
Abstract
Vitamin D activity is associated with the modulation of a wide variety of biological systems, in addition to its roles in calcium homeostatic mechanisms. While vitamin D is well known to promote gastrointestinal calcium absorption, vitamin D also plays a role in attenuating and/or preventing the progression of several gastrointestinal diseases including Crohn’s disease, ulcerative colitis, and colorectal cancer, and may also play a role in chemotherapy-induced intestinal mucositis. The pro-differentiation, immunomodulatory, and anti-inflammatory effects of vitamin D, which has been reported in numerous circumstances, are key potential mechanisms of action in the prevention of gastrointestinal disorders. While the debate of the effectiveness of vitamin D to treat bone pathologies continues, the clinical importance of vitamin D therapy to prevent gastrointestinal disorders should be investigated given current evidence, using both nutritional and pharmaceutical intervention approaches.
Impact statement
The non-skeletal functions of vitamin D play an important role in health and disease. The anti-inflammatory properties and maintenance of intestinal function fulfilled by vitamin D impact other systems in the body though downstream processing. This review provides insight into the mechanisms underpinning the potential benefits of vitamin D in both maintaining intestinal homeostasis and associated diseased states.
Introduction
The vitamin D endocrine system plays a key role in the maintenance of calcium homeostasis via its actions on several organs, including the intestine to stimulate calcium absorption. 1 However, vitamin D has also been shown to play numerous non-calcemic roles within other biological systems, targeting several organs such as bone, 2 kidney, 3 skin,4,5 brain, 6 and the cardiovascular system. 7 In addition to its intestinal activity to stimulate calcium absorption, vitamin D also regulates intestinal stem cell development, 8 intestinal epithelial differentiation9,10 and barrier function, 10 and intestinal detoxification. 11 Furthermore, vitamin D regulates inflammation and immunity in association with reducing the disease symptoms of several gastrointestinal diseases, including inflammatory bowel diseases (IBD) Crohn’s disease (CD) and ulcerative colitis (UC), 12 and colorectal cancer (CRC). 13 It is plausible that vitamin D may also contribute to reducing chemotherapy-induced gastrointestinal mucositis, based on recent findings for oral mucositis which has similar underlying mechanisms. 14 This review describes the role of vitamin D in preventing gastrointestinal disorders and highlights the potential value in establishing therapeutic strategies to using nutritional or pharmaceutical approaches to enhance vitamin D activity.
Classical actions of vitamin D in intestinal function
Vitamin D, obtained via exposure of the skin to UV light or through nutritional means, undergoes multistep enzymatic conversion to form its active metabolite, 1,25-dihydroxyvitamin D [1,25(OH)2D]. 15 One of the vital roles of 1,25(OH)2D is to maintain the homeostasis of calcium (Ca2+), acting via vitamin D receptor (VDR)-dependent target gene expression regulation. The intestinal absorption of Ca2+ is the most well-known 1,25(OH)2D/VDR-dependent process, which occurs primarily in the proximal intestine. 16 With the exception of pregnancy,17–19 intestinal calcium absorption has been shown to be vitamin D-dependent process. In mice, a network of 1,25(OH)2D-responsive genes in the proximal intestine has been identified to facilitate Ca2+ absorption. 20 The absorption of Ca2+ normally occurs through two defined transport systems: active transcellular transport system and passive paracellular transport system. 11 In transcellular Ca2+ transport, Ca2+ is absorbed through luminal membrane transient receptor potential vanilloid subfamily member 6 (TRPV6), a 1,25(OH)2D-inducible epithelial calcium-selective channel. 11 Cytosolic calcium binding protein, calbindin-D9k (CaBP-9k), is also 1,25(OH)2D-inducible and is involved in cytosolic transfer of Ca2+ to the basolateral membrane. 21 In Vdr–null (Vdr−/−) mice, a marked reduction of intestinal calcium absorption is associated with a 50% and 90% decline in intestinal CaBP-D9k and Trpv6 mRNA levels, respectively.22–24 Cytosolic calcium is then extruded at the basolateral membrane against a concentration gradient by the intestinal plasma membrane ATPase (PMCA1b) 25 (Figure 1). The upregulation of PMCA1b under low dietary calcium condition and by 1,25(OH)2D has been reported previously.26,27
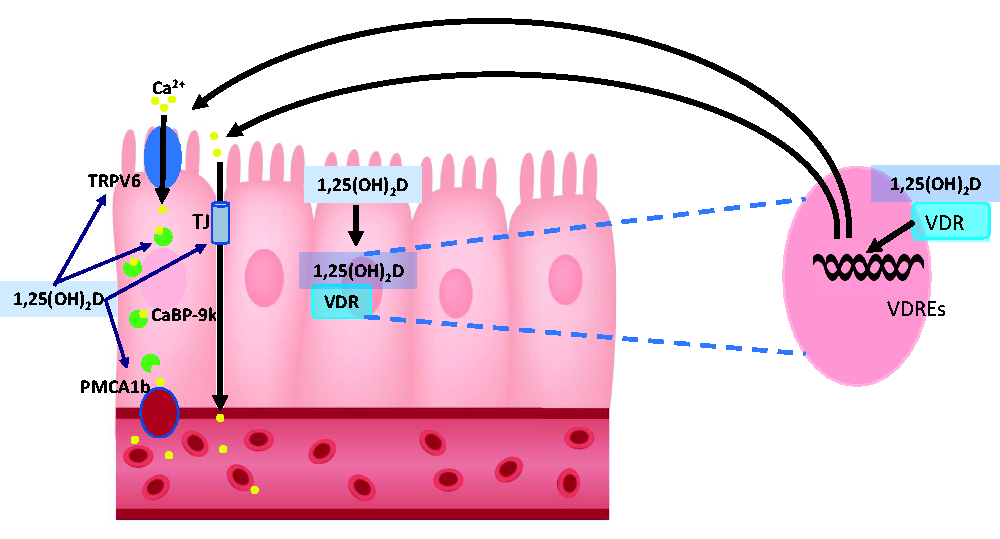
Role of vitamin D in transcellular Ca2+ transport. Ca2+ absorption occurs through 1,25(OH)2D-inducible TRPV6, followed by cytosolic transfer of Ca2+ to the basolateral membrane by calcium binding protein, 1,25(OH)2D-inducible calbindin-D9k (CaBP-9k). Cytosolic calcium is then extruded at the basolateral membrane against a concentration gradient by the intestinal plasma membrane ATPase (PMCA1b). (A color version of this figure is available in the online journal.)
Paracellular Ca2+ transport occurs through intestinal tight junctions. This system is specifically active during the first weeks after birth, and then is gradually replaced by the active transport system. 11 It has been suggested that 1,25(OH)2D can play a mediatory role in transporting Ca2+ in paracellular fashion by influencing the expression of tight junctions28,29 (Figure 1). Expression of claudin-2 and -12 (major transmembrane components of tight junctions) was downregulated in the jejunum, ileum, and colon of Vdr−/− mice, when compared to wild type (WT) mice,28,29 and upregulated by 1,25(OH)2D in Caco-2 cells, 30 suggesting 1,25(OH)2D-dependent regulation of these proteins. Furthermore, the expression of other tight junction-related proteins including cadherin-17, important for cell–cell contact, and aquaporin-8, a tight junction channel, have been reported to be affected by 1,25(OH)2D.28,29 Therefore, these findings suggest 1,25(OH)2D upregulates paracellular calcium transportation, in addition to stimulating transcellular calcium absorption.
Similar to Ca2+ absorption, 1,25(OH)2D regulates intestinal phosphate absorption, through active transport.31–34 Active transport of phosphorus absorption becomes more important under conditions of low phosphate intake and hypophosphatemia. 33 In Vdr−/− mice, hypophosphatemia and compromised phosphate homeostasis were detected.22,24 However, compared to Ca2+ absorption, 1,25(OH)2D is considered to have a less important role in intestinal phosphate absorption, as the human diet is usually rich in phosphate and phosphate absorption is mainly paracellular. 11 Transcellular transport of phosphate is mediated through sodium-dependent phosphate cotransporters. 35 In the intestine, NPT2b (a type II sodium-dependent phosphate cotransporter), expressed on the apical surface of intestinal epithelial cells, is involved in the transcellular absorption of sodium and phosphate. 35 1,25(OH)2D-treated rats (6 μg/kg body weight) showed upregulation of Npt2b, and increased intestinal Na–phosphate uptake. 36 However, studies in mice suggest that the 1,25(OH)2D-mediated regulation of sodium and phosphate can be site- and age dependant.33,37,38 Collectively, 1,25(OH)2D seems, at least partially, to play a role for regulation for phosphate absorption, albeit temporally and spatially.
Vitamin D and intestinal inflammatory disease
Vitamin D has been illustrated to be crucial for reducing the progression of several gastrointestinal diseases, including IBD, CRC, and potentially chemotherapy-induced intestinal mucositis.
IBD
IBD, a group of inflammatory disorders of small intestine and the colon, occur due to complicated interactions between different environmental and genetic factors. UC and CD are two main types of IBD, and manifest as inflammation with distinct, but often overlapping clinical characteristics, and intestinal barrier dysfunction. 39 There are multiple hypotheses describing the etiology of IBD. The hygiene hypothesis is a frequently described key hypothesis, as is barrier dysfunction. The hygiene hypothesis suggests improper exposure to micro-organisms, and excessive hygiene in childhood could lead to abnormal immune response and thus IBD due to disturbance of the intestinal microbiome. A recent modification to the hygiene hypothesis states that vitamin D status may be a crucial environmental factor for IBD incidence/progression, contributing to immune system development and function, 40 and may also contribute to barrier dysfunction through its role in modulating tight junction integrity. 41 However, evidence to suggest a causal link between vitamin D deficiency (or insufficiency) and IBD has yet to be established, despite many studies suggesting that this is the case. In a recent study by Opstelten et al., pre-diagnostic serum 25(OH)D levels were not associated with the development of CD or UC, and neither was dietary vitamin D intake, 42 suggesting vitamin D does not play a significant role in the etiology of IBD. Prior to this, only one prospective cohort study has been carried out, where it was reported that higher vitamin D status was associated with lower risk of incident CD and UC. However, this study used predicted rather than measured serum 25(OH)D levels, raising questions around accuracy. 43 A meta-analysis conducted by Del Pinto et al. showed IBD to be associated with higher odds of vitamin D deficiency, compared with IBD absence. 44 However, this meta-analysis was based entirely on observational studies with confounders not measured, baseline data not adjusted, and results generally not stratified to include important parameters, including surgical procedures, location of disease, or disease activity; therefore, the authors recommend to use the findings cautiously. Vitamin D deficiency may also occur post-diagnostically with IBD, potentially due to vitamin D malabsorption resulting from mucosal damage and can be intestinally impactful, resulting in reduced bacterial clearance, reduced tight junctions, and increased inflammation. 45
The role of vitamin D and its receptor in severity of IBD has been examined in mouse models of IBD. These models include dextran sodium sulfate (DSS)-induced colitis (ranging from 0.5 to 3.5% DSS, driven by innate immune response) in Vdr−/− mice,46,47 2,4,6-trinitrobenzene sulphonic acid (TNBS)-induced colitis (100 mg/kg TNBS, Th1 driven), 48 oxazolone-induced colitis (5 µL/g body weight of 5% oxazolone, similar pathogenesis to UC), 48 interleukin (IL)-10 knockout (Il-10−/−) models, with variables of vitamin D deficiency, and VDR−/− with a high calcium diet,49–51 and T-cell transfer IBD in Il-10−/−, Vdr−/−, or double knockout Il-10−/−/Vdr−/− (DKO) mice.50,52,53 The results showed that the vitamin D deficiency and Vdr−/− mice represented the most severe IBD, while 1,25(OH)2D (or analogs) treatment reduced the severity of the IBD to a mild level in DSS-induced IBD (50 ng/d 1,25(OH)2D in diet or administered rectally starting one week prior and continuing during DSS administration), TNBS-induced IBD (0.5 µg/kg body weight paricalcitol injected intraperitoneally (i.p.) 30 min prior to TNBS, days 1, 3, and 5 following TNBS), 48 and T-cell transfer models, and to a moderate level in Il-10−/− model (0.005 µg/d–0.2 µg/d dietary 1,25(OH)2D from four weeks of age, or from appearance of colitis symptoms). However, a vitamin D analog was not effective in oxazolone-induced colitis (0.5 µg/kg body weight paricalcitol injected i.p. 30 min prior to oxazolone, days 1, 3, and 5 following oxazolone), with no improvement in disease activity, histology, or inflammatory cytokine levels observed. 48
Vitamin D can be beneficial for IBD treatment through multiple mechanisms, including immunomodulatory and anti-inflammatory features, and decreasing disease severity through modulating gut commensal microbiota. 54 Therefore, administration of vitamin D, its analogs, or active metabolites of vitamin D might be a safe and effective therapeutic approach for the treatment of IBD. 12 Two small open label trails (1000 IU/d vitamin D and escalating by 1000 IU/d every two weeks until serum 25(OH)D reached 40 ng/mL, or a dose of 5000 IU/d was reached, 55 and 37 CD patients in remission received two doses of 0.25 µg alfacalcidiol daily for one year 54 ) reported a reduction of Crohn’s Disease Activity Index (CDAI). Similarly, in a large double-blind placebo-controlled study, the IBD relapse rate was insignificantly decreased (p = 0.056) by vitamin D intervention. 56 However, as a standard of care, evaluation of vitamin D status has not been established in patients with IBD, and there are no established guidelines for treatment of IBD patients when vitamin D deficiency is detected. 57
CRC
CRC, which results from malignant transformation of the epithelium of the large intestine, 13 is the fourth leading cause of death from cancer, and third most frequent malignancy worldwide. 58 CRC is driven primarily by mutations resulting in loss of function of the adenomatous polyposis coli (APC) gene, defects in DNA repair, or mutations in β-catenin. 13 APC acts as a tumor suppressor, removing β-catenin from the nucleus. Loss of APC therefore allows accumulation of β-catenin in the nucleus (Figure 2). In vitro studies demonstrate that 10−8 M 1,25(OH)2D, and induced full length APC promoted β-catenin-VDR interactions in HT-29 and Caco-2 cell lines, with greatest interaction occurring when both 1,25(OH)2D and APC were present (p < 0.001), 59 suggesting that 1,25(OH)2D works with APC to stimulate interaction between VDR and β-catenin.
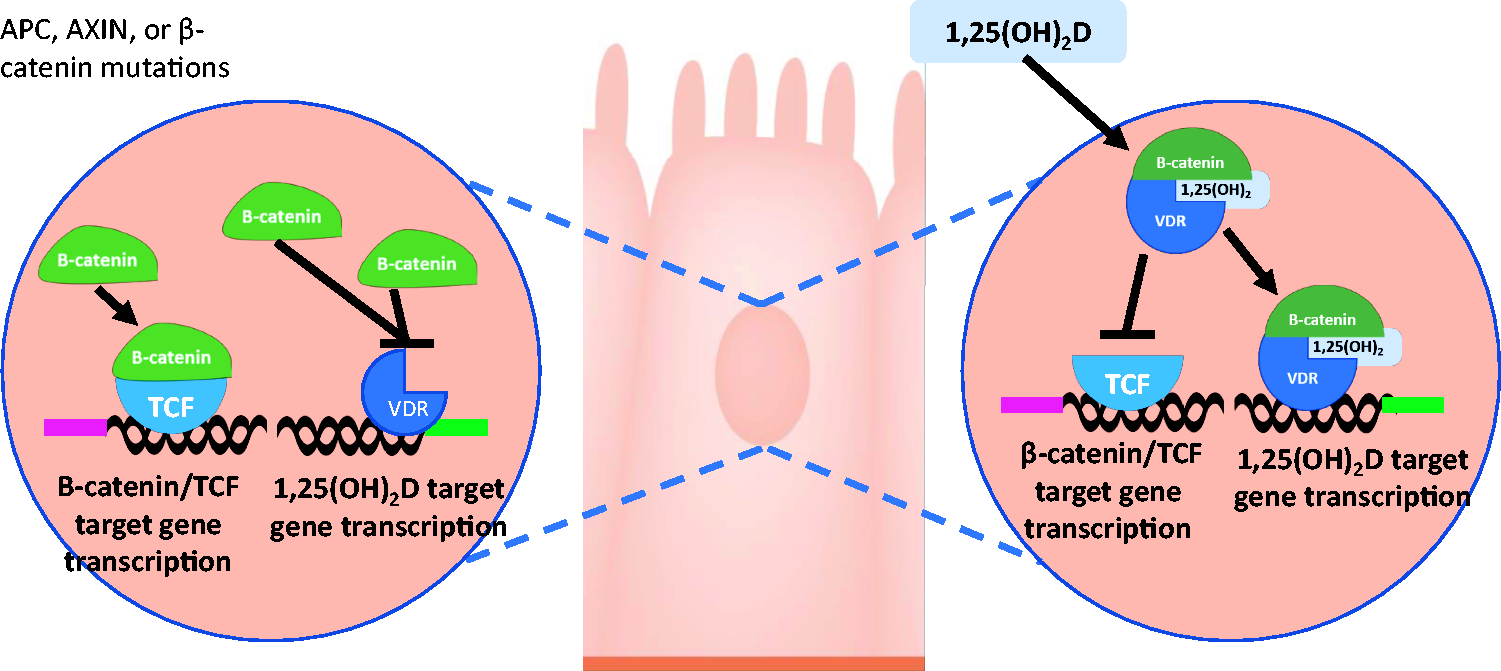
In normal cells (right), one way 1,25(OH)2D inhibits β-catenin/TCF transcriptional activity is by stimulating VDR/β-catenin binding, and inhibiting binding of β-catenin to TCF. In CRC cells (left), when APC, AXIN, or β-catenin genes undergo mutation, and 1,25(OH)2D is reduced, β-catenin accumulates in the nucleus, where it can bind to TCF, upregulating target gene transcription. (A color version of this figure is available in the online journal.)
One of the main signaling pathways involved in CRC progression is Wnt/β-catenin, a suppressor pathway in CRC progression. Wnt/β-catenin is activated as a consequence of mutation of tumor suppressor genes, such as AXIN2 or APC, or CTNNB1/β-catenin oncogene. 13 Abnormal activation of Wnt/β-catenin signaling leads to accumulation of β-catenin proteins in the cytosol, and translocation of some β-catenin into the nucleus, where it binds DNA-bound T-cell factor (TCF), leading to expression of multiple genes inhibited by TCF when the β-catenin is absent. 60 However, vitamin D can interfere with Wnt/β-catenin signaling in CRC cells, acting as an antagonist for Wnt/β-catenin pathway, resulting in enhancement of VDR/β-catenin interaction, reducing binding of β-catenin to TCF 61 (Figure 2). 1,25(OH)2D also upregulates the expression of E-cadherin in CRC cells, suppressing invasion, and can upregulate expression of Dickkopf-related protein (DKK)1, and downregulate the expression of DKK4, Wnt signaling inhibitor and inducer, respectively. 61
Other potential mechanisms of action of vitamin D which interfere with the progression of CRC have been proposed.
Preclinical studies investigating the action of vitamin D on xenografted tumors have shown induction of tumor cell apoptosis by 1,25(OH)2D analogs (1 or 5 µM, six days of exposure), 62 and decreased tumor size in mice injected with MC26 CRC cells with dietary supplementation of 1,25(OH)2D (500,000 IU/kg diet cholecalciferol, 0.47% calcium and 0.3% phosphorus). 63 In xenograft models overexpressing CYP24A1, tumors are more aggressive, and dietary 1,25(OH)2D (2500 IU/kg diet) has little to no effect. 64 There are many clinical studies where inverse correlation between serum level of vitamin D and risk of CRC, or adenoma have been suggested. 61 However, these studies have all claimed limitations and confounding factors, 61 therefore more robust randomized clinical trials are required to establish clear conclusions in this area.
Despite the anti-cancer potential demonstrated for vitamin D, the AMATERASU clinical trial reported no significant improvement of relapse-free survival in post-operative cancer patients with vitamin D supplementation of 2000 IU/d for the duration of the trial. 65 The SUNSHINE phase 2 clinical trial reported no significant difference in median progression-free survival between high (8000 IU/d for chemotherapy cycle 1, followed by 4000 IU/d for remaining cycles) and low-dose (400 IU/d for all cycles) vitamin D supplementation combined with mFOLFOX chemotherapy (2400 mg/m2 5-fluorouracil infusion over 46–48 h, 400 mg/m2 5-FU bolus, 400 mg/m2 leucovorin and 85 mg/m2 oxaliplatin, with 5 mg/m2 intravenous bevacizumab every 14 days) in advanced or metastatic CRC. 66 The link between CRC response to therapy and prognosis and polymorphisms of genes of the vitamin D system (GC, CYP27B1, VDR, CYP2R1, DHCR7 and CYP24A1) have been also reported.67–70 In a clinical study investigating immunohistochemical staining of VDR, nuclear VDR is observed most intensely in normal colonic mucosa, with lower intensity in low-grade and high-grade dysplastic adenoma, and almost no nuclear VDR is observed in CRC. Cytoplasmic VDR is observed most intensely in CRC and high-grade dysplastic adenoma, with lower intensity in normal colonic mucosa. 71
Furthermore, the expression of VDR gene is repressed in advanced stage CRC due to overexpression of EMT genes, particularly SNAIL1 and SNAIL2, believed to facilitate cancer cell metastasis.72,73 Therefore, the benefits and effectiveness of vitamin D may vary in the prevention or treatment of early stage CRC compared with advanced stage CRC.
Intestinal cancer stem cells
Human cancers can be regarded as a stem cell disease, and accordingly, it is hypothesized that CRC is a stem cell-driven tumorigenesis in colon cancer. 74 CRC stem cells are defined by representing similar features to normal colonic stem cells, with ability to self-renew, but also tumorigenic characteristics and ability to aberrantly differentiate into cells that generate tumors. 74 It has been speculated that the first mutation occurs within the normal colonic stem cells located at the base of the crypt, followed by further accumulated mutations over an extended period of time. 74 Once transformed, CRC stem cells can divide symmetrically and asymmetrically to give rise to the cancer cells. 74 CRC stem cells are defined by high expression of cluster of differentiation (CD)44 and CD133, 75 and expression of LGR5, CD26, and epithelial cell adhesion molecule (EpCAM). 76
Vitamin D may directly or indirectly impact CRC stem cell function. As mentioned above, the activation of Wnt/β-catenin signaling plays a crucial role in CRC progression. Similarly, in CD133-positive CRC stem cells, activation and inhibition of Wnt signaling enhanced tumorigenicity, 77 and reduced the expression of CD133, 78 respectively. Previous in vitro studies showed that PRI-1906 and PRI-2191, both vitamin D analogs, modified the gene expression profile of CRC stem cells by reducing anti-apoptotic gene, Bcl-2, and proliferation-related genes including cyclin D1, MYC, BRAF, KRAS, and EGFR. 79 Interestingly, these analogs and PRI-2205 (another vitamin D analog) were reported to downregulate the expression of CRC stem cell-related genes including OCT3/4, EpCAM, NANOG, Notch1, and C-X-C chemokine receptor type 4 (CXCR4). 79 By affecting the microenvironment of CRC, vitamin D can also indirectly affect the function of colon cancer stem cells, as VDR has been detected in patient-derived colon normal fibroblasts, and colon cancer associated fibroblasts, 80 suggesting the potential role of inhibiting colon cancer stem cells via regulation of stromal fibroblasts. Therefore, vitamin D/VDR signaling may affect CRC stem cell function through regulation of Wnt/β-catenin signaling and/or interactions within CRC microenvironment.
Cancer treatment-induced mucositis
Mucositis is a frequently reported side effect in cancer patients receiving cancer treatments (chemotherapy and/or radiotherapy, and/or targeted therapies, and/or immunotherapies), occurring in the mouth (oral mucositis, OM) and/or intestine (gastrointestinal mucositis, GM). However, therapeutic approaches are currently very limited, and specific to tumor types and treatment regimens. 81 Studies in rats have demonstrated the progression of intestinal pathology to consist of chemotherapy-induced crypt hypoplasia, followed by rebound crypt hyperplasia, then re-establishment of normal tissue.82–84
It has been reported that nuclear factor kappa-light-chain-enhancer of activated B cells (NFкB) signaling plays a substantial role in intestinal mucositis progression, as peaks in NFкB expression were demonstrated to precede peaks in pro-inflammatory cytokines in intestinal tissue following chemotherapy. 85 It is also speculated that the upregulation of pro-inflammatory cytokines drives positive feedback by activating NFкB, amplifying the injury.86,87 Following irradiation, NFкB has also reported to upregulate cyclooxygenase 2 (COX2), an enzyme with pro-inflammatory characteristics. 88 Overexpression of COX2 was detected in submucosal fibroblasts and endothelial cells of the oral mucosa 10 and 16 days after targeted irradiation. 88 Association of other pro-inflammatory cytokines, including tumor necrosis factor (TNF)-α, IL-1β, and IL-6 were reported in the progression of chemotherapy-induced gastrointestinal mucositis.89–91 In addition, two clinical studies performed on oral (buccal) mucosal biopsies following chemotherapy demonstrated that basal layer apoptosis was increased in the tissues, 92 and the loss of tissue architecture was attributed to the high level of NFκB and COX2 expression. 93 Therefore, inflammation is implicated as a key player of chemotherapy-induced mucositis, and therapeutic approaches targeting anti-inflammatory and immunomodulatory pathways, such as vitamin D analogs, might provide a novel treatment for affected individuals.
Vitamin D has been widely shown to possess immunomodulatory and anti-inflammatory features. 94 The Vitamin D/VDR signaling pathway is important in mitigating the progression of intestinal diseases, including IBD and CRC. 95 Studies in Cyp27−/− mice (1,25(OH)2D deficient) demonstrated significantly increased levels of inflammatory cytokines IL-1α, IL-1β, IL-6, and IL-8 in Cyp27−/− mice, suggesting a role for 1,25(OH)2D in maintaining low levels of these cytokines under normal conditions. 10 In addition, studies in VDR−/− mice show increased expression of TNF-α and IL-1β following induction of colitis. 96 Therefore, it is logical to hypothesize that vitamin D-related treatments might also be beneficial for reducing, if not preventing, chemotherapy-induced intestinal mucositis incidence and severity, where these cytokines are prevalent. Previous preclinical studies have shown promising effects of anti-inflammatory agents for attenuating the severity of mucositis in animal models.97–99 Moreover, vitamin D deficiency or insufficiency has been detected in CRC patients undergoing chemotherapy, 100 and 1,25(OH)2D treatment in combination with calcium carbonate showed a reduced proliferative index and increase of VDR expression in colonic mucosa. 101
Vitamin D may also diminish chemotherapy-induced intestinal mucositis through its effects on gut microbiota. It has been shown that chemotherapy alters microbiome composition throughout the gastrointestinal tract, shifting from predominantly commensal bacteria to a more pathogenic profile, specifically Salmonella spp. and Escherichia coli.101–104 Cyp27B1−/− mice (unable to produce active vitamin D) and Vdr−/− mice (lacking capacity for vitamin D to bind) have significantly higher levels of Proteobacteria and Bacteroidetes phyla, often reported to be associated with diseased states, and significantly lower levels of Firmicutes phyla, often reported to be associated with good health outcomes, compared to WT littermates. 105 The general overall finding when taking studies into consideration is that the gut microbiota are responsive to vitamin D deficiency, and also vitamin D supplementation, potentially by regulating gut immune responses to regulate the microenvironment where commensal bacteria reside. Vitamin D supplementation may therefore be a promising new pathway to investigate in terms of reducing the burden of chemotherapy-induced intestinal mucositis in affected patients through contributing to modulation of the gut microbiome response to chemotherapy and other cancer treatments.
Intestinal stem cell development, epithelial cell differentiation, barrier function, and detoxification
Intestinal stem cell development
Potential roles for vitamin D in intestinal stem cell fate have been reported in a variety of studies. Lgr5+ crypt base columnar cells, providing stem cell-associated functions, have the ability to differentiate into all cell lineages in the villi. 8 Vitamin D/VDR signaling pathway was found to be an essential signaling pathway for Lgr5+ stem cell functions, as Ki67 (cell proliferation marker)-negative Lgr5+ cells were detected in the crypt base of VDR-deactivated mice, suggesting vitamin D may promote intestinal cell maturation. 8 In addition, by comparing the level of protein expression in the intestine of Vdr−/− and WT mice, it was demonstrated that vitamin D, directly and/or indirectly, regulates proteins involved in intestinal epithelial cell proliferation, migration and stress response. 106 It is also well accepted that exposure to 1,25(OH)2D has a profound influence on intestinal stem cell function, thought to be the cell of origin of intestinal tumors. 107 Wnt/β-catenin signaling may also be a crucial pathway for intestinal stem cell development. 108 Therefore, it is logical to hypothesize that vitamin D plays an important role in intestinal stem cell development, potentially by regulating Wnt/β-catenin signaling in these cells. These findings suggest that vitamin D/VDR signaling is likely to be important for intestinal stem cell development.
Epithelial cell differentiation and barrier function
Vitamin D may also be effectively involved in differentiation of epithelial cells. 1,25(OH)2D promotes intestinal epithelial cell differentiation by regulating caudal-related homeobox transcription factor 2 (CDX-2) activity. CDX-2 plays a vital role in intestinal development by mediating intestinal cell differentiation 109 and is considered as a tumor suppressor gene in CRC. 110 Studies have shown that CDX-2 protein binds the human VDR gene promoter, inducing the transcription of VDR gene. 111 Interestingly, a polymorphism in the CDX-2 binding site of the VDR promoter negatively affects the transcription and the activity of VDR and is believed to be associated with CRC occurrence.112,113 In addition, 1,25(OH)2D has been reported to enhance the formation of microvilli by increasing the expression and activity of several brush border enzymes, including maltase and alkaline phosphatase, regarded as differentiation markers in the small intestine. 11 1,25(OH)2D has also been shown to enhance the expression of a plasma membrane calcium ATPase isoform associated with differentiation, through analysis of alkaline phosphatase activity in CRC cells. 114
A well-differentiated intestinal epithelium requires a proper pre-established barrier between the internal environment of the host, and the lumen, providing protection against infection. This barrier is constituted of several adhesion structures, including adherens junctions, gap junctions, tight junctions, and desmosomes between adjacent epithelial cells. 11 1,25(OH)2D is actively involved in maintaining the function of the epithelial barrier by regulating the expression of proteins involved in the establishment of the barrier. In addition, 1,25(OH)2D maintains the differentiated adhesive phenotype of intestinal epithelial cells by upregulating several adherens junction proteins, such as E-cadherin, and tight junction proteins including claudin-2, -7 and -12, occludin, and zonula occludens (ZO)-1 and -2.30,115–117 1,25(OH)2D was shown to increase the mucosal transepithelial resistance,47,117 an indicator of barrier integrity and mucosal permeability, and to restore intestinal tight junction integrity following damage by bacterial lipopolysaccharide (LPS) in vitro. 118 Furthermore, to maintain barrier integrity, 1,25(OH)2D was reported to regulate TNF-α-induced permeability of intestine 119 and block TNF-α-induced intestinal cell apoptosis. 120 Therefore, vitamin D may play a key role in maintaining the integrity and permeability of the intestine.
Intestinal detoxification
Vitamin D may contribute to detoxification of toxic and chemical compounds, maintaining gut homeostasis. Expression of a wide range of enzymes, including phase I metabolic enzymes (3α- and 17β-hydroxysteroid dehydrogenases, CYP1A1, CYP3A1, CYP3A3, CYP4F5, CYP2B2, and CYP3A9) phase II metabolic enzymes (sulfotransferases and UDP-glucuronosyltransferases), and antioxidant enzymes (epoxide hydrolase, glutathione peroxidase, S-transferases, γ-glutamyl transpeptidase, and Pi-class glutathione) can be regulated by 1,25(OH)2D. 35 In CRC cells, 1,25(OH)2D induces the expression of CYP3A4, recognized as the most important drug metabolizing enzyme in humans. 121 Lithocholic acid (LCA) is a secondary bile acid that causes DNA damage and inhibits DNA repair in colonic cells and is found in CRC patients, which has led to its use as an experimental CRC inducer. 122 LCA can be removed by SULT2A1 (a phase II sulfotransferase), CYP3A4, and the multidrug resistance-associated protein (MRP) 3, whose expressions are induced by 1,25(OH)2D.123,124 In addition, VDR can function as a receptor for LCA, although with a lower affinity than 1,25(OH)2D, 125 promoting SULT2A1, CYP3A4, and MRP 3 through positive feedback to eliminate LCA. 61 Collectively, by regulating the expression of enzymes involved in intestinal detoxification, 1,25(OH)2D contributes to removal and/or modification of harmful substances from the intestine.
Fine tuning of immune system
In addition to its classic effects in calcium absorption and mineralization, great attention has been given to the effects of vitamin D on the immune system. VDR is expressed on immune cells, including B cells, T cells, and antigen presenting cells, enabling these cells to synthesize active vitamin D metabolites, and thus act by autocrine and/or paracrine mechanisms in an immune environment. 94 It is known that macrophages can recognize bacterial LPS through their toll-like receptors (TLR), and upon binding, increase expression of 1-α-hydroxylase and VDR, and formation of 1,25 D-VDR-RXR heterodimer, leading to the production of antimicrobial peptides. 126
Vitamin D inhibits proliferation of B cells and blocks B-cell differentiation and maturation. 127 It also suppresses T-cell proliferation, 128 and induces production of anti-inflammatory cytokines, such as IL-10, by affecting T-cell maturation129,130 and facilitating induction of T regulatory (Treg) cells. 131 Other immune cell types, such as monocytes and dendritic cells, are also affected by vitamin D. Vitamin D not only inhibits production of inflammatory cytokines, such as TNFα, IL-1, IL-6, IL-8, and IL-12 by monocytes, 132 but also inhibits differentiation and maturation of dendritic cells. 127
In IBD, it has been suggested that vitamin D activates innate immunity by regulating the production of antimicrobial peptides, 133 and suppressing associated inflammation and adaptive immunity. 134 Vitamin D-deficient mice exhibit suppression of the colonic expression of angiogenin (ANG)-4, which is an antimicrobial peptide produced by Paneth cells. 135 As the dysregulated innate immune response to gut microbiota has been proposed to initiate tissue inflammation in some types of IBD, 136 vitamin D may have a preventive effect for IBD through the induction of -4 against the enteric bacteria. 134 Antimicrobial peptides could be a therapeutic approach for IBD, suggested by studies administering antimicrobial peptides to mice. 137 Furthermore, aberrant migration of CD8+ α–α cells in the gut has been demonstrated in mice administered antimicrobial peptides, which could suggest enhanced risk of IBD in these animals. 53 Vitamin D has been previously shown to reduce the symptoms of IBD in transgenic mice by increasing calcium intake, resulting in strengthening the mucosa barrier 138 ; therefore, increasing 1,25(OH)2D-mediated calcium absorption may play a role in suppressing the progression and severity of IBD. However, the extent to which vitamin D and its receptor modulate the immune system and whether this has any role in triggering or preventing immune disorders, such as IBD is yet to be elucidated.
Regulation of inflammatory pathways
Vitamin D can regulate inflammatory pathways in the gut, and thus can potentially reduce the burden of gut diseases. Prostaglandins (PG), such as PGE2, are synthesized by prostaglandin-endoperoxide synthase 2 (PTGS2), also known as COX2. These PG are important stimulators of inflammation, leading to regulation of cell activation, cell migration,139,140 and promotion of colon cancer cell proliferation.141,142 Studies have shown that 1,25(OH)2D decreased prostaglandin levels and inhibited COX2 expression in prostate cancer cells, 143 and although not yet reported in colon cancer cells, there is likely be a similar effect. In colon cancer cells, TNF-α can promote Wnt signaling 144 and activate NF-κB pathway. 145 IL-6 treatment of colon cancer cells resulted in increased cell proliferation, 146 and upregulation of MMP2 and MMP9, proposed to facilitate metastasis of cancer cells.147,148 Similarly, the treatment of CRC cells with IL-8 not only increased proliferation of cancer cells, but also migration and metastasis, and induced angiogenesis.149,150 However, in mouse models of CRC, 1,25(OH)2D was demonstrated to inhibit the activity of NF-κB and increased the expression of NF-κB inhibitor (IκB). 151 This phenomenon results in inhibition of pro-inflammatory gene expression, including COX2, IL-6, IL-8, and TNF-α, which biologically possess anti-apoptotic and pro-tumorigenic properties. 152 Production of other inflammatory cytokines, including IL-17, might also be regulated by 1,25(OH)2D. Although it remains to be investigated in CRC, it has been suggested that 1,25(OH)2D can inhibit synthesis of IL-17 by immune cells. 153 IL-17 is produced by both Paneth cells and immune cells, and elimination of these cells in a CRC mouse model showed the prevention of metastasis and reduction of pro-inflammatory cytokine gene expression (such as IL-1β, COX2, and IL-6), and thus attenuation of tumor progression.154,155 Taken together, these findings suggest that vitamin D inhibits the production of inflammatory cytokines (Figure 3), reducing inflammation, which may attenuate the severity/progression of intestinal diseases.
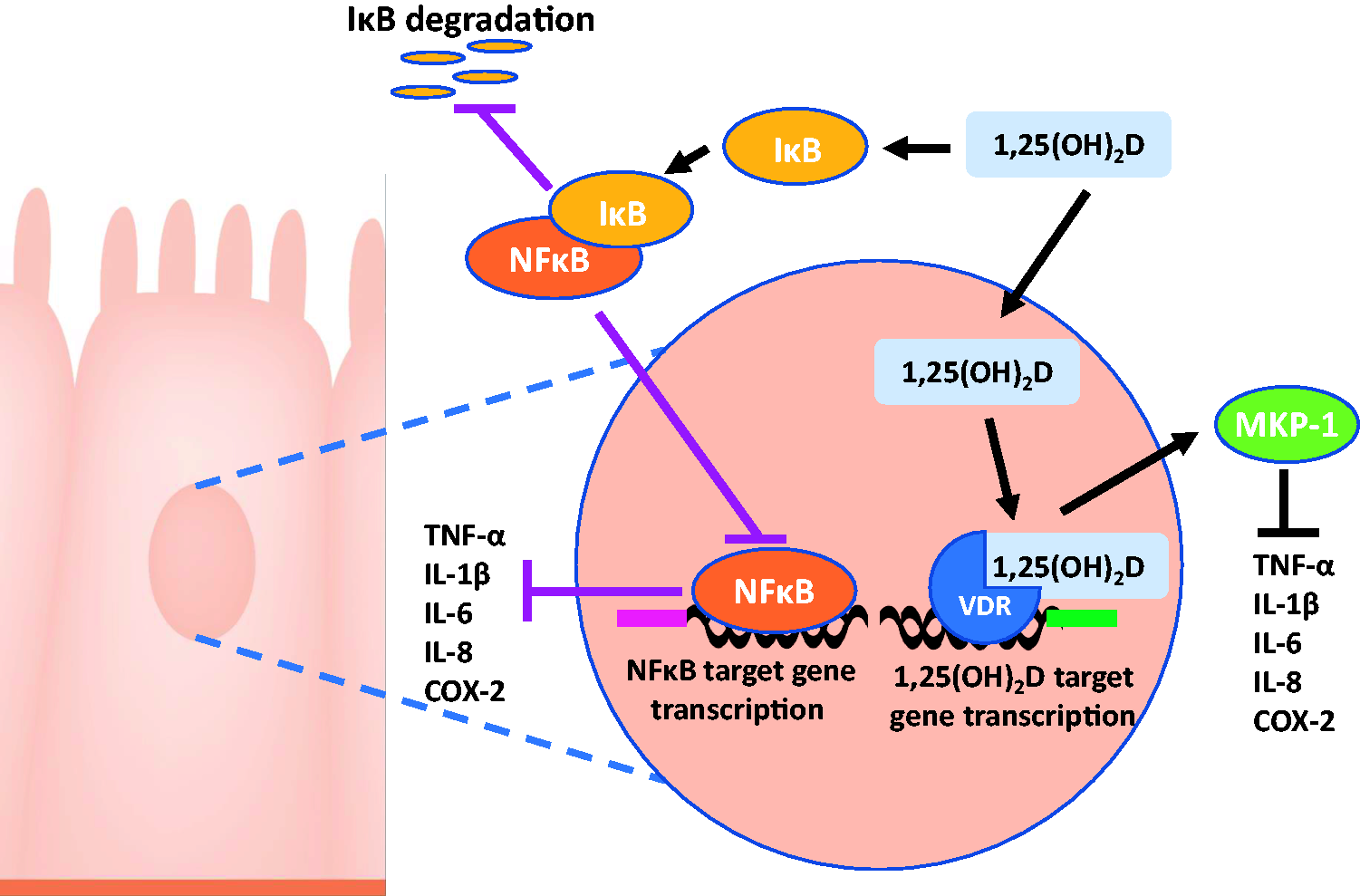
1,25(OH)2D binds to VDR, stimulating production of MKP-1, inhibiting pro-inflammatory cytokines. 1,25(OH)2D also stimulates production of IκB, inhibiting NFκB, also inhibiting production of pro-inflammatory cytokines. (A color version of this figure is available in the online journal.)
Regulation of intestinal microbiota
Regulation of the immune response to the intestinal microbiota and changes to the intestinal microbiota are additional biological features of vitamin D, which is mediated by the host.41,105 Cyp−/− or Vdr−/− mice have been shown to have more bacteria present from the Proteobacteria and Bacteroidetes phyla, and less bacteria from the Firmicutes phylum compared to WT. Moreover, these mice are more susceptible to colitis compared to WT. 105 Previous studies have shown differences in bacterial genera, including Bacteroides, Salmonella, and Eubacterium, between Vdr−/− and WT mice, 156 and a study by Jin et al. suggested that the dysbiosis-induced alterations in crucial pathways of the gut flora in Vdr−/− mice may affect gut detoxification, cancer, infection, and other diseases. 157 Moreover, a vitamin D-deficient diet in mice showed an aggravated dysbiosis, intestinal barrier dysfunction, and inflammation, 158 while treatment with 1,25(OH)2D was suggested to raise Citrobacter rodentium load in the colon. 159
VDR−/− and Cyp27B1−/− mice showed an increased susceptibility to colitis as the result of gut microbiota alteration. 105 This study further demonstrated increase in rates of Proteobacteria and colitis-causing Helicobacteraceae family members. Vitamin D supplementation in human and Vdr−/− mice, on the other hand, reduced Proteobacteria and enhanced the number of beneficial micro-organisms including the members of the Firmicutes phyla.105,160 Vitamin D metabolism can be regulated by gut microbiota as shown in germ-free (GF) mice, where either the conventionalization, or colonization of GF mice with specific commensals enhanced serum 24,25(OH)2D, and 25(OH)D levels to those of conventional mice, and reduced fibroblast growth factor (FGF)-23 levels. 161 Conversely, infection of GF mice with Citobacter rodentium (a murine pathogen) decreased serum 24,25(OH)2D and 25(OH)D levels. 161 Blocking FGF-23 in GF mice increased serum 1,25(OH)2D levels, suggesting that FGF-23 may play a role in the microbial regulation of vitamin D metabolism. 161
In a clinical study where seven vitamin D-deficient patients with ileocolonic CD in remission were administered 20,000 IU/d cholecalciferol orally from days 1 to 3, then every other day for a total treatment time of four weeks, there was a specific shift in intestinal microbial communities to a potentially beneficial profile, with increased Actinobacteria, Firmicutes, and Bacteroidetes, while no microbial changes were observed in healthy controls, 162 suggesting that administration of vitamin D may only be effective in modulating intestinal microbiota when microbial composition is altered. Other clinical studies revealed that circulating 25(OH)D levels increased following oral supplementation with probiotic Lactobacillus reuteri NCIMB 30242. In addition, the level of circulating 25(OH)D was reported to be associated with differences in stool microbial composition, as the abundancy of Roseburia, Blautia, Ruminococcus and Dorea (Firmicutes phylum, Clostridia class) decreased in people with higher 25(OH)D concentration. 163 In addition, by analyzing the genome of approximately 1800 individuals, VDR gene was identified as a host factor that impacts the gut microbiota. 164
A recent systematic review showed that 22 out of 24 studies reviewed demonstrated an association between vitamin D and gut microbiota, and similarities in findings between mouse and human studies. 165 This remains a significantly understudied area, as many human studies are observational only, and reported observational findings in humans suggest a mutual interaction between vitamin D and gut microbiota. Therefore, adequately powered studies are required to determine the mechanisms of association that contributes to maintenance of intestinal homeostasis.
Conclusions
Vitamin D is involved in orchestrating various physiological processes, such as regulating intestinal microbiota, and calcium and phosphate absorption. In addition, vitamin D plays a substantial role in reducing or preventing several pathological conditions, including bone diseases, IBD, CRC, and chemotherapy-induced intestinal mucositis. Prevention or attenuation of the aforementioned conditions may be attributed to the immunomodulatory and anti-inflammatory characteristics of vitamin D. However, more preclinical and clinical studies are required to address the mechanisms of action by which vitamin D can be beneficial for particular pathological condition, and provide the optimum dosage of vitamin D, or its analogs, for each particular condition.
Footnotes
Authors’ contribution
MH conducted literature searches and wrote the majority of the article. PA wrote sections on skeletal functions of vitamin D, and edited article. CS contributed to the writing of the article, and edited article, AS contributed to the detailed planning of article content, writing of the article, and editing of the article.
DECLARATION OF CONFLICTING INTERESTS
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
FUNDING
CS is supported by a University of South Australia Postgraduate Award (USAPA).