
Abstract
Microtubules are cytoskeletal elements known as drivers of directed cell migration, vesicle and organelle trafficking, and mitosis. In this review, we discuss new research in the lens that has shed light into further roles for stable microtubules in the process of development and morphogenesis. In the lens, as well as other systems, distinct roles for characteristically dynamic microtubules and stabilized populations are coming to light. Understanding the mechanisms of microtubule stabilization and the associated microtubule post-translational modifications is an evolving field of study. Appropriate cellular homeostasis relies on not only one cytoskeletal element, but also rather an interaction between cytoskeletal proteins as well as other cellular regulators. Microtubules are key integrators with actin and intermediate filaments, as well as cell–cell junctional proteins and other cellular regulators including myosin and RhoGTPases to maintain this balance.
Impact statement
The role of microtubules in cellular functioning is constantly expanding. In this review, we examine new and exciting fields of discovery for microtubule’s involvement in morphogenesis, highlight our evolving understanding of differential roles for stabilized versus dynamic subpopulations, and further understanding of microtubules as a cellular integrator.
Introduction
The cytoskeleton is a structure that helps maintain cell shape,1–6 organizes and transports organelles,1,7–12 provides mechanical support,1,5,13–16 and allows for cell division and movement.2,17–20 Amongst these cytoskeletal elements, microtubules specifically are known regulators of directional cell migration,21–26 vesicle and organelle trafficking,8,27,28 and mitosis.29–32 These polarized hollow cylinders are composed of α-tubulin and β-tubulin heterodimers and nucleated at the microtubule organizing center at their minus ends, with the plus ends growing outward.33–35 Movement along microtubules to allow for trafficking and placement of cellular contents is directed by molecular motor proteins kinesins and dyneins, with kinesins moving toward the microtubule plus end and dyneins moving toward the microtubule minus end.27,36–40
Microtubules are characteristically defined by a constant cycling between growing and shrinking referred to as dynamic instability.41–44 Subpopulations of microtubules can be stabilized by post-translational modifications including acetylation45–53 or detyrosination48,49,54 as well as their interactions with microtubule-associated proteins (MAPs).55–58 This stabilization of microtubules is important for several cell processes, with acetylated microtubules found at the leading edge of actively migrating cells assisting in directional migration. 54 Acetylated microtubules are also the foundation of primary cilia, which are generally referred to as the antennae of the cell, involved in sensing the cellular environment, cell signaling, liquid flow, cell polarity and multiple sensory organ functions including smell, sound, and sight.45,47,59–63
Microtubules’ role in cell functioning greatly depends on its ability to coordinate the functioning of other cellular elements, including other cytoskeletal components.64–68 This has proven especially true in the field of cellular migration, with microtubules shown to have both direct and indirect effects on the actomyosin machinery in motile cells, partly through regulation of their stability.64–68 Newer research has also demonstrated that microtubule interaction with cellular elements includes not only cytoskeletal elements and cytoplasmic organelles, but also cell–cell junctional proteins.68–72 Here, we look at the expanding role of microtubules in cellular functions and how microtubules may serve as a keystone of cytoskeletal coordination.
Microtubules in the lens: A developmental role
The lens is a tissue whose structure defines its function. A transparent tissue devoid of vasculature and nerves, it serves to focus light rays onto the retina to allow for vision. While generally looked upon as a simple clear gelatinous structure, the lens is actually defined by a precise cellular arrangement within the lens epithelium and among the differentiated fiber cells as well as between these two primary cell types of the lens.73–76 It is this interaction between and among these two different cell types that allows for the generation and maintenance of a three-dimensional structure that can adapt and accommodate to allow for proper vision.77,78 This precise cytoarchitecture’s establishment and maintenance relies heavily on cytoskeletal signaling networks in conjunction with cell–cell junctions.79–83 This reliance on cell–cell interaction makes the lens an ideal system for studying the processes of development, morphogenesis, and regeneration.
Additionally, the lens is a structure that continues to develop and grow throughout an organism’s life.84–86 Given that the lens lacks blood and nervous supply, 87 it is the structural architecture and cell–cell communications that allow for this tissue to retain integrity across the course of an animal’s life. The disruption of this system leads to opacities such as cataracts, often accompanied by changes in protein density, cellular depolarization, osmotic disruption, and oxidative stress damage leading to cellular degeneration, membrane disruption, and loss of tissue architecture.88–92 This makes the lens a system also well-suited to examining pathogenesis, at how cellular and structural changes affect tissue development and maintenance and how this tissue and those around it respond to injury.
Despite early research into the presence and potential role of microtubules in lens culture systems,93–96 very little was understood about the role of microtubules in the in vivo lens. In this early work, microtubules were demonstrated to orient themselves along the axis along which they would elongate and were found to be present in both the epithelium and fiber cells, 97 and were abundant in elongated fiber cells. 97 Early work from the Beebe lab looked at lens epithelial cell elongation and found that only certain microtubule inhibitors, namely colchicine, prevented epithelial cells in culture from elongating. 98 However, these in vivo studies looked at epithelial cells in isolation under the influence of fetal calf serum to promote differentiation rather than looking at how the microtubules in actively elongating fiber cells may play a role in overall lens development. Moreover, microtubule inhibitors caused significant inhibition of lens cell elongation and cataract56,99 and cataractous lenses were often found to demonstrate loss of microtubules 97 demonstrating that microtubules must play some role in lens development and maintenance.
More recently, a form of autosomal-recessive congenital cataract was linked to a mutation in a mediator of microtubule plus-end-directed vesicle transport, FYCO1, suggesting that there is a role for microtubule trafficking in proper lens development. 56 FYCO1 is also associated with autophagosomes, and thus microtubules’ role may also involve trafficking of autophagosomes for organelle-free zone formation, 100 similar to its role in other systems.63,101–103 Additional studies have also suggested that microtubules may play a role in lens fiber cell differentiation related to their known function in transporting proteins, membrane, and organelles. 9
New findings using the embryonic lens as a model demonstrate that microtubule isoforms are expressed abundantly in both epithelial and fiber cells68,104 unlike other cytoskeletal elements, in particular lens-specific intermediate filaments, that are greatly enriched in either epithelial or fiber cell compartments.104–108 Instead, it is the post-translational modifications that are highly localized in the lens, with stable, acetylated microtubules concentrated in newly differentiated fiber cells and the apical aspects of lens equatorial epithelial cells. 68 Disruption of dynamic microtubules does not impede normal fiber cell elongation and morphogenesis, but depolymerization of the stable acetylated microtubules results in opacities. 68 These opacities coincide with separation along the transition zone of the apical surfaces of lens equatorial epithelial cells and newly differentiating fiber cells at the epithelial-fiber interface (EFI), 68 revealing a requisite role for the stable microtubule population in lens morphogenesis. In addition, these studies demonstrated that loss of acetylated microtubules resulted in aberrant directionality of movement of the anterior tips of lens fiber cells as they elongated, as well as dysregulation of actin organization along lateral cell–cell borders of both lens epithelial and fiber cells and increased activation of myosin. 68 These studies using the lens as a model system both reinforce previously known functions of microtubules, while also providing greater insight into the developmental process as a whole and how cytoskeletal elements’ interactions and interrelation underlie crucial cellular processes.
Microtubule stability and dynamics: Expanding understanding of roles
Microtubules are traditionally characterized by their property of dynamic instability41–44 (Figure 1) as described in a variety of cell populations including brain, kidney, and cell lines. However, tubulin itself is a highly modifiable protein, subject to a variety of post-translational modifications, including detyrosination,109,110 glutamylation,111,112 polygyclation, 113 acetylation,114,115 and phosphorylation of serine112,116 and tyrosine 117 residues. Many of these have been examined in brain tissue, which greatly relies on axonal plasticity, with dynamic growing and shrinking and directionality, as well as incorporation of a neurofilament network as an essential cytoskeletal component. These modified forms of tubulin are generally found to accumulate in the subpopulation of stabilized microtubules within a cell48,118–121 (Figure 1), as demonstrated in brain, kidney, ovary, intestine, and fibroblast cells. These post-translational modifications and changes in microtubule stability also provide several alterations in terms of microtubule functioning including microtubule interaction with other cytoskeletal elements122,123 as well as microtubule roles in cellular processes such as morphogenesis48,124,125 including brain development and regeneration, 126 barrier function127,128 of the epidermis and vasculature, cytokinesis, 129 and cell migration 130 as described in multiple systems including neurons and fibroblasts.
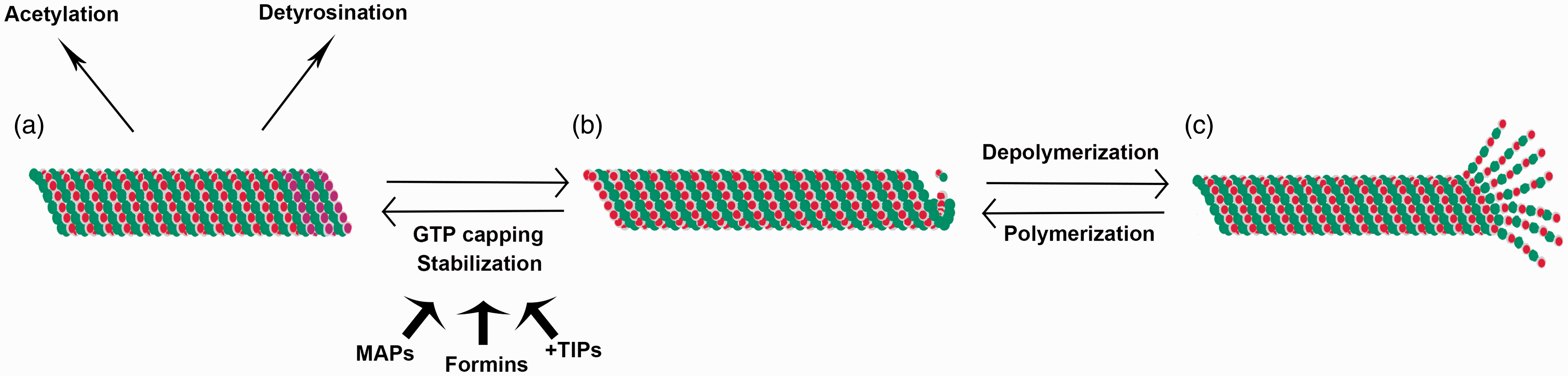
Microtubule stabilization and dynamic instability. Microtubules are typically characterized by cycles of polymerization and depolymerization, termed dynamic instability (b, c). However, some subpopulations of microtubules can be stabilized, a process that involves GTP capping of terminal α-tubulin monomers and is promoted by microtubule associated proteins (MAPs), formins and microtubule plus-end binding proteins (+TIPs) (a, b). This stabilization can then give rise to post-translational modifications of microtubules including acetylation and detyrosination (a).
Early work initially demonstrated the presence of a distinct stable microtubule population131,132 that proves resistant to depolymerization.133,134 From the beginning, stability was linked to differences in MAPs 135 as well as post-translational modification.49,110 In multiple systems, both acetylation as well as detyrosination have been shown to correlate with microtubule stability.49,118,136,137 It has been difficult to identify whether the posttranslational modifications are the cause of increased stability or a consequence of stable microtubules being a longer lived population due to prior modification such as end capping.49,121,138 Tyrosination or detyrosination involves the removal or re-addition of a tyrosine residue on the C-terminus of α-tubulin. 48 Detyrosination is increased artificially when microtubules are exposed to stabilizing treatments54,139 and detyrosinated microtubules show resistance to depolymerization by microtubule antagonistic drugs, 137 preventing microtubule disassembly by blocking interaction with microtubule depolymerizing motor proteins.140,141 These detyrosinated microtubules have specialized organization and distribution in both morphogenesis and cellular migration. In migrating cells, those cells responsible for leading wound healing have been shown to be rich in detyrosinated microtubules and are responsible for appropriate orientation of migration. 54 Additionally, the presence of detyrosinated microtubules has been linked to morphogenetic change and differentiation.142–147 The distribution of detyrosination may act as a cellular cue to help direct cellular polarity. 148 More recent work has shown that tyrosination/detyrosination of tubulin impacts other cellular functions, including interaction of microtubules with microtubule plus end tracking proteins50,140 as well as molecular motors, such as kinesin-1. 149 As will be discussed further later, these detyrosinated microtubules have also been shown to preferentially interact with intermediate filaments, specifically vimentin, and help to localize vimentin. 150
Although detyrosination has been the most studied tubulin modification, acetylation has also been greatly linked to microtubule stabilization. Acetylated microtubules are highly enriched in certain cellular structures including the mitotic spindle, centrioles, and cilia 51 as well as being highly accumulated in stable microtubule subpopulations.118,151–153 Changes in acetylation have been linked to several disease states including Huntington’s, 154 polycystic kidney disease, 155 Alzheimer’s, 156 and Parkinson’s. 157 Acetylated microtubules have been shown to play a role in cellular polarity52,158,159 as well as contact inhibition of proliferation and cellular adhesion. 160 Recent work also shows a functional role for acetylated microtubules in lens fiber cell morphogenesis, elongation, and directionality. 68 Acetylated microtubules also show differential interactions with microtubule motors 51 and organelles.161–163 Interestingly, formin proteins, traditionally known to function as actin nucleators, have been shown to act as microtubule stabilizers and help control microtubule acetylation.164,165 Formins promote microtubule stabilization by promoting end capping 166 while also promoting expression of the tubulin acetylation protein α-TAT1 gene.167,168 The overlapping functions of formin proteins on both actin and microtubule proteins demonstrate a potential coordinate activity between cytoskeletal elements in cellular functioning. 167
Beyond these two better studied post-translational modifications associated with microtubule stability, newer work has also investigated the roles for polyamination and transglutamination in microtubule stability. 169 This was examined in neuronal axons, where stable microtubules serve as the structural framework and tracks for axonal transport, while dynamic microtubules are more linked to reorganization and repair during neurite growth and remodeling.170–172 Transglutaminase activity and polyamine levels were known to increase with brain maturation and neuronal differentiation, 173 and were found to be modulators of microtubules by promoting stability.53,169 Further research will likely further develop our understanding of how post-translational modifications and their link to microtubule stability impact microtubule function. New approaches may help to differentiate between microtubule posttranslational modification and microtubule stability to better understand the differential roles these play in microtubule functions.
In addition to the correlation between certain post-translational modifications and microtubule stability, certain MAPs play a role in microtubule stability as well (Figure 1). Among these are several structural MAPs that in binding to microtubules affect their stability and polymerization. 174 In particular, all MAP1 proteins have been associated with a stabilization function,58,175–177 with MAP1B playing a larger role in modulating microtubule stability and catastrophe depending on its phosphorylation state.178–180 Other MAPs, including MAP4, also enhance tubulin polymerization and stability, potentially by sterically blocking disassembly,180–182 again with phosphorylation of the MAP regulating its activity.183–185 In addition to these, tau, a MAP highly studied in neurodegeneration, also promotes axonal microtubule stabilization,186–188 and work has demonstrated that tau stabilizes a protofilament conformation that favors the formation of microtubules, and increases microtubule stability.189,190 Recent work has shown that this tau binds at the interface between tubulin heterodimers to promote stability.57,191
Microtubule plus end binding proteins (+TIPs) also have a role to play in microtubule stability. Regulators of prototypical +TIPs including CLIP-170, specifically a family of proteins called CLASPs, have been identified as microtubule stabilizers and also help to orient stable microtubules toward the leading edge of migration.192–194 Other +TIPs involved in microtubule stabilization include APC, which has been shown to be necessary for axonal outgrowth.195,196 Additionally, end binding proteins, particularly EB1, are involved in the formation of the EB-stabilizing cap at microtubule plus ends.197–201 Formins, previously recognized as promoters of microtubule stability, can also interact with +TIP proteins, including EB1, APC, and CLIP-170, and these interactions may also serve to promote microtubule stabilization downstream of Rho activity.152,202,203 Interestingly, new research also demonstrates a role for certain microtubule motor proteins in promoting microtubule stability as well 204 and that in particular kinesin Kif4 can induce microtubule stability by interactions with the formin-EB1 microtubule stabilization pathway. 205
Importantly, this subpopulation of stable microtubules, as opposed to those more characteristic dynamically unstable microtubules, has differing functions. In many cell types, there is a characteristic distribution of stable versus dynamic microtubules.45,68,129,206 Partly, this works to help establish cellular polarity, with stable microtubule populations generally found at the cell front versus dynamic microtubules more likely to be present at the rear of the cell.48,54 Further work to better understand microtubule stability and its various roles in the cell is essential to global understanding of microtubule function.
Microtubules as a keystone of cytoskeletal coordination
The cytoskeleton is comprised of three major subtypes—microtubules, actin, and intermediate filaments—that have distinct functional and morphological properties that are important for their individual functions. However, despite these distinctions, all three interact to form a complex network necessary for proper cell functioning. Dynamic interactions between these groups are critical for multiple cellular processes as well as maintaining cytoplasmic homeostasis. These interactions are just the beginning of our understanding of how microtubules, through interaction with other cellular elements, play critical roles in a variety of cellular functions.
Previous work has shed light on various coordinate roles for actin and microtubules, with their interaction being critical for such processes as cell division, migration, vesicle and organelle transport, axonal growth, and wound healing67,207,208 (Figure 2). These roles often rely on actin’s ability to guide as well as stabilize microtubules. 209 Previous work has shown that microtubules can grow along actin bundles210,211 and when actin becomes disorganized, microtubular growth can be inhibited, with redistribution and misdirectionality.21,195,212,213 More recent studies show that in turn microtubule depolymerization causes upregulation and reorganization of the actin cytoskeleton, 68 implying a mutual regulatory process.
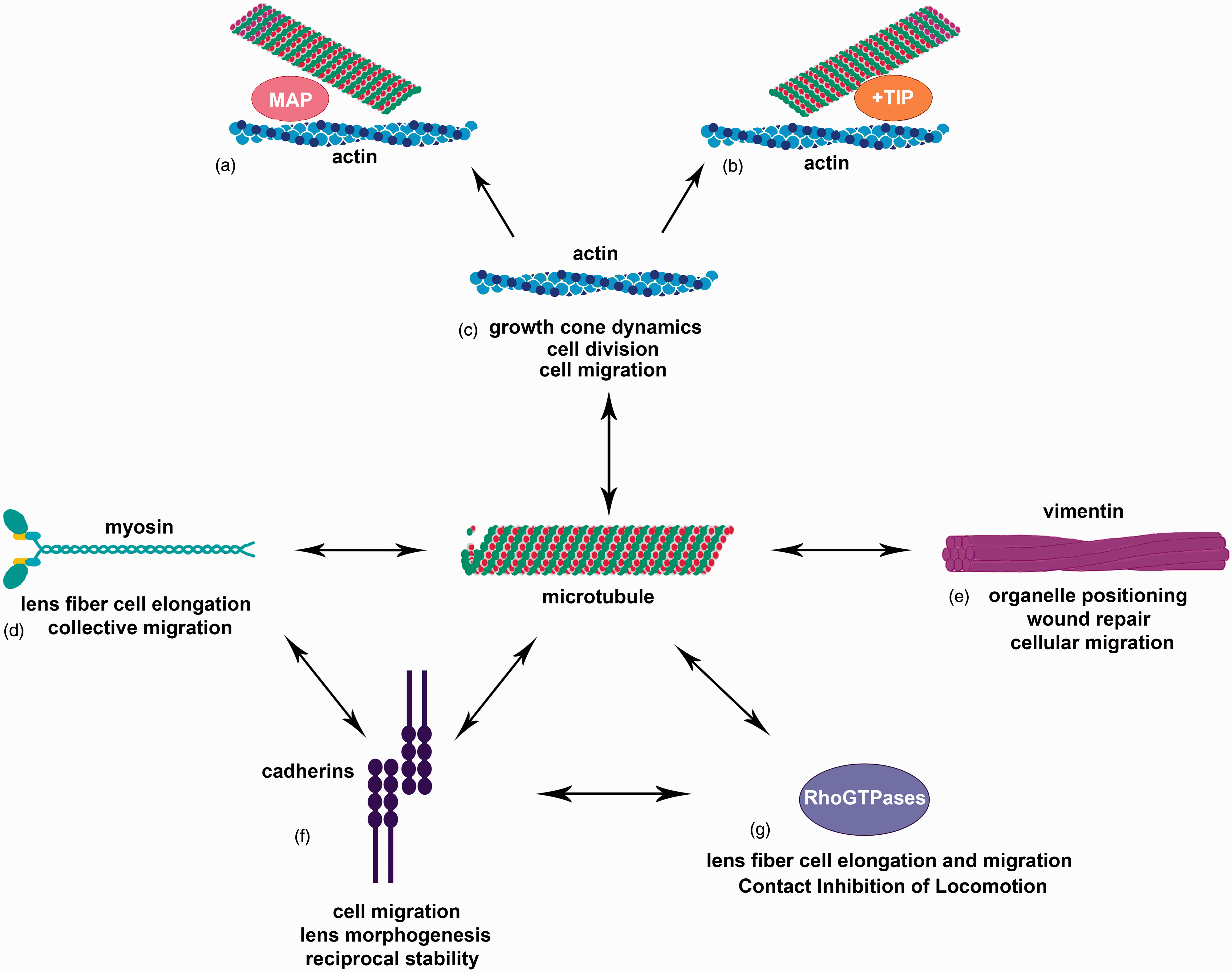
Microtubules as a cornerstone of cellular interaction. Microtubules interact with a variety of cellular components (a–g). Interaction with actin is important for growth cone dynamics, cell division, and cell migration (c) and is facilitated by microtubule-associated proteins (MAPs) (a) and microtubule plus-end binding proteins (b). Interactions with myosin have been seen in collective migration and lens fiber cell elongation (d). Intermediate filaments, including vimentin, interact with microtubules for organelle positioning, wound repair, and cellular migration (e). Cadherins, especially N-cadherin, have been shown to interact with microtubules as well as myosin and RhoGTPases (d, f, g). Cadherins and microtubules reciprocally regulate one another’s stability (f) and the interaction of cadherins with microtubules as well as myosin and RhoGTPases is important for cell migration and lens morphogenesis (d, f, g). RhoGTPases are also direct interactors with microtubules for processes such as lens fiber cell elongation and migration as well as Contact Inhibition of Locomotion.
One of the more studied aspects of this interaction has been in the field of growth cone steering and dynamics. In growth cones, actin filaments exhibit dynamicity, with a constant cycle of polymerization and severing, that is necessary for rapid axonal outgrowth.214–216 It is this instability of the actin cytoskeleton that is able to facilitate microtubule extension. 217 Microtubules in growth cones have been primarily linked as guidance sensors in growth cones controlling directionality as well as forward progression.218–221 For this to occur, F-actin networks provide tracks for microtubule growth, while actin dynamics allow for targeted distribution of microtubules to help oversee directionality.222–226
Actin filaments and microtubules can be crosslinked by direct linkage mediated by crosslinking proteins227,228 or indirectly via protein complexes or signaling molecules. Direct linkage by crosslinking proteins is usually considered to be static interactions, whereas indirect interactions mediated by motor protein complexes or signaling molecules are thought to be dynamic in nature. 229 Some of the key mediators of these interactions are MAPs, which come in multiple types. Among these are structural MAPs, including MAP 1, MAP 2, MAP4 and tau, that bind along the surface of microtubules and enhance stability and assembly, motor proteins such as dyneins and kinesins and microtubule plus end tracking proteins (+TIPs) that interact with the growing (plus) end of microtubules as well as other protein complexes. 229
For many structural MAPs, their expression is developmentally regulated, including MAP1, MAP2, and tau230–235 particularly in neurons. Previously, it was shown that these proteins indirectly regulated actin by modulating microtubule dynamics and can act as structural links to hold these filaments together.236–238 However, studies have also shown that these proteins can act to bundle as well as crosslink actin filaments independent of their actions on microtubules, with differential abilities among different classes of structural MAPs.229,239,240 The properties of these MAPs, including their regulation and interaction with cytoskeletal proteins, are also impacted by post-translational modifications, similar to microtubules themselves.241,242 Additionally, tau-mediated changes in actin organization have been linked to the development of neurodegenerative disorders such as Alzheimer’s and taupathies. 243 Future research to better understand the in vivo role that these proteins play in dynamic coordinated interactions is needed to better understand the mechanisms of their function.
+TIPS are the other major subgroup of microtubule interacting proteins that can regulate crosslinking with other cytoskeletal proteins. These proteins can interact not just with microtubules but also actin filaments and regulatory kinases, which can induce microtubule-actin crosslinking.195,196,223,244–248 They have been linked as mediators of actin-microtubule interaction for growth cone guidance and regulated distribution of +TIPS can promote interaction of F-actin and microtubules that allows for effective growth cone steering.244,246,247,249–251 Some of the more studied of these +TIPS include CLASP, which binds actin and microtubules through a shared domain 247 and whose loss results in both disrupted actin morphology as well as impaired axon outgrowth. 250 Additionally, +TIPS such as APC can act as microtubule stabilizers as discussed above, which can promote their outgrowth.195,196 These +TIPS do not only act on microtubules, but also may have a role in microtubule-F-actin coupling.252,253 Other studies have looked at end-binding (EB) proteins as regulators of microtubule dynamics254,255 and studies within the lens demonstrate a coordinate localization between EB1 and acetylated stable microtubules. 68 These proteins are also important scaffolding proteins that can recruit actin modulators as well as MAPs that alter actin dynamics.175,245,256–258 Of note, one of the major classes of proteins known to interact with both microtubules and actin cytoskeletal elements is spectraplakins.259–264 Some of these essential interactions are facilitated by spectraplakin binding to cytoskeleton through interaction with end binding proteins such as EB1.244–265 Just as importantly, +TIPS can also act as microtubule-actin uncouplers, including LIS1. 266
Newer evidence also expands the potential role of +TIPS in terms of its role in actin functioning, including serving as sites that facilitate F-actin nucleation. This is likely related to interactions between APC and mDia, a formin linked to actin remodeling267–269 and also an influencer of microtubule stability.153,270,271 Future research will likely reveal expanding ways by which different cytoskeletal elements impact one another and evolving roles that microtubule-actin crosslinking plays in a variety of cellular processes.
It is not only actin that has been shown to interact with microtubules, but also certain intermediate filament subtypes, in particular, vimentin (Figure 2). In earlier work, it was demonstrated that distribution of vimentin corresponded to microtubule distribution 272 and depolymerization of microtubules causes retraction of the vimentin network to the perinuclear region of the cell.273,274 Additionally, studies suggest that vimentin as well as other intermediate filaments, including neurofilaments, can be rearranged by microtubule-associated motor proteins, kinesin, and dynein, implying that vimentin distribution relies on microtubule transport.275–280 Further research demonstrates that vimentin filament transport is dependent on microtubules and their acetylation state but not impacted by microtubule dynamics or polymerization. 281 Additionally, these studies also showed that vimentin motility was also linked to interactions between vimentin and polymerized actin that may in turn be impacted by microtubules.281–283
This interaction between microtubules and vimentin is necessary for several cellular processes. This includes proper positioning of organelles requires the combined efforts of microtubules as well as intermediate filaments such as vimentin and neurofilaments.284–287 Studies in a mock cataract surgery model demonstrated that collapse of intermediate filaments, and in particular, vimentin, following microtubule depolymerization results in impaired wound closure. 288 These studies, as well as others, demonstrate a role for vimentin-microtubule interaction in cellular migration. Vimentin-null fibroblasts demonstrate impaired locomotion and it is possible that normal locomotion relies on the associations between vimentin and MTs and their molecular motors to provide for rapid turnover and reorganization of the vimentin network.276,289,290 Recent work on astrocytes has shown that APC, already known to interact with microtubules and control their organization in migrating astrocytes291–293 is necessary for microtubule interaction with intermediate filaments in these cells. 294 More importantly, it also controls the microtubule-dependent rearrangement of IFs during astrocyte migration, as well as having a role in vimentin organization in other cell types as well. 294
Beyond interaction with cytoskeleton, microtubule functioning through reciprocal regulation with cell–cell adhesion molecules is becoming better understood. Both N-cadherin and microtubules have been demonstrated to be critical in regulating cellular migration processes.38,66,67,70,72,295–299 In many instances, the cell–cell adhesions and cytoskeleton have been shown to work coordinately in regulating these processes, with classical and non-classical cadherins traditionally associated with the actin and intermediate filament elements of the cytoskeleton. However, there are several insights into how microtubules and cadherin junctions may interact to function together in cellular processes. 245
Microtubules have been shown to directly interact with cadherin junctions in other systems300,301 and potentially regulate the presence of cadherin junctions at the cell surface for cell–cell contact302,303 (Figure 2). In addition to this, microtubule depolymerization has also been shown to disrupt integrity of cadherin junctions as well.304–306 This effect is not limited solely to microtubule depolymerization but also applies to impedance of microtubule dynamics or overstabilization. 303 It is possible that the function microtubules play in maintaining cadherin junction integrity is intrinsic to the role that microtubules play in regulating vesicle transport, with N-cadherin having been shown to traffic along microtubules.307,308 Clearly, there is a dependence of N-cadherin on microtubules for proper function.
Conversely, cadherin junctions have also been shown to regulate microtubules. Microtubule polymerization has been linked to interaction with the actin cytoskeleton for guidance, with cadherin junctions playing roles in anchoring actin bundles at the cell surface.309,310 Additionally, cadherins have been shown to also play direct roles in recruiting microtubules to sites of cell–cell contact. 303 Microtubules have been shown to anchor to cadherin junctions304,305 and p120 catenin, an atypical catenin found at cadherin junctions, has also been demonstrated to directly interact with microtubules.311,312 These interactions can play a role in regulating microtubule dynamics by either increasing or decreasing microtubule stabilization.69,313–315 New work has shed insight into how interaction between cadherins and microtubules, along with actin, may be critical in how cells sense guidance cues and underlie mechanoresponsive behavior316,317 and suggest that polarization of cell–cell junctions as well as the actin cytoskeleton that is crucial cellular processes including migration and angiogenesis are dependent on microtubule dynamics as well as RhoGTPases. 318
It is therefore not surprising that systems such as the lens would have interactions between the microtubule cytoskeleton and N-cadherin junctions. Recent findings show that microtubules, especially acetylated microtubules, and N-cadherin junctions interact strongly in the cortical fiber cell region, the site of active lens fiber cell elongation, and that stable microtubules are necessary for appropriate localization and likely functioning of N-cadherin junctions. 68 However, loss of N-cadherin does not disrupt the presence and assembly of these acetylated microtubules, nor does the presence of these microtubules alone prevent dysmorphogenesis demonstrating the necessity of both proteins. 319
Since the loss of either N-cadherin or stable microtubules also causes alteration in actin distribution as well as an aberrant overactivation of myosin II, it is likely that the regulation of the actomyosin network plays a critical role in N-cadherin and microtubule coordinate activity within the lens68,319 and plays a part in the process of lens fiber cell elongation and migration, given active myosin’s role in cell contractility and movement.79,320–323 Microtubule functioning therefore relies not just on interactions with other cytoskeletal proteins but also further expands to include other cellular networks (Figure 2).
Myosin II activity has been shown to serve as a regulator of both microtubule stability and N-cadherin distribution,64,295,324–326 both of which have proven necessary for lens fiber cell elongation. Myosin activation has been shown to antagonize microtubule acetylation, with the converse also proven true. 64 Additionally, myosin activity has been shown to weaken the ability of cells to migrate coordinately when intercellular junctions are compromised; a block in myosin activity can rescue collective migration even when cadherins are lost. 325 Beyond this general role in the process of collective migration, active myosin II has been shown to regulate the concentration of cadherin junctions at cell–cell interfaces326,327 and this myosin activity may depend on dynamic microtubules. 303 This may explain the stable microtubule dependent, myosin activity mediated N-cadherin distribution in the lens epithelium and fiber cells.
The roles myosin may play in relation to microtubules and N-cadherin are likely also dependent on actin regulation. While myosin can be activated by myosin light chain kinase, its activity is also regulated by Rho-dependent kinase (ROCK). Through this downstream regulation, ROCK has been shown to mediate effects on the stability of microtubules 65 and the distribution of N-cadherin 326 (Figure 2). In addition to this downstream regulation of myosin activity, RhoGTPases also play a large role in modulating N-cadherin and microtubule functions.
Interestingly, the regulation of the stabilization of microtubules has been linked to the RhoGTPase family.152,153,270,328–333 Multiple reports have shown that RhoA can promote the stabilization of microtubules,270,328 while Rac1 can inhibit the destabilization of microtubules.334–336 The active growth and stabilization of microtubules have been shown to promote activation of Rac1 and lamellipodial protrusions,315,337,338 creating a potential feedback loop that allows for cellular migration which may tie microtubules to Rac1’s role in lens fiber cell elongation and migration.
Rac1 involvement in lens fiber cell elongation and migration has been previously documented, 82 with lens conditional deletion of Rac1 preventing elongation of lens fiber cells along both the posterior lens capsule and the epithelial fiber cell interface and is now confirmed in our N-cadherin knockout system. RhoGTPases are also linked to regulation of cadherin cell–cell adhesion.339–345 It is likely therefore that there may exist an integral role for the multifold regulation of these RhoGTPases by cell–cell adhesion molecules, the cytoskeleton as well as signaling cascades including FGF 346 that allows for proper lens fiber cell elongation and migration, which may explain the high association of N-cadherin junctions and microtubules particularly in the area of fiber cell elongation. 68 This interconnectivity may also provide insight into the effects of microtubule depolymerizing agents on the stability and solubility of N-cadherin junctions within the lens.
The RhoGTPases’ role in migration has also been greatly tied to Contact Inhibition of Locomotion (CIL), a process that relies on both cadherins and microtubules as mediators. In CIL, N-cadherin-mediated cell–cell contact formation regulates RhoGTPase activity, causing a switching in Rac1 versus RhoA localization and activation within the cell that can help determine the directionality of further motion.296–298,347–349 This requires an alteration of cell polarity, mediated by RhoA, Rac1, and ROCK regulation of microtubule dynamics, causing a switch in the localization of stable microtubules to lead the new directionality of motion.350–352 Given the importance of this mechanism for collective movement of cells during development, it is possible that this interplay between N-cadherin and microtubules mediated through RhoGTPases may be a critical part of overall lens morphogenesis.
Conclusions
Microtubules are a critical aspect of the cellular cytoskeleton. Newer studies in the lens have given insight into the continuously evolving roles of microtubules, both dynamic and stable subpopulations, in a variety of cellular processes. The dynamic versus stable microtubular interplay is an evolving field that leads to a deeper look into how microtubule modifications and interaction with other cellular elements also are integral in cellular functioning. More and more research points to the cytoskeletal network of the cell as highly interactive. Microtubules are becoming more and more understood as a key integrator with other cytoskeletal proteins including actin and intermediate filaments, as well as cell–cell junctional proteins and other cellular regulators including myosin and RhoGTPases. For appropriate cellular homeostasis, all of these elements must interact in precise balance, with microtubules playing an integral role in this balance.
Footnotes
Authors’ contributions
CML wrote the manuscript and designed the figures with critical oversight input, and review from ASM.
DECLARATION OF CONFLICTING INTERESTS
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
FUNDING
This work was supported by grants EY014258 and EY 026478 from NEI to ASM.