
Abstract
Gastrointestinal toxicity arising from cancer treatment remains a key reason for treatment discontinuation, significantly compromising remission. There are drawbacks to the currently used in vitro and rodent models, and a lack of translatability from in vitro to in vivo work. A screening-amenable alternative in vivo model such as zebrafish would, therefore, find immediate application. This study utilized a transgenic reporter line of zebrafish, Tg(cyp2k18:egfp), that shows eGFP induction as an indicator of drug-induced pathology. Here, we investigate its utility as an alternative vertebrate model to bridge the gap between simple in vitro cellular studies and complex in vivo models for understanding gastrointestinal toxicity induced by chemotherapy and targeted therapy. Transgenic zebrafish larvae were administered afatinib or SN38, and assessed for viability and eGFP induction. Adult zebrafish were administered afatinib via oral gavage, and SN38 via intraperitoneal injection. Fish were killed after 24 h, and had gastrointestinal tracts removed and assessed for histopathological damage, goblet cell changes, and apoptosis. While treatment with either compound did not induce eGFP in the gastrointestinal tract of larvae, SN38 caused histopathological damage to adult intestines. The lack of eGFP induction may be due to poor solubility of the drugs. Chemotherapy agents with high solubility and permeability would be more amenable to these models. Further progress in this area would be greatly facilitated by the generation of robust and reproducible genetic models of zebrafish intestinal toxicity that mimic the known pathobiological pathways in rodents and humans, and can be readily induced in a short time-frame.
Impact statement
Gastrointestinal toxicity secondary to cancer treatment remains a major reason for the termination of cancer drug candidates in the development pipeline as well as withdrawal or restrictions of marketed drugs. Current cancer treatment-induced gastrointestinal toxicity models available are limited to in vitro and rodent models that lack translatability and are prohibitively expensive and time consuming. An alternative model to study cancer treatment-induced gastrointestinal toxicity that allows rapid, miniaturized, multi-organ toxicity, screening-amenable testing is therefore warranted. The newly developed Tg(cyp2k18:egfp) zebrafish reporter line was found to induce eGFP in the gastrointestinal tract if toxicity was induced in this area. This paper explored utilizing this reporter line for cancer treatment-induced gastrointestinal toxicity, but found that it was not a useful reporter line in this setting. Further progress in this area would be greatly facilitated by the generation of robust and reproducible genetic models of zebrafish intestinal toxicity that mimic the known pathobiological pathways.
Introduction
Gastrointestinal toxicity arising from systemic cancer treatment such as chemotherapy and targeted therapies remains a key reason for treatment cessation, significantly compromising chances of remission. These side effects are frequently associated with traditional chemotherapy drugs, and now are increasingly recognized to be associated with targeted therapies. 1 Patients commonly report gastrointestinal toxicity as being the most impactful on quality of life, influencing their willingness and ability to comply with treatment. 2 This results in dose reductions, interruptions and discontinuation, reducing remission rates. 2
Currently, the animal models used to study gastrointestinal toxicity arising from cancer treatment are primarily rodent models. 3 These models have provided extensive information on mechanisms, and prompted specific hypotheses about the effect of cancer treatment on any regions of the gastrointestinal tract. 3 However, there are challenges associated with rodent models including being prohibitively expensive, time consuming, and the relative difficulty of genetic manipulation. In vitro models are also utilized to study gastrointestinal toxicity. Epithelial cell lines derived from the intestine are cultured as monolayers, in an attempt to mimic the intestinal epithelium. 4 However, there are many limitations to in vitro work, and while they can be useful to understand simple mechanisms, they lack stromal, neural, and immune signaling which is of key importance for modeling gastrointestinal toxicity. Further, they cannot recapitulate the complex systemic metabolism of compounds and thus do not assay the full spectrum of compound derivatives found in vivo. Considering the constraints of in vivo and in vitro models currently used, an alternative model to study cancer treatment-induced gastrointestinal toxicity that allows rapid, miniaturized, multi-organ toxicity, screening-amenable testing is warranted.
Recently, a new zebrafish transgenic eGFP reporter, Tg(cyp2k18:egfp), was developed by identifying highly upregulated genes as biomarkers of liver toxicity. 5 Although this transgenic zebrafish reporter line was initially developed for liver toxicity, upon further testing, it was identified that the line also induced eGFP in the gastrointestinal tract following drug treatment, highlighting this organ as a major detoxification site. Detailed investigations have also reported the intestinal morphology and genetic expression of the zebrafish, demonstrating it as a useful model for gastrointestinal disease models.6–8 Transcriptome profiling has demonstrated that the large anterior portion (58%) of the zebrafish intestine has a similar RNA profile to the human small intestine, followed by a small transitional zone (14%), a tissue resembling the caecum (14%) and lastly a profile resembling the rectum (14%). 8 Zebrafish do not have separated stomach or colon sections, and instead have a continuous tube with segments expressing some genes characteristic of human colon and rectum. 8
Here we exploit this newly developed transgenic zebrafish reporter line to investigate its utility as an alternative vertebrate model to bridge the gap between simple in vitro cellular studies and complex in vivo models for understanding gastrointestinal toxicity induced by SN38 and afatinib. These agents are associated with high levels of gastrointestinal toxicity, manifesting as diarrhea, and represent two classes of anti-cancer agents; classical cytotoxic chemotherapy (SN38, a topoisomerase 1 inhibitor) and targeted agents (afatinib, a pan-HER tyrosine kinase inhibitor).9–13 Thus, this study aimed to assess the efficacy of zebrafish as a platform to study gastrointestinal toxicity second to anticancer chemotherapy and targeted therapy.
Materials and methods
Fish husbandry
Experiments were conducted under the authority of the Institutional Animal Care and Use Committee (IAUCUC) of the Biological Resource Centre, A*STAR (Protocol number 120751), mandated by National Advisory Committee for Laboratory Animal Research Guidelines of the Agri-Food and Veterinary Authority, Singapore.
Zebrafish (Danio rerio) were housed in the zebrafish facility of the Institute of Molecular and Cell Biology, a division of A*STAR, Singapore. Larvae were obtained through natural crosses and staged as previously described by Kimmel et al. 14 Embryos and larvae were raised and treated in water containing 60 µg/mL sea salt (Red Sea Aquatics, UK), and 1% methylene blue.
Zebrafish used included wildtype (AB strain), Tg(cyp2k18:egfp) reporter line and Tg(BACmpx:gfp) reporter line. The development of both the Tg(cyp2k18:egfp) reporter line and Tg(BACmpx:gfp) line have been described previously.5,15
Drug treatment
For treatment of both larvae and adults, SN38 (Tocris bioscience, 2684) was used as the chemotherapeutic agent, and afatinib (AdooQ, A10141) was used as the HER-TKI agent. Both were administered to mimic clinical administration (afatinib orally, and SN38 via intraperitoneal injection). Diclofenac (Sigma D6899) was used as the positive control to cause hepatic and gastrointestinal toxicity reported by Tg(cyp2k18:egfp) zebrafish line (larvae only). Larvae were treated at three days post fertilization (dpf). Drug compounds were dissolved in DMSO, and added to the egg water in a 12-well plate (volume 2 mL/well), with 10 larvae/well. All treatments were conducted in duplicate. Compounds were dissolved in DMSO, and diluted in egg water for larvae treatment. DMSO exposure to larvae or adult fish did not exceed 0.01%. SN38 and afatinib were administered to larvae at a range of concentrations from 0.5 to 900 µM for 24 hours, and optimal dose for treatment was determined as 300 µM. Positive controls (Tg:cyp2k18 line only) were administered diclofenac at a concentration of 13 µM for 24 h as this is known to induce eGFP in this line. 5 Controls were administered the corresponding solvent concentration (DMSO, 0.01%).
Adult zebrafish were weighed and administered 10, 20, 30, and 40 µg/g afatinib via oral gavage, with the maximum volume administered not exceeding 1% of the fish bodyweight. Afatinib was dissolved in DMSO, and diluted in PBS. DMSO exposure did not exceed 0.01%. Controls were administered the corresponding solvent concentrations (DMSO, 0.01%). The oral gavage technique was modified from Collymore et al. 16 A soft, flexible 20 µL ultra-micro tip (eppendorf) was trimmed to 5 cm in total length, and the cut edge was assessed under a dissection microscope to ensure cut edges were blunt, with no beveled or sharp edges. A sponge was cut with a groove and soaked in facility system water to hold zebrafish during gavage procedure. The ultra-micro tip was attached to a pipette and appropriate volume drawn up, ensuring no bubbles were present, and gently inserted into mouth, below the gills. To ensure procedure was effective, Casper transparent zebrafish 17 were gavaged with phenol red (1:10 dilution with a 0.5% phenol red solution in Dulbecco’s PBS). This indicated that the solution was entering the intestinal bulb and not exiting via the gills, and that fish did not expel the solution through their mouth (Supplementary Figure 1). Fish were killed 24 h following gavage.
Adult zebrafish were weighed and administered 10, 20, 30, 40 µg/g SN38 via intraperitoneal injection, with the maximum volume administered not exceeding 1% of the fish bodyweight. SN38 was dissolved in DMSO, and diluted in PBS. DMSO exposure did not exceed 0.01%. Controls were administered the corresponding solvent concentrations (DMSO, 0.01%). Using a 10 µL micro syringe (Hamilton), the correct volume was injected into the fish on the ventral body wall, posterior to the pelvic girdle and anterior to the anus, roughly midway along the length of a pelvic fin. The tip of the needle was pointing rostrally, and was inserted shallowly. Fish were killed 24 h following injection.
Imaging and tissue preparation
For imaging, larvae were anaesthetized with 0.02% Tricane (buffered to pH 7.0) and placed on a Petri dish under a Leica MZ16FA. Only larvae were imaged for eGFP induction as they are transparent.
Adult fish were culled by placing in 0.02% Tricane. A shallow longitudinal incision on the ventral side, from the gills to the anus, was made to remove the intestines which were uncoiled and the anterior 60% of the intestines were fixed in 4% PFA overnight before processing and embedding in paraffin.
Histopathological analysis
Hematoxylin and eosin (H&E) staining was performed on 4 µm sections of intestine, cut on a rotary microtome and mounted onto glass Superfrost® microscope slides (Menzel-Gläser, Braunschweig, Germany). Slides were scanned using a NanoZoomer™ (Hamamatsu Photonics, Japan) and assessed with NanoZoomer Digital Pathology software view.2 (Histalim, Montpellier, France). The occurrences of six histological criteria in the intestine were examined to generate a total tissue injury score. 18 These criteria were villous fusion, villous atrophy, disruption of brush border and surface enterocytes, infiltration of polymorphonuclear cells and lymphocytes, dilation of lymphatics and capillaries, and edema. Each parameter was scored as present = 1, or absent = 0, in a blinded fashion (YZAVS).
Goblet cell analysis
Alcian blue (1% Alcian blue 8GX (CI 74240) in 3% glacial acetic)/periodic acid Schiff staining was performed on 4 µm sections the intestine. Sections were oxidized in 1% periodic acid before washing then treated in Schiff’s reagent. Slides were scanned using a NanoZoomer™ (Hamamatsu Photonics, Japan) and assessed with NanoZoomer Digital Pathology software view.2 (Histalim, Montpellier, France). Data presented as average per villus. All analysis was done in a blinded fashion (YZAVS).
Immunohistochemistry
Immunohistochemical analysis was performed for apoptosis analysis (caspase 3; BD Pharmingen #559565). Change in caspase-3 is a validated marker for altered tissue kinetics in previous models of cancer therapy-induced gastrointestinal toxicities.19–22 Immunohistochemical analysis was performed using Dako reagents on an automated machine (AutostainerPlus™, Dako, Denmark) following standard protocols supplied by the manufacturer. Briefly, sections were deparaffinized in histolene and rehydrated through graded ethanol before undergoing heat-mediated antigen retrieval using an EDTA/Tris buffer (0.37 g/L EDTA, 1.21 g/L Tris; pH 9.0). Retrieval buffer was preheated to 65°C using the Dako PT LINK (pre-treatment module). Slides were immersed in the buffer and the temperature raised to 97°C for 20 min. After returning to 65°C, slides were removed and placed in the Dako AutostainerPlus and stained following manufactures guidelines. Negative controls had the primary antibody omitted. Caspase 3 was quantified by counting the number of positively stained cells. Data presented as average positively stained cells per villus. All analysis was done in a blinded fashion (YZAVS).
Statistics
Data were compared using Prism version 7.0 (GraphPad® Software, San Diego USA). A D’Agostino Pearson omnibus test was used to assess normality. When normality was confirmed, a one-way analysis of variance (ANOVA) with appropriate post hoc testing was performed to identify statistical significance between groups. In other cases, a Kruskal–Wallis test with Dunn’s multiple comparisons test and Bonferroni correction was performed. Differences were considered significant when P < 0.05.
Results
Dose titration
Larvae zebrafish
Zebrafish larvae (n = 10/group) were treated with varying doses of afatinib and SN38, ranging from 100 µM to 1 mM to determine optimal treatment dose. SN-38 did not cause mortality in larvae at any concentration tested. In contrast, afatinib induced dose-dependent mortality (Figure 1). The optimal dose of afatinib for larvae was determined as LC25 (300 µM).
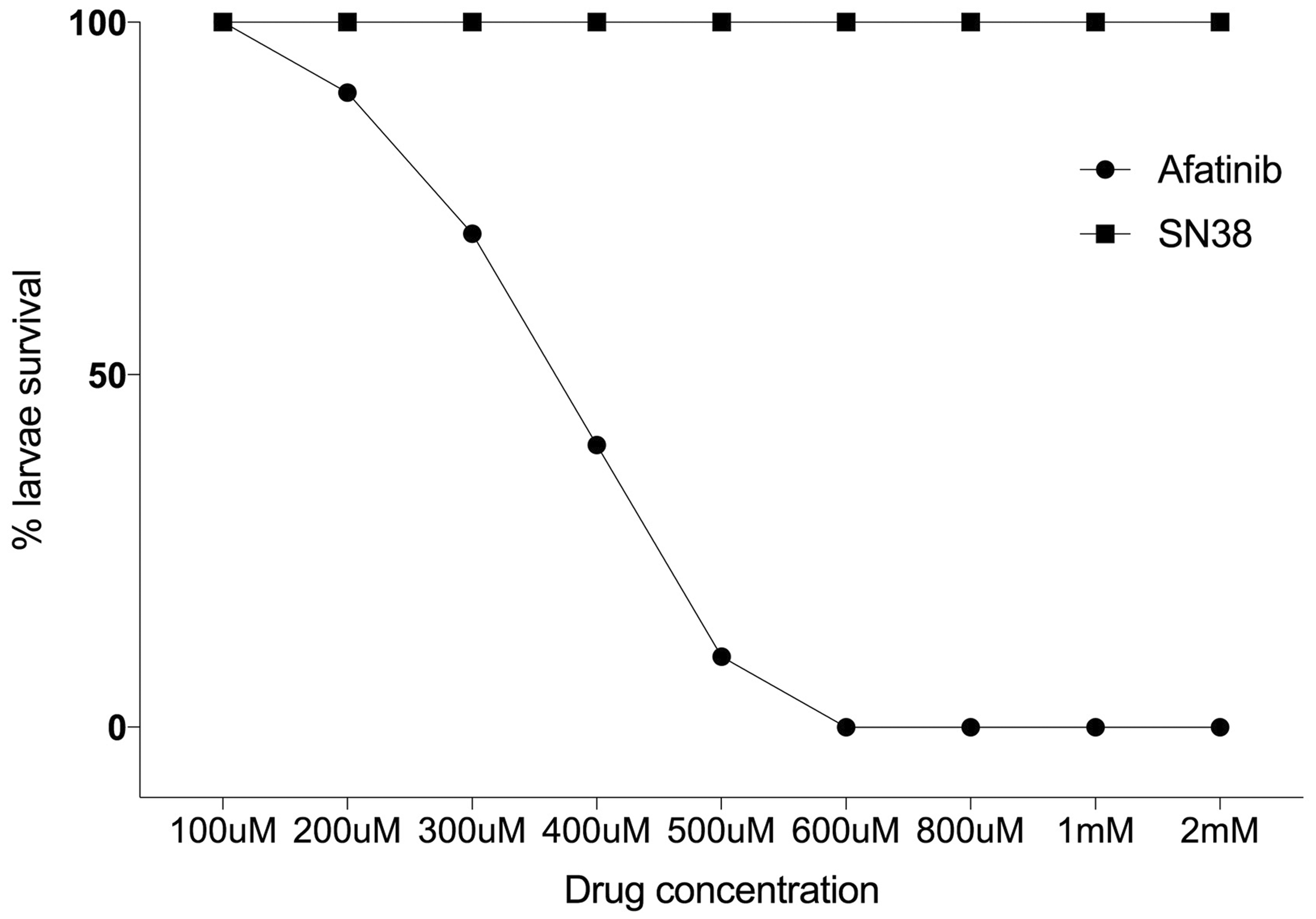
Percent death of larvae. Administration of varying concentrations of afatinib caused lethality in zebrafish larvae. At doses below 100 µM, no larvae died. Doses above 600 µM caused lethality in 100% of larvae. LC25 (dotted line) was 300 µM, and this was determined as the optimal dose for further treatment. SN38 did not cause death at any concentration.
Adult zebrafish
Adult zebrafish (n = 6/group) were treated with SN38 (by intra peritoneal injection) or afatinib (by oral gavage) at 10, 20, 30, and 40 µg/g based on previous publications. 23 SN-38 and afatinib did not cause mortality or noticeable morbidity in adult fish, and hence 40 µg/g was dosed for the remainder of the study.
Transgenic zebrafish larvae fluorescence imaging
To assess the effectiveness of zebrafish Tg(cyp2k18:egfp) reporter line as a high-throughput screening for gastrointestinal toxicity of cancer treatment, larvae were treated with SN38 and afatinib, anti-cancer compounds with known gastrointestinal toxicity profiles; and positive control, diclofenac.
5
Larvae were imaged for induction of eGFP in the gastrointestinal tract of Tg(cyp2k18:egfp) reporter line. As expected, diclofenac induced eGFP in the gastrointestinal tract of the Tg(cyp2k18:egfp) zebrafish larvae (Figure 2(a)), whereas untreated, SN38 and afatinib did not elicit a response (Figure 2(b) to (d)). To assess gut inflammation, the neutrophil reporter line, Tg(BACmpx:gfp) was also exposed to both compounds. Untreated controls had neutrophils primarily contained in the vasculature, with 11.6
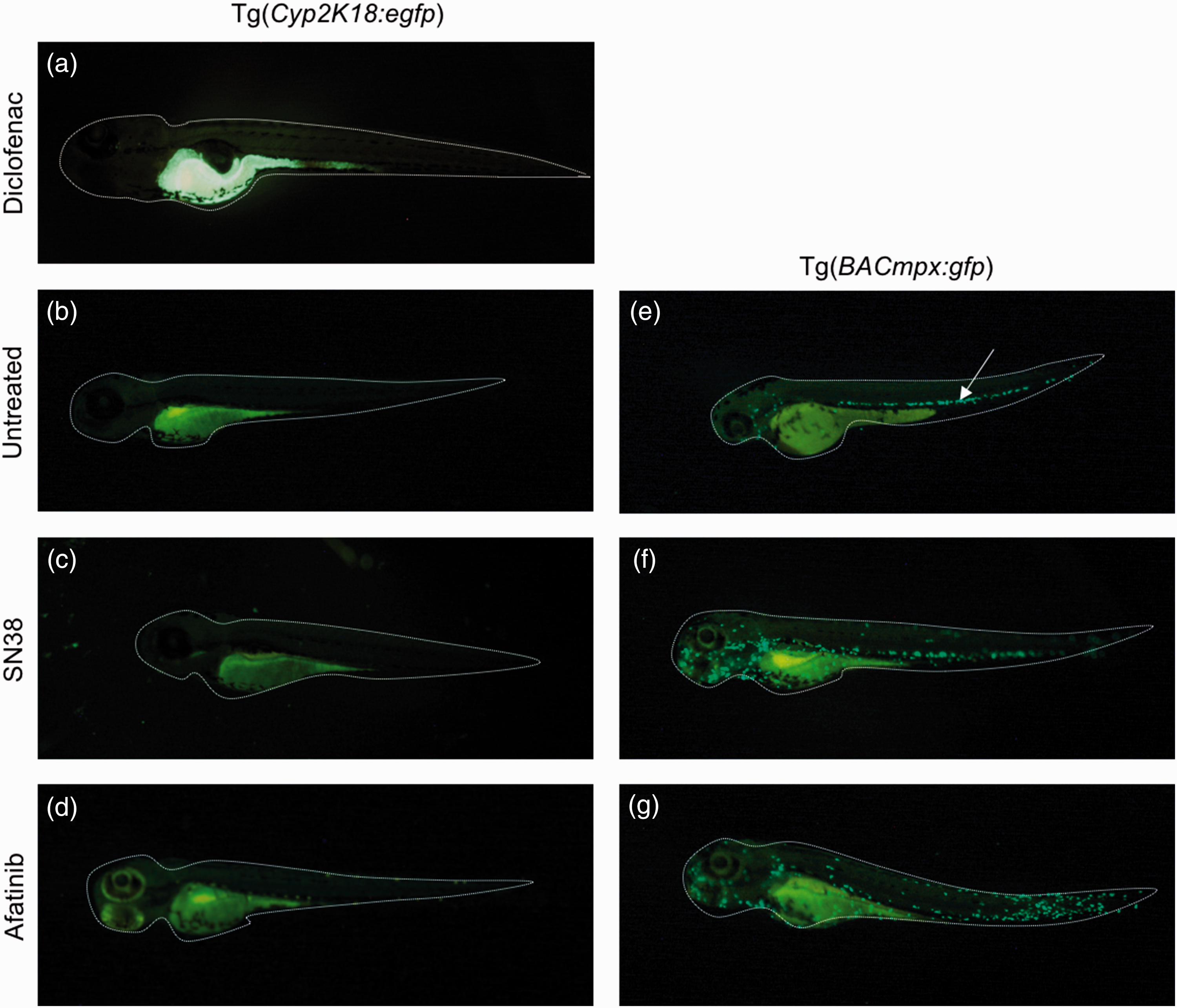
Fluorescent microscope still images of Tg(cyp2k18:egfp) and Tg(BACmpx:gfp) larvae zebrafish at 4dpf. (a) Diclofenac (positive control) induced eGFP in gastrointestinal tract of Tg(cyp2k18:egfp). (b) Untreated larvae did not display eGFP in gastrointestinal tract Tg(cyp2k18:egfp). (c) SN38 treatment did not induce eGFP in gastrointestinal tract Tg(cyp2k18:egfp). (d) Afatinib treatment did not induce eGFP in gastrointestinal tract Tg(cyp2k18:egfp). (e) White arrow: Untreated Tg(BACmpx:gfp) displayed neutrophils contained to the vasculature. (f)SN38 treated Tg(BACmpx:gfp) displayed some neutrophil translocation from the vasculature. (g) Afatinib treated Tg(BACmpx:gfp) displayed neutrophil translocation from the vasculature. (A color version of this figure is available in the online journal.)
Histopathological intestinal analysis in adult zebrafish
To assess the effect of cancer drugs on the histopathology of the gastrointestinal tract, six key markers of damage were assessed in adult zebrafish: disruption of brush border and surface enterocytes, infiltration of polymorphonuclear cells and lymphocytes, dilation of lymphatics and capillaries, edema, villus fusion and villus atrophy. Both lines of adult zebrafish (Tg(cyp2k18:egfp) and wildtype AB) (n = 6/group) treated with afatinib did not display histopathological damage to the intestines (P > 0.05) (Figure 3(a)). Both lines of adult zebrafish (Tg(cyp2k18:egfp) (P = 0.0076) and wildtype AB (P = 0.0407)) treated with SN38 had significantly increased histopathological scores (Figure 3(b) and (c)) compared to controls (Figure 3(d)). The adult Tg(cyp2k18:efp) zebrafish were not different from the wild type (AB) with DMSO control or SN38 or afatanib treatment (P > 0.05). To assess the effect of cancer drugs on goblet cells in the gastrointestinal tract, AB-PAS staining was analyzed. There was no significant difference between either the transgenic (5.736 ± 1.49 cells per villus in DMSO IP; 7.446 ± 1.092 cell per villus in DMSO gavage) or AB controls (5.923 ± 2.06 cells per villus in DMSO IP; 7.625 ± 1.205 cells per villus DMSO gavage). There was no significant difference in fish treated with either SN38 (8.454 ± 1.229 per villus in AB fish; 6.528 ± 2.135 per villus in Tg(cyp2k18:egfp) fish), or afatinib (5.508 ± 1.482 cells per villus in AB fish; 6.392 ± 1.957 cells per villus in Tg(cyp2k18:egfp) fish), (P > 0.05). Caspase 3 staining to identify apoptotic cells in the intestine showed no significant difference between either the transgenic or wildtype line in controls and fish treated with either SN38 or afatinib (P > 0.05; data not shown).
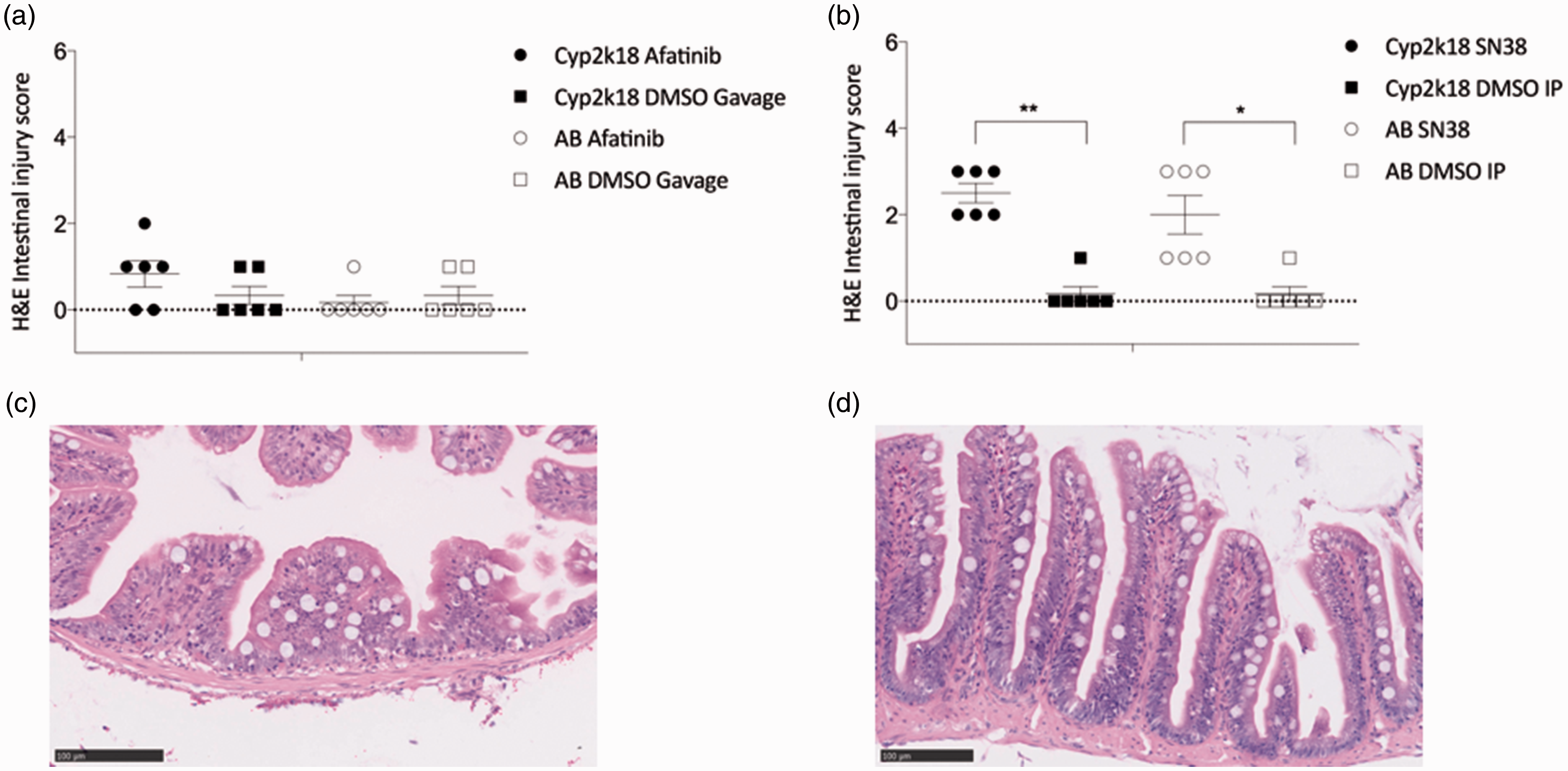
Histopathological injury in zebrafish. (a) Adult Tg(cyp2k18:egfp) and adult AB zebrafish treated with SN38 had significant histopathological intestinal injury compared to controls (**P = 0.0076 and *P = 0.0407, respectively). (b) Adult zebrafish treated with afatinib did not develop histopathological intestinal injury (P > 0.05). (c) Representative image of adult Tg(cyp2k18:egfp) zebrafish treated with DMSO control. (d) Representative image of adult Tg(cyp2k18:egfp) zebrafish treated with SN38. (A color version of this figure is available in the online journal.)
Discussion
Gastrointestinal toxicity is a common side effect of cancer therapies, and can restrict administration dose, and thus treatment efficacy. There is a pressing need for diverse models and assays to better understand the mechanisms and identify toxicity early in drug development. The zebrafish model has been recognized as a powerful tool for human disease modeling, toxicology and drug screening assays, based largely on its ease of handling, fecundity, low cost, rapid external development, amenability to genetic manipulation as well as its genetic similarity to higher vertebrates. 24 There has been minimal use of the zebrafish model to study gastrointestinal toxicity, with one model reporting mucositis findings when developing a hand-foot disease model with PEGylated doxorubicin. 23 Here, we aimed to utilize a newly developed transgenic reporter line as a novel model to screen anti-cancer drugs that induce gastrointestinal toxicity.
The Tg(cyp2k18:egfp) reporter line was developed to screen liver toxicity drugs; however, on testing it was noted that eGFP was also induced in the gastrointestinal tract if toxicity was induced. 5 Non-steroidal anti-inflammatory agents such as diclofenac have long been reported to induce gastrointestinal injury. 25 It was therefore logical to test the ability of this line as a reporter line for cancer treatments that induce both hepatotoxicity and gastrointestinal toxicity. In this model, two agents known to induce severe gastrointestinal-toxicity were used to assess the viability of the Tg(cyp2k18:egfp) line as a high throughput, low-cost screening tool. However, upon testing, it was noted that following treatment with both chosen drugs, SN38 and afatinib, no induction of eGFP was seen in larvae. It is now becoming appreciated that each toxicant is likely to induce a unique set of transcriptional responses and that a global toxic reporter is unlikely to exist. It was noted previously that diverse toxicity reporter lines each responded differently to hepatotoxicants. 5 Ideally any toxicity assay would involve a number of independent reporter lines. Unbiased transcriptional profiling of the toxic response to afatinib and SN38 in the zebrafish might yield novel markers for transgenic reporter development.
A key drawback in using zebrafish for the study of toxicity is that larvae must receive the compound in water. Drug solubility is therefore a major factor to be considered when utilizing this model. While this study overcame this limitation in adult fish by administering drugs via oral gavage or intraperitoneal injection, such approaches are not feasible in larvae nor are adults readily screenable for reporter eGFP induction due to inherent opacity. While it is understood that SN38 and afatinib engage a different pathway for metabolism than Cyp2K18, these agents have been shown to induce hepatotoxicity in other models, the precise mechanisms of which are unknown.26,27 Therefore, the potential confounding issue in this model may be that sufficient concentrations were not achieved to induce liver or gastrointestinal toxicity due to the low solubility, and thus lack of absorption. The solubility of SN38 has previously been identified as problematic, and hence development of soluble SN38 is now being investigated, and may be useful in this model in the future. 28 Nevertheless, afatinib was soluble, and indeed, caused some lethality at doses above 200 µM; however, it is unclear if the lethality was of gastrointestinal origin.
Adult zebrafish treated with afatinib did not show histopathological damage. There are a number of reasons to potentially explain this: (1) histopathological damage is only occasionally reported as a feature of targeted-therapy-induced gastrointestinal toxicity; (2) when histopathology is reported, it is most commonly only in the ileum, and it is possible that given the differences in zebrafish gastrointestinal anatomy this was not seen. The zebrafish lacks a stomach, with the regionalization of the adult zebrafish being more gradual. The morphology of the zebrafish intestine is analogous to the villus structure of mammalian intestines, with capacious folds of the epithelium that protrude into the lumen, increasing the surface area with the finger-like projections, however do not contain crypts 8 ; (3) afatinib is typically administered daily over many weeks, and so a single dose may not cause gastrointestinal toxicity. Further, studies are therefore needed to further investigate this model with a time-course; however, the tolerance of daily gavage to zebrafish is unclear. Adult zebrafish treated with SN38 showed histopathological damage that is consistent with that seen in mammalian models. The well-characterized features of cancer treatment-induced gastrointestinal histopathological include villous fusion, villous atrophy, disruption of brush border and surface enterocytes, crypt loss/architectural disruption, disruption of crypt cell, infiltration of polymorphonuclear cells and lymphocytes, dilation of lymphatics and capillaries and edema. 29
While a robust and tractable genetic model of gastrointestinal toxicity is not available yet, the identification of genes required for the rapid proliferation of zebrafish intestinal epithelial cells during development has highlighted a number of essential genes that could be targeted to disable gastrointestinal toxicity. 30 Moreover, appreciation of the utility of zebrafish to study intestinal inflammation is gaining momentum. In particular, zebrafish provide opportunities to investigate the integrity of intestinal epithelial barrier function, a key emerging marker of gastrointestinal toxicity secondary to cancer treatment, with FITC-dextran gavage. With currently available tools, the interplay between epigenetic regulators, intestinal injury, microbiota composition, and innate immune cell mobilization can be analyzed in exquisite detail. 30 This provides excellent opportunities to define critical events that could be targeted therapeutically. Furthermore, the use of zebrafish larvae as hosts for xenografts of human tissue, while still in its infancy, holds great promise that zebrafish could provide a practical, preclinical personalized medicine platform for the rapid assessment of the drug sensitivity of the patient. Furthermore, there are nascent, yet encouraging, efforts to generate humanized transgenic zebrafish lines for more accurate reconstitution of human drug metabolism. 31
Conclusion
Zebrafish are providing several productive avenues for toxicology research; however, a timely screening model for gastrointestinal toxicity second to cancer treatment is yet to be developed. Here, we aimed to utilize a newly developed transgenic reporter line to address this need; however, the transgenic line did not report gastrointestinal toxicity using the compounds tested. This may be explained due to poor solubility, providing an unclear picture of whether these larvae were induced for any type of injury. We therefore had to concentrate our attention on the adult fish in the direct delivery of the compounds. Chemotherapy agents with high solubility and permeability would be more amenable to these models. In addition, different drugs may invoke highly unique genomic responses, and hence we did not see induction of eGFP in larvae in response to these compounds. Further progress in this area would be greatly facilitated by the generation of robust and reproducible genetic models of zebrafish intestinal toxicity that mimic the known pathobiological pathways in rodents and humans, and can be readily induced in a short time-frame. This would include identification of more gastrointestinal toxicity reporter genes in zebrafish from these and other drugs. Suitably apt models will have the potential to enhance the development of novel cancer drugs by providing a platform for high-throughput chemical screens.
Supplemental Material
Supplemental material for Use of zebrafish to model chemotherapy and targeted therapy gastrointestinal toxicity
Supplemental Material for Use of zebrafish to model chemotherapy and targeted therapy gastrointestinal toxicity by Ysabella ZA Van Sebille, Rachel J Gibson, Hannah R Wardill, Thomas J Carney and Joanne M Bowen in Experimental Biology and Medicine
Footnotes
Authors’ contributions
This research was first published in abstract format “Harnessing transgenic zebrafish as a novel platform for the study of chemotherapy- and targeted therapy-induced gastrointestinal toxicity” in Abstracts of the MASCC/ISOO Annual Meeting 2018 in the Supportive Care in Cancer Journal. 32 All authors participated in the design, interpretation of the studies and analysis of the data and review of the manuscript; YZAVS wrote and received the grants for this study, developed the concept, conducted the experiments and wrote the manuscript, RJG and HRW reviewed manuscript drafts, TJC developed the transgenic reporter line and supplied critical reagents and reviewed manuscript drafts, JMB was involved in developing the concept, and reviewing manuscript drafts.
DECLARATION OF CONFLICTING INTERESTS
The author(s) declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: Ysabella Van Sebille declares that she has no conflict of interest. Rachel Gibson declares that she has no conflict of interest. Hannah Wardill declares that she has no conflict of interest. Thomas Carney declares that he has no conflict of interest. Joanne Bowen declares that she has no conflict of interest.
FUNDING
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was funded by the Florey Medical Research Foundation Project Grant in Cancer Research. Dr Ysabella Van Sebille was supported by The Australian Postgraduate Award, The Global Learning Travel Grant, The Research Abroad Scholarship, and The Walter and Dorothy Duncan Trust Travel Grant.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.