 Those compounds were proved to be a safe alternative for estrogen replacement therapies.
The evidenced estrogenic activity of natural sources of phytoestrogens like isoflavones from soybean (Glycine max)7,8 has motivated the scientific interest to evaluate potential estrogen-like natural compounds.
Colocasia esculenta (taro), Dipteryx odorata (cumaru), and Physalis angulata (camapu) have been popularly recognized by the Brazilian female population, who uses them to treat the unpleasant symptoms of menopause. Tubers of C. esculenta as well fruits of P. angulata have been included in diary food intake for decades, with no risk to health. However, further studies to validate the efficacy of constituents from these species in hormone replacement therapy are necessary. 9
We previously reported on the isolation of isoflavones from Dipteryx odorata10,11 and also on the isolation of ergostan-secosteroids (physalins) from P. angulata. 12 In this work, phytochemical procedures using LC-MS2 and GC-MS techniques to analyze an acetate fraction from C. esculenta crude methanol tuber extract, allowed the detection of glycosylated flavonoids and steroids capable of acting as estrogen-like compounds.
To investigate the estrogenic/antiestrogenic activity of structurally heterogeneous groups of phenolic compounds obtained from P. angulata roots, D. odorata callus, and C. esculenta roots, prepubescent, pubescent, ovariectomized, and non-ovariectomized adult Wistar rats were treated with the isoflavone-rich fraction of D. odorata extract, ergostane-type steroids from P. angulata or the flavonoid glycoside-rich fraction (FG) of C. esculenta root extract. The animals were evaluated to determine body weight during treatment, weight of the uterus and endocrine glands. Moreover, secondary sexual characteristics such as vaginal opening and vaginal epithelium, the presence or absence of renal and hepatic toxicity and the possible mechanism of action of phytocomplexes that exhibited a trophic effect on the uterine endometrium were investigated.
Materials and methods
Plant material
Callus in vitro cultures of D. odorata (Voucher HUAM 7276, Herbarium of Universidade do Amazonas), established and subcultured over years, were the source of the isoflavone-rich fraction which was extracted as described by Fernandes et al. 10 Dried and powdered roots of P. angulata (Voucher 173388, Herbarium of Instituto Nacional de Pesquisas Agronômicas – INPA) stored in the Medicinal Plant Collection at the UNAERP provided the physalin-rich fraction. Sliced and dried tubers of C. esculenta grown on Sítio Bela Vista in Pratápolis, MG, were the source of the FG fraction.
Plant extracts
Dipteryx odorata isoflavone-rich extract
D. odorata callus cultures were established and extracted as described by Januario et al. 11 In short, freeze-dried callus was extracted with chloroform at room temperature. The CHCl3 extract was concentrated in a rotary evaporator, fractionated by silica gel column chromatography using ethyl acetate and methanol solvent mixtures, and further analyzed by HPLC (Supplemental data Figure S1 – Chemical profile of plant phytocomplex). Fractions rich in isoflavones (CH3 COOC2H5:CH3OH 1:1) (41.35 ± 1.95 mg/g dry weight) were dried, resuspended in a hydroalcoholic solution (methanol:water, 1:1), and used in the pharmacological assays.
P. angulata ergostan-rich extract
P. angulata roots cultured in vitro were dried, powered, and extracted following the procedures described by Januario et al. 12 Crude CHCl3 extract was then fractionated by silica gel column chromatography using hexane and ethyl acetate solvent mixtures and further analyzed by HPLC. Fractions rich in physalins (C6H14:CH3COOC2H5 3:7) (150.00 ± 3.20 mg/g dry weight) with a defined HPLC (Supplemental data – Figure S2) profile were dried, resuspended in a hydroalcoholic solution, and used in the pharmacological assays.
C. esculenta flavonoid glycoside-rich extract
C. esculenta tubers were sliced, dried in an air circulating oven at 50°C, powdered, and maintained in the dark free of humidity. The powdered material (2 kg) was macerated in methanol for 48 h, filtered, and evaporated under vacuum. The crude MeOH extract was resuspended in hydroalcoholic solution and successively partitioned with hexane and ethyl acetate solvents to afford the FG fraction used in the in vivo biological assays. Analyses developed in GC-MS and HPLC MS2[M − H]− were carried out to determine the chemical profile of C. esculenta FG.
Animals
After approval of the experimental protocol by the Animal Ethics Committee of the Universidade de Ribeirão Preto (UNAERP), a first in vivo assay was carried out with female Wistar rats in different stages of the life cycle (n = 75). For a subsequent experiment, adult and ovariectomized Wistar rats (n = 38) were used. The rats were kept in the animal room at the UNAERP under controlled conditions of humidity (70%), temperature (21°C±2), and light/dark photoperiod (12/12 h), receiving a balanced diet and water ad libitum.
Oral administration of solutions
Animals were grouped and received oral treatments for 3, 7, or 21 days in the approximate volume of 400 µl/animal, through the gavage procedure, using a rounded and curved tip needle, rat-specific, which is inserted into the animal's mouth and carefully pushed through the esophagus to the stomach to ensure that the animal receives the full dose administered. 13
The animals received hydroalcoholic solution (methanol and water, 1:1), Dipteryx odorata (160 mg/kg), 10 P. angulata (160 mg/kg), 14 C. esculenta (80 mg/kg), 15 C. esculenta flavonoid glycoside-rich extract (80 mg/kg), estrogen (50 µg/kg estradiol benzoate), and raloxifen (0.75 mg/kg).
Experiment with extract fractions from three plant species administered to rats in different stages of maturity
Experimental groups of female Wistar rats, divided according to life cycle stage, were fed with C. esculenta FG (80 mg/kg), D. odorata isoflavones (160 mg/kg), and P. angulata physalins (160 mg/kg) for different periods of time. Ovariectomized adult rats (60 days old) were treated for 7 days, and prepubescent rats (21 days old) were treated for 3 or 21 days. To evaluate the estrogenic/antiestrogenic action of tested compounds, the negative control group received corn oil (0.1 ml, s.c.) and hydroalcoholic solution by gavage and the positive control received estradiol cypionate (10 µg/0.1 ml/animal s.c. Pfizer, Paulinia, SP, Brazil) 16 and hydroalcoholic solution by gavage (Table 1). Following the designed protocol (Figure 1), all rats were anesthetized with 2.5% tribromoethanol (TBE) administered intraperitoneally (1.0 ml/100 g body weight) before opening of the abdominal cavity and diaphragmatic rupture. The animals were then euthanized in accordance with international standards for euthanasia, and their uteruses, adrenal and pituitary glands, kidneys, and liver were removed for evaluation of morphological alterations. The uterine weight of treated and untreated animals was compared on the last day of the experiment. After perforation, the organs were pressed between two sheets of filter paper for liquid drainage as described by Clode, 17 and the uterine dry and wet weights were determined.
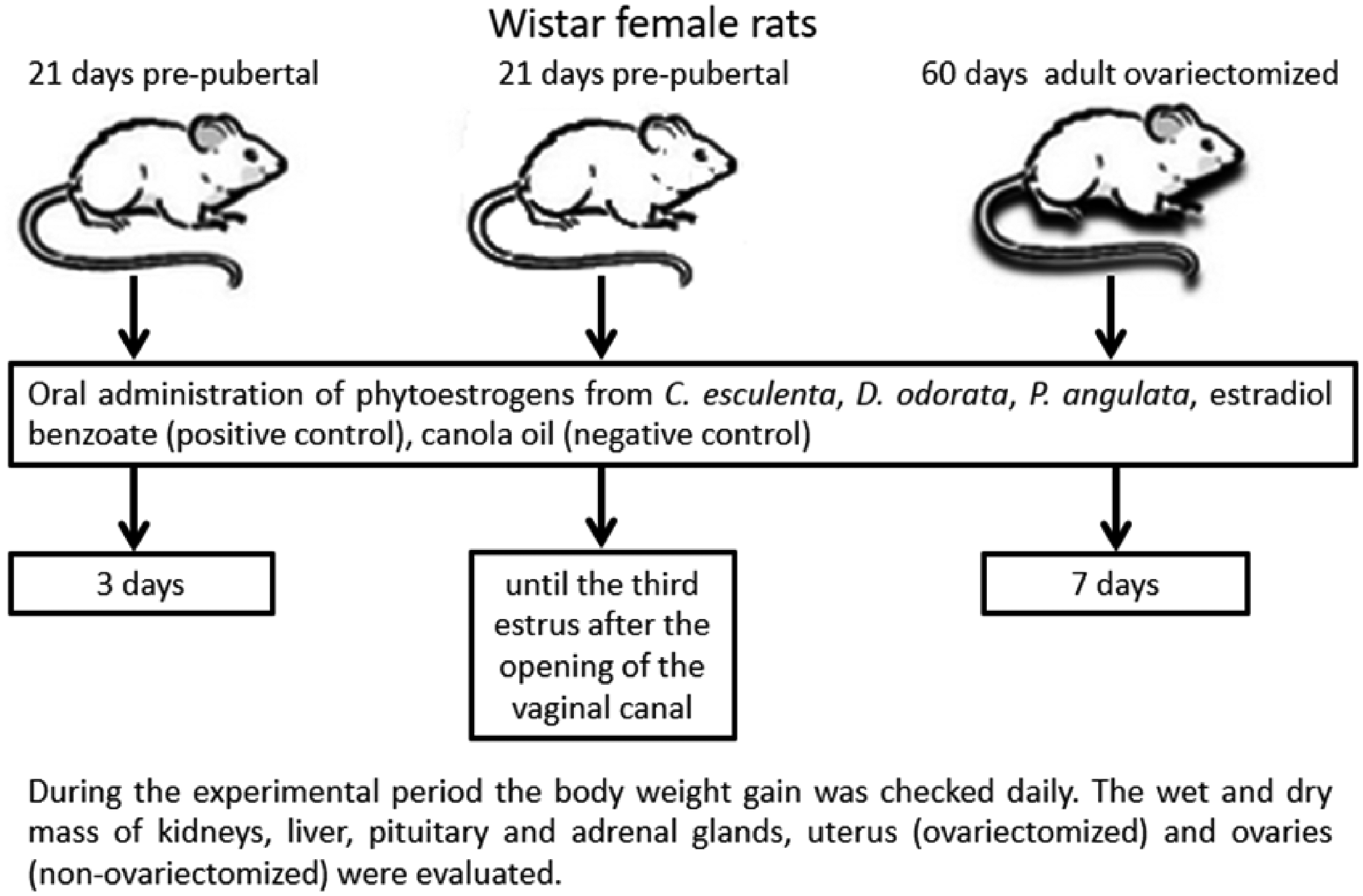
Design of the experimental protocol.
Type and dosage of drugs administered for different periods to five groups of rats at three different stages of life cycle: adult ovariectomized (seven days); pre-pubescent rats (three days); pre-pubescent/pubescent rats treated until the third estrus after opening of vaginal canal (21 days).
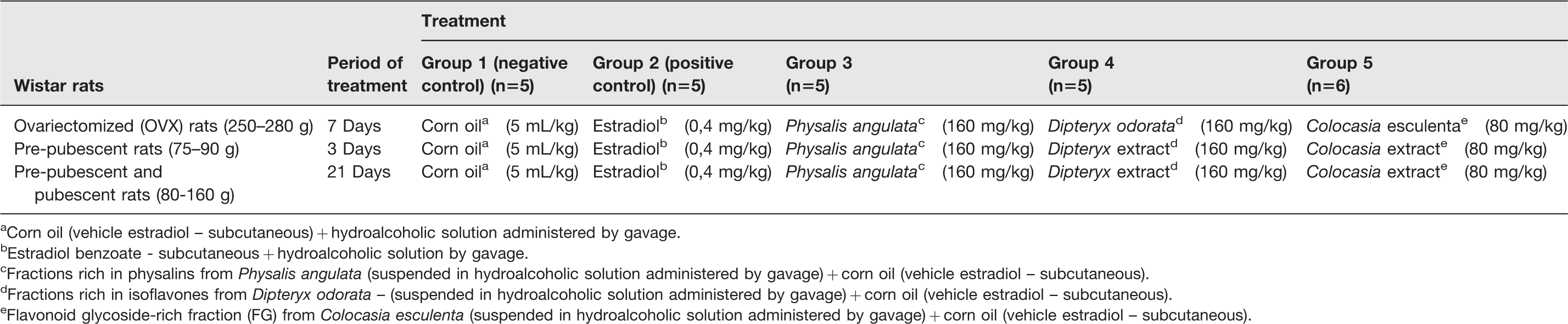
aCorn oil (vehicle estradiol – subcutaneous) + hydroalcoholic solution administered by gavage.
bEstradiol benzoate - subcutaneous + hydroalcoholic solution by gavage.
cFractions rich in physalins from Physalis angulata (suspended in hydroalcoholic solution administered by gavage) + corn oil (vehicle estradiol – subcutaneous).
dFractions rich in isoflavones from Dipteryx odorata – (suspended in hydroalcoholic solution administered by gavage) + corn oil (vehicle estradiol – subcutaneous).
eFlavonoid glycoside-rich fraction (FG) from Colocasia esculenta (suspended in hydroalcoholic solution administered by gavage) + corn oil (vehicle estradiol – subcutaneous).
Experiment with C. esculenta flavonoid glycoside-rich extract using ovariectomized rats
Sixty-day-old (adult) Wistar rats (n = 38) were anesthetized intraperitoneally (1 ml/100 g) with 2.5% TBE, and bilateral ovariectomy was performed. After surgery, all rats were treated with a prophylactic dose of pentabiotic (0.1 ml/rat, Fort Dodge, Campinas, Brazil) and analgesic flunixin meglumine (2.5 mg/kg i.m., Banamine®, Schering-Plough, Rio de Janeiro, Brazil). 16 Fourteen days after surgery, the animals were distributed in six groups as follows and treated orally for 21 days: group 1: 0.1 ml propylene glycol + 0.1 ml hydroalcoholic solution; group 2: 50 µg/kg estradiol benzoate; group 3: 0.75 mg/kg raloxifene; group 4: 80 mg/kg C. esculenta FG; group 5: 50 µg/kg estradiol + 0.75 mg/kg raloxifene, and group 6: 0.75 mg/kg raloxifene + 80 mg/kg C. esculenta FG.
Vaginal smear cytology
On the day of euthanasia, exfoliative vaginal cytology analysis was carried out to verify the mucosa cells from vaginal epithelium. Fresh vaginal material was then collected and analyzed by optical microscopy. Vaginal smear cytology allows estimating the estrogen action on the vaginal epithelium, considering the morphological alterations in the vaginal epithelium as well the proportion of different cell types in the vaginal smear. Circulating estrogen acts on the rat vaginal epithelium and stimulates cell proliferation and keratinization. 18 The epithelium morphology and cell types characterize the estrous cycle phase and the correlated level of reproductive hormone of the rats: In estrous phase, the epithelium is squamous, stratified, and with round and nucleated keratinized cells. In metestrous, the epithelium is squamous and stratified with loss of the keratin layer. A leukocyte infiltration, extending from the base to the surface of the epithelial layer is also present. 19
Trophic effects on pituitary glands and reproductive organs
All the rats were anesthetized with 2.5% TBE administered intraperitoneally (1.0 ml/100 g body weight) and euthanized by rupture of the diaphragm. Subsequently, pituitary gland, uterus, liver, kidneys, ovaries, and adrenals were removed for analysis. The weight of each organ was measured and calculated to the animal’s body weight (weight organ/100 g body weight) corresponding to the last day of the experiment. The uterus was submitted to double evaluation: wet and dry weight. Estrogen has a trophic effect on specific structures of the hypothalamic–pituitary–gonadal (HPG) axis and are used to assess the estrogenic or antiestrogenic effect of some hormones.20,21 The changes in weight of the pituitary, adrenal glands, in dry and wet weight of the uterus, liver, kidney, and ovaries of rats treated for 3 and 21 days and of ovariectomized adult rats treated for 7 and 21 days were determined.
Statistical analyses
The statistical analyses were performed using analysis of variance (ANOVA), followed by Tukey’s test. The analyses were performed using the statistical software GraphPad Prism 5.0, with significance level fixed at p < 0.05.
Results
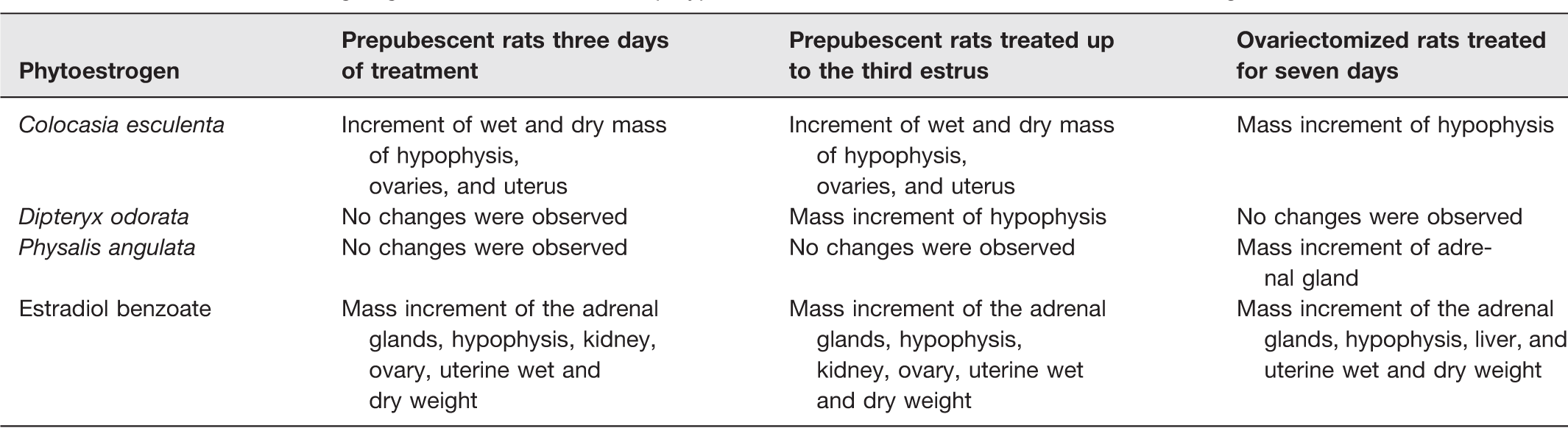
The assays to investigate the effects of P. angulata, D. odorata, and C. esculenta bioactive compounds on the estrous cycle of prepubescent rats revealed that C. esculenta flavonoid glycoside-rich extract (C. esculenta FG) exerted the most significant estrogenic effect on opening of the vaginal canal and on estrogenic action on the vaginal epithelium at the end of treatment (Table 2).
Mass increment of organ/gland of rats treated with polyphenolics from C. esculenta, D. odorata, and P. angulata.
The results obtained showed that 66% of prepubescent rats treated for 21 days remained in the estrus/proestrus stages of the estrous cycle, evidencing the estrogenic effects of the constituents of the phytocomplex on rat estrous cycle (Table 3). Further experiments were then carried out using C. esculenta FG, which was administered for 21 days to ovariectomized adult rats to validate its estrogenic effect and to investigate its mechanism of action compared to raloxifene, a selective estrogen receptor modulator (SERM) which exhibits agonist or antagonist effects, depending on the affected organ/gland. Six treatment groups of ovariectomized rats were examined.
Estrous cycle stages in prepubescent rats treated with plant crude extracts for 21 days.

aVaginal opening did not occur.
A significant agonist effect, characterized by an increase in adrenal weight, was observed in rats treated with estradiol and with the combination of estradiol plus raloxifene (Figure 2(a)). Moreover, there was a significant increase of pituitary weight in rats treated with C. esculenta FG, estradiol, C. esculenta FG plus raloxifene, or raloxifene plus estradiol. The results indicated that C. esculenta FG displayed a trophic action on the pituitary when compared to the control group treated only with vehicle like the trophic effect induced by estradiol, though the estradiol exhibited a greater trophic effect than the C. esculenta FG. Raloxifene exhibited an estrogen agonist effect on the pituitary, promoting a weight increase probably due to its SERM characteristics. Co-administration of FG plus raloxifene or raloxifene plus estradiol did not promote effects different from those induced by the individual compounds (Figure 2(b)).
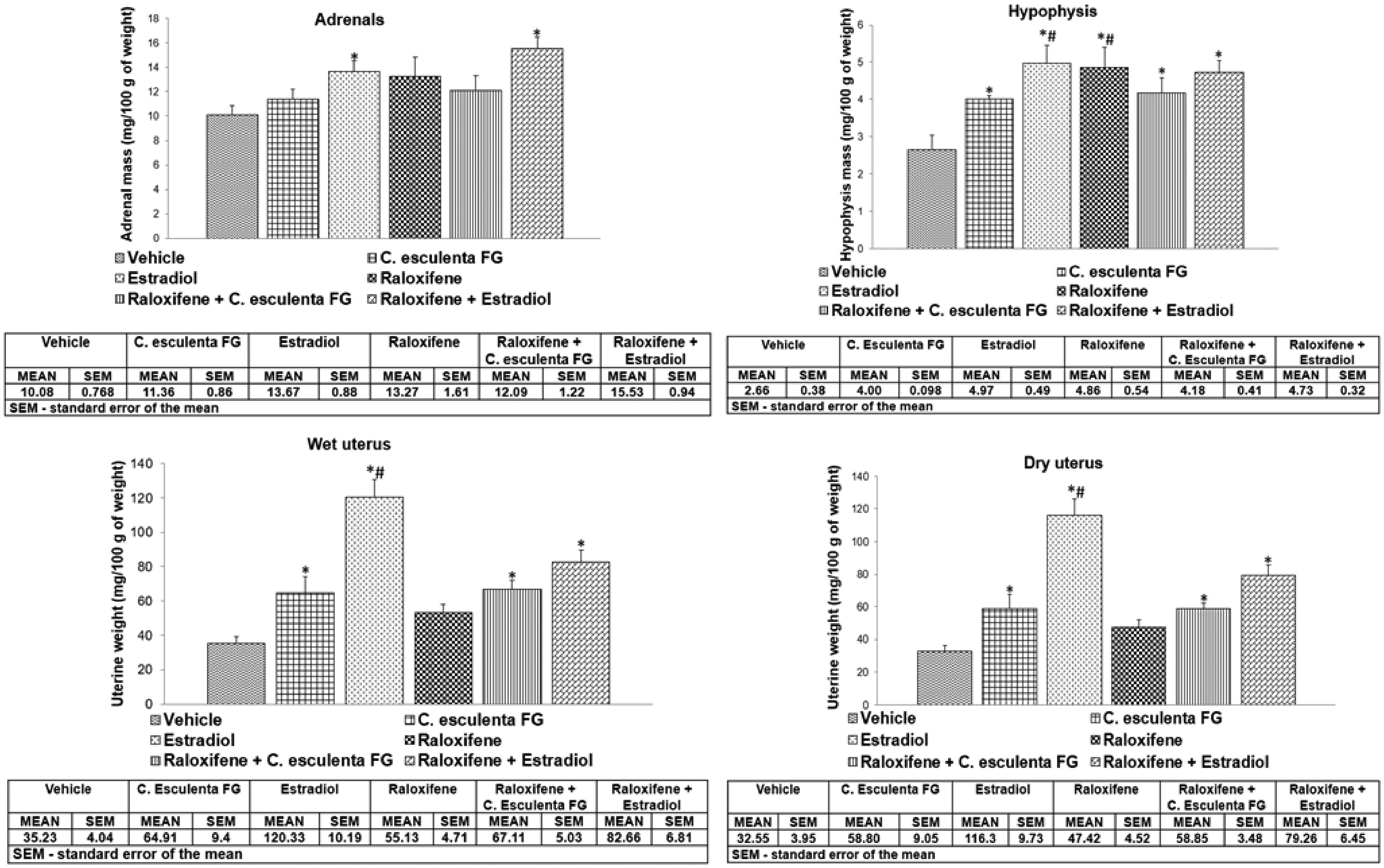
Changes in the weight increase of adrenal glands, pituitary and uterus dry and wet weight of ovariectomized adult rats treated with raloxifene, estradiol or Colocasia esculenta FG for 21 days. The statistical analyses were performed using analysis of variance (ANOVA), followed by Tukey’s test. The analyses were performed using the statistical software GraphPad Prism 5.0, with significance level fixed at p < 0.05.
An increase in uterine dry and wet weight was also detected in rats treated with C. esculenta FG and estradiol, while raloxifene did not exert a significant uterotrophic effect. Rats treated with C. esculenta FG, estradiol, raloxifene + FG, or raloxifene + estradiol had an increased uterine weight compared to rats treated with vehicle. As expected, the treatment with estradiol caused a more pronounced effect compared to the other treatments. Rats treated only with raloxifene did not show uterine weight increases, and when combined with estradiol, partially blocks the trophic action of this hormone on the uterus, preventing gain in uterine weight, suggesting that this compound functions as an antagonist to estrogen in the uterus. Differently, co-administration of raloxifene plus C. esculenta FG did not inhibit or reduce the trophic action of C. esculenta FG on the uterus, suggesting that the estrogen receptors for the C. esculenta compounds differ from those for raloxifene and estradiol (Figure 2(c) and (d)).
Cytological analysis of the vaginal smears of ovariectomized adult rats treated for seven days with the phytocomplexes showed that the animals remained in the metestrous phase. Prolonged treatment (21 days) of ovariectomized rats with C. esculenta FG fraction altered the proportion of the different types of vaginal epithelial cells in these animals (Figure 3), indicating an interference of these constituents with the hormone levels controlled by the HPG system (Table 4).
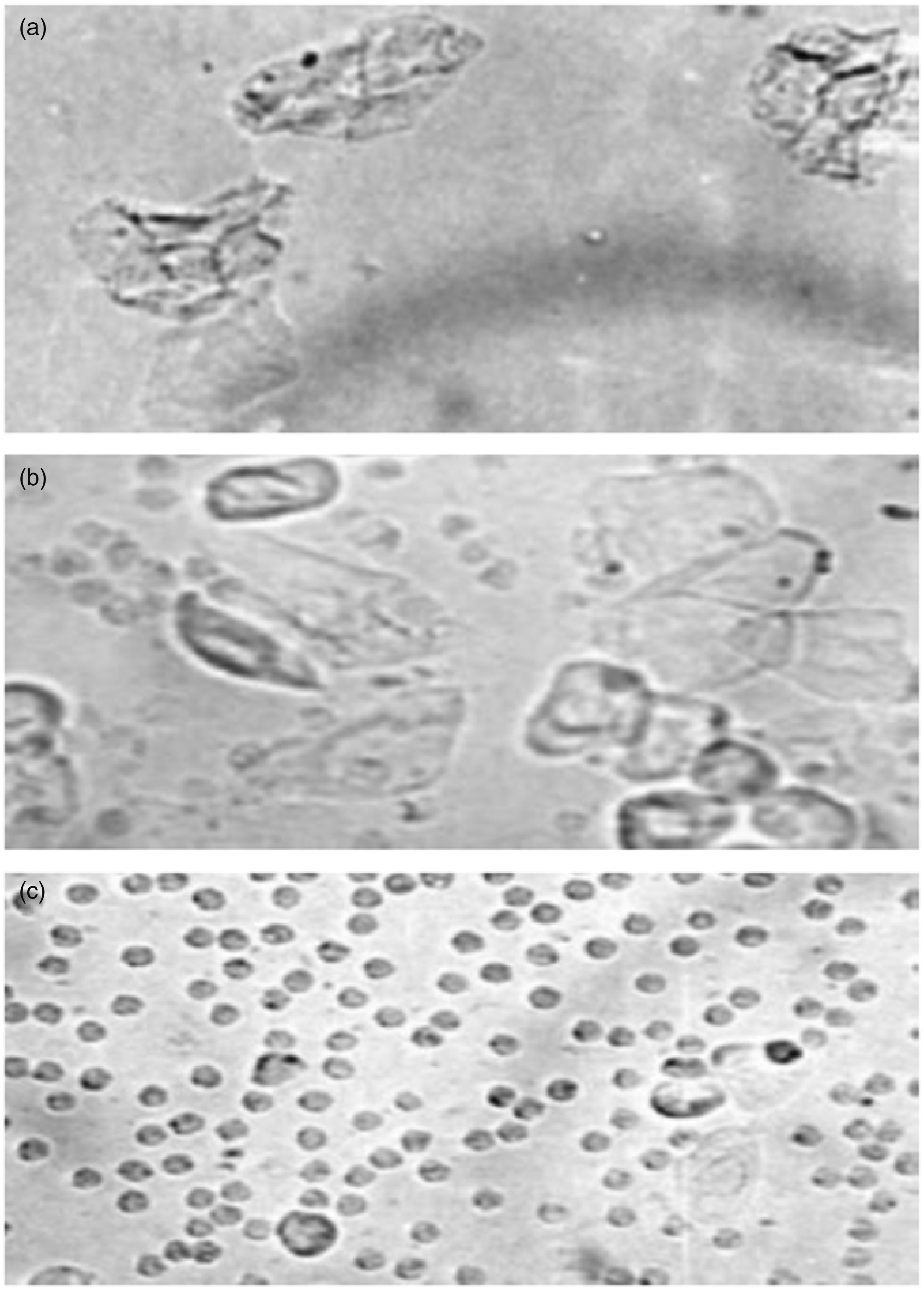
Photomicrographs of vaginal smears obtained from ovariectomized rats. (a) Rat treated with estradiol (estrus phase of the cycle characterized by a predominance of anucleated cornified cells); (b) rat treated with Colocasia esculenta FG (metaestrus phase of the cycle characterized by the presence of leukocytes, cornified cells and nucleated epithelial cells); (c) rat treated with vehicle (diestrus phase of the estrous cycle characterized by a predominance of leukocytes).
Percentage of OVX rats presenting vaginal smear cytological characteristics at the final period of the experiment, typical of different stages of the estrous cycle of pubescent rats, according to the treatment administered.

OVX: ovariectomized.
aVehicle for dilution of Raloxifene.
bVehicle for dilution of C. esculenta FG.
cLow levels of reproductive hormones.
dHigh levels of reproductive hormones.
Images of kidney tissue sections obtained from rats treated with the C. esculenta FG fractions revealed that convoluted tubule cells were unaffected, exhibiting normal characteristics, while kidney tissue of rats treated with raloxifene (0.75 mg/kg) + estradiol (50 µg/kg) presented areas of vascularization and cell degeneration. In addition, the treatment with raloxifene and estradiol caused loss of integrity of cuboidal cells in the renal cortex region where filtration occurs (Figure 4). Images of liver tissue sections of treated rats showed preserved hepatocytes, organized hepatic parenchyma, and centrilobular vein of regular appearance (Figure 5). The results obtained validate the safe use of C. esculenta FG to mimic or interfere with the action of estrogen, with reduced harmful effects on specific tissues.

Images of histological slides of renal tissue sections stained with hematoxylin-eosin of control (a) and rats treated with purified (b) Colocasia esculenta FG; (c) raloxifene (d) raloxifene + C. esculenta FG; (e) raloxifene + estradiol. Notice that cortex and renal pyramids maintain normal appearance. (A color version of this figure is available in the online journal.)

Images of histological slides of liver tissue sections stained with hematoxylin-eosin of control (a) and rats treated with purified (b) Colocasia esculenta FG; (c) raloxifene (d) raloxifene + C. esculenta FG; (e) raloxifene + estradiol. Note that images of liver tissue sections of treated rats showed preserved hepatocytes, organized hepatic parenchyma and centrilobular vein of regular appearance. (A color version of this figure is available in the online journal.)
GC-MS analysis of the acetate fraction of tuber crude methanol extract of C. esculenta showed the steroids stigmastrol and β-sistosterol present in abundance in FG. (Supplemental data – Figure S3)
LC-MS2 data analysis from the acetate fraction of tuber crude methanol extract of C. esculenta present is showed in Supplemental Information. These data were obtained in a comparison with Bruker HMDB Metabolite Library and also with the literature. 22 Analyses evidenced the presence of flavonoid derivatives and glycosylflavonoids: apigenin (m/z 270); eriodictyol-hexose (m/z 450); chrysoeriol-hexose-pentose (m/z 606); apigenin-glycoside (m/z 414); luteolin-pentose (m/z 430), catechin derivative (m/z 385; native catechins m/z 289), naringenin (aglycone m/z 272), orientin (m/z 448), quercetin-hexose-pentose (m/z 610; aglycone m/z 302), and proved to be a useful technique to characterize the chemical profile of the main components of C. esculenta FG (Supplemental data – Figure S4).
Discussion
During the experiments conducted, we found that the crude extracts of C. esculenta and D. odorata contain substances that induced morphological alterations and weight changes in the pituitary, adrenal glands, and uterus of all treated rats and also affected ovarian function of prepubescent rats compared to controls. These structural changes may have been triggered by the action of estrogenic compounds produced by C. esculenta 23 and D. odorata. 10
Menopausal women are usually deficient in endogenous estrogens hormones and their body weight tends to increase. According to Kirchengast et al., 24 this occurs because estrogen increases the consumption of body energy, which contributes to body weight reduction. Thus, they waste less energy and consequently have increased body weight, demonstrated by adipose tissue gain and a reduction of lean body mass.25,26 Synthetic hormone replacement therapy using estradiol benzoate tends to reduce obesity.27,28 In this work, we did not observe changes in gain weight among the studied groups, only a tendency to reduce weight gain in animals treated with estradiol. In women, the role of estrogen in controlling food intake and energy expenditure is demonstrated by the fact that after menopause, there is an increase in adiposity and food intake.26,29 It was also observed that the removal of the ovaries induced hyperphagia and weight gain in the animals, demonstrating the estrogen inhibitory tonic effect on the feeding behavior, and this effect was reversed by the treatment of these animals with estrogen.26,30 According to Torrezan et al., 31 isoflavones trigger a lipolytic effect on adipocytes and reduce lipogenesis in white adipose tissue of ovariectomized rats. Additionally, Russell et al. 32 demonstrated an interaction of dietary isoflavone and estradiol on body weight regulation in ovariectomized rats. No change in weight gain between the studied groups was observed, probably because of the treatment duration, since the estrogen-treated group showed a trend towards decreased weight gain at the end of treatment (Figure S5). Thus, a longer period of treatment might be necessary, so that we could have observed a reduction in weight gain in the animals treated with estrogen and in the extracts studied.
Makella et al. 33 reported that isoflavones exhibit a moderate estrogenic effect, which may be associated with the ineffectiveness of isoflavones to interact with estrogen receptors present in the endometrium, perhaps because estrogen interacts more easily with estrogen receptor alpha (ER-α). This may indicate that isoflavones have a selective action and exhibit estrogenic activity only in specific tissues. Mosquette et al. 34 described uterine changes in ovariectomized rats when isoflavones at doses of 300 mg/kg/day or higher were administered to rats over a 21-day period. Furthermore, Ribeiro et al., 35 treating hypoestrogenic rats with soybean isoflavones (100 mg/kg/day) during a prolonged period, found that a low dose of isoflavone was not sufficient to promote uterine development. These findings suggest that the potential of isoflavone to induce uterine weight gain is associated with the phytohormone dosage. Moreover, Carbonel et al., 36 investigating the action of the isoflavone genistein (42–250 mg/kg) administered to ovariectomized rats, observed an increase in uterine weight, endometrial gland hyperplasia, and an augmented number of eosinophils depending on the dose tested. Higher doses of genistein caused uterine squamous metaplasia. The results obtained in this work demonstrated that oral administration of the D. odorata isoflavone-rich phytocomplex caused a weight increase in the rat’s pituitary gland that plays a role in endocrine axis functionality. The weight increase of the uterus and pituitary in rats treated with FG indicated a hypertrophic action of this class of substances, which contribute to the estrogenic activity attributed to the C. esculenta phytocomplex.
Phytoestrogens possibly act through initial binding of alpha (ER-α) and beta (ER-β) estrogen receptors and interaction with key steroidogenic enzymes. 37 In the light of the pleiotropic effects of FG observed in this work, we further investigated the probable mechanism of action of FG. Experiments were carried out to compare the effects of raloxifene, an SERM, 38 and FG. Additional results were obtained from the co-administration of raloxifene and C. esculenta FG to ovariectomized rats. FG actually exhibited a trophic action on uterine tissue but raloxifene partially inhibited the mechanism of action of FG in the same way as it partially inhibited the effects of estradiol. This probably occurred due to direct stimulation of estrogen receptors present therein, with ligand–receptor complex conformational alterations being responsible for the key pharmacological differences exhibited by the SERMs. Alternatively, this may be due to a competition between FG and raloxifene for the estrogen receptors, with FG probably interacting better with those receptors than raloxifene. The same effect was observed when the weight increase of pituitary tissues was analyzed in rats treated with FG or with the combination of raloxifene and FG. Our results demonstrate that both estrogen and C. esculenta FG supplementation promoted increased pituitary weight compared to controls (treated with vehicles). Studies have demonstrated that estrogens control many aspects of the pituitary gland biology, including regulation of lactotroph homeostasis and synthesis and secretion of prolactin. Moreover, they display a trophic effect on the pituitary gland, especially on the proliferation of lactotrophs, the most abundant cell type in anterior pituitary gland.20,21 Obtained results indicate that C. esculenta FG may have an estrogen-like effect on the pituitary gland causing hypertrophy.
The hepatic and renal safety verified in this study corroborates the findings reported by Lewu et al. 39 who fed Wistar rats cooked C. esculenta-based diets for 28 days. Histological and biochemical analysis of liver and kidney tissues confirmed that oral ingestion of C. esculenta extracts presents no risk of toxicity.
To the best of our knowledge, there are no previous reports on the estrogen-like effects of C. esculenta despite its traditional use by the population to treat symptoms of reproductive hormone disorders. Experiments carried out in this study using raloxifene permit the prediction of the probable mechanism of action of C. esculenta flavonoid glycosides as SERMs and open an avenue to investigate a variety of plants that, like taro-inhame, are popularly used as natural phytoestrogen-containing diets.
Understanding the mechanism of action of estrogen-like natural products traditionally used may lead to the development of more effective formulations using innovative bioactive compounds, which may be indicated for menopaused women who do not tolerate conventional hormone replacement therapy. 40
Conclusions
The overall results indicated that a synergistic action between phytoestrogens and endogenous estrogens only occurs in female rats that still produce endogenous estrogens. Therefore, in order to ensure the effectiveness of phytoestrogens, it is crucial to establish the optimal maturity period to initiate hormone replacement therapy in order to prevent the unpleasant symptoms of menopause so that the appropriate physiological conditions for the occurrence of this synergistic action are guaranteed.
Data from this work support the hypothesis that senescent rats benefit from the prolonged treatment with C. esculenta flavonoid glycosides, as demonstrated by the positive effects induced in cells of the vaginal epithelium that displayed proliferation and keratinization similar to those of pubescent rats in the proestrus/estrus phase of the estrous cycle. Consequently, it is necessary to determine the most appropriate time from the onset of menopause to start an alternative hormone replacement therapy and to establish the duration of treatment.
Taken together with other data reported in the last decade, our results reinforce the potential use of natural products of different chemical classes as phytoestrogens to treat menopause symptoms as long as the mechanism of action and toxicity of plants rich in compounds with estrogenic activity are completely elucidated. This study demonstrated the estrogenic effects of C. esculenta flavonoid glycosides and no toxicity was detected on the hepatic and renal tissues evaluated.
Supplemental Material
Supplemental Material1 - Supplemental material for Effects of estrogen-like plant compounds on the vaginal epithelium pituitary, adrenal glands, and uterus of rats
Supplemental material, Supplemental Material1 for Effects of estrogen-like plant compounds on the vaginal epithelium pituitary, adrenal glands, and uterus of rats by Gismar MC Rodrigues, Bruno DB Borges, Leticia Gabriela Q Moreira, Érica Aparecida G Rossete and Suzelei de Castro Franca in Experimental Biology and Medicine
Supplemental Material
Supplemental Material2 - Supplemental material for Effects of estrogen-like plant compounds on the vaginal epithelium pituitary, adrenal glands, and uterus of rats
Supplemental material, Supplemental Material2 for Effects of estrogen-like plant compounds on the vaginal epithelium pituitary, adrenal glands, and uterus of rats by Gismar MC Rodrigues, Bruno DB Borges, Leticia Gabriela Q Moreira, Érica Aparecida G Rossete and Suzelei de Castro Franca in Experimental Biology and Medicine
Supplemental Material
Supplemental Material3 - Supplemental material for Effects of estrogen-like plant compounds on the vaginal epithelium pituitary, adrenal glands, and uterus of rats
Supplemental material, Supplemental Material3 for Effects of estrogen-like plant compounds on the vaginal epithelium pituitary, adrenal glands, and uterus of rats by Gismar MC Rodrigues, Bruno DB Borges, Leticia Gabriela Q Moreira, Érica Aparecida G Rossete and Suzelei de Castro Franca in Experimental Biology and Medicine
Supplemental Material
Supplemental Material4 - Supplemental material for Effects of estrogen-like plant compounds on the vaginal epithelium pituitary, adrenal glands, and uterus of rats
Supplemental material, Supplemental Material4 for Effects of estrogen-like plant compounds on the vaginal epithelium pituitary, adrenal glands, and uterus of rats by Gismar MC Rodrigues, Bruno DB Borges, Leticia Gabriela Q Moreira, Érica Aparecida G Rossete and Suzelei de Castro Franca in Experimental Biology and Medicine
Supplemental Material
Supplemental Material5 - Supplemental material for Effects of estrogen-like plant compounds on the vaginal epithelium pituitary, adrenal glands, and uterus of rats
Supplemental material, Supplemental Material5 for Effects of estrogen-like plant compounds on the vaginal epithelium pituitary, adrenal glands, and uterus of rats by Gismar MC Rodrigues, Bruno DB Borges, Leticia Gabriela Q Moreira, Érica Aparecida G Rossete and Suzelei de Castro Franca in Experimental Biology and Medicine
Footnotes
Authors’ contributions
Conceived and designed the study: SdCF. Performed the pharmacological assays: GMCR, BDBB, EAGR. Histological analysis: EAGR. Chemical analysis of plant extracts: LGQM. Analyzed the data: SdCF, GMCR. Wrote the paper: SCF, GMCR, BDBB.
Acknowledgements
DECLARATION OF CONFLICTING INTERESTS
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
FUNDING
This work was sponsored by UNAERP and GMCR was granted with a scholarship (91864968672) from the Coordination for the Improvement of Higher Education Personnel (CAPES).
Availability of data and material
All data generated or analyzed during this study are included in this published article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.