
Abstract
We provide a review of Spectrin isoform function in the cytoplasm, the nucleus, the cell surface, and in intracellular signaling. We then discuss the importance of Spectrin’s E2/E3 chimeric ubiquitin conjugating and ligating activity in maintaining cellular homeostasis. Finally we present spectrin isoform subunit specific human diseases. We have created the Spectrinome, from the Human Proteome, Human Reactome and Human Atlas data and demonstrated how it can be a useful tool in visualizing and understanding spectrins myriad of cellular functions.
Impact statement
Spectrin was for the first 12 years after its discovery thought to be found only in erythrocytes. In 1981, Goodman and colleagues 1 found that spectrin-like molecules were ubiquitously found in non-erythroid cells leading to a great multitude of publications over the next thirty eight years. The discovery of multiple spectrin isoforms found associated with every cellular compartment, and representing 2-3% of cellular protein, has brought us to today’s understanding that spectrin is a scaffolding protein, with its own E2/E3 chimeric ubiquitin conjugating ligating activity that is involved in virtually every cellular function. We cover the history, localized functions of spectrin isoforms, human diseases caused by mutations, and provide the spectrinome: a useful tool for understanding the myriad of functions for one of the most important proteins in all eukaryotic cells.
Introduction
The discovery of nonerythroid spectrin in 1981, by Goodman and colleagues 1 , has led to a large number of publications describing first its structure and functions within the plasma membrane and cytoplasm and then within the nucleus of eukaryotic cells. As spectrin contributes 2-3% of the total protein in eukaryotic cells2–4, is found associated with every organelle and major cytoskeletal component 5 , and has its own E2/E3 chimeric ubiquitin conjugating ligating activity6–9, it should come as no surprise that this ubiquitous scaffold protein belies a myriad of cellular functions that reside within the nucleus and cytoplasm and integrate these cellular compartments. In this minireview, we introduce the concept of the spectrinome, or spectrin interactome, and explain how this is a useful construct to think about its integrative functions. We will raise questions that should keep those interested in spectrin busy for decades to come. We begin the discussion by presenting the earliest discovered spectrin that was found within the erythrocyte on the cytoplasmic surface of the plasma membrane.
The Erythrocyte Membrane Skeleton
Erythrocyte spectrin was first isolated from human red blood cells (RBCs) by Marchesi and Steers Jr. in 1968 10 . They said “The functional role of this protein is unknown, but it appears to be involved in maintaining the structure of the red cell membrane”. Over the next 20 years, the structure and function of the RBC spectrin membrane skeleton was elucidated (Figure 1). This membrane skeleton does, as suggested by Steers and Marchesi, provide structural integrity to the erythrocyte, but also is responsible for its elasticity and flexibility that allows a biconcave disc of 8 μm diameter to continuously pass through a circulatory system that narrows to less than 2 μm in venules.
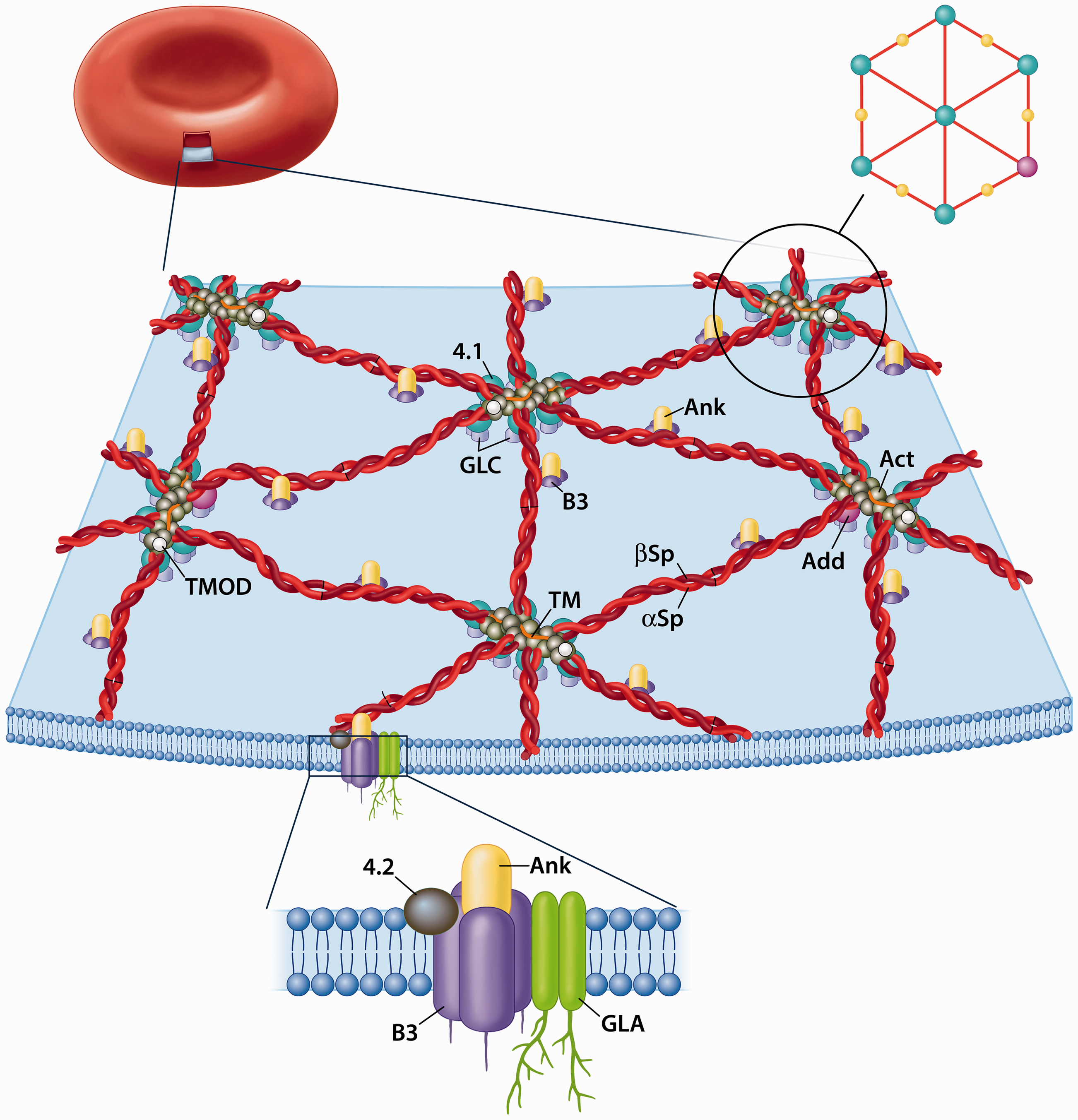
The Red Blood Cell Membrane Skeleton
RBC Spectrin is composed of an α and β subunit of 280 kDa and 246 kDa respectively. The simplest form of RBC spectrin is an antiparallel αβ heterodimer, but on the cytoplasmic surface of the plasma membrane it primarily takes the form of an (αβ)2 tetramer formed by head-to-head interaction of two heterodimers.11,12 As such the two tail regions of the heterodimer are chemically equivalent and can bind and crosslink short actin protofilaments (∼14 g actin monomers long).13–15 The binding of spectrin to these short actin protofilaments is strengthened by the binding of protein 4.116–19 and adducin20–23 to the tail regions, each forming its own ternary complexes with spectrin and actin. Electron micrographs of spread membrane skeletons demonstrated a hexagonal array (Figure 1) with actin protofilaments at the vertices and center, interconnected by spectrin tetramers. The RBC spectrin tetramers isolated at low ionic strength and visualized by negative staining and electron microscopy appear as flexible rods that are 200 nm long13–15. As will be discussed below these rods can condense to 55-65 nm. This provides the membrane skeleton and plasma membrane with its properties of elasticity and flexibility. The spectrin tetramers, and hence the membrane skeleton are associated to the bilayer through spectrin-ankyrin-band 324–28 (another early name for ankyrin was syndeins 25 ) and spectrin-4.1-glycophorin C interactions29,30 (with accessory proteins) as shown in Figure 1.
Spectrin Isoforms
In 1981, Goodman and colleagues described “Spectrin-like” proteins in embryonic chicken cardiac myocytes (ECCMs), cultured mouse fibroblast 3T3 cells, and rat hepatoma HTC and HMOA cells. 1 They immunoprecipitated two peptides of 240kDa and 230 kDa apparent molecular weight from the ECCMs, with anti-RBC spectrin antibody. These spectrin-like proteins were stained with spectrin antibody on immunoautoradiography and generated similar chymotryptic peptides to the α and β subunits of embryonic chicken RBC α and β spectrin 1 . Goodman’s description of nonerythroid spectrin was confirmed by multiple laboratories in 1982.31–34 This seminal finding led to a large number of publications from many laboratories bringing us to our current understanding that it is present in all eukaryotic cells.
One of the first tissues that was explored for the structure, location and function of spectrin was brain.33,35,36 Several laboratories isolated brain spectrin that contained α and β subunits with apparent molecular weight of 240 kDa and 235 kDa.33,35,36 Reiderer et al 37 prepared two antibodies against [1] mouse brain spectrin, which were isolated from an enriched fraction containing synaptic/axonal membranes, and [2] mouse RBC spectrin. They then cleaned these antibodies against the reverse antigen: [1] mouse RBC spectrin and [2] mouse brain spectrin isolated as described above. 37 These antibodies that had very high specificity to their own antigen were used by Reiderer et al 37 to determine localization of these two spectrin antigens in mouse cerebellum. What was found was that the brain spectrin antibody stained axons and presynaptic terminals, while the RBC spectrin antibody stained specifically the soma, dendrites, and post synaptic terminal of all neurons. 37 This was the very first description of distinct spectrin isoforms, which Reiderer et al called brain spectrin 240/235 and brain spectrin 240/235E (for erythroid). 37 As will be described below, today these isoforms would be called αSpIIΣ1/βSpIIΣ1(nonerythroid spectrin isoform) and αSpIΣ1/βSpIΣ2 (the erythroid spectrin isoform). Over the next five to six years Goodman, Reiderer, Zagon, Zimmer and colleagues would go on to demonstrate that this differential localization was true for neurons throughout the central nervous system5,37–43 and that these two isoforms had very different expression during brain ontogeny.44–46 The nonerythroid spectrin isoform was present at the earliest stages of prenatal mouse brain development and constantly increased during brain ontogeny while the erythroid spectrin isoform arose beginning in the second week of mouse brain postnatal development44–46.
A major step in the localization of the brain spectrin isoforms was the first immuno-electron microscopic study on mouse cerebellum by Zagon et al. 5 As described by Zagon et al, 5 and shown in Figure 2, spectrin isoforms were compartmentalized as described by Reiderer el al. 37 At the electron microscope level of resolution, we found the spectrin isoforms not only on the cytoplasmic surface of the plasma membrane, but also on the cytoplasmic surface of all organelle membranes (nuclear envelope, endoplasmic reticulum, mitochondria, etc.). The spectrin isoforms were also in the cytoplasm crosslinking cytoskeletal elements (actin filaments, intermediate filaments and microtubules) to each other and to membrane surfaces. We also observed granular staining within the nucleus. Brain spectrin 240/235 was in the presynaptic terminal staining the plasma membrane, cytoskeletal structures and small spherical synaptic vesicles, while spectrin 240/235E was found in the post synaptic terminal associated with the plasma membrane, mitochondria, cytoskeletal elements and post-synaptic densities. 5
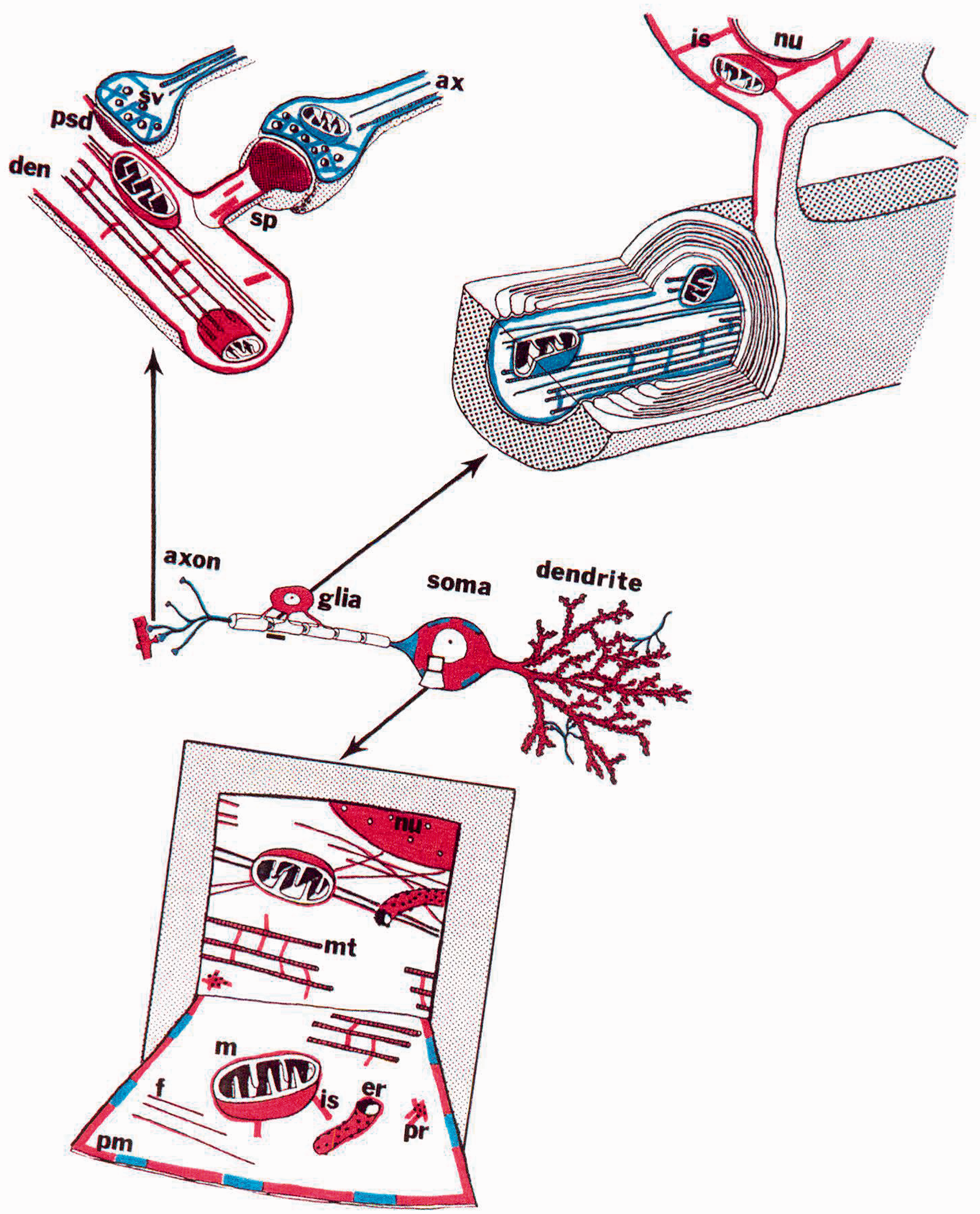
Immunoelectron microscopy location of spectrin isoforms
Today we know that there are two alpha spectrin genes, SPAT1 and SPTAN1, that encode the erythroid (αSpI) and nonerythroid (αSpII) spectrin isoforms respectively. There are also five beta spectrin genes SPTB, SPTBN1, 2, 4, 5 which encode the erythroid (βSpI) and nonerythroid (βSpII, III, IV, and V) spectrin isoforms. Table 1 gives the protein nomenclature, names in the literature, molecular weight, human gene symbol, and human chromosome locus for the seven spectrin genes and their products. Alternate splice forms of the spectrin genes give rise to isoforms indicated with the symbol Σ. Note that a common alternate name for the non-erythroid spectrin is Fodrin. 47 This name was coined when Levine and Willard found two axonally transported proteins of 240 kDa and 235 kDa apparent molecular weight that failed to react with spectrin antibodies. 47 Spectrin was given several other names before the recognition of its relationship to spectrin.
Spectrin Isoforms

The nonerythroid spectrin isoforms are found in all eukaryotic cells, except RBCs. The erythroid spectrin isoforms are found in RBCs, neurons, cardiac and skeletal muscle.2–4,34,48 The spectrin tetramers are usually composed of either complexed erythroid α and β subunits or non-erythroid α and β spectrin subunits. 49 Clark et al demonstrated that a smaller percentage of spectrin tetramers in brain tissue are hybrid where αSpI complexes with βSpII, III, IV, or IV or αSpII complexes with βSp1Σ2. 50
Spectrin Structure
As described above, erythrocyte spectrin is found in situ primarily as (αβ)2 tetramers, although higher order oligomers exist. 51 This is also true for the nonerythroid spectrins. 4 In both cases head to head linkage of two heterodimers forms a tetramer with chemically equivalent tail regions. As shown in Figure 3, the heterodimers contain α and β subunits that are antiparallel with the N-terminus of the β subunit and the C-terminus of the α subunit at the tail regions of the tetramer. The original protein sequencing of a segment of this αSPI followed by secondary structure determination defined a spectrin repeat unit of approximately 106 amino acids. 52 Several X-ray crystallography studies53–55 have indicated that the spectrin repeats maintain the 3 dimensional structure shown in Figure 3A. What is presented in this Figure are two adjacent αSPI repeats 16 and 17. Repeat 16 shown in Figure 3 A contains three α helices, where Helix A (dark blue) and C (green) are parallel and B (light blue) is antiparallel. As can be seen in the illustration Helix B is a little longer than the other two helices and is kinked in the middle due to a conserved proline residue. Repeat 16 and 17 are associated through the association of Helix C in repeat 16 with Helix A in Repeat 17 (also colored green to show the continuity). Helix A and B in each Repeat are associated by coiled flexible regions. The individual spectrin repeats are separated by regions which can bend in αSPI and βSpI and less so in αSpII and βSpII. Atomic Force Microscopy has indicated that the erythrocyte spectrin molecule can condense down to approximately 55-65 nm,56,57 and it has been recently suggested that it does so by creating a quaternary structure resembling a Chinese finger trap. 58 The ability of erythrocyte spectrin tetramers to condense and expand, from 55 to 200 nm is what provides the erythrocyte membrane skeleton with its properties of elasticity and flexibility.
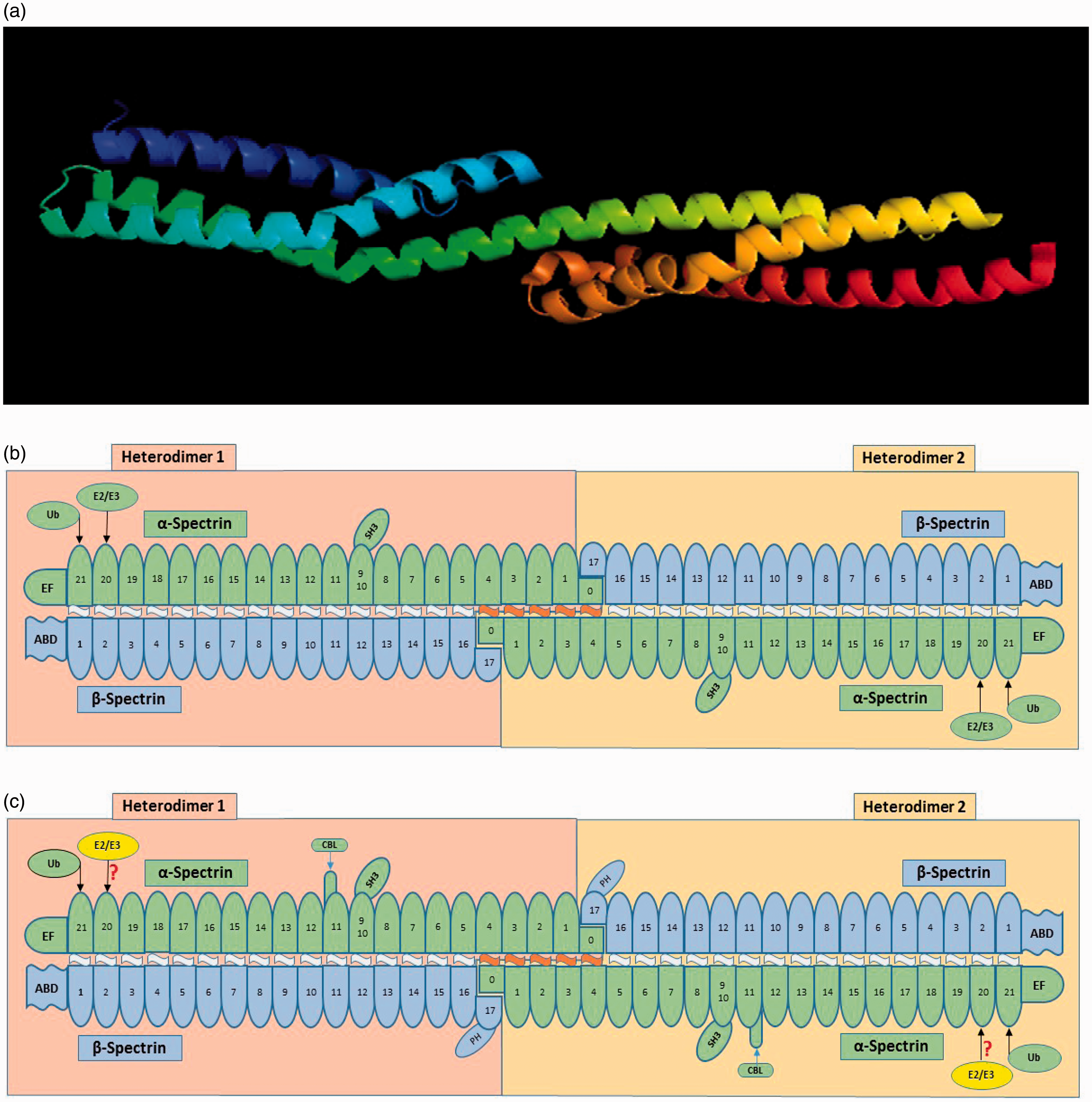
Spectrin Structure
The complete sequence of mouse and human erythrocyte αSpI and βSpI59–61 and nonerythroid αSpII and βSpII47,62–65 have been determined by cloning and DNA sequencing. αSpI, the α subunit of human αSpIΣ1/βSpIΣ1, contains 2419 amino acids and has a mass of 280,014 daltons. As shown in Figure 3b, αSpI has 20 spectrin canonical repeats while repeat 21 contains EF Hand domains between residues 2271 and 2386. EF Hand domains bind Ca2+. 66 Between repeat 9 and 10 there is an SH3 domain involved in signal transduction.67–69 Interestingly, in repeat 20 there is a E2 ubiquitin conjugating site with 70% identity to all known E2 active sites and the canonical cysteine (C2071) that can transfer ubiquitin to itself in a lysine rich segment of repeat 21.6–9 Spectrin’ s chimeric E2/E3 ubiquitin conjugating/ligating activity was demonstrated to target other spectrin binding proteins (ankyrin, protein 4.1, 4.2, the anion transport channel and an unknown protein gi13278939) as well.70,71 This will be discussed in far more detail below. αSpI also has an N-terminal sequence that does not have the typical repeat structure (labeled 0) that associates with the C-terminal sequence of βSpI (labelled 17) creating the head-to-head linkage creating the tetramer.
The βSpI sequence, beta erythrocyte spectrin βSpIΣ1, contains 2,137 residues and has a molecular weight of 246,468 daltons. βSpIΣ1has 16 canonical repeats where repeat 17 has a pleckstrin homology (PH) domain. 72 PH domains are involved in cell signaling, cytoskeletal organization and phosphinositide binding. Goodman’s laboratory found that the N-terminal, non-repeat, segment of βSpI contains the actin-binding domain (ABD). 73 They demonstrated that residues 47 to 186 contains the binding site, and that this sequence was highly conserved in other actin binding proteins such as dystrophin, α-actinin, and many others. 73 Consistent with this seminal work, later studies confirmed the ABD and found two Calponin Homology (CH) domains within this ABD region, which appear to bind f-actin. 74 CH1 (residues 1-160) can bind f-actin on its own; but CH2 cannot bind f-actin on its own, unless the first 20 amino acids are truncated (leaving residues 191-280).75–77
αSpII, or nonerythroid spectrin αSpIIΣ1, contains 2,472 residues and has a molecular weight of 284,539 daltons. As shown in Figure 3C, αSpIIΣ1 has 21 spectrin repeats with similar EF Hands, SH3, E2/E3-Ubiquitin-Conjugating/Ligating and target sites as αSpιΣ1 described above. What is unique is the Calmodulin Binding Loop (CBL) that is a 36 amino acid insert that has a Ca2+-dependent calmodulin binding site and can be cleaved by calpain and caspases 2, 3 and 7.78–83 This cleavage is blocked by phosphorylation of tyrosine1176.80,81 The overall sequence identity for αSpIIΣ1 and αSpιΣ1 is approximately 60%.
βSpII or nonerythroid spectrin βSpIIΣ1 contains 2,364 residues and a molecular weight of 274,699 daltons. βSpII, like βSpI, has 17 spectrin repeats with a similar ABD and PH. They share about 60% sequence identity. The C-terminus of βSpIIΣ1 extends 207 amino acids beyond the end of βSpI. Ma et al 64 who performed the sequence analysis on mouse found a hydrophobic section of βSpIIΣ1, that has very high sequence homology with the heme binding domain of globins. This laboratory went on to demonstrate that non-erythroid spectrin has the ability to bind hemin in vitro. 64 This observation could be of major importance to Oxygen and Nitric oxide localization and function intracellularly and is worthy of follow up.
Constructing the Spectrinome
As spectrins are known to be multifunctional scaffold proteins located both in the nucleus, cytoplasm and all cytoplasmic facing membranes (including the plasma membrane), we felt it would be of great value to build a spectrinome using existing human interactome data and then placing the proteins within either the nuclear or cytoplasmic compartments using the human atlas data.
The Homo sapiens gene-gene interaction and Homo sapiens protein-protein interaction files were downloaded from the Reactome Pathway Database (https://reactome.org/). 84 In addition to the Reactome files, we downloaded the Hallmark and C2 pathways from the Molecular Signatures Database from the Gene Set Enrichment Analysis set created by the Broad Institute (http://software.broadinstitute.org/gsea/msigdb/collections.jsp). 85 The files were combined to create a list of all Homo sapiens interactions. 86 We created a program in C++ to collect all interactions that involve SPTA1, SPTAN1, SPTB, SPTBN1, SPTBN2, SPTBN4, or SPTBN5. Each subset of reactions were saved into a csv file. The seven files were then combined to create a master list of all seven spectrins. These files were used to create an edge and vertex list for each interaction graph. The graphs were created in R using ggnet2. Secondary interactions were gathered from the reactome database using the previous C++ program, and used to create secondary interaction graphs for SPTA1 and SPTAN1. Locations of each protein inside human cells were extracted from the Human Protein Atlas (https://www.proteinatlas.org/). 87 The proteins were separated into four categories for the interaction graphs: nucleus, cytoplasm, both areas, and spectrin.
The 1st order interactions and 2nd order interactions for SPTA1 (αSpI) and SPTAN1 (αSpII) displayed in Figure 4 are extremely informative. 1st order are those proteins that interact directly with the spectrin isoform. 2nd order interactions are proteins that interact with the 1st order set of interacting proteins. Some of the major points are: [1] both erythroid and nonerythroid α spectrins can bind all five β spectrin gene products, but cannot associate with each other. As mentioned above, while the primary form of spectrin tetramers are formed from either α and β erythroid or α and β nonerythroid subunits, Goodman’s laboratory demonstrated that hybrid forms are also present in neurons. 50 [2] The β-spectrin isoforms cannot associate with each other, but can self-associate. [3] SPTA1 (αSpI) and SPTAN1 (αSpII) associate with different sets of proteins in the nucleus, in the cytoplasm, and proteins in both compartments. Therefore, it is quite likely that all seven spectrin gene products will ultimately be found in both compartments. Many of these localizations have already been demonstrated. [4] The 1st order inner ring α-spectrin isoform interacting proteins are the same, except for the other spectrin isoform (Table 2). This will be discussed further below. [5] The outer ring interactions for SPTA1 (αSpI) and SPTAN1 (αSpII) are different except for the following common interactions: ADD1, CASP3, CDC42, COL4A3, EVL, EZR, F3, FGA, KDM4B, TANK, and UBE21. [4] The 2nd order interactor proteins for SPTA1 (αSpI) and SPTAN1 (αSpII) display very different interactomes. A very prominent difference is the dense protein cloud surrounding polyubiquitin (UBC). Ubiquitin has a well-documented association with SPTA1 (αSpI).6–9,88–90 Sangerman et al reported the association of ubiquitin with SPTAN1 (αSpII) in neurons, 90,91 but this association does not appear in the interactome database. This is an important point. While the spectrinome is very useful, it will become even more so when the protein and interaction databases are more complete. The current human Reactome Pathway database contains approximately 16,000 proteins which represents roughly three quarters of the human proteome (see Table 3). As the number of proteins and interactions grow in these databases, the Spectrinome will become increasingly robust. The current reactome database is very accurate, but not 100%. Occasionally it assigns interactions to both α- and β-spectrin subunits when the assignment should be to only one. An example are the ANK interactions which should only be assigned to the β-spectrin isoforms. On rare occasions there is a miss-assignment. An example is synapsin I (SYN1) which associates with SPTBN1, not SPTAN1. The Spectrinome is dependent on the accuracy of the machine learning tools that scan existing literature, the yeast two hybrid accuracy, and the expertise of those curators who review the data. The Spectrinome is currently of enormous value and its coverage and accuracy will improve with advances in machine learning.

The α Spectrinome
Spectrinome 1st Order Interactions
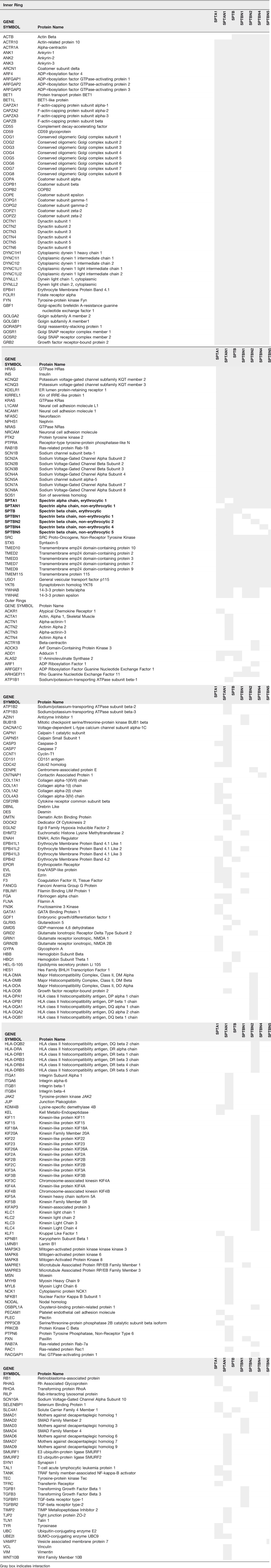
Gray box indicates interaction
Protein Nodes and Interactions
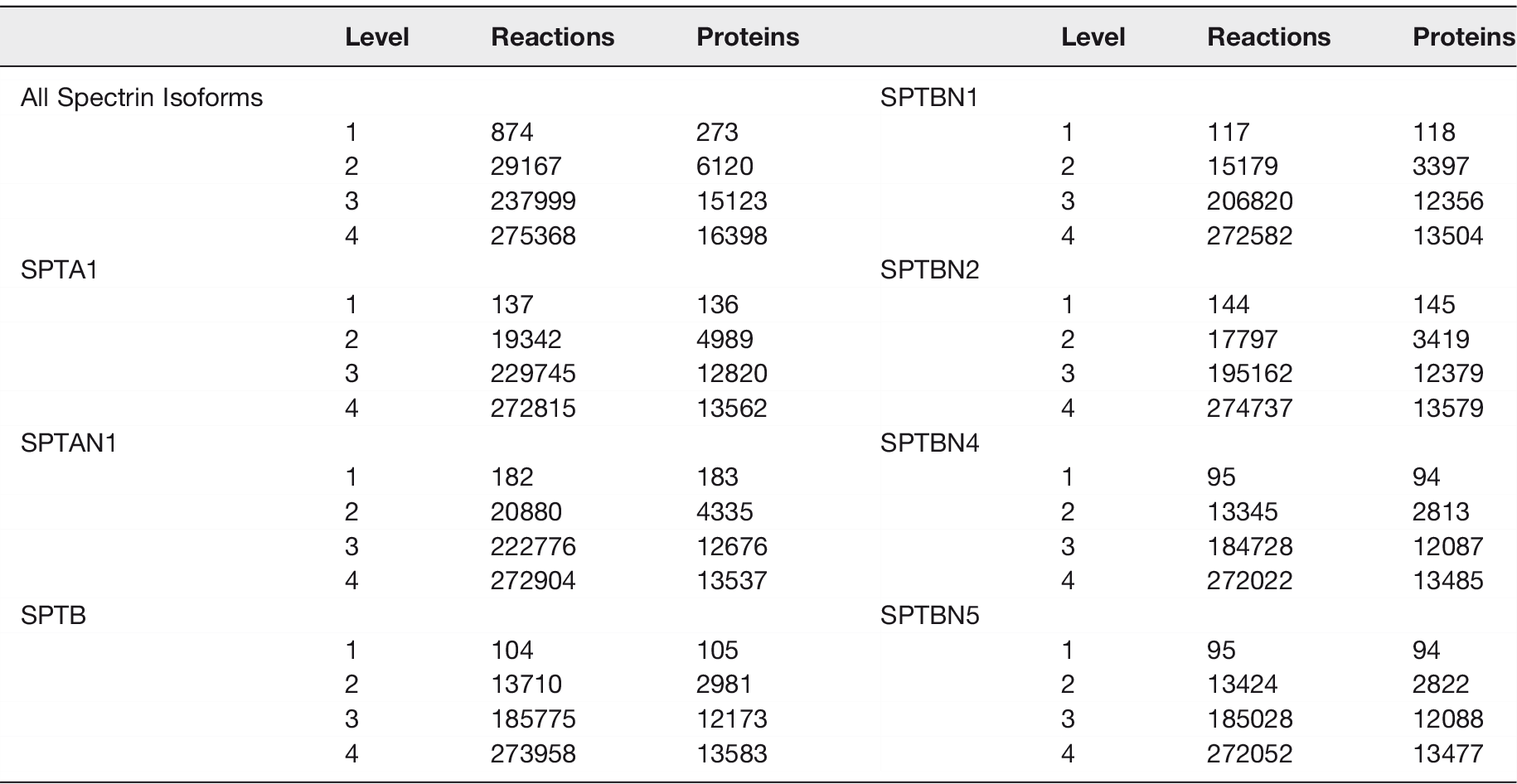
Figure 5 contains the 1st order interactomes for five β spectrin gene products. The five beta spectrin genes SPTB, SPTBN1, 2, 4, and 5 encode the erythroid (βSpI) and nonerythroid (βSpII, III, IV, and V) spectrin isoforms. The major points observed are: [1] all five beta gene products associate with both SPTA1 (αSpI) and SPTAN1 (αSpII). [2] The interacting proteins for each beta are found in the nucleus, cytoplasm and both compartments. As spoken of above this indicates that all five are likely to be found in both compartments. [3] Many of the 1st order interacting proteins for the five beta spectrins are in common, but the following are not: the β-spectrin isoforms cannot associate with each other, and all do not bind with YWHAB and YWHAE. [4] The outer ring β-Spectrin isoforms interactions are unique except for ARHGEF11 (Table 2).

The β Spectrinome
The complete 1st order Spectrinome was created by linking the two α spectrin and five β spectrin interactomes together. A remarkable visual picture emerges as shown in Figure 6. We provide a list of first order binding partners for all seven individual spectrin gene products in Supplemental Lists 1 to 7.
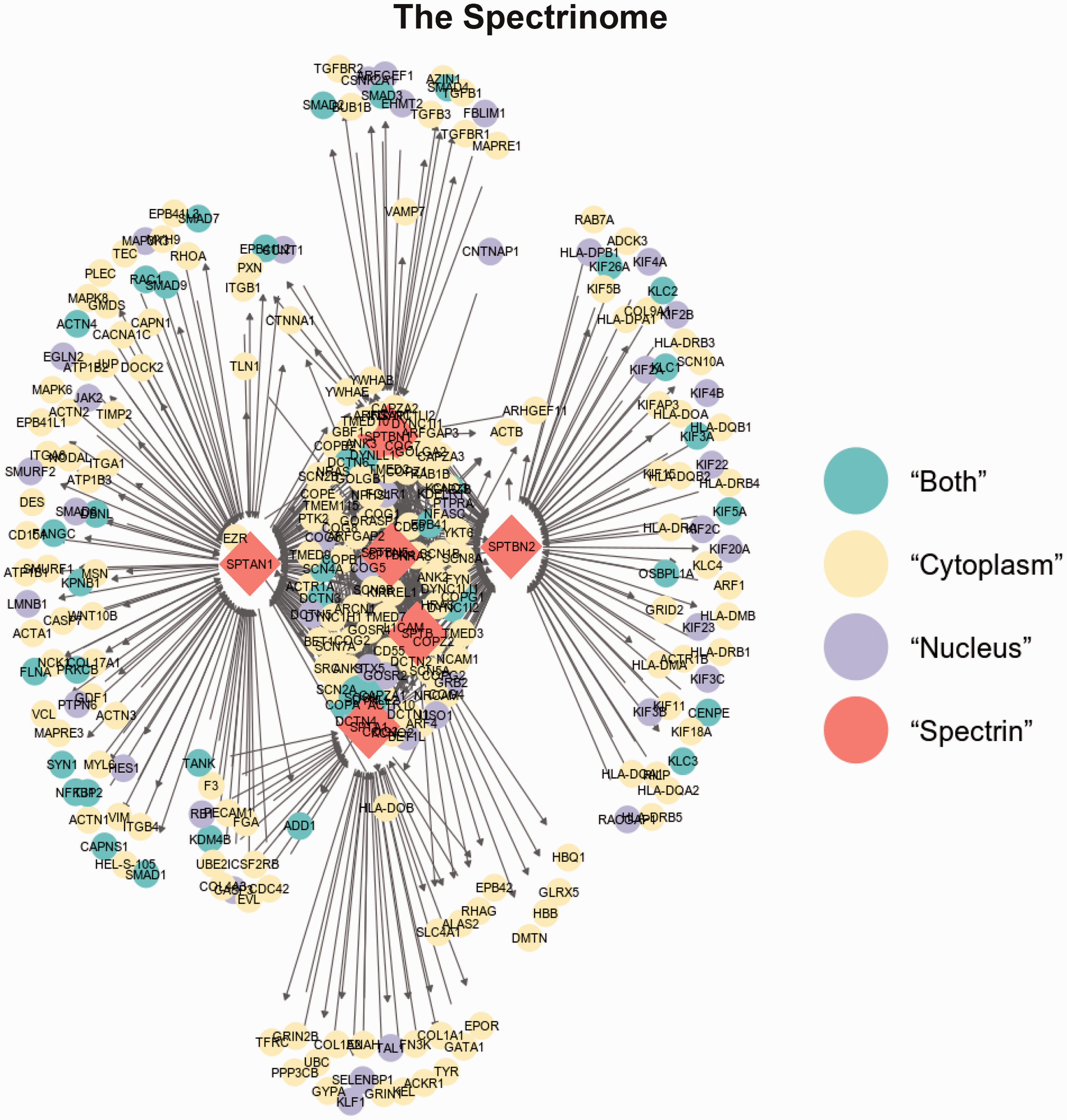
The Entire First Order Spectrinome
The 273 proteins in the 1st order Spectrinome fall into a large number of pathways that are presented for each spectrin gene product in Table 4. These pathways are involved in [1] functions within the cytoplasm (including all membrane surfaces facing the cytosol), [2] essential nuclear functions, [3] signaling from the cell surface to within the nucleus, and [4] extracellular cell – cell interactions. Given the number of pathways in Table 4 a comprehensive discussion of every pathway is beyond the scope of this minireview. In the coming sections we will give several examples for each of the four categories above.
Spectrin Isoform Pathway Analysis
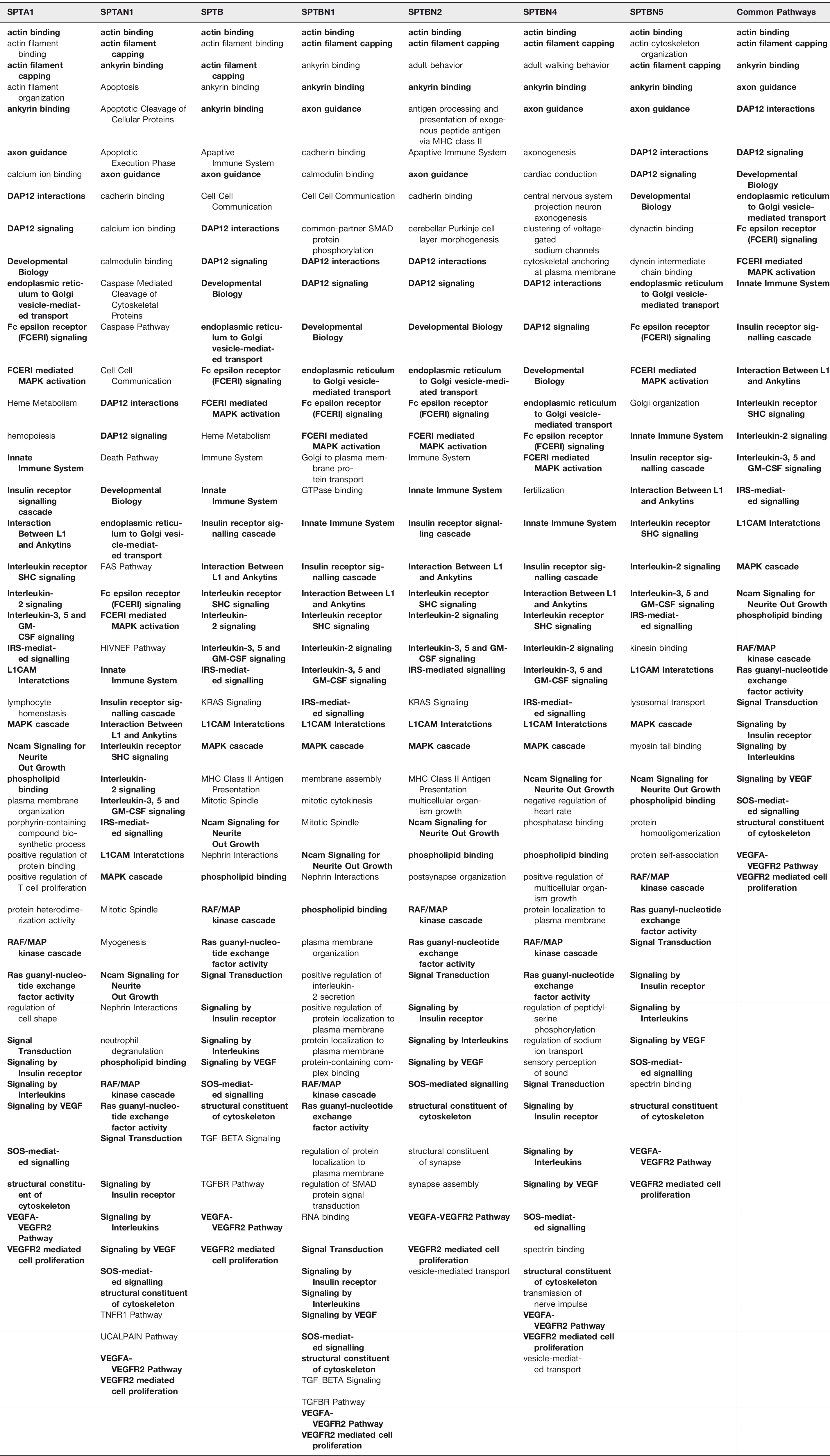
Functions within the Cytoplasm including Cytosol Facing Membrane Surfaces
Just as there are multiple spectrin gene products the same is true for ankyrin, and protein 4.1. Nonerythroid ankyrin was first described by Bennett and Davis, 93 and we now know that there are three distinct ANK genes and proteins: ANK1 or Ankyrin R (human chromosome 8); ANK2 or Ankyrin B (human chromosome 4); and ANK3 or Ankyrin G (human chromosome 10). 94 Goodman and colleagues described a protein 4.1 analogue in brain, that had an apparent molecular weight of 87 kDa, and localized in the identical pattern as the erythroid form of spectrin in mouse cerebellum. 95 It was demonstrated to bind to Spectrin, be distinct from Synapsin I, and was given the name Amelin, 96 but we now refer to this form as 4.1R. Zimmer et al found a form of Amelin (protein 4.1) that localized to axons in mouse cerebellum. 97 Much like the spectrin story we realized that there were both erythroid and nonerythroid 4.1 isoforms, which co-localized with the erythroid and non-erythroid spectrin isoforms in neurons. 98 Today we know there are four 4.1 isoforms: 4.1R (EPB41), 4.1N (EPB41L1), 4.1G (EPB41L2) and 4.1B (EPB41L3). 99 The 4.1 isoforms interact with many important transmembrane proteins. One example is the store-operated Ca2+ channel in endothelial cells.100–102
ANK1, 2 and 3 94 and 4.1 R, G and N 99 can each bind to a wide range of transmembrane proteins, and these associations are essential for many functions including the ion channels, transporters and several enzymes and these interactions and functions have been well chronicled. In this review, we will focus on the direct interactions of spectrin with other proteins and the functional pathways that are influenced by these interactions.
All forms of ankyrin bind to each β spectrin isoform in the β15 repeat between residues 1776 and 1898.103–105 All forms of protein 4.1 bind to each β spectrin isoform in the CH2 domain between residues 171 and 282. 106 Heme binds to βSpII in the β11-12 region between residues 1450 and 1494. 64 The Synapsin I/II binding domain within βSpII has been assigned to a very specific CH2 domain region of residue 211-235. 107 Important to this discussion is that fact the protein 4.1 can compete with Synapsin I and II for binding to intact βSpII. 108 The importance of this spectrin- synapsin interaction to synaptic transmission is described below.
The best-known cytoplasmic function of spectrin comes from our knowledge of the erythrocyte membrane skeleton, which provides membrane stability, shape, flexibility and elasticity.109–111 This raises the question of whether there are similar structures as the spectrin membrane skeleton on the cytoplasmic surface of organelle membranes and, if so, does it play similar functions described above? We know that each of the organelle membrane cytoplasmic components contain isoforms of spectrin, ankyrin and protein 4.1.2,4,9,25,92,94,99 But can they assemble into structures similar to the erythrocyte membrane skeleton and confer similar properties? The answer is yes. In neurons spectrin, actin, adducin, and associated proteins (4.1 and ankyrin) form what has been referred to as membrane associated periodic skeleton (MPS), observed using the super resolution stochastic optical reconstruction microscopy (STORM) methodology. MPS are rings around the cytoplasmic surface of axons, with the rings connected by spectrin tetramers that provide a 190nm periodicity112. The MPS is thought to supply structure, flexibility and mechanical support to axons. MPS also controls the lateral mobility of transmembrane proteins defining which proteins are confined to different axonal segments, and provides stability to axonal microtubules. In more recent studies, MPS have also been found in dendrites. Of substantial interest, Han et al 113 demonstrated a 2D polygonal lattice that reminded the authors of the erythrocyte membrane skeleton in the somato-dendritic compartment of cultured hippocampal neurons using antibodies against βSpII and βSpIII and the STORM methodology. The authors conclude “Taken together, these results suggest a membrane skeleton with a 2D polygonal lattice structure is formed in the somatodendritic compartment of neurons”. This was a clear demonstration of an erythrocyte membrane skeleton-like structure in neurons, and demonstrates that different compartments of neurons can contain this structure (somato-dendrite) or MPS rings (axons and some dendrites). What would be very interesting would be for this study to be performed with βSpI and/or αSpI antibodies, which should recognize the major isoform of spectrin in soma and dendrites of hippocampal neurons (αSpI/βSpIΣ2). As diverse nonerythroid cells and their organelles carry out different functions it is not at all surprising that the spectrin membrane skeleton will take on different structures required for specific functional needs.
Inspection of the spectrinome demonstrates a large number of cytoplasmic proteins involved in membrane traffic. Spectrin isoforms are involved in both anterograde and retrograde transport along microtubules, utilizing kinesin family motors and cytoplasmic dynein motors respectively. The earliest example that spectrin was involved in anterograde transport occurred before fodrin was known to be a non-erythroid spectrin isoform (now called αSpII/βSpII or αSpII /βSpIII). Levine and Willard found a protein with subunits of 250kDa and 240kDa apparent molecular weight in the cortical cytoplasm of neuronal and non-neuronal tissues. 47 Because they felt that this protein looked like a lining of the cell they named it based on the Greek word fodros, meaning lining, fodrin. They had previously found that these two proteins, originally called 26 and 27, travelled the axon from the soma of retinal ganglion neurons to the presynaptic terminal at two speeds: 40 mm/day along with mitochondria and other membranous vesicles, and at slower velocities along with actin, myosin, neurofilament and microtubule proteins. Levine and Willard noticed that the apparent molecular weights of 250 and 240 kDa was similar to those found for filamin and spectrin, so they tried to show a relationship using their fodrin antibody. Unfortunately, they were using a rather insensitive Ouchterlony immunodiffusion technique, where you looked for a precipitin line between the antibody well and the antigen wells. While they clearly observed such a precipitin line with fodrin, they observed no precipitin line with filamin or spectrin. It is historically interesting why they missed that they were studying non-erythroid spectrin, but does not at all diminish the importance of this study. Levine and Willard had demonstrated for the first time that what we now know as non-erythroid spectrin is intimately involved in anterograde rapid and slow transport down the axon. 47 Today we know that the slow transport is actually not slow at all, but instead involves multiple stops and starts along the microtubule pathway.
A quick look at the Spectrinome indicates SPTBN2 (or βSpIII) association with a large number of kinesin superfamily members, referred to as KIFs: KIF 11, 15, 18A, 20A, 22, 23, 26A, 2A, B, C, 3A, B, C, 4A, B, 5A, 5B, AP3. These Kinesin superfamily proteins use the energy of ATP hydrolysis to move cargo along microtubules, most (the N-Kinesins) in the plus-end directed anterograde transport. Different cargos utilize distinct Kinesin Superfamily members (those in bold are associated with
We have known for over 30 years that spectrin, ankyrin, and protein 4.1 isoforms can be found on Golgi cisternae where they form an erythrocyte-like two-dimensional skeleton regulating it’s shape and vesicle transport.5,116 Salcedo-Sicilia et al have demonstrated that βSpIII is enriched in the distal Golgi compartments where it provides structural support and shape and is required for anterograde transport. 117 Further they demonstrated that PI4P is required for βSpIII membrane association.
Spectrin also plays an essential role in retrograde movement of cargo, via the ATP dependent motor called cytoplasmic dynein. Cytoplasmic dynein is involved in the retrograde microtubule based axonal transport from the presynaptic terminal to the soma, vesicle trafficking from the Golgi to the ER, and mitotic spindle assembly (discussed later). A look at the Spectrinome inner ring indicates several key proteins essential for these functions: Alpha-centractin or Arp1 (ACTR1A), the Dynactin subunits (Dynactin1-6), and the cytoplasmic Dyneins (DCTN1-6 plus the heavy, intermediate, and light chains subunits). The Spectrinome outer ring proteins include: Beta-centractin (ACTR1B) which associates with SPTBN2 or βSpIII. Cytoplasmic Dynein contained dimerized heavy chains (DYNC1H1) associated with two light intermediate chains (DYNC1LI1 and DYNC1LI2). Dynactin, a 23-subunit complex, associates with Dynein activating the motor. Dynactin contains a 37 nm actin like filament that is referred to as the ARP1 filament. The ARP1 filament is composed of eight Arp1 subunits (ACTR1A), one β-actin, one Arp 11 (ACTR10), which interacts with p25, 27, and 62 (DCTN 5, 6 and 4) to form the pointed minus end of dynactin. CapZa and CapZb cap the barbed end, while two p24 (DCTN3) subunits and four p50 (DCTN2) subunits and two p150Glued (DCTN1) bind to the pointed end. There are several activator and adaptor proteins that are beyond the scope of this discussion. 118 Again, we would like to give one exemplar of spectrin function related to retrograde transport of vesicle within the cell cytoplasm. Holleran et al demonstrated that βSpIII interacts with Arp1 directly and that brings the Dynactin complex and Dynein motor to associate with the spectrin associated cargo. 119 They demonstrate that in neurons this is involved in retrograde vesicular traffic, while in interphase cells the βSpIII and dynactin colocalize with vesicles in the perinuclear region of the cytoplasm, and the developing cleavage furrow and mitotic spindle of dividing cells. 119
To complete this section, we will describe the association of spectrin (αSpII/βSpII) with synaptic vesicles and the essential role that this plays in synaptic transmission. Goodman and colleagues over a period of approximately 15 years demonstrated by immunoelectron-microscopy that αSpII/βSpII was associated with small spherical 50 nm synaptic vesicles, 5 that this interaction of αSpII/βSpII was via synapsin I, 120 which bound to the tails of the βSpII subunit, 108 and was independent of synapsin phosphorylation. 121 Sikorski et al 122 demonstrated that a peptide specific antibody, directed against the region of βSpII [A207- V445] predicted by Ma et al 64 to be the synapsin I binding domain, could block synaptic transmission in paired hippocampal neurons while peptide specific antibodies against closely flanking regions had no significant effect. 122 Building on these studies, Zimmer et al, by utilizing recombinant peptides demonstrated that the synapsin I binding domain resided within a 25 amino acid stretch of the βSpII CH2 domain (residues 211-235). 107 Further, peptide specific antibody FAB fragments against this 25 amino acid domain rapidly inhibited synaptic transmission in the same paired hippocampal neuron preparation. 107 By this series of studies, recognized as one of the most important contributions to 20th century neuroscience, 123 Goodman and colleagues had established an essential role for non-erythroid spectrin (αSpII/βSpII) in synaptic transmission. 123 Goodman et al have proposed, in the casting the line hypothesis, 49 that the interaction of synapsin I (and possibly synapsin II) with spectrin at the active zone cause the dimpling in of the presynaptic membrane towards the vesicle, thus allowing αSpII/βSpII to serve as a scaffold on which the fusogenic v-snares, t-snares and Ca+2 regulatory proteins can assemble to allow exocytosis of neurotransmitter upon local elevation of Ca+2.
Essential Nuclear Functions
Last year Muriel Lambert, the Special Editor of this Thematic Issue, published a comprehensive description in this journal covering the functions of spectrin, and its partners in the nucleus. 124 We refer the reader to this well-crafted EBM review for detailed coverage of this topic, as we will focus on a few examples of spectrin isoform specific function. Historically it is interesting to ask why the study of nuclear spectrin lagged behind the study of cytoplasmic (including membrane) spectrin by more than a decade. We believe that the answer is two-fold. First, the expectations of what spectrin might be doing in non-erythroid cells was influenced by what we knew about the erythrocyte membrane skeleton. But perhaps more importantly the staining of a wide range of cells and tissues by immunofluorescence and immuno-electron microscopy always showed more intense staining in the cytoplasm as compared to the nucleus when using anti-erythroid or anti-nonerythroid spectrin antibodies.5,37,42,125–130 This is only of historical interest, as we now know that spectrin isoforms play essential roles in the nucleus.
Lambert and colleagues have demonstrated a critical scaffold function for spectrin in DNA repair. 124 To repair interstrand cross-links (ICLs), αSpII binds to DNA at the site of ICL damage. FANCG binds to the SH3 domain of αSpII, followed by ERCC1 binding to FANCG. XPF, an endonuclease, then binds to ERCC1 and incises the DNA damage and repair occurs.124,131–142 Fanconi Anemia is a bone marrow failure disorder, where there is a deficiency in αSpII leading to diminished DNA ICL repair. 124 It has been proposed that Fanconi Anemia (FA) proteins, FANCG and FANCA, can bind to the αSpII SH3 domain protecting αSpII from calpain cleavage. 140 Consistent with this concept, there are patient derived mutations in at least one FA gene, FANCG, which result in the FANCG protein having a defect in the motif that binds to the SH3 domain. 140
Telomeres are regions at the end of chromosomes with repetitive nucleotide sequences that protect genomic stability.144–146 αSpII plays a similar role as described above in serving as a scaffold required for repair of ICL damaged telomeric DNA.124,147 αSpII localizes to telomeres during S phase, where it stimulates the association of TRF1 and TRF2 required for the binding of XPF, demonstrating an important role in DNA repair at the replication fork.124,147
The nuclear envelope contains and outer membrane, inner membrane and a perinuclear space. A nuclear pore complex allows the bidirectional movement of molecules through the nuclear envelope, and a LINC (linker of nucleoskeleton and cytoskeleton) complex allows association between the nuclear skeleton and the cytoskeleton. The LINC complex plays a mechano-sensory function that allows the cell to react to modulation of homeostasis within the extracellular, cytoplasmic or nuclear compartments.148–151 It is composed of several major proteins, including SUN1/2 that forms heterotrimers which are in contact with the nuclear lamina on the nuclear side of the inner membrane, but is also in contact with the Nespirin family KASH (Klarsicht, ANC-1, and Syne homology) domain in the perinuclear space.151,152 The nespirins are transmembrane proteins that cross the outer nuclear membrane. They contain varied numbers of spectrin repeat units, and some contain actin binding CH domains (Nespirin 1 and 2) and all contain KASH domains. Nespirin 1 and 2 can directly bind to cytoplasmic actin filaments, with or without accessory proteins. The smaller Nespirin 1α2, which lacks an actin binding domain, can associate directly with Kinesin 1 thereby associating the nuclear envelope to cytoplasmic microtubules. Nespirin 3 can associate indirectly to cytoplasmic intermediate filaments via proteins such as plectin (also contains spectrin repeat units).151–153 Several transmembrane inner nuclear membrane proteins associate with the lamins and/or chromatin. Lamins A/C and B nuclear lamina intermediate filament interact directly with heterochromatin. Heterochromatin also bind to Emerin via HDAC3. The Lamin B receptor associates with lamin B and heterochromatin via HP1 (heterochromatin protein 1). αSpII associates with Lamin B1 and Plectin (Table 2). The nuclear lamina, an intermediate filament structure, made up of lamins A and B has multiple functions including involvement in gene expression and maintenance of chromatin and nuclear structure.151–159
The nucleoskeleton consist of the nuclear lamina just below the inner nuclear membrane and an interacting spectrin/actin skeleton that also includes myosin and protein 4.1. 123 αSpII has been reported to interact with several nucleoskeleton proteins, based on coimmunoprecipitation from HeLa and human lymphoblastoid cells, including βSpIVΣ5, βSpII, actin, protein 4.1, lamin A, emerin and nuclear myosin1c.160,161 Holaska and Wilson, 161 using an recombinant bead-conjugated emerin to pull down emerin complexes from HeLa nuclear extract, observed seven complexed proteins: nuclear αII-spectrin, non-muscle myosin heavy chain alpha, Lmo7 (a predicted transcription regulator), nuclear myosin I, β-actin, calponin 3 and SIKE (suppressor of IKKε- and TBK1-activation of interferon response elements). These authors went on to further immuno-pulldown and binding assays to determine an emerin proteome. Emerin plays multiple roles in the spectrin-actin cortical network including minus end capping of actin filaments, linking centrosomes to the nuclear inner membrane, and regulating β-catenin activity.157–159 The data taken together from Sridharan et al 160 and Holanska and Wilson 161 suggest that these nuclear spectrin complexes include a spectrin/actin skeleton associated with the nuclear lamina, the inner nuclear membrane via emerin, myosin and transcription factors.
Historically, the structure and function of the nuclear lamina was understood well before the discovery of nuclear spectrin.124,162 As discussed above the nucleoskeleton is an integrated combination of the nuclear lamina and the spectrin-actin network. The nucleoskeleton provides important mechanosensory properties to the nucleus. The spectin-actin lattice provides the properties of elasticity and response to compression, reminiscent of the functions of the erythrocyte membrane skeleton.163,164 The nuclear lamina provides mechanical stiffness to the nuclear membrane.165–168 In summary nuclear spectrin is involved in DNA repair, telomere integrity, nuclear architecture, properties of mechanical stability and elasticity of the nuclear envelope, and signaling from the extracellular space into the nucleus. The later will be the subject of the next section.
Signaling From the Cell Surface to Within the Nucleus
The Spectrinome shows the interaction of αSpII with several SMADs (SMAD 1, 6, 7, 9), SMURF 1 and 2; while βSpII interacts with SMAD 2, 3, 4, TGFB1 and B3, TGFBR1 and 2. This clearly demonstrates a central role of non-erythroid spectrin isoforms in the TGF-β signaling pathway. The TGF-β signaling pathway is involved in many functions of normal cells including, but not limited to, pattern formation during embryonic development, ECM production, angiogenesis, tissue repair, immune regulation, cell differentiation, cell growth and proliferation, and apoptosis. 169 For such an impactful set of cellular processes the pathway, as it relates to spectrin is rather simple.111,170,171 The TGF-β receptor 2 (TGFBR2) is a Ser/Thr receptor kinase. When TGF-β binds to TGFBR2, it results in association and phosphorylation of the TGF-β receptor 1 (TGFBR1). TGFBR1 then recruits and phosphorylates SMAD2 or SMAD3. The phosphorylated SMAD3 dissociates from the receptor and associates with SMAD4 (a known tumor suppressor) and βSpII. The SMAD3/4- βSpII complex enters into the nucleus together and associate with specific transcription factors and a TGF-β-response element in target genes. This in turn leads to increased specific target gene transcription. As an example, if the function is cell cycle regulation then examples of targets are Cyclin dependent kinase inhibitors, which inhibit cyclin D/CDK4/6 activity blocking the cell cycle at the G1 phase; and p21 which mediates the anti-proliferative response and provides a partial explanation for tumor-suppression by the TGF-β signaling pathway. The SMURFS are E3 Ubiquitin ligases which polyubiquitinate the SMADS, leading to their turnover by 26S proteasomes.111,169–171
As mentioned at the start of this section, αSpII also is associated with several SMADs and SMURF 1 and 2. Metral et al demonstrated that decreasing αSPII in a cell line derived from human melanoma cells resulted in cell cycle arrest at the G1 checkpoint. 172 They further demonstrated that αSpII deficiency led to increased expression of the cyclin-dependent kinase inhibitor: p21Cip. 172 This suggests that both αSpII and βSpII are involved in cell cycle control via TGF-β/SMAD signaling.
Extracellular Cell – Cell Interactions
Pollerberg et al 173 demonstrated by fluorescence recovery after photobleaching (FRAP) technology, that NCAM140 moved more rapidly than NCAM180 in the plasma membrane of a neuroblastoma cell line. In this study, NCAM180 was suggested to be restricted in its mobility by interaction with the cytoskeleton, and brain spectrin appeared to copurify with NCAM180 but not NCAM140. In a related article from Pollerberg et al 174 , NCAM180, but not NCAM140 or NCAM120, was demonstrated to be concentrated in areas of cell-cell contact in both the neuroblastoma cell line and primary cultures of cerebellar neurons. Further, this study directly tested the association of the NCAMs and brain spectrin. Brain spectrin bound only to NCAM180, via its C-terminal cytoplasmic domain, and it did not bind to NCAM140 or NCAM 120 that lack this domain. 174 This demonstrated a direct association of spectrin with NCAM180.
The NCAMs are members of the Immunoglobulin superfamily adhesion molecules, which also includes the L1 family. 175 The Immunoglobulin superfamily are cell surface transmembrane proteins that have one to five Ig domains and three fibronectin III (FNIII) homologous domains in their extracellular domain. Their extracellular domains are involved in hemophilic and heterophilic interactions with adhesion molecules at sites of cell contact and ion channels respectively. They serve many functions in neurodevelopment, and synaptic functions essential for learning and memory; and interact with various ion channels, neurotransmitter and cytokine receptors. 175 The Spectrinome shows several Immunoglobulin superfamily members in the inner ring, associating with all spectrin isoforms.
For a few examples of the published interactions: NCAM180 binds to αSpI/βSp1Σ2;174,175 L1 binds via ANK B to αSpII/βSpII; CHL1 associates directly with βSpII or indirectly via ankyrin; Neurofascin binds to αSpII/βSpII via ANK G; and SynCAM 1 associates with αSpII/βSpII via 4.1B.111,170,175 The association of NCAM180 to αSpI/βSp1S2 recruits spectrin associated-NMDA receptors and CAM Kinase IIa to the spectrin scaffold at post synaptic densities. 176 The NMDA Receptor is an ionotropic glutamate receptor, that is essential for controlling synaptic plasticity and is linked to memory and learning.175,176 We showed in our immuno-electron microscopy (see Figure 2 for summary) that αSpI/βSpIS2 is highly enriched in post-synaptic densities, 5 and Sytnyk et al demonstrated that NCAM180 drives both spectrin and CAM Kinase IIα to this post-synaptic complex. 176 In the NCAM-/- mice that they utilized the NMDA receptor–dependent form of long-term potentiation (LTP) was impaired. 176 Therefore defects in this spectrin scaffold could be critical for psychiatric disorders such as bipolar disease and schizophrenia. 175
Why is Spectrin’s Chimeric E2-E3 Activity Important to Spectrin Isoform Functions?
There have been several reviews that describe our discovery that erythroid spectrin is not only an essential scaffold protein serving all the functions described in previous sections of this review, but also has an essential function as an E2/E3 chimeric ubiquitin conjugating and ligating enzyme.8,9,92 A relatively recent EBM review by Goodman et al 9 , covered the history of this discovery of erythroid spectrin’s E2/E3 activity, but also the very likely probability that αSpII has the very same function with a greater number of target proteins. Therefore, we will cover the history in just enough detail to be able to describe just how important we believe this enzymatic function will be for normal cellular homeostasis and disease states that are accompanied by oxidative stress.
Kakhniashvili et al first demonstrated that human alpha spectrin (αSpΣ1) had an E2 ubiquitin conjugating activity that had the ability to target itself. 6 We proposed, based on sequence and structural analysis, that cysteine 2071 would be the essential E2 site as it was found within a 17 amino acid sequence that had 70% identity to all known consensus E2 active sites. 6 We also proposed, based on structural analysis, that cysteine 2100 could play a role in ubiquitin transfer. Both cysteine 2071 and cysteine 2100 are found within αSpI repeat 20 and we predicted that a lysine rich region within repeat 21 could be the target site. 6 At this time, we did not yet know whether a separate E3 ligating enzyme was required or whether spectrin was capable of ubiquitinating other associated proteins. Over the following four years both questions were answered.
Hsu et al 7 , using a recombinant αSpI GST-protein (2005-2415) which contained the αSp repeats 20 and 21 and reached to the C-terminus of αSpI, demonstrated that this domain of spectrin had the ability to ubiquitinate itself in the presence of purified E1 enzyme and ATP, but in the absence of any E3 ligase. Hsu et al had demonstrated that this predicted fragment did contain a chimeric E2/E3 activity capable of ubiquitinating itself. Further mutating the six cysteines in this protein, either individually or collectively, demonstrated that Cysteine 2071 is the cysteine that can serve as the E2/E3 site.7,8 Cysteine 2100 serves a backup E2/E3 function in humans only, as this cysteine is converted to a tyrosine or glutamine in other mammalian species.7–9
Chang et al70,71,177 demonstrated that spectrin was capable of ubiquitinating not only alpha spectrin, but also ankyrin, band 3, protein 4.1 and a protein of unknown function (gi 13278939). The ability of a site on αSpI repeat 20 to ubiquitinate protein 4.1 could be explained by the proximity to the 4.1 binding domain. However, ubiquitination of ankyrin and band 3 required either: [a] that one spectrin molecule can ubiquitinate sites on an adjacent spectrin molecule, or [b] that the condensation of spectrin down to 55 nm brings the enzymatic site in close proximity to the target protein, or [c] that the ubiquitination of these other proteins requires a separate E3 ubiquitin ligase. Possibility [c] was ruled out as no exogenous E3 was added in these ubiquitination assays.70,71,177 During the preparation of this review, we searched the existing literature to gain possible insight into the possible role of the unknown protein gi 13278939. What we found holds special interest for this review. Two studies using human embryonic kidney HEK293 cells and either Proliferating Cell Nuclear Antigen (PCNA) affinity resin 178 or PCNA proximity labelling and pull down 179 , followed by proteomics and interactomics, have demonstrated an association between gi 13278939 and PCNA. In the case of Ohta et al gi 13278939 was pulled out of a nuclear extract with PCNA resin. 178 PCNA is essential for chromatin establishment and remodeling, DNA replication, DNA repair and sister chromatid cohesion. It functions as a homo-trimer that encircles DNA, tethering DNA polymerases to DNA. Therefore, to summarize, a target of SpI E2/E3 ubiquitin conjugating/ligating activity is a nuclear binding partner to PCNA which is essential to numerous nuclear functions.
Goodman’s laboratory went on to demonstrate that membrane protein ubiquitination, including αSPI, is diminished in erythrocytes from Sickle Cell Disease patients. 177,180 Furthermore, we demonstrated that this was due to decreased spectrin E2/E3 activity in SCD RBCs. 177 SCD RBCs are exposed to increased oxidative stress, with decreased protection by reduced glutathione, which results in cysteine oxidation and glutathiolation leading to blockage of αSpI C2071 (and other cysteines), and oxidative damage to other proteins.6–9,177,181–187 Goodman and colleagues have demonstrated that diminished mono-ubiquitination of αSpI has no impact upon spectrin dimer to tetramer interconversion 188 but results in the diminished ability of spectrin-4.1-actin 189 and spectrin-adducin-actin 190 complexes to dissociate at physiologic conditions. These effects, plus oxidation of actin leading to filaments which will not depolymerize191–194, leads to the slow dissociation of the membrane skeleton191,195 and is the molecular basis of the irreversibly sickle cell (ISC). 183 Caprio et al have found that diminished spectrin ubiquitination is also the basis for RBC rigidity in multiple organ dysfunction syndrome (MODS). 196
As explained in our previous reviews, and shown in Figure 7, there is every reason to believe that αSpII has the same chimeric E2/E3 activity as αSpI.8,9 As this has been previously reviewed, we will focus on the reasons demonstrated by Figure 7. Human αSpII has a 16 amino acid segment in repeat 20 that contains a cysteine 2120 which is surrounded by the consensus E2 sequence found in all E2 enzymes. This stretch in αSpII has 63% identity to the same stretch in human αSpI (Figure 7A), and a 100% identity to the same stretch in αSpII from mouse, chicken and worm.
9
There is a single conserved amino acid change in Drosophila that is distinct from the human version (Figure 7B). It will be a simple study to test the αSpII E2/E3 activity

Spectrin E2/E3 Consensus Active Sites
So why is the spectrin E2/E3 activity important for normal physiology and pathophysiology of all human cells? We already know that SPTA1 is present not only in RBCs, but also in the cytoplasm and nucleus of neurons and skeletal and cardiac muscle (see Spectrinome figures 4 and 6). While the human erythrocyte proteome contains approximately 2300 proteins, 110 the human proteome will contain about ten times that number of proteins. 87 The number of Spectrinome proteins with 1st order interactions with SPTA1 are 137 (Supplementary Table 1), and these databases contain only about 75% of the human proteome. All of these proteins can potentially be ubiquitinated by αSpI. For SPTN1 the number of proteins in the 1st order Spectrinome is 183, again with a database which is about 75% complete. SPTAN2 is in all human cells except RBCs. We have demonstrated ubiquitination of αSpI and αSpII in rat hippocampal neurons in culture and in human brain slices from deceased Alzheimer’s, Parkinson’s, and Age matched control human brains. In Alzheimer’s and Parkinson’s disease both α-spectrins were major components of the ubiquitinated inclusions.91,92
Homeostasis of every protein pathway discussed in this review can potentially be regulated by Spectrin’s chimeric E2/E3 activity. While we have demonstrated that RBC spectrin can be mono-ubiquitinated, and this regulates protein-protein interactions; 6,189,190 examination of our earlier articles (ex, ref 6 Figure 1) shows a ladder of spectrin-ubiquitin adducts when RBC membranes are blotted with Ubiquitin antibodies. This suggests that while spectrin can mono-ubiquitinate itself, that this single ubiquitin can become a poly-ubiquitin chain. We would suggest that it is likely that spectrin’ s E2/E3 activity can lead to poly-ubiquitination, thereby involving proteasomes in the regulation of those pathways impacted by the various spectrin scaffolds. This is an exciting area for future investigation.
Sangerman et al studied the synthesis and turnover of spectrin isoform specific subunits in primary cultures of hippocampal neurons. 91 What was found was quite interesting. The αSpIΣ1 and βSpIΣ2 erythroid subunits all turned over with a half-life of 16 to 24 minutes, independent of whether the spectrin subunits were in the soluble pool or the particulate pool. However, for the nonerythroid subunits αSpIIΣ1 and βSpIIΣ1 the turnover of the particulate (membrane or nucleo/cyto-membrane skeletal) forms was 30 to 34 minutes, but the turnover of the soluble forms turned over much slower: 80 (αSpII) and 53 minute (βSpII) half-life. Do these rates of turnover reflect the distribution of proteasomes in the cytoplasm versus nucleus, or membrane versus cyto- or nucleo-plasm? Spectrin could be keeping both the cytoplasmic and nuclear proteasomes very busy. This in uncharted territory for future investigation.
Calpain cleavage of αSpIIΣ1 between Tyr1176 and Gly1177 at the hypersensitive site, in response to Glutamate binding to the NMDR receptor and elevated Ca2+, is a normal part of regulating neuronal development and synaptic plasticity.80,81,83,197–199 But during brain injury caused by TBI, hypoxia-ischemia or neurodegeneration, Ca2+ increases to levels where in addition to the calpain cleavage of αSpII, we also see caspase 3 cleavage of both αSpII and βSpII leading to cell death.83,198–200 How are the calpain spectrin signature breakdown products (SPDPs) replaced by new αSpII? Does this require ubiquitin directed proteasomal degradation of these membrane bound SBDPs? Is this also involved in control of synaptic plasticity? There are lots of studies to be done. Neurodegeneration and many other diseases discussed in this review, including the following section, are characterized by altered redox status. Does this cause altered spectrin E2/E3 activity in these non-erythroid cells and tissues, where the critical cysteines would again be oxidized or glutathiolated?
There is much to be done, but when you have a scaffold protein, representing 2-3% of all human cells, present in all cellular compartments, playing a myriad of functions, and this protein has the ability to ubiquitinate its partners, how could this not be central to normal and/or pathologic cell function?
Spectrin and Human Disease
In this section, we will focus on Human Disease caused by a spectrin specific genetic defect or diminished levels of the protein. We have given several examples in Table 5. Although a very interesting subject, we will not focus on genetic disorders related to spectrin binding partners. As an example, we refer the reader to several reviews that discuss the relationship of defects in ankyrin isoforms to human cardiomyopathies and neurologic disorders.201–205
Spectrin Isoforms and Human Diseases
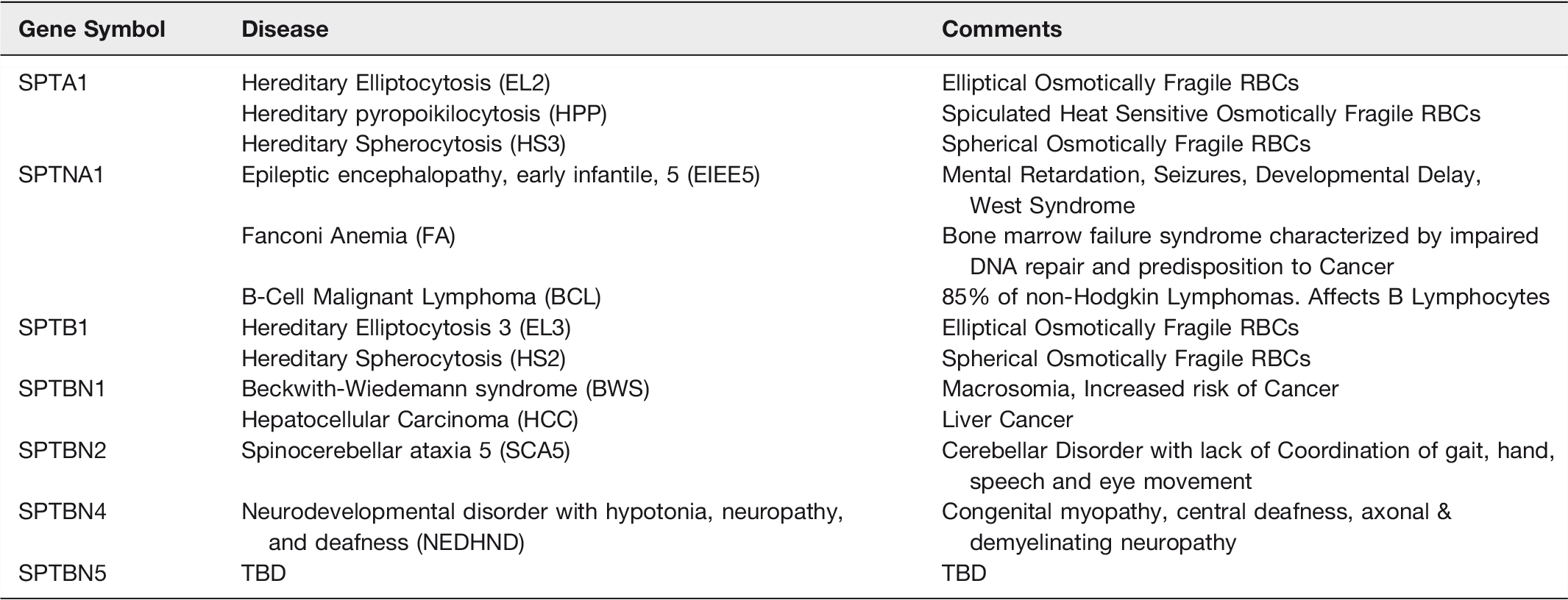
Defects in SPTA1 and SPTB1 lead to the well-known RBC disorders where red cells take on abnormal shapes, possess friable membrane skeletons and are osmotically fragile. Hereditary Elliptocytosis (HE) are a heterogeneous group of genetic disorders where the RBC takes on an extended oval or elliptical shape. The most common causative genetic defects are found near the n-terminus of αSpI (EL2) or the c-terminus of βSpI (EL3) and disrupt the head-to-head interaction of spectrin heterodimers leading to an inability to form tetramers.206,207 In some cases, the genetic defect can be more distal from the n-terminus of αSpI, and be caused by stabilization of a closed dimer conformation leading to diminished spectrin tetramers. 208 Most of HE patients are heterozygotes and have a mild form of the disease. Some patients are homozygous or compound heterozygotes and have a more severe form of this disorder that is referred to as Hereditary Pyropoikilocytosis (HPP) where the RBCs are oval and spiculated and heat sensitive.206,207 HPP patients have a higher content of abnormal αSpI as compared to HE.
Hereditary Spherocytosis (HS) is also heterogeneous hemolytic anemia where the RBCs are round and osmotically fragile. Most cases demonstrate a partial deficiency in spectrin leading to blebbing off vesicles and loss of membrane surface area. The diminishment of spectrin can be due to direct genetic defects in α-spectrin (SPTA1), β-spectrin (SPTB1) or, alternatively in the proteins that are involved in linking spectrin to the bilayer (ankyrin (ANK1), band 3 (SLC4A1), and protein 4.2 (EPB42)).206,207,209 There have also been cases of HS described that are caused by defects in the 4.1/actin binding domain of βSpI leading to diminished formation of the spectrin-4.1-actin ternary complex.210,211
As discussed in the prior section dealing with the functions of spectrin in the nucleus, αSpII (SPTAN1) plays an essential scaffolding role in DNA interstrand crosslink repair and maintaining chromosomal stability. Diminished levels of αSpII (35-40% of normal) are found in cells from patients with the bone marrow failure disorder Fanconi’s Anemia.124,133,136 The hallmarks of Fanconi’s anemia are bone marrow failure, inability to repair DNA damage and a higher likelihood of developing cancer.124,212–216 Given its role in DNA repair it is not surprising that diminished or mutationally altered αSpII will be found in various forms of human cancer. In fact, it has been suggested that αSpII plays a tumor suppressor role for a broad range of cancers. 217 One example is the observation of diminished αSpII in bone marrow samples from patients with acute myeloid leukemia. 218 Mutations in SPTAN1 have also been demonstrated to lead to early onset West syndrome, or Severe Early Infantile Epileptic Encephalopathy (EIEE5). EIEE5 is characterized by early onset seizures, brain atrophy, mental retardation, difficulties with walking, speech and vision.219–221 The mutations in several cases were in the nucleation site that allows normal spectrin heterodimer formation.
SPTBN1, or βSpII, genetic defects lead to several human disorders including Hepatocellular Cancer. 222 As discussed earlier, βSpII serves an essential role in the TGF-β signaling pathway. Dysregulation of TGFβ signaling is thought to contribute to dysfunctional differentiation leading to the development of cancers. As mentioned above, βSpII associates with the Smad3/Smad4 complex and plays a role in its entry into the nucleus. This drives TGFβ-mediated tumor suppression. The defective or diminished βSPII leads to disruption of TGFβ signaling and has been demonstrated to play a key role in the development of gastrointestinal cancers. Zhi et al demonstrated that knockdown of βSpII expression led to stem cell-like features in hepatocellular carcinoma cells and to malignant tumor progression. 223 Chen et al recently showed that βSPII expression correlates with the differentiation of hepatocytes in culture. They further demonstrated that hepatocyte differentiation induced by βSPII decreased the production of liver cancer stem cells. 224 . This suggests that enhancing the levels of βSpII in liver cancer stem cells could be an efficacious treatment strategy for treating hepatocellular carcinoma.
Ikeda et al have characterized the mutations in German, French and American families with spinocerebellar ataxia type 5 (SCA5). 225 Interestingly, the American family was descended from the paternal grandfather of President Abraham Lincoln. What was found were mutations in the actin-binding domain of βSpIII in the German family and in repeat 3 in the American and French families. This group found a loss of the glutamate receptor (EAAT4) from the plasma membrane of Purkinje cells due to the defect in βSpIII. 225 This would lead to altered glutamate signaling and may explain the characteristics of SCA5. These characteristics include: mildly progressive atrophy of the cerebellum, ataxia impacting gait, and slurred speech.
Several cases of mutations in βSpIV (SPTBN4 gene) result in a human neurodevelopmental disorder with hypotonia, neuropathy, and deafness (NEDHND). 226 βIV-spectrin protein is known to be critical for securing Na+ channels to their position in the initial segment of an axon, as well as nodes of Ranvier.227,228 In this regard, Kopp-Scheinpflug and Tempel have shown mutations in mice that affect the βIV-spectrin protein result in temporal alterations in central auditory neuronal signaling that could be a mechanism of auditory processing disorder. 229 More recently, Liu et al., 230 have outlined the critical role of spectrin in hearing development and deafness. Not surprisingly, they found that specific spectrin subunits, namely, αII- and βII-spectrin, were critical for the structural organization of rodent hair cells. Further, βII-spectrin knockout mice demonstrated extreme deafness.
The observation that βIV-spectrin is important for the control of initial segment and nodal Na+ channel clustering provides insights into other sensory organ dysfunction that may result from key mutations. Although no specific forms of olfactory dysfunction have been associated with such mutations, the observations of Kosaka et al 231 suggest that such a relationship should exist. The main olfactory bulb is by-in-large anaxonic. Nonetheless, these investigators identified sodium channel clusters associated with βIV-spectrin ‘‘hot spots’’ on dendritic segments of external plexiform layer neurons in the rodent main olfactory bulb. Thus, it is intriguing to speculate that mutations that lead to channelopathies or altered cytoskeletal/scaffolding in this sensory organ would also lead to altered processing and dysfunction.
We have purposely focused these discussions on human diseases that have been linked to spectrin isoforms. Many of these assignments were based on prior work in animal models. Although we have not assigned a human disease to SPTBN5 in table 5, that could change soon. Usher Syndrome (USH) is the major cause of human deaf-blindness. One of the defective gene products causing a severe form of this disease (USH1B) is the actin-based motor protein: myosin VIIa. Mutant mice that are deficient in this motor protein provide a phenotype similar to the hearing defect observed in USH1 subjects. It has now been demonstrated that βSpV (SPTBN5) homodimers couple USH1 proteins to myosin VII actin-based motors; and could play a key role in USH1B. 232
Postlude
Erythrocyte Spectrin was first described over 50 years ago by Marchesi and Steers, 10 we are approaching 40 years since Goodman et al described “Spectrin-like molecules in non-erythroid cells” 1 , and it has been over 30 years ago since Reiderer et al provided the first description of multiple spectrin isoforms. 37 The great abundance of publications in the field over the past 50 years made it challenging to describe the functions of spectrin isoforms in the cytoplasm and nucleus, and signaling between the two compartments, in a minireview. We found the spectrinome to be a useful tool for visualizing and considering the enormous number of roles played by these isoforms and hope that it is useful to workers in the field, as they chart their future studies. We have made enormous progress in understanding the role of these spectrin isoforms in cellular physiology and pathophysiology, and there is plenty that remains to be done by current and future spectrin researchers. With the enormity of five decades worth of information, portrayed by the spectrinome, we had to pick and choose examples to discuss. There are many elegant studies that we could not cover in this review, due to page limitations, and we hope you will understand that we have read and appreciate all of the outstanding work that has preceded this review.
Supplemental Material
Supplemental Table1 - Supplemental material for The Spectrinome: The Interactome of a Scaffold Protein Creating Nuclear and Cytoplasmic Connectivity and Function
Supplemental material, Supplemental Table1 for The Spectrinome: The Interactome of a Scaffold Protein Creating Nuclear and Cytoplasmic Connectivity and Function by Steven R. Goodman, Daniel Johnson, Steven L. Youngentob and David Kakhniashvili in Experimental Biology and Medicine
Supplemental Material
Supplemental Table2 - Supplemental material for The Spectrinome: The Interactome of a Scaffold Protein Creating Nuclear and Cytoplasmic Connectivity and Function
Supplemental material, Supplemental Table2 for The Spectrinome: The Interactome of a Scaffold Protein Creating Nuclear and Cytoplasmic Connectivity and Function by Steven R. Goodman, Daniel Johnson, Steven L. Youngentob and David Kakhniashvili in Experimental Biology and Medicine
Supplemental Material
Supplemental Table3 - Supplemental material for The Spectrinome: The Interactome of a Scaffold Protein Creating Nuclear and Cytoplasmic Connectivity and Function
Supplemental material, Supplemental Table3 for The Spectrinome: The Interactome of a Scaffold Protein Creating Nuclear and Cytoplasmic Connectivity and Function by Steven R. Goodman, Daniel Johnson, Steven L. Youngentob and David Kakhniashvili in Experimental Biology and Medicine
Supplemental Material
Supplemental Table4 - Supplemental material for The Spectrinome: The Interactome of a Scaffold Protein Creating Nuclear and Cytoplasmic Connectivity and Function
Supplemental material, Supplemental Table4 for The Spectrinome: The Interactome of a Scaffold Protein Creating Nuclear and Cytoplasmic Connectivity and Function by Steven R. Goodman, Daniel Johnson, Steven L. Youngentob and David Kakhniashvili in Experimental Biology and Medicine
Supplemental Material
Supplemental Table5 - Supplemental material for The Spectrinome: The Interactome of a Scaffold Protein Creating Nuclear and Cytoplasmic Connectivity and Function
Supplemental material, Supplemental Table5 for The Spectrinome: The Interactome of a Scaffold Protein Creating Nuclear and Cytoplasmic Connectivity and Function by Steven R. Goodman, Daniel Johnson, Steven L. Youngentob and David Kakhniashvili in Experimental Biology and Medicine
Supplemental Material
Supplemental Table6 - Supplemental material for The Spectrinome: The Interactome of a Scaffold Protein Creating Nuclear and Cytoplasmic Connectivity and Function
Supplemental material, Supplemental Table6 for The Spectrinome: The Interactome of a Scaffold Protein Creating Nuclear and Cytoplasmic Connectivity and Function by Steven R. Goodman, Daniel Johnson, Steven L. Youngentob and David Kakhniashvili in Experimental Biology and Medicine
Supplemental Material
Supplemental Table7 - Supplemental material for The Spectrinome: The Interactome of a Scaffold Protein Creating Nuclear and Cytoplasmic Connectivity and Function
Supplemental material, Supplemental Table7 for The Spectrinome: The Interactome of a Scaffold Protein Creating Nuclear and Cytoplasmic Connectivity and Function by Steven R. Goodman, Daniel Johnson, Steven L. Youngentob and David Kakhniashvili in Experimental Biology and Medicine
Footnotes
Author Contributions
S.R. Goodman was responsible for the Spectrinome concept and contributed to the writing of all sections of this mini-review. D. Johnson did all of the bioinformatics work on the Spectrinome and wrote part of Constructing the Spectrinome section. S.L. Youngentob wrote part of the Spectrin and Human Disease section. D. Kakhniashvili did much of the RBC proteomic studies and spectrin E2/E3 studies that are described, and contributed to the construction and writing of the Why is Spectrin’s Chimeric E2/E3 Activity Important to Spectrin Isoforms Function? section.
DECLARATION OF CONFLICTING INTERESTS
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
National Institutes of Health (AA017823).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.