
Abstract
Annona atemoya is a hybrid of Annona squamosa and Annona cherimola that grow in several subtropical or tropical areas such as Florida in the US, Philippines, Cuba, Jamaica, Taiwan, and Jeju in South Korea. We report that the A. atemoya leaves (AAL) have inhibitory effects on the pathogenesis and regulatory mechanisms of Alzheimer’s disease (AD). Ethanol extract of AAL prevented amyloid-β (Aβ) aggregation and increased free radical scavenging activity. In addition, AAL extract exerted protective effects against neuronal cell death in HT22 hippocampal cells. Moreover, oral administration of AAL extract significantly improved memory loss in the passive avoidance task and Y-maze test, as well as downregulated the expression of neuronal markers neuronal nuclei and brain-derived neurotrophic factor in Aβ-injected AD mice. To verify the molecular mechanisms responsible for anti-AD actions of AAL, we conducted the antibody microarray analysis and found that epidermal growth factor receptor/G protein-coupled receptor kinase 2 signaling was activated in neuronal cells and AD-like mouse models. Additionally, quantitative analyses of the six standard compounds using high-performance liquid chromatography revealed that rutin is the most abundant compound of AAL. Furthermore, efficacy analyses of six standard compounds showed that rutin and isoquercitrin had significant inhibitory activity on Aβ aggregation. Taken together with biological activity and the content of compounds, rutin maybe a bioactive compound of AAL in the AD pathogenesis. Overall, our findings provide the first scientific support for the therapeutic effects of AAL in AD and AD-related disorders.
Impact statement
Our study was aimed to find a novel candidate drug for Alzheimer’s disease (AD) using natural products. We assessed the effects of Annona atemoya extracts on crucial events in the pathogenesis of AD. A. atemoya leaf (AAL) extract significantly inhibited amyloid-β aggregation, oxidative stress, neuronal cell death, and memory impairment through the epidermal growth factor receptor/G protein-coupled receptor kinase 2 pathway. Simultaneous analysis using HPLC determined six standard compounds of AAL extract, and rutin was identified as a bioactive compound. Of note, the anti-AD activity of AAL extract was more significant compared to other extracts from medicinal plants of which efficacy was previously reported. The potential of AAL extract as an anti-AD agent may provide insight into the new drug development for AD treatment.
Introduction
Alzheimer’s disease (AD) is a progressive neurodegenerative disease characterized by cognitive disruption. AD is considered life-threatening; yet, no cures for this disease exist. Currently, there are five medications approved by the US FDA for AD: tacrine, rivastigmine, galantamine, donepezil, and memantine. These drugs are sufficient to offer symptom relief 1 but have side effects such as diarrhea, nausea, muscle cramps, vomiting, fatigue, lowered blood pressure, insomnia, and loss of appetite. 2 Molecular targets of the approved medications are acetylcholinesterase (AChE) or N-methyl-D-aspartate (NMDA) receptors. However, the limitations of AChE- or NMDA receptor-targeting drugs are becoming evident as they lack the ability to cure AD.
Amyloid-β (Aβ) is a crucial mediator in AD and is considered an attractive target molecule for developing novel AD medications. According to the amyloid hypothesis, the overproduction or accumulation of Aβ in extracellular senile plaques occurs early in disease and is a central event in AD progression. Thus, controlling the production or accumulation of Aβ plaques could be an effective treatment and help to prevent AD. The amyloid hypothesis is regarded by many as a central description for AD progression, although recent failures of anti-Aβ drugs in clinical trials have been reported.3,4 Actually, the neuropathogenesis of AD caused by abnormal Aβ deposition is primarily initiated and occurred by oxidative stress. 5 Thus, we investigated to find novel AD drug candidate(s) targeting Aβ with robust antioxidant activity.
Annona atemoya, also known as atemoya, is a hybrid between A. squamosa (sugar apple) and A. cherimola (cherimoya) that was first crossed by Wester in 1908, a horticulturist at the USDA’s Subtropical Laboratory in Miami. A. atemoya is distributed in the subtropics and tropics such as Florida in the US, Philippines, Cuba, Jamaica, Taiwan, and Jeju in South Korea. A. atemoya fruit (AAF) is heart-shaped or round with pale-green and bumpy skin. It is used as an ingredient in juices, desserts, ice creams, or in natura.6–8 Bullatacin, an acetogenin isolated from AAF, has been reported to have anti-cancer activity in hepatoma cells.9,10 A. atemoya seed (AAS) was recently reported to have anti-angiogenic properties. 11 However, there are no reports on the biological activity of A. atemoya leaves (AAL). In the present study, we demonstrate that AAL extract possess anti-AD effects, and we demonstrate its molecular mechanisms using in vitro and in vivo experimental models. In addition, we established the simultaneous analysis methods of six standard compounds for quality control and identified the bioactive compound from AAL extract.
Materials and methods
Preparation of ethanol extract from A. atemoya, Ginkgo biloba (GB), Lycium chinense (LC), and Angelica gigas (AG)
A. atemoya was supported by Jeju Sunny Farm (Jeju, South Korea). The materials (AAL, AAF, and AAS) were dried and 2.7 kg of each was extracted twice with ethyl alcohol (60 L) for 3 h using the COSMOS-660 electric extractor (Kyungseo Machine Co., Incheon, South Korea). The extracted solutions were filtered, evaporated, and freeze-dried for making powdered extracts. GB, LC, and AG were obtained from the Naemome Dah (Ulsan, South Korea). Each material was dried and 50 g of each was extracted twice with ethyl alcohol (0.3 L) by refluxing for 2 h. The extracts were filtered and evaporated using a rotary evaporator.
Solvent fractionation of AAL
The powdered AAL extract (10 g) was suspended in H2O (0.2 L) and in turn partitioned with n-hexane (HX), ethyl acetate (EA), and n-butanol (BU) to give extracts of 0.413 (4.13%), 0.424 (4.24%), and 1.373 g (13.73%), respectively (Figure 3(a)).
Aβ aggregation assay
Aβ aggregation assay was performed using SensoLyte® Thioflavin T β-Amyloid aggregation kit (AnaSpec, Fremont, CA). Aβ1–42 peptides were stored at −80°C and used at 100 µg/mL for the assay. Thioflavin T dye was prepared by dissolving in the Assay buffer included in the kit. AAL extract was dissolved in the Assay buffer (100 µg/mL of final concentration). Thioflavin T (85 µL) and AAL sample solution (5 µL) were mixed and incubated in 96-well black microplate at 37°C. Fluorescence readings were expressed as the relative fluorescence units. All assays were completed in triplicates. The inhibition rate (%) of Aβ aggregation was assessed as described previously. 12
Free radical scavenging activity
The free radical scavenging activity of ABTS and DPPH was used in the modified protocol of Re et al.
13
and Moreno et al.,
14
respectively. The absorbance of control or tested samples was assessed at 734 and 517 nm, respectively, and the activity for radical scavenging was calculated using the following equation
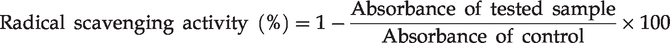
Cell lines, maintenance and treatment
Murine hippocampal cell line HT22 were cultured in Dulbecco's modified Eagle's medium (Hyclone/Thermo, Rockford, IL), containing 10% fetal bovine serum (Hyclone/Thermo) and antibiotics penicillin/streptomycin at 37°C in a 5% CO2 atmosphere. HT22 cells were concurrently treated with AAL, AAF, or AAS, and hydrogen peroxide (H2O2, 250 µmol/L, Sigma-Aldrich, St Louis, MO) for 6 h.
Cell viability test
The cytotoxic effects of AAL, AAS, and AAF extracts were evaluated using CCK-8 assay kit (Dojindo Molecular Technology, Kumamoto, Japan). HT22 cells were seeded onto the 96 well micro-culture plates (3 × 104/well) and treated with AAL, AAS, or AAF extracts for 24 h and incubated with CCK solution for additional 4 h. Cell viability was determined using Epoch Microplate Spectrophotometer (BioTek Instruments, Inc., Winooski, VT)

Brain-derived neurotrophic factor (BDNF) ELISA
An ELISA kit for BDNF was commercially obtained from R&D Systems (Minneapolis, MN). The culture supernatant was collected and applied to each well containing anti-BDNF. The amount of BDNF of each sample was determined using a BDNF standard curve.
Phospho-specific protein microarray analysis
Antibody array was performed using Full Moon Biosystem’s Phospho Explorer Antibody Array kit (Sunnyvale, CA). Data normalization was conducted using the Genowiz 4.0 (Ocimum Biosolutions, Gaithersburg, MD) and determined the average intensity of samples comparable to that of all genes. For statistical analysis, MeV software ver. 4.9.0 (TM4 Development Group, Rockville, MD) was used for one-sample t tests. UniProt DB was used for annotation of the protein data and the phosphorylation ratio was determined as described previously. 15
Western blotting
HT22 cell lysates were prepared using CelLytic™ MT Cell Lysis Reagent (Sigma-Aldrich). Tissues from mouse hippocampus were lysed in lysis buffer containing 1% Triton X-100, 1% deoxycholate, 0.1% sodium azide (Sigma-Aldrich), and protease inhibitors (Roche Diagnostics, Mannheim, Germany). Lysates were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis using the 4–20% Mini-PROTEAN TGX Precast gels (Bio-Rad, Hercules, CA) and transferred to polyvinylidene fluoride membranes. For blocking the membranes, 5% skim milk in Tris-buffered saline containing Tween 20 (TBST) was used. The membranes were let an overnight incubation at 4°C with the desired primary antibodies for anti-phospho-Ca2+/calmodulin-dependent protein kinase II (CaMK2) β/ν/δ, phospho-epidermal growth factor receptor (EGFR), phospho-G protein-coupled receptor kinase 2 (GRK2), phospho-Myc, phospho-ferric reductase (FER), phospho-caveolin-1, phospho-nuclear factor kappa B (NFκB) p65, phospho-myosin regulatory light chain (MLRN) 2 (Cell Signaling Technology, Beverly, MA), and glyceraldehyde 3-phosphate dehydrogenase (GAPDH; Santa Cruz Biotechnology, Santa Cruz, CA). After washing the membranes with TBST, they were incubated with anti-mouse or rabbit secondary antibody conjugated with horseradish peroxidase (Jackson Immuno Research, West Grove, PA) at room temperature for 1 h. After washing the membranes with TBST, they were developed using the Pierce ECL Western Blotting Substrate (Thermo Scientific, Rockford, IL).
Animals
Eight-week-old imprinting control region (ICR) male mice were obtained from Dae Han Bio Link (Eumseong, South Korea). Mice were bred in environmentally managed specific pathogen-free area at 22°C with light and dark cycling (12 h each) with providing water and standard food pellets ad libitum. Mice were grouped into five (n = 7/group): normal control, Aβ only, Aβ + AAL extract (50 or 100 mg/kg/day), and Aβ + morin (10 mg/kg/day). All animal experiments were carried out according to the National Institutes of Health Guidelines for the Care Use of Laboratory Animals. In addition, all assays were approved by the Institutional Animal Care and Use Committee of the Korea Institute of Oriental Medicine (Approval number: #17-034). Animal handling was conducted according to the dictates of the National Animal Welfare Law of Korea.
Intracerebroventricular (icv) injection of Aβ1-42 aggregates
Aβ1-42 peptide (AnaSpec) was dissolved in PBS to 1 mM concentration and incubated at 37°C for five days to form Aβ aggregates. Aβ (10 µmol/mouse) was infused (coordinates from bregma: anteroposterior = −0.5 mm, mediolateral = 1 mm, and dorsoventral = 2.5 mm) based on the brain in stereotaxic coordinates. A Hamilton syringe (26-gauge stainless-steel needle) was used to deliver Aβ aggregates (3 µL/mouse) at a rate of 1 µL/min into the icv region. Body temperature was maintained at 36.5 ± 0.5°C. Normal control group was treated by the same procedure, except that the same volume of PBS was injected instead.
Passive avoidance test
A test box with two of equally sized chambers (40 × 20 ×30 cm3) divided by a guillotine door (5 × 5 cm2); one part was lit with a 60-W bulb and the other part was dark. Floor in the dark box was composed of several stainless steel rods (5 mm) set 1 cm apart. All mice were daily fed either vehicle or AAL extract for 23 days. Two weeks after Aβ administration, each mouse was placed into the light side and allocated to travel for training, and the guillotine door was opened to allow the mice to move to the dark area. The same processes with above described were repeated next day with an electric foot shock (0.5 mA) for 3 s for immediately delivering to a mouse via the grid floor after moving the dark side. Latency time to the dark was recorded up to 300 s.
Y-maze test
The Y-maze test was conducted at day 18 after Aβ injection. Y-maze was composed of three black colored wood arms (40 long × 12 high × 3 wide cm3 at the bottom, and 10 cm wide at the top). The arms converged at a symmetrical triangular central point. A mouse was allowed to place at one arm and allocated to freely travel through the maze for 8 min. Percent of the alternation behavior was determined as a successive entry into each arm on overlapping triplet sets
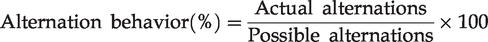
Immunohistochemistry
Mice were immediately anesthetized after finishing behavioral tests and transcardially perfused with 0.05 mol/L PBS and ice-cold 4% paraformaldehyde in 0.1 mol/L phosphate buffer. Removed mouse brains were post-fixed in 4% paraformaldehyde in 0.1 mol/L phosphate buffer at 4°C for overnight and immersed in 30% solution of sucrose in 0.05 mol/L PBS. A freezing microtome (Leica Instruments GmbH, Nussloch, Germany) was utilized to obtain serial coronal sections (30 µm thick) and, the sections were kept in cryoprotectant (25% ethylene glycol, 25% glycerol, and 0.05 mol/L phosphate buffer) at 4°C. For immunohistochemical assays, briefly rinsed brain sections were incubated with rabbit anti-neuronal nuclei (NeuN) (Merck Millipore, Darmstadt, Germany) at 4°C for overnight with normal goat serum and 0.3% Triton X-100. Then, the sections were reacted with anti-rabbit biotinylated IgG for 1 h, followed by incubation with ABC at room temperature for 1 h. After developing using DAB in 0.05 mol/L Tris-buffered saline (pH 7.6) to determine the peroxidase activity, histomount medium was added on gelatin-coated slides.
Chemicals and reagents
The AAL standard compounds (+)-catechin, chlorogenic acid, epicatechin, rutin, and isoquercitrin were provided by ChemFaces Biochemical Co., Ltd. (Wuhan, China); and kaempferol-3-O-rutinoside was obtained from Biopurify Phytochemicals (Chengdu, China). Purities of the chemicals were ≥98.0%. The high-performance liquid chromatography (HPLC)-grade solvents acetonitrile and water were purchased from J. T. Baker Chemical Co. (Phillipsburg, NJ). Trifluoroacetic acid (TFA) was purchased from Sigma-Aldrich (St Louis, MO).
Preparation of standard and sample solutions for HPLC analysis
For preparing sample solution, AAL extract was dissolved in 90% aqueous methanol (10 mg/mL of final concentration) and filtered using a syringe filter with 0.45 µm pore size. The stock solutions of six standard compounds were dissolved in methanol (1.0 mg/mL of final concentration). Working solutions of the standard compounds were prepared by diluting the stock solution to a final concentration of 0.1 mg/mL.
Chromatographic conditions
For HPLC analysis, Waters Alliance e2695 HPLC system (Waters Corp., Milford, MA) with a photodiode array (PDA) detector was used. Acquired data were processed using Empower software version 3. Chromatographic separation of the standard compounds was obtained using a Luna C18 analytical column (250 × 4.6 mm, 5 µm; Phenomenex, Torrance, CA) at 30°C. The mobile phases consisted of 0.1% (v/v) aqueous TFA (A) and acetonitrile (B). Conditions of the gradients were 10% B for 0–10 min, 10–50% B for 10–50 min, 50–100% B for 50–51 min, and 100% B for 51–60 min (injection volume: 10 µL, and flow rate: 1.0 mL/min). Ultraviolet (UV) wavelength of the PDA detector ranged from 190 to 400 nm.
Statistical analysis
Values were represented as mean ± SEM. Statistical significance was analyzed using one-way analysis of variance and Dunnett’s multiple comparisons test. Difference was considered if P value was less than 0.05.
Results
Effects of AAL extract on Aβ disaggregation and antioxidation
Amyloid deposition in the fibrillary aggregated form of Aβ is a crucial pathogenic mechanism underlying AD. 16 To explore whether A. atemoya had inhibitory effects on Aβ aggregation, ethanol extracts from AAL, AAS, and AAF were prepared. At 100 µg/mL, AAL and AAF extracts increased the relative percentage of Aβ disaggregation by 91.35% and 38.92%, respectively. In contrast, AAS had no effect on Aβ disaggregation (Figure 1(a), left). As AAL extracts were the most effective at promoting Aβ disaggregation, we further explored its effect on Aβ aggregation at a broad range of concentrations. As shown in Figure 1(b) (left), AAL extract exerted a dose-dependent inhibition of Aβ aggregation.
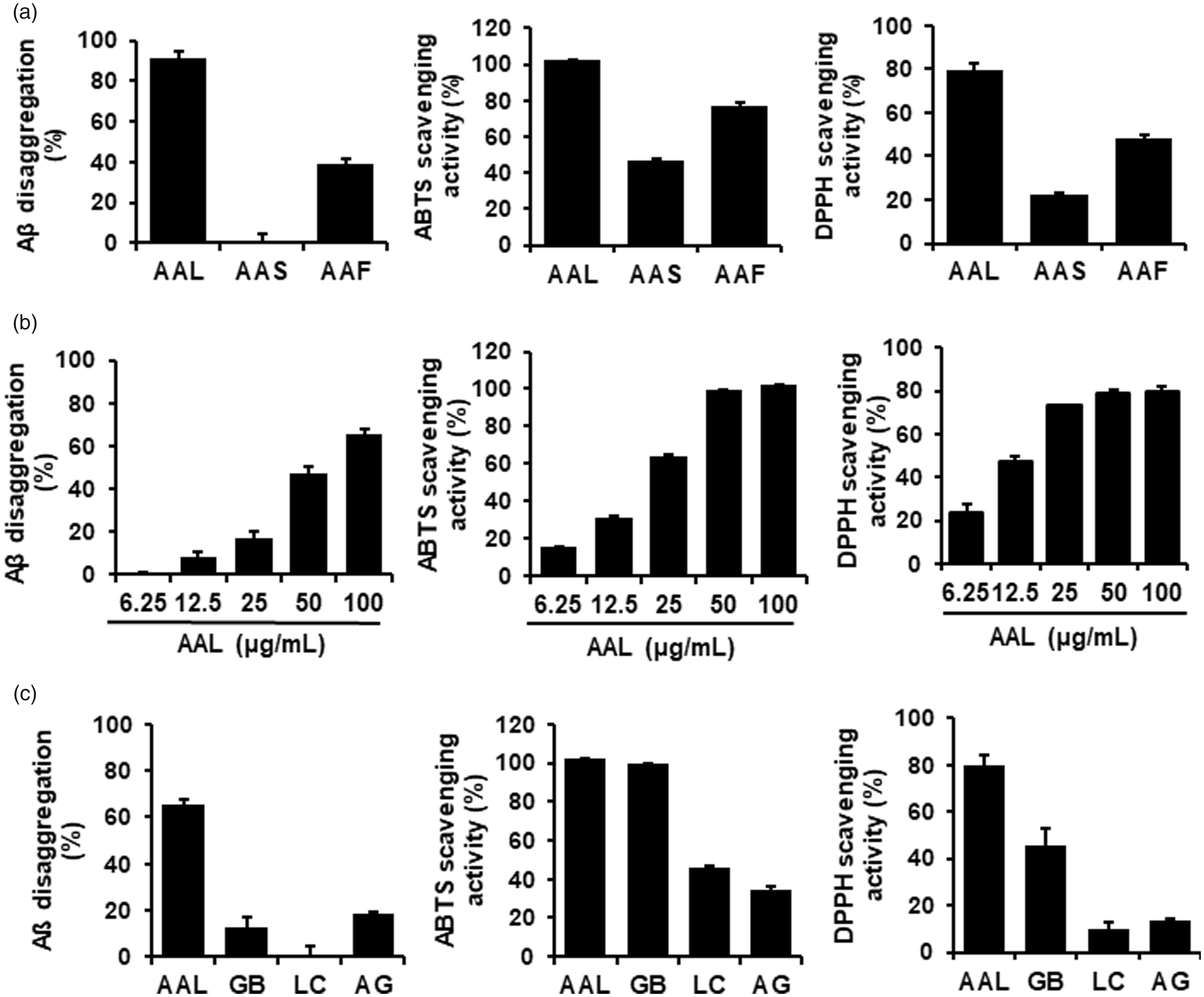
Effects of AAL extract on Aβ aggregation and antioxidation. Assays were conducted by reaction with AAL, AAS, or AAF extracts at 100 μg/mL (a); various concentrations (6.25, 12.5, 25, 50, or 100 μg/mL) of AAL extract (b); and AAL, GB, LC, or AG extract (c). (Left column) Aβ aggregation assay was performed using the SensoLyte® Thioflavin T β-Amyloid aggregation kit. Samples were reacted with Aβ (1–42) peptides, followed by the addition of Thioflavin T. Fluorescence of Thioflavin T was measured at an excitation wavelength of 440 nm and an emission wavelength of 485 nm. (Middle and right columns) Antioxidant activity was assessed by a free radical scavenging assay for ABTS (middle column) and DPPH (right column). Percent activity is equal to percent inhibition of ABTS and DPPH. Each value is presented as the mean ± SEM (n = 3). AAL: Annona atemoya leaf, AAS: A. atemoya seed, AAF: A. atemoya fruit, GB: Ginkgo biloba, LC: Lycium chinense, and AG: Angelica gigas.
Neurotoxic factors including Aβ in AD cause oxidative stress in the brain by perturbing the balance between oxidants and antioxidants. 17 The scavenging activities of A. atemoya extracts against ABTS or DPPH were measured to examine antioxidant activities. Consistent with the results of the Aβ aggregation assay, ABTS and DPPH scavenging activity was exhibited by AAL, AAF, and AAS extracts (Figure 1(a), middle and right, respectively) in decreasing order. AAL extract dose-dependently enhanced scavenging activity against ABTS and DPPH radicals (Figure 1(b), middle and right, respectively).
We compared the effects of AAL extract with GB, LC, and AG extracts on inhibition of Aß aggregation and activation of free radical scavenging. GB, LC, and AG are known medicinal plants that are effective on anti-AD.18–20
As shown in Figure 1(c), AAL showed robust Aß disaggregation activity compared with that of other extracts. Effects of AAL extract on antioxidation were similar to that of GB extract but higher than those of LC and AG extracts.
Neuroprotective effects of AAL extract on H2O2-damaged HT22 neuronal cells
Neuronal cell death is a major cause of neurodegenerative diseases including AD.21,22 To examine the effects of A. atemoya extracts on neuronal death, HT22 hippocampal cells were used. To determine the cytotoxicity of A. atemoya extracts, CCK assay, a sensitive colorimetric assay for the determination of cell viability in cytotoxicity assay, was performed. As shown in Figure 2(a), at concentrations up to 100 µg/mL, AAL and AAF extracts had no significant effect on cell viability. In contrast, AAS extract significantly reduced cell viability at 100 µg/mL by 56.2% compared with that of untreated control.
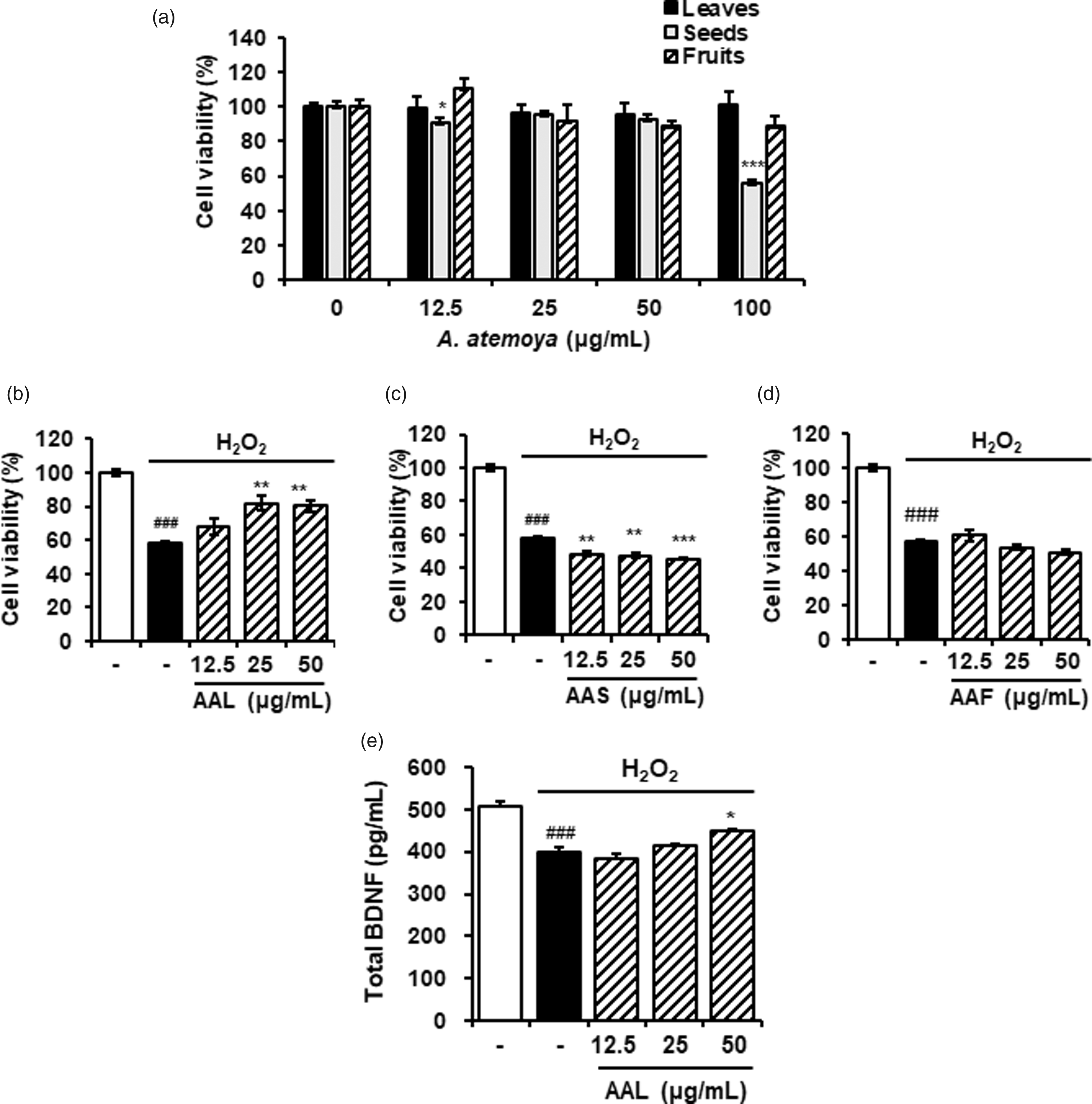
Neuroprotective effects of AAL extract in H2O2-damaged HT22 cells. (a) Cytotoxicity of AAL, AAS, or AAF extracts was measured using the CCK assay against HT22 cells. The cells were treated with the extracts for 24 h. (b) Cells were exposed to H2O2 in the absence or presence of AAL, AAS, or AAF extracts for 6 h. Cell viability was determined using a CCK assay kit. (e) The production of BDNF was measured in the culture supernatant and assessed using ELISA. The results are expressed as mean ± SEM of three independent experiments. ###P < 0.001 vs. vehicle control cells; *P < 0.05, **P < 0.01, and ***P < 0.001 vs. H2O2-treated cells. AAL: Annona atemoya leaf, AAS: A. atemoya seed, and AAF: A. atemoya fruit.
Neuronal cell damage was mediated by exposing the cells to H2O2 (250 mol/L), followed by the treatment with AAL, AAS, or AAF extracts at the range of non-toxic concentrations. The treatment with H2O2 alone significantly reduced the HT22 cell viability by 58.3% compared to untreated control. Of note, AAL extract significantly reversed H2O2-induced neuronal cell death at 25 or 50 µg/mL (Figure 2(b)) whereas AAS and AAF extracts did not inhibit H2O2-mediated cellular damage (Figure 2(c) and (d), respectively), indicating the neuroprotective effects of AAL, but not AAS or AAF. We further examined the neuroprotective effects of AAL by measuring the level of BDNF, a neuronal marker, 23 in H2O2-treated HT22 cells. H2O2 treatment significantly reduced the amount of BDNF compared to that of untreated control. In contrast, AAL extract prevented the H2O2-mediated inhibition of BDNF production at 50 µg/mL when compared to that of treatment with H2O2 alone (Figure 2(e)).
Effects of BU fraction on Aβ aggregation and neuroprotection
To determine which fraction of AAL extract exerted the greatest effects on Aβ aggregation and neuroprotection, we prepared solvent fractions of HX, EA, BU, and H2O from AAL ethanol extracts. All tested fractions increased the inhibition of Aβ aggregation in a dose-dependent manner. The inhibition percentages of HX, EA, BU, and H2O at 100 µg/mL were 34.9, 71.4, 90.2, and 66.0%, respectively (Figure 3(b) to (e)).
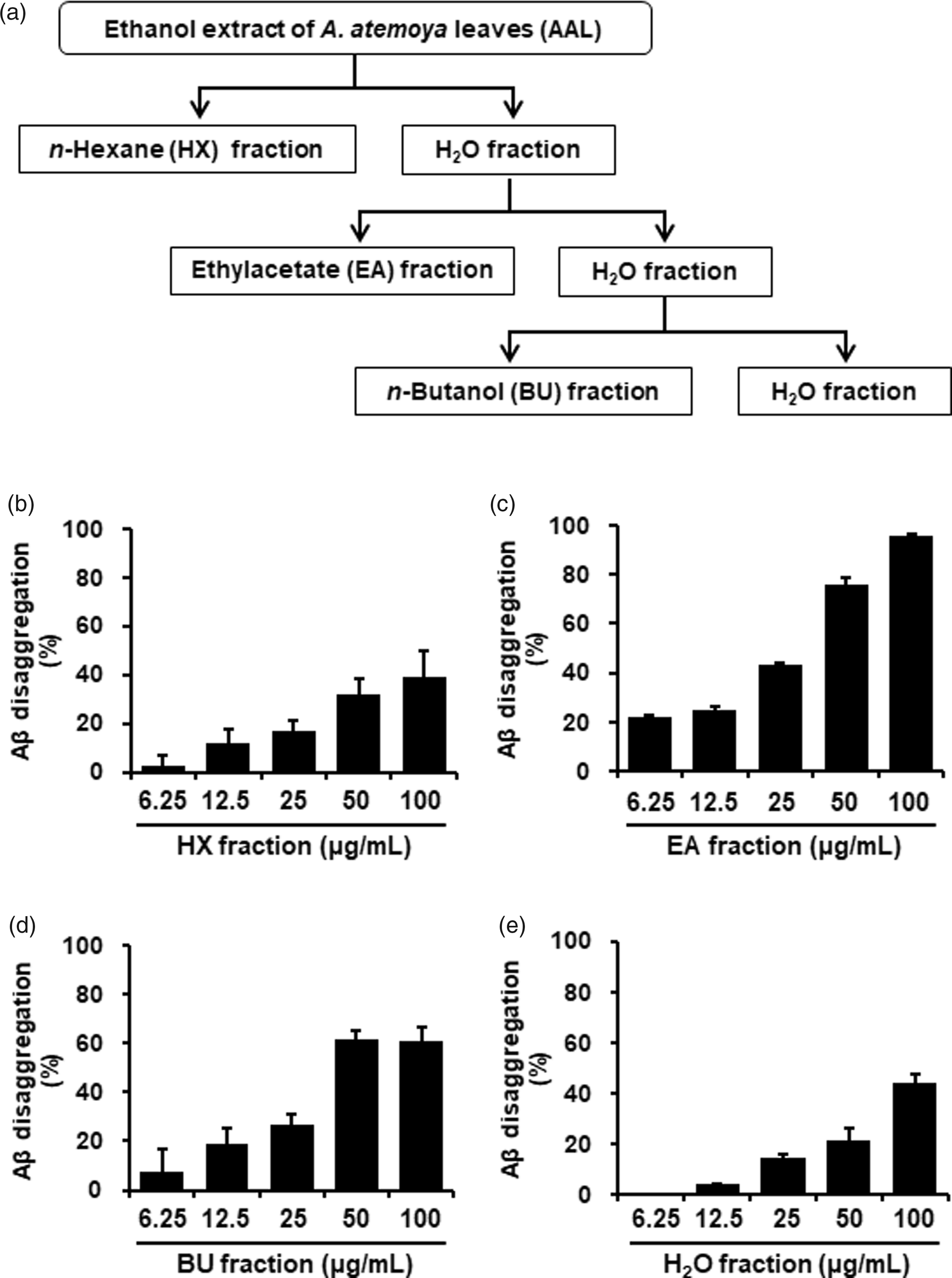
Effect of solvent fractions of AAL extract on Aβ aggregation and neuronal cell damage. (a) Flow diagram of fractionation of AAL extract. (b) Aβ aggregation assay was performed using the SensoLyte® Thioflavin T β-Amyloid aggregation kit. Various concentrations (6.25, 12.5, 25, 50, or 100 μg/mL) of HX (B), EA (c), BU (d), and H2O (e) fractions of AAL extract were reacted with Aβ (1–42) peptides, followed by the addition of Thioflavin T. Fluorescence of Thioflavin T was measured at an excitation wavelength of 440 nm and an emission wavelength of 485 nm. Each value is presented as the mean ± SEM (n = 3). AAL: Annona atemoya leaf.
We tested the neuroprotective effects of each fraction of AAL extracts in H2O2-treated HT22 cells. The cytotoxicity of the fractions was evaluated by CCK assay. Treatment with BU and H2O fractions up to 100 µg/mL had no significant effect on cell viability. In contrast, HX and EA fractions significantly decreased the cell viability in a dose-dependent manner (Figure 4(a)). Subsequent assays were performed using non-toxic concentrations.
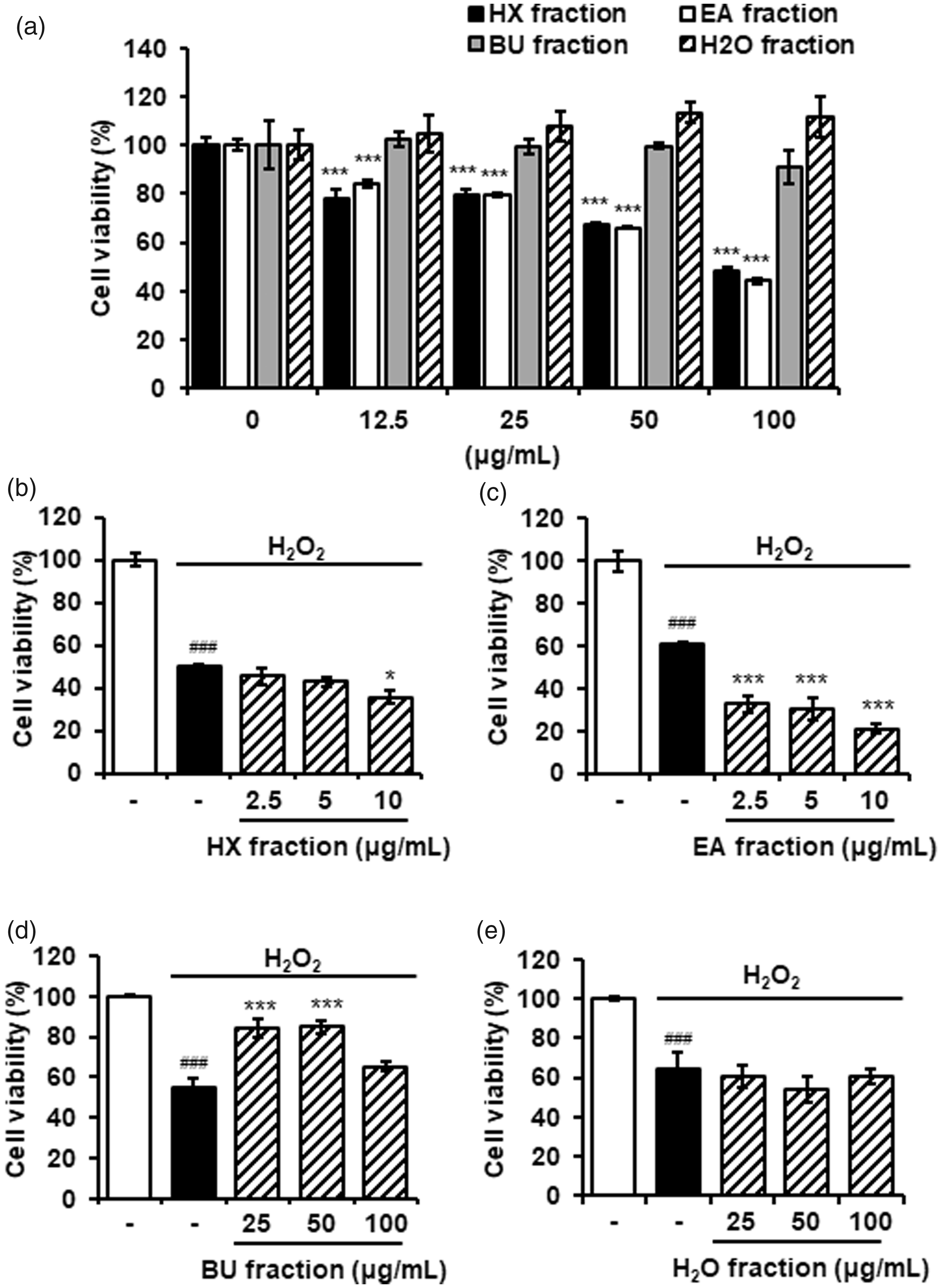
Neuroprotective effects of solvent fractions of AAL extract in H2O2-damaged HT22 cells. (a) Cytotoxicity of n-hexane (HX), ethylacetate (EA), n-butanol (BU), and water (H2O) fractions was measured using the CCK assay against HT22 cells. The cells were treated with the extracts for 24 h. The results are expressed as mean ± SEM of three independent experiments. ***P < 0.001 vs. untreated control cells. (b) to (e) HT22 cells were exposed to H2O2 in the absence or presence of HX (b), EA (c), BU (d), and H2O (e) fractions for 6 h. Cell viability was determined using a CCK assay kit. The results are expressed as mean ± SEM of three independent experiments. ###P < 0.001 vs. vehicle control cells; *P < 0.05 or ***P < 0.001 vs. H2O2-treated cells. AAL: Annona atemoya leaf.
As described above, neuronal cell damage was induced by adding H2O2 to HT22 cells. BU fraction significantly blocked H2O2-induced damage of HT22 cells. All other fractions had no significant effect on cell damage (Figure 4(b) to (e)).
AAL extract ameliorates memory impairment in Aβ-injected AD-like mice model
Memory loss is a main symptom of AD. 24 The effect of AAL extract on memory impairment induced by Aβ was evaluated using the passive avoidance and Y-maze tests (Figure 5(a)). Aβ aggregates were injected icv into male ICR mice to induce an AD-like mouse model. In the passive avoidance task, Aβ aggregates significantly shortened the transfer latency compared to that of vehicle (P < 0.01). Administration of AAL extract at 100 mg/kg significantly increased transfer latency compared to that following Aβ treatment (P < 0.001; Figure 5(b)). In the Y-maze test, AAL extract significantly reversed the spontaneous alternative behavior of Aβ-injected mice at 100 mg/kg (Figure 5(c)). Morin was used as a positive control.
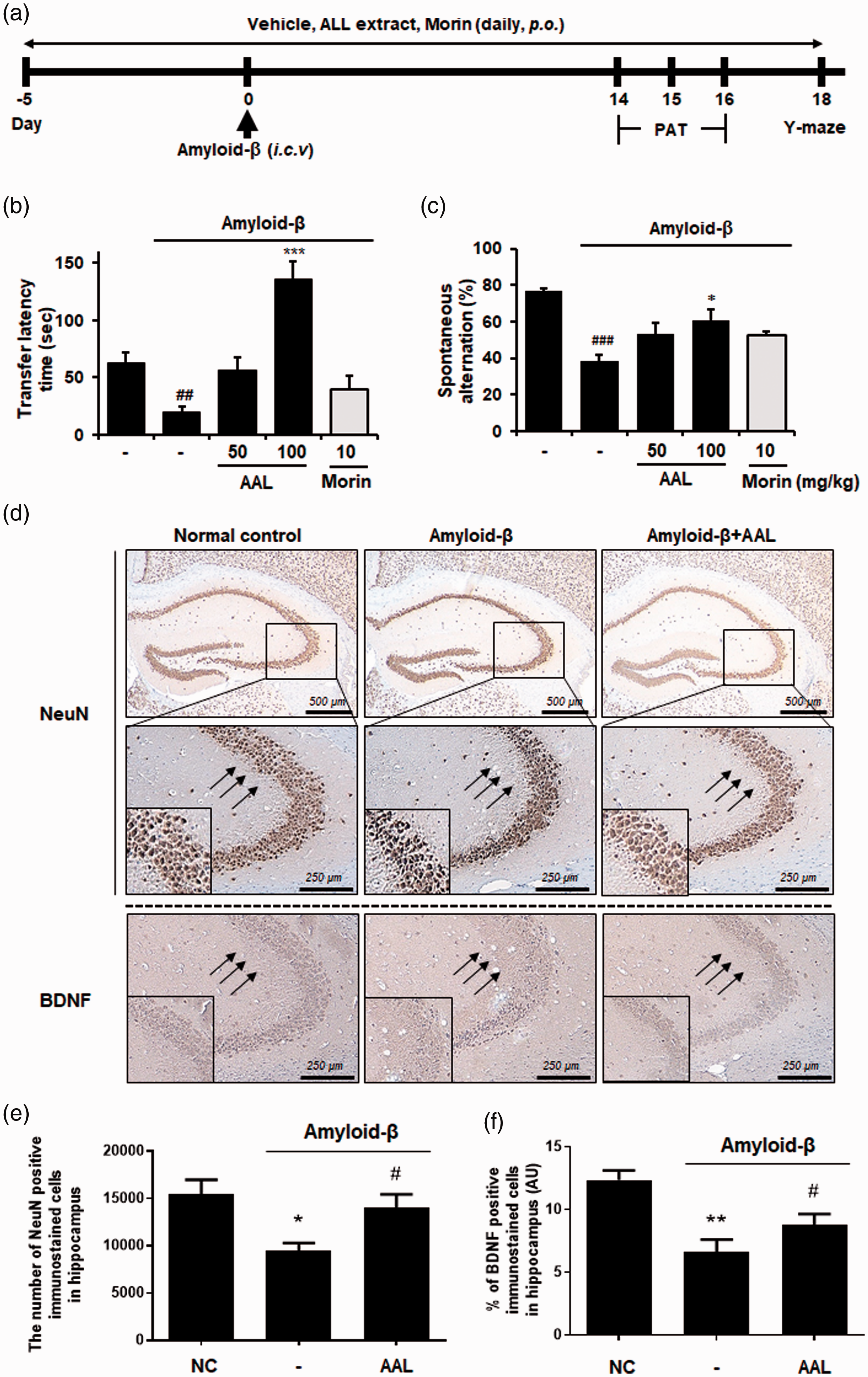
Effects of AAL extract on memory impairment in an Aβ-induced AD-like mouse model. (a) Time schedule of the animal experiments. (b) Male ICR mice were assigned to five groups (n = 7/group). Aβ aggregates (50 pmol per 10% DMSO in PBS) were injected into the intracerebroventricular region. Vehicle (saline), AAL (50 or 100 mg/kg), or morin (10 mg/kg) was orally administered for 23 days after adaptation for one week. (b) For the passive avoidance test, mice were trained on a one-trial step-through passive avoidance task on day 14 after Aβ injection (retention trial). The testing trial was given two days after the training trial (acquisition trial). The latency time was recorded at the retention and acquisition trials. (c) Y-maze was performed on day 18 after Aβ injection. Spontaneous alternation behavior was measured during an 8-min session. The data are presented as means ± SEM. ##P < 0.01 vs. normal group. *P < 0.05, and ***P < 0.001 vs. Aβ control group. Morin was used as a positive control for inhibitor of Aβ aggregation. (d) Expression of NeuN and BDNF in hippocampus was determined by immunohistochemistry (magnification ×200). (e) Quantification of the BDNF positive stained cell density within the hippocampus (proportion of BDNF positive staining over the hippocampus area) was compared between experimental groups (section from n = 3 brains were analyzed per group). (f) The number of neurons in different areas of the brain was estimated by automated cell counting (particle analysis) using the Image J software. NC: normal control, AAL: Annona atemoya leaf. (A color version of this figure is available in the online journal.)
Immunohistochemistry was performed against NeuN and BDNF in hippocampal tissues to elucidate the effects of AAL extract on neuronal marker expression. As shown in Figure 5(d) and (e), the decreased number of NeuN-positive cells was observed in the hippocampus of the Aβ-administered group compared with the normal group. In contrast, AAL administration with Aβ injection considerably reversed the effects of Aβ treatment. Consistently, Aβ injection reduced the immunoreactivity with anti-BDNF antibody whereas AAL administration inhibited Aβ-mediated loss of BDNF expression in Aβ-injected mouse. These data show the neuroprotective effect of AAL in a mouse model.
Molecular mechanisms underlying anti-AD effects of AAL extract
To investigate the regulatory mechanisms responsible for anti-AD effects of AAL, antibody microarray was performed using H2O2-treated HT22 cells with or without AAL extract. Of 1318 phospho-specific proteins, we observed that phosphorylation of four proteins (CaMK2 β/ν/δ, GRK2, EGFR, and Myc) was increased, and that of four proteins (FER, caveolin-1, NFκB 65, and MLRL 2) was decreased by H2O2 treatment at specific tyrosine or serine residues (≥1.5-fold change). Concurrent treatment with AAL and H2O2 reversed phospho-levels of the proteins compared to that of treatment with H2O2 alone (Figure 6(a)). Total protein quantification was used to normalize the phosphoprotein levels. Internal positive controls GAPDH and β-actin were constitutively expressed in all samples.
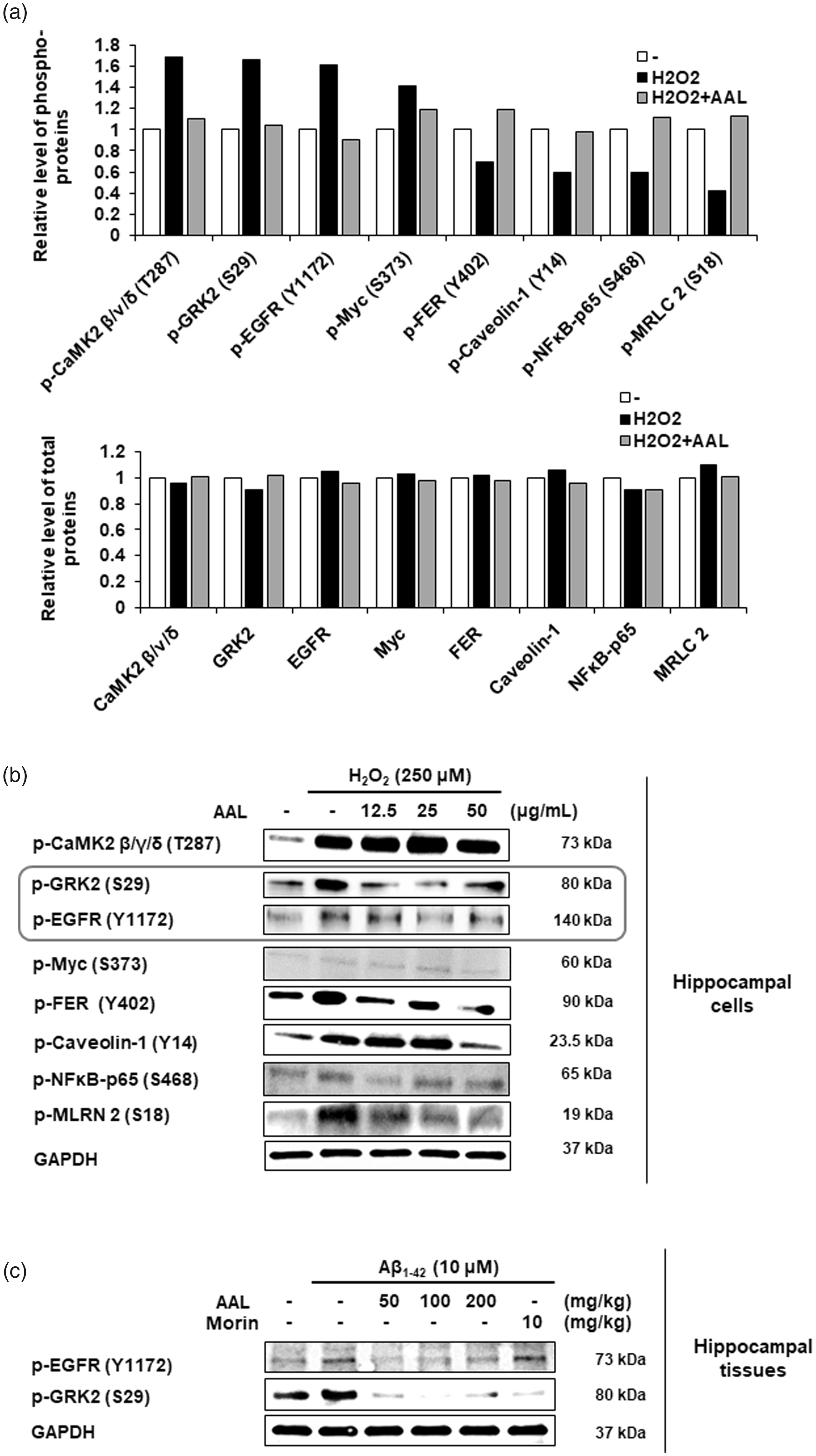
Molecular mechanisms responsible for anti-AD effects of AAL extract. (a) HT22 cells were exposed to H2O2 in the absence or presence of AAL extract (50 μg/mL) for 6 h. Antibody microarray assay was performed using the phospho-specific antibody microarray slide (Full Moon BioSystems). For data acquisition, GenePix 4100A scanner (Axon Instrument, USA) was used. The normalization data were analyzed using Genowiz 4.0™ (Ocimum Biosolutions). The phosphorylation ratio was calculated and represented as fold changes of indicated phosphoproteins after H2O2 treatment normalized to total protein expression (upper panel). Total protein quantification is shown (lower panel). (b) HT22 cells were exposed to H2O2 with or without various concentrations of AAL extract (0, 12.5, 25, or 50 μg/mL) for 6 h. Cell lysates were prepared from HT22 cells and equal amounts of protein were subjected to Western blotting using anti-phospho-CaMK2 β/ν/δ, GRK2, EGFR, Myc, FER, caveolin-1, NFκB p65, and MLRN 2 antibodies to validate the Ab microarray. (c) Protein extracts were prepared from hippocampal tissues in an Aβ-induced AD mouse model. Vehicle or various concentrations of AAL extract (50, 100, or 200 mg/kg) were administered to Aβ mice for 23 days. Western blotting was performed for anti-phospho-EGFR and phospho-GRK2. The validity of the two phospho-antibodies was determined using pre-stained protein marker (Bio-Rad). GAPDH was used as an internal control. Shown blots are representative results from three independent experiments.
Western blotting was performed to validate the antibody microarray results using the anti-phospho-specific antibodies. AAL treatment reduced the phosphorylation of EGFR and GRK2 in H2O2-treated HT22 cells (Figure 6(b)). However, the results of Western blotting for other six phosphoproteins were not consistent with antibody microarray. The effect of AAL on phosphorylation of EGFR and GRK2 was confirmed in an AD-like mouse model. As shown in Figure 6(c), phosphorylation of EGFR and GRK2 was increased in Aβ-stimulated mouse tissue compared with that of normal controls. In contrast, AAL administration reduced the Aβ-mediated phosphorylation of EGFR and GRK2 compared with that of Aβ administration alone.
HPLC determination of the six standard compounds in AAL
Optimized HPLC method was used for the simultaneous determination of the six standard compounds in AAL extract. The results showed well-separated chromatograms using mobile phases consisting of 0.1% (v/v) aqueous TFA (a) and acetonitrile (b). The UV wavelengths were 254 nm for rutin, isoquercitrin, and kaempferol-3-O-rutinoside, 280 nm for (+)-catechin and epicatechin, and 330 nm for chlorogenic acid. The six standard compounds were resolved within 27 min. The retention times for (+)-catechin, chlorogenic acid, epicatechin, rutin, isoquercitrin, and kaempferol-3-O-rutinoside were 11.39, 12.31, 17.8, 24.4, 25.45, and 26.73 min, respectively. HPLC chromatograms of AAL extract and standard compounds are shown in Figure 7.
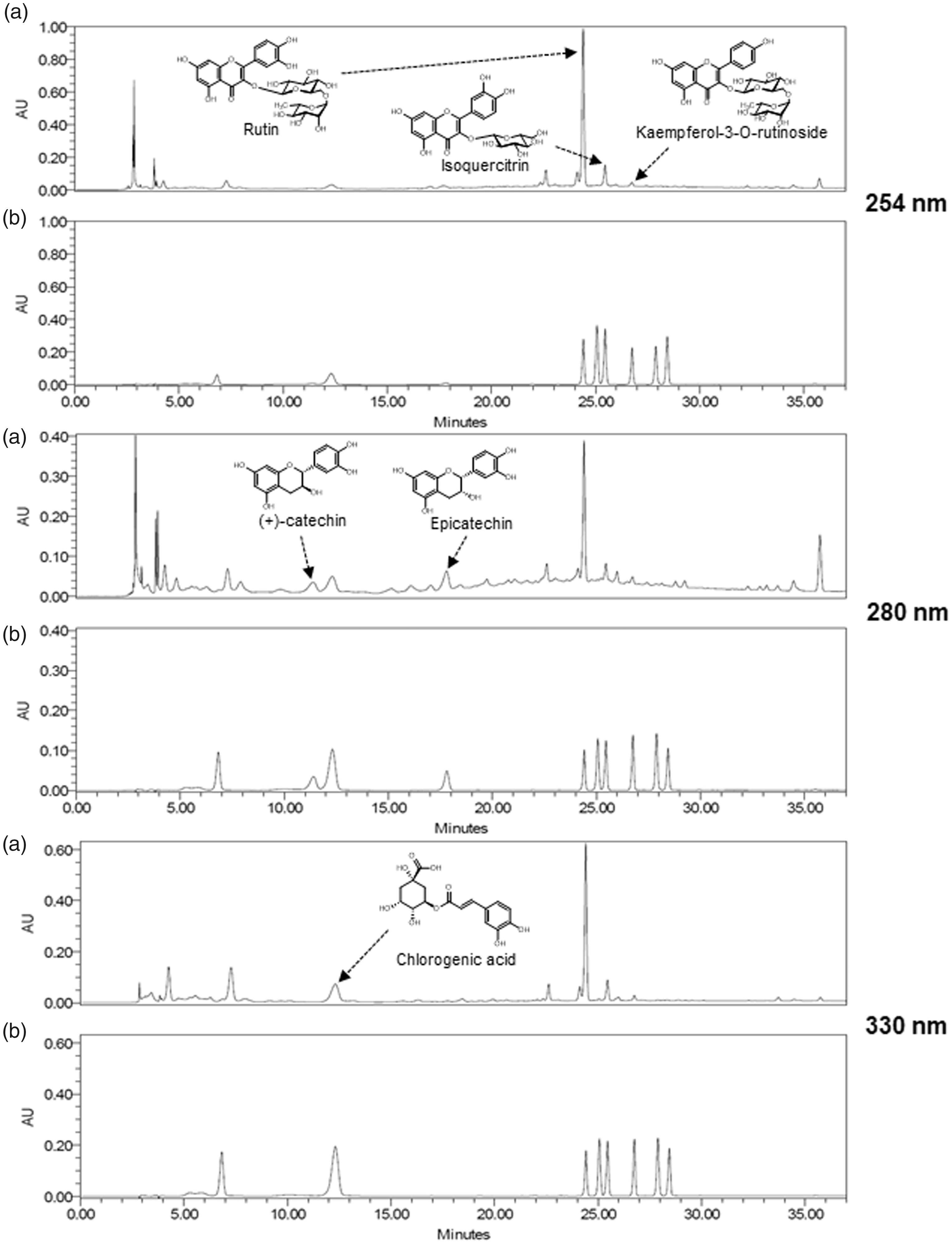
HPLC chromatograms of the ethanol extract of AAL (a) and standard mixture (b) at 254, 280, and 330 nm.
Effect of standard compounds of AAL extract on the Aβ disaggregation
To determine biologically active compound of AAL extract, we conducted the Aβ aggregation assay using the AAL standard compounds. Among six compounds, isoquercitrin (Figure 8(a)) and rutin (Figure 8(b)) increased the ability of Aβ disaggregation in a dose-dependent manner and reached 73.7% and 81.8% at 100 µM, respectively. The maximum ratio of Aβ disaggregation by (+)-catechin (Figure 8(c)), kaempferol-3-ruinoside (Figure 8(d)), epicatechin (Figure 8(e)) was less than 35%. Chlorigenic acid (Figure 8(f)) had no effect on the Aβ disaggregation. Notably, we confirmed rutin and isoquercitrin were contained in BU layer, an active fraction of AAL extract, using HPLC analysis (Figure 8(g)).
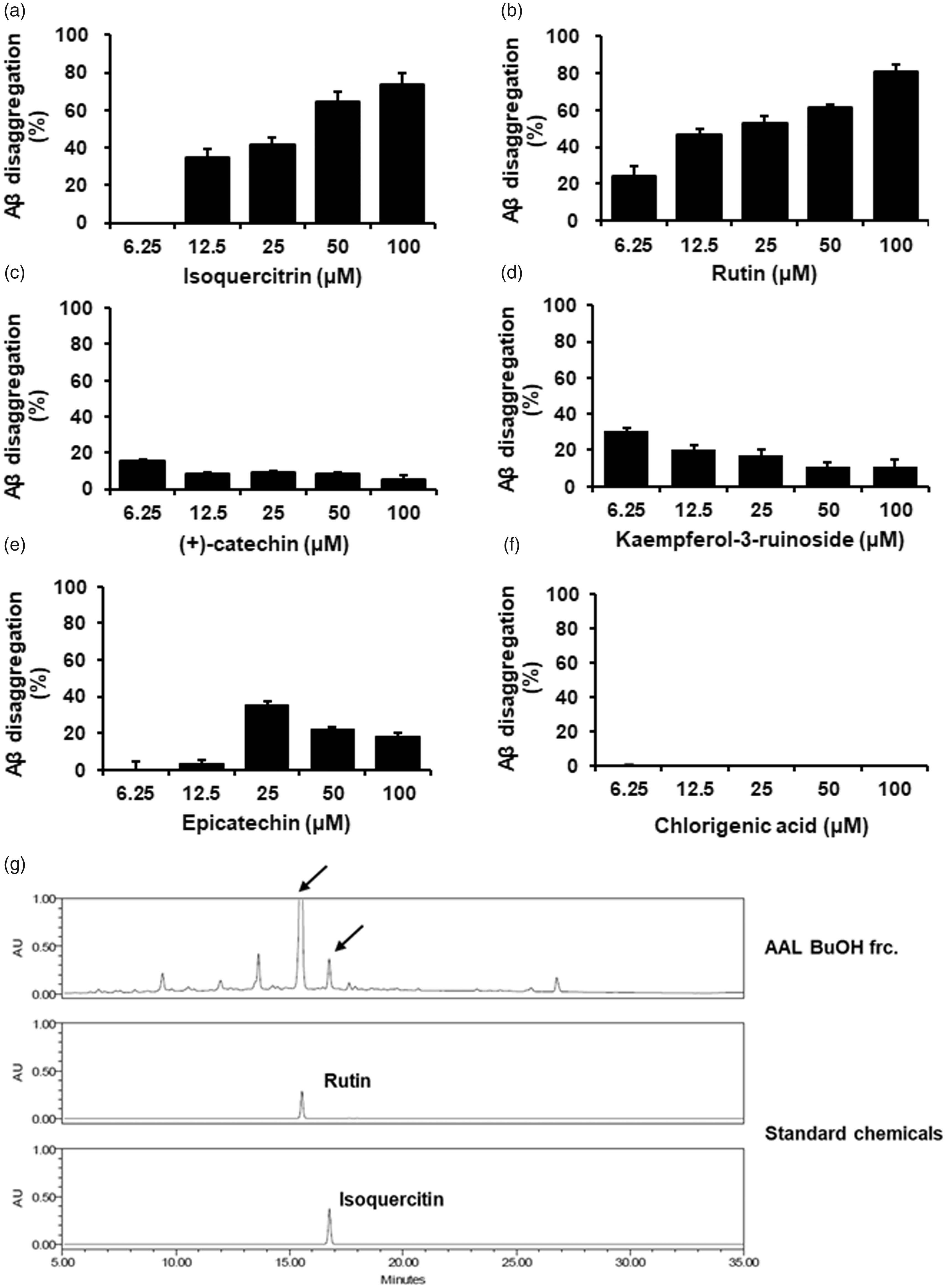
Effect of standard compounds of AAL extract on the Aβ disaggregation. Aβ aggregation assay was performed using the SensoLyte® Thioflavin T β-Amyloid aggregation kit. Various concentrations (6.25, 12.5, 25, 50 or 100 μmol/L) of AAL standard compounds isoquercitrin (a), rutin (b), (+)-catechin (c), kaempferol-3-ruinoside (d), epicatechin (e), and chlorogenic acid (f) were reacted with Aβ (1–42) peptides, followed by the addition of Thioflavin T. Fluorescence of Thioflavin T was measured at an excitation wavelength of 440 nm and an emission wavelength of 485 nm. Each value is presented as the mean ± SEM (n = 3). (g) HPLC chromatogram of BU fraction from AAL extract at 245 nm. Rutin and isoquercitrin were used as standard compounds.
Discussion
Natural products are naturally occurring substances that include phytochemicals and herbal medicines. 25 Accumulating evidences demonstrate the powerful antioxidant activity of many natural products.26–28 An optimal level of reactive oxygen species (ROS) is essential for regulation of cellular processes. However, excessive ROS generation is harmful to biomolecules and induces oxidative stress in various chronic diseases. 29 Therefore, elevating antioxidant activity by activating antioxidant enzymes and free radical scavenging rates may be a beneficial therapeutic approach for chronic diseases. 5 In this regard, natural products are attractive materials for drug discovery and as functional foods. In addition, most natural products have lower toxicity and less side effects compared to those of synthetic drugs.30,31
Our study explored the efficacy of AAL using neuronal cells and an AD-like mouse model. A. atemoya grows and is marketed in subtropical and tropical areas. AAF is generally harvested in fall and winter depending on the region; leaf-dropping occurs from fall to spring.6,7 AAF is eaten raw or utilized for juice, ice cream, desserts, and liquors. 7 Unlike AAF, there are no reports on the usage and biological effects of AAL. Here, we report that AAL extract has the therapeutic potential for AD or AD-related diseases. Our data revealed that AAL extract inhibited Aß aggregation, the hallmark of AD pathology, 16 and enhanced antioxidant activity compared to that of AAF and AAS extracts. In addition, we observed that AAL extract possessed greater potential than that of other medicinal plants such has GB, LC, and AG that have previously been reported to have anti-AD effects.18–20
To date, five drugs were approved by the US Food and Drug Administration for AD. Unfortunately, the drugs do not act as therapeutics for AD. They delay the progression or provide relief of symptoms, but are accompanied by adverse effects such as muscle cramps, bradycardia, and gastrointestinal disorders. The drugs are AChE inhibitors (donepezil, tacrine, galantamine, and rivastigmine) or NMDA receptor antagonists (memantine).1,2 Thus, new drug targets have been sought as drugs to treat AD. Recent studies have explored drugs that inhibit Aß aggregation as a potential therapeutic for AD. Abnormal Aß accumulation in the brains of AD patients is significantly associated with oxidative damage.5,32 The oxidative stress in AD brains is caused by the imbalance between oxidants and antioxidants due to over-production of free radicals. 5 As mentioned above, natural products such as herbal medicines have robust antioxidant capacity. AAL extract dramatically enhanced the scavenging rates against ABTS and DPPH, indicating antioxidant activity of AAL. Excess concentration of Aß acts as a neurotoxin thereby inducing neuronal cell death and oxidative stress. 33 We thus investigated whether AAL extract had neuroprotective effects using HT22 mouse hippocampal cells. We and others have used H2O2-treated HT22 cells in AD studies.34–36 AAL extract, but not AAF or AAS, significantly prevented H2O2-induced neuronal cell death. Our findings suggest that AAL extract has protective effects in neuronal cells damaged by oxidative stress.
Memory loss associated with aging is known as age-related memory impairment. 37 Memory impairment is a main symptom in the pathogenesis of AD and AD-related diseases, that are controlled by different brain mechanisms. 21 Several AD-related animal models involving genetically and/or non-genetically engineered animals have been established, and their pathophysiological characteristics have been identified. Recently, we reported that Aß-injected mice, 5xFAD mice, and initial phase bilateral common carotid artery occlusion rats have similar molecular features compared to those of human AD patients, supporting their validity as models for AD. 38 In the present study, we utilized an Aß-injected AD-like mouse model. AAL extract was orally administered for 23 days, and behavioral tests were conducted. AAL administration displayed a significant preventive effect on memory impairment induced by Aß aggregates in the passive avoidance task and Y-maze test. The effects of AAL at 100 mg/kg/day were greater than those of morin, a positive control for Aβ inhibition. 39
The hippocampus plays an important role in learning and memory by interplaying with the prefrontal cortex. 40 In AD and other dementia patients, the hippocampus is the first brain region to suffer from oxidative stress and related damage. 41 Neuronal markers NeuN and BDNF are altered in the hippocampi of AD-like mice model.42,43 In our immunohistochemical analysis, Aß treatment suppressed the expression of neuronal markers NeuN and BDNF in hippocampal tissues. AAL reversed the effect of Aβ on NeuN and BDNF expression. Taken together, our results provide evidence for anti-AD activity of AAL via inhibition of Aß aggregation, antioxidation, neuronal cell protection, and prevention of memory impairment.
To expand our investigations into the molecular mechanisms underpinning anti-AD effects of AAL, we performed a phosphoprotein antibody microarray consisting of >1000 phospho-antibodies from >30 signaling pathways. In HT22 hippocampal cells, H2O2 stimulation increased the phosphorylation of EGFR and GRK2 among the phosphoproteins tested, while AAL treatment in H2O2-stimulated cells showed similar levels of phospho-EGFR and GRK2 compared with those of untreated control. Antibody microarray results were validated by immunoblotting in vitro and in vivo. Consistently, AAL extract blocked the phosphorylation of EGFR and GRK2 in H2O2-damaged neuronal cells and hippocampal tissues from Aβ-injected mice. EGFR is a transmembrane protein that plays pivotal roles in cell proliferation and differentiation. 44 EGFR signaling in the brain is important in central nervous system development. 45 GRK2, also known as ß-ARK (beta adrenergic receptor kinase), is a serine/threonine intracellular kinase. Activation of EGFR stimulates GRK2 activity. 46 In relation to the study of AD, neuronal EGFR signaling is associated with presenilin 1, the most common cause of familial AD, in the regulation of neuroprotection in AD patients. 47 Several studies reported dysfunction of GRK2 in AD, especially at early stages.48–50 However, the association of EGFR and GRK2 in AD has yet to be identified. Further investigations are required to elucidate the roles of EGFR/GRK2 and their related signaling factors in AD pathogenesis at both protein and gene levels.
We finally determined the biologically active compound of AAL to regulate anti-AD effect. Among the six standard compounds (five flavonoids: (+)-catechin, epicatechin, rutin, isoquercitrin, and kaempferol-3-O-rutinoside, one phenolic acid: chlorogenic acid), isoquercitin and rutin markedly increased inhibitory activity on the Aβ aggregation in a dose-dependent manner. We also identified that both isoquercitrin and rutin were included in BU fraction, an active fraction of AAL extract. Considering biological activity and the content of compound, rutin maybe the active compound of AAL.
Conclusions
Overall, it is of note that we demonstrate the biological activity of AAL for the first time. AAL extract strikingly prevented Aβ aggregation and attenuated memory impairment through inhibition of the EGFR/GRK2 phosphorylation in vitro and in vivo. Rutin was identified as an active compound of AAL to regulate anti-AD activity. Our findings demonstrate that AAL has potential as a therapeutic drug candidate for AD or AD-related disorders as well as functional food for improving cognitive or memory impairment.
Footnotes
Authors’ contributions
H-SL and S-JJ conceived and designed the experiments. H-SL, ES, and S-JJ conduced to the cell experiments. H-SL, YJK, ES, and S-JJ assisted with the animal experiments. H-SL, B-YK, and S-JJ analyzed the data and performed the statistical analysis. YJK conducted AAL extract and analysis using HPLC. H-SL and S-JJ wrote and helped to modify the paper. All authors read and approved the final version of the manuscript.
ACKNOWLEDGMENTS
We thank Drs Chang-Seob Seo and Mee-Young Lee in Korea Institute of Oriental Medicine for kind donation of plant materials.
DECLARATION OF CONFLICTING INTERESTS
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
FUNDING
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by a research grant (KSN1515293) from the Korea Institute of Oriental Medicine (KIOM).