
Abstract
Hypertensive status induces modifications in the respiratory profile. Previous studies have indicated that hypertensive rats show increased respiratory-sympathetic coupling compared to normotensive rats. However, these effects and especially the mechanisms underlying such effects are not well known. Thus, we evaluated the influence of high blood pressure and autonomic dysfunction on a ventilatory pattern associated with lung injury and on the ubiquitin-proteasome system of the diaphragm muscle. Autonomic cardiovascular modulation (systolic BP variance and low-frequency band and pulse interval variance) and arterial blood gases patterns (pH, pO2, HCO3, SpO2), can be changed by hypertension, as well exacerbated chemoreflex pressor response. We observed that the diaphragm muscle of SHR showed increase in type I cross-sectional fiber (16%) and reduction in type II cross-sectional fiber area (41%), increased activity of the ubiquitin-proteasome system and lipid peroxidation, with no differences between groups in the analysis of ubiquitinated proteins and misfolded proteins. Our results showed that hypertension induced functional compensatory/adverse alterations associated with diaphragm fiber type changes and protein degradation as well as changed autonomic control of circulation. In conclusion, we believe there is an adaptation in ventilatory pattern in regarding to prevent the development of fatigue and muscle weakness and improve ventilatory endurance.
Impact statement
It was well known that hypertension can be driven by increased sympathetic activity and has been documented as a central link between autonomic dysfunction and alterations in the respiratory pattern. Our study demonstrated the impact of hypertension in ventilatory mechanics and their relationship with diaphragm muscle protein degradation. These findings may assist us in future alternative treatments to prevent diaphragm fatigue and weakness in hypertensive patients.
Introduction
Arterial hypertension (AH) is a multifactorial disease associated with functional and structural alterations in diverse organs beyond cardiovascular organs, constituting a well-established risk factor for important clinical manifestations. 1 However, the changes induced in non-cardiovascular organs (e.g changes in respiratory pattern and changes in the pulmonary vascular bed, as well as respiratory muscle disorders) remain poorly understood.
The autonomic nervous system (ANS) works to ensure the maintenance of homeostasis. 2 An appropriate ventilatory response is essential to the maintenance of homeostasis, as ventilation constitutes one of the main components modulated by parasympathetic activity. 3 On the other hand, a growing number of studies have demonstrated a central link between autonomic dysfunction and alterations in the respiratory pattern. 4
The autonomic nervous system (ANS), composed of the sympathetic and parasympathetic, works independently or in counter-balance to ensure the maintenance of homeostasis. Some experimental studies have demonstrated reciprocating and mechanistic links between inflammation and autonomic dysfunction.5–8 Increase in circulating pro-inflammatory cytokines increases sympathetic activity, 5 reduces baroreflex sensitivity 9 and heart rate variability (HRV) 2 ; and yet, the activation of cytokines such as TNF-α, IL-6, IL-1b among others contributes to muscle wasting and cachexia. 10
The impairment in different target organs accompanying the time-course development of hypertension can culminate in heart failure. In this sense, oxidative stress and increased protein degradation are usually found in cardiovascular disease-induced tissue damage. 11
Ubiquitin-proteasome system (UPS) is a proteolytic pathway for disposal of damaged proteins, that may be overactivated in skeletal muscle atrophy, as observed in heart failure. 12 However, there are few studies addressing UPS activation associated with oxidative stress and respiratory muscles in hypertension.
In this regard, some functional, morphological, and molecular changes have been observed in diaphragm muscle in humans and in animal models of heart failure and pulmonary hypertension, in which fiber atrophy and contraction strength impairment have been described.13–15 Increase in type I and reduction in type II fibers were described in heart failure and have been associated with the respiratory overload and with the increase of diaphragm fatigue. 16 Although the peripheral phenotypic transition in skeletal muscle is well known in AH, studies addressing changes in the diaphragm structure are rare and less consistent.
In fact, it is well known that respiratory pattern can also influence HR and heart rate variability (HRV), inducing modulation of both sympathetic and parasympathetic output to vessels and other structures. 17 According to Simms et al., 18 SHR have augmented respiratory-sympathetic coupling in comparison with normotensive control in all ages, suggesting that the combination of these increased respiratory-related bursts of sympathetic activity reaching arterial system and the increased BP variability can play an important role in arterial hypertrophy and in other adaptative adjustments of diaphragm and heart muscle to the hypertensive process.
In the present study, changes in BP, HR, chemoreflex sensitivity and cardiovascular variability, as well as ubiquitin-proteasome activity, and oxidative stress were studied to better understand the adjustments of these parameters in spontaneous hypertension that contribute to systemic damage. Therefore, we hypothesized that the imposition of functional overload caused by AH favors some sort of adjustment in the diaphragm.
Methods
Animals and experimental groups
Twenty-four adult male spontaneously hypertensive rats (SHR, 18 weeks old) and adult male Wistar rats (18 weeks old) obtained from the Animal Facility of University of São Paulo (USP) were equally distributed into: hypertensive group (H) and normotensive group (N). Procedures were approved by the Ethical Committee University of Sao Paulo (Protocol 0773/09
Experimental protocol
Femoral arteries and veins were catheterized in order to determine the gasometric values from arterial blood samples, hemodynamic, and autonomic evaluations. In addition, left ventricle (LV) function was assessed by direct catheterization and measurement of lung elastance was conventionally assessed during mechanical ventilation. At the end of the experiment, animals were euthanized and the lungs and diaphragm were removed for analysis. A hemidiaphragm was quickly frozen in liquid nitrogen for fiber cross-sectional area analyses. Another hemidiaphragm was used for molecular evaluations. The lungs were removed and submitted to classical histological routine.
Respiratory rate
The plethysmograph (Buxco Europe, Winchester, UK) was used to assess the ventilation in conscious rats. For this, the animals with spontaneously breathing were placed in the main chamber of the plethysmograph, as previously described. 19
Systemic hemodynamic assessments
The anesthetized rats were implanted with two catheters (Tygon+PE10) into the femoral vein and artery for drug administration and direct measurements of arterial pressure. Rats were observed for at least 24 h after catheter placement; the rats were conscious and allowed to move freely during the experiments.2,20 The recorded data were analyzed on a beat-to-beat basis to quantify changes in mean arterial pressure and heart rate (HR). The parameters for analysis in the time domain consisted of calculating heart rate variability (HRV) by pulse interval variance (PIV) and blood pressure variability (BPV) by systolic blood pressure variance (SBPV) and low frequency band (LFSBP).
Arterial blood gases analysis
After 24 h of catheterization procedure, the blood sample from femoral artery was collected (0.4 mL) with an heparin syringe (Heparin, heparin 5000 UI/mL, Cristália) and the following parameters were measured by using ABL 800 Flex device (Radiometer Copenhagen): potential of hydrogen ion (pH), partial pressure of oxygen (PO2), partial pressure of carbon dioxide (PCO2), bicarbonate concentration in plasma (HCO3), and oxygen saturation (SpO2).
Invasive evaluation of left ventricle function
Three days after hemodynamic evaluations, the evaluation of LV function was done. For this, two catheters were inserted into LV by carotid artery in anesthetized rats. As previously described, 21 the acquisition of LV pressure signals was made using a transducer and the changes in LV pressure were recorded by analysis from beat-to-beat. At the end of the experiment, we were able to obtain; maximum rate of LV pressure rises and fall and LV end diastolic pressure.
Lung elastance measurement
The lung elastance was measured as previously described. 22 The rats were assessed under mechanical ventilation after being anesthetized, tracheostomized, and intubated. The cannula was connected to a mechanic ventilator with 60 breaths/min, 2 mL of tidal volume, and a pneumotachograph and a differential pressure transducer recorded the ventilatory flow. To evaluate the static and dynamic elastances, we used the airway occlusion method and close or open chest wall for total respiratory system and lung elastance. 22
Histology of the lungs – Remodeling of pulmonary arterioles
At the end of experiments, the lungs were fixed with 10% formaldehyde for 24 h. Following this, to perform histologic sections, we embedded the lungs in paraffin. For collagen quantification, we did slices of 5 µm thick and stained with picrosirius red. 23 Histological images were taken with a light microscope with a 40× objective lens and we used the Image-Pro Plus 6.0 for windows software to acquire vascular morphometric.
Histology of diaphragm – Cross-sectional area
We harvested the diaphragm muscle and stored in liquid nitrogen after snap-frozen it in isopentane. We cut the muscle in cross sections of 10 μm and incubated in alkaline solution for assessing the myofibrillar ATPase activity. The muscle type fiber was identified, and the fiber CSA was further analyzed by Image-Pro Plus software. All analyses were performed blindly, by a single observer.
Western blot
To determine the protein concentrations, we used Bradford assays and included the diaphragm samples in SDS-PAGE (10% polyacrylamide gel). We electrotransferred the proteins to a nitrocellulose membrane after electrophoresis. We monitored the samples loading and efficiency of transfer using the Ponceau S staining by 0.5% on the blot membrane. After this, we blocked the blotted membrane at 4°C with 5% low-fat milk in TBS-T for 1 h and subsequently incubated overnight with primary antibody against ubiquitinated proteins (Biomol International, USA). Membranes were then washed and added HRP-conjugated anti-rabbit secondary antibody, and then incubated for 1 h at room temperature. After this, membranes were washed again and incubated for 1 min with ECL. We used the Scion Image software for the quantification analysis of blots. Protein expressions were normalized against Ponceau S staining.
Slot blot analysis of misfolded proteins
The diaphragm tissue was homogenated with 25 mcg of protein and normalized before the slot blotted on a PVDF membrane and then the membranes were washed with T-TBS and blocked with T-TBS + 5% milk. We incubated the membranes with an antibody (anti-soluble oligomer) and the sample loading was normalized by Ponceau staining.
26S proteasome activity assay
The total lysate from diaphragm was assayed for determining the proteasomal chymotrypsin-like activity. We conducted the assay by diluting 50 µg of protein into 200 µL of 10 mM MOPS, pH 7.4 containing 25 µM LLVY-MCA, 2.5 mM ATP, and 5.0 mM Mg2+ in a microtitier plate. Peptidase activities were determined by the release of amino-4-methylicoumarin after the degradation was detected by fluorometer.
Lipid hydroperoxides
We homogenized the diaphragm samples in cold phosphate-buffered saline and centrifuged for 20 min at 4°C. We precipitated the proteins with trichloroacetic acid (10% w/v) and mixed the supernatant with ferrous oxidation-xylenol orange reagent and after incubation of 30 min, the absorbance was read at 560 nm. 24
Statistical analysis
Data are presented as mean ± SEM. We used The Kolmogorov–Smirnov test to determine the normal distribution of variables and the Student t test for comparisons between groups. Significance level was established as *P < 0.05, **P < 0.01, ***P < 0.001.
Results
Hemodynamic and autonomic parameters, arterial blood gases, and respiratory rate
As expected, diastolic and systolic blood pressures were higher in hypertensive (H) rats compared to control rats. Moreover, systolic blood pressure variability and the vascular sympathetic activity were also increased in the H group. Heart rate and cardiac sympatho-vagal balance were similar between groups. LVEDP increased in H rats when compared to N rats. However, we did not observe differences in LV− or +dP/dt (Table 1).
Arterial blood gases, hemodynamic, cardiovascular autonomic modulation and invasive evaluation of left ventricular function in studied groups.
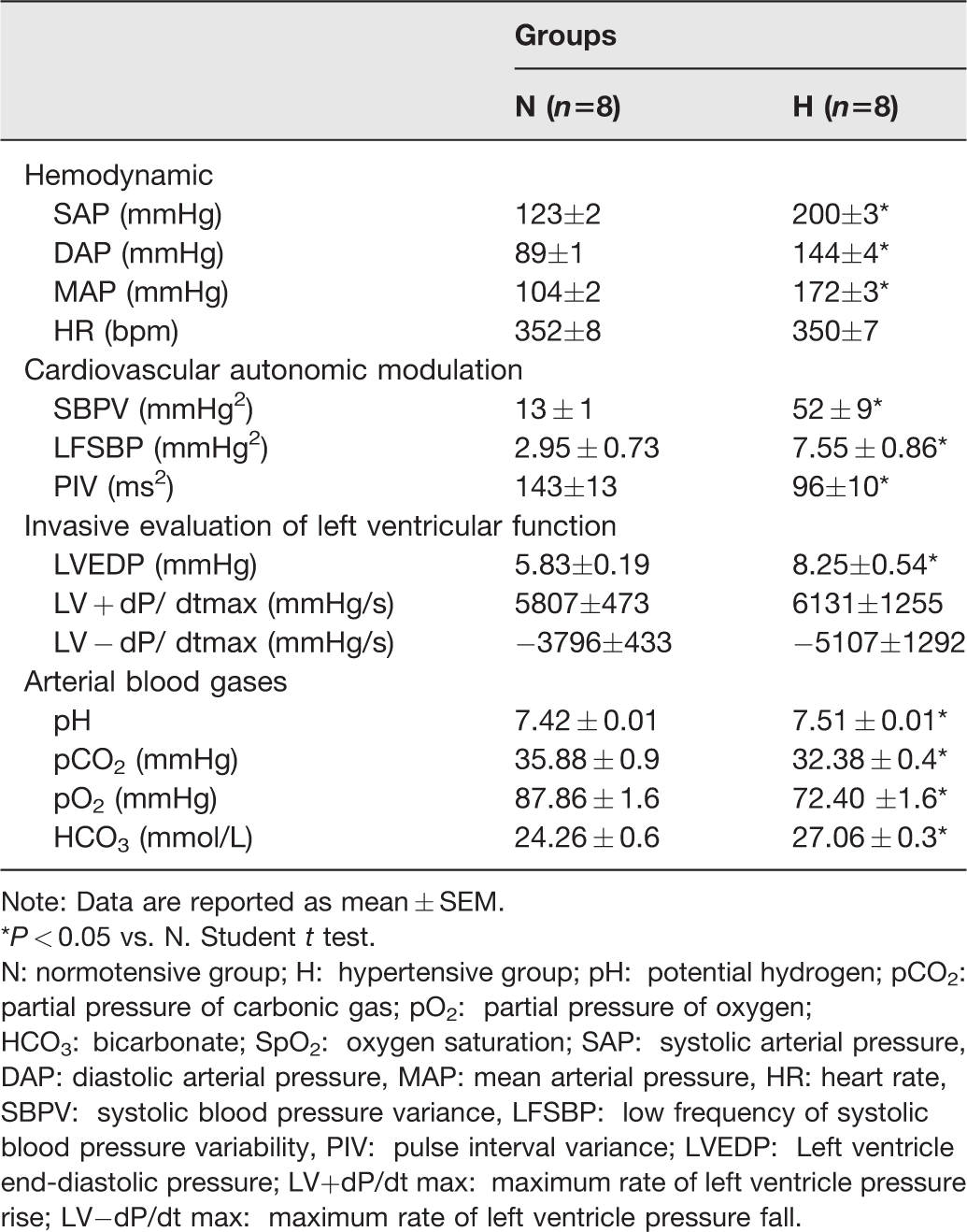
Note: Data are reported as mean ± SEM.
*P < 0.05 vs. N. Student t test.
N: normotensive group; H: hypertensive group; pH: potential hydrogen; pCO2: partial pressure of carbonic gas; pO2: partial pressure of oxygen; HCO3: bicarbonate; SpO2: oxygen saturation; SAP: systolic arterial pressure, DAP: diastolic arterial pressure, MAP: mean arterial pressure, HR: heart rate, SBPV: systolic blood pressure variance, LFSBP: low frequency of systolic blood pressure variability, PIV: pulse interval variance; LVEDP: Left ventricle end-diastolic pressure; LV+dP/dt max: maximum rate of left ventricle pressure rise; LV−dP/dt max: maximum rate of left ventricle pressure fall.
There was a significant change in the arterial gases profile in H animals, characterized by an increment in pH and HCO3, and a reduction in pCO2, pO2 levels and SpO2 as compared to control rats. These alterations were associated with a high respiratory frequency in H as compared to normotensive rats (H: 158 ± 4 vs. N: 119 ± 6 bpm) (Figure 1).
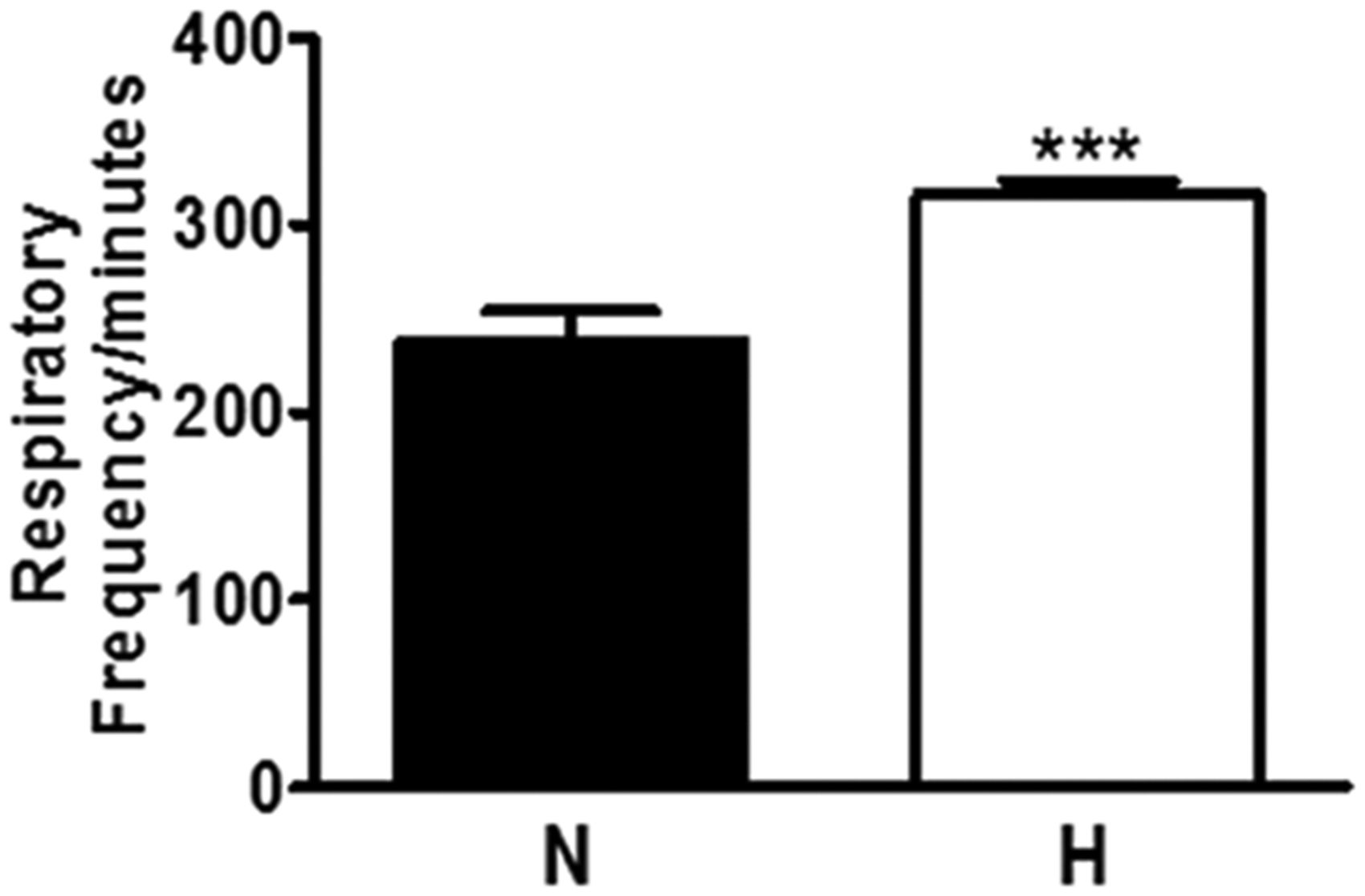
Baseline characteristics of respiratory frequency in normotensive and hypertensive group. Data are reported as mean ± SEM. N: normotensive group; H: hypertensive group.***P < 0.001 vs. N. Student t test.
Pulmonary morphometric evaluations
Hypertensive animals at 18 months of age had a lower body weight compared to their age-matched controls (N) (Figure 2(a)). However, the lung-to-body weight ratio of H was higher compared to normotensive rats (Figure 2(b)). Lung wet-to-dry weight ratio was similar in both groups (H: 79 ± 0.4 vs. N: 79 ± 0.3).
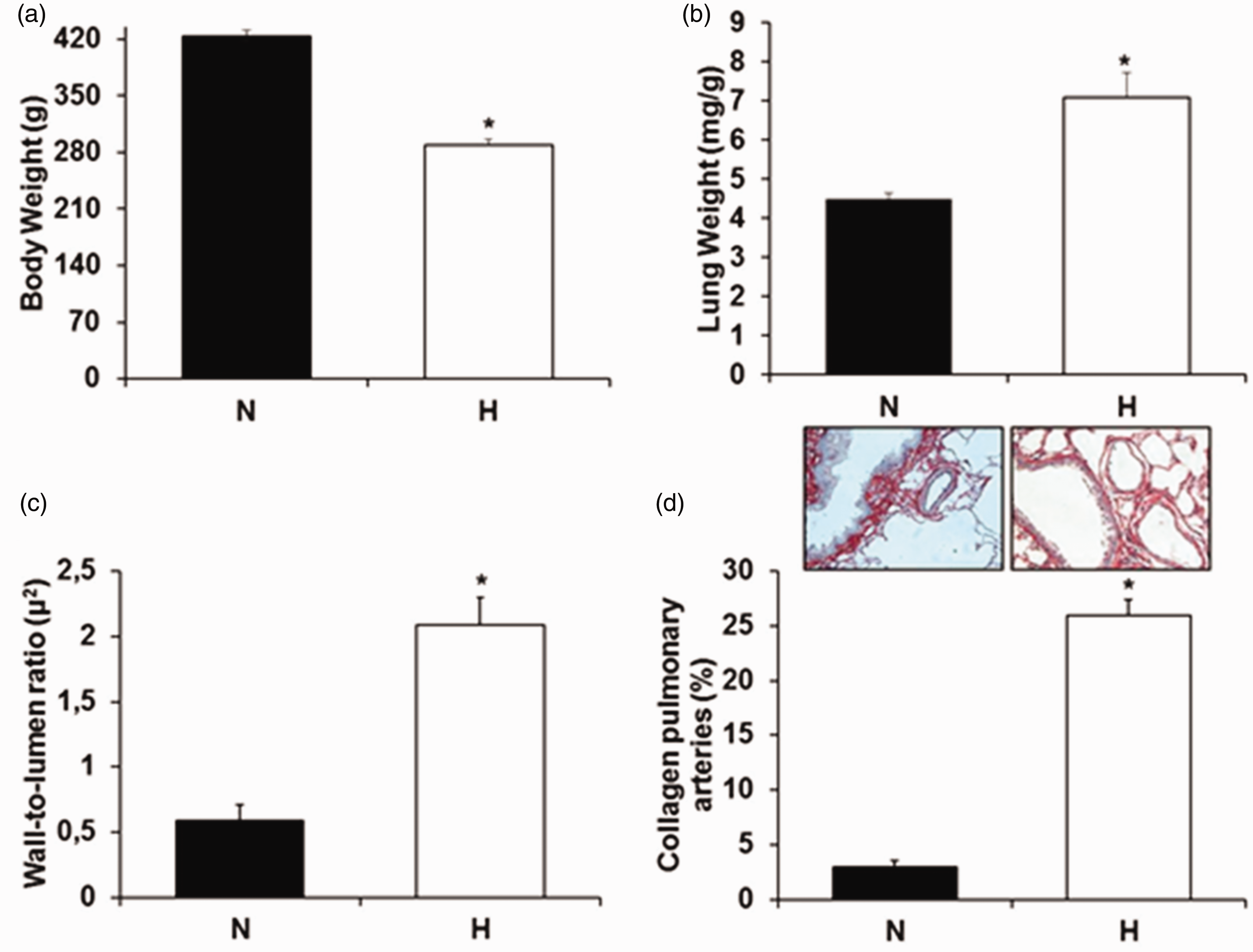
(a) Body Weight and (b) Lung Weight among the animals. (c) Wall-to-lumen ratio of the pulmonary artery and (d) percentage of collagen in the pulmonary artery of the intra-acinar groups normotensive and hypertensive. Data are reported as mean ± SEM. White column: normotensive group; Black column: hypertensive group. *P < 0.05 vs. N. Student t test. (A color version of this figure is available in the online journal.)
Hypertensive group showed a higher wall-to-lumen ratio compared to control group (H: 2.09 ± 2.21 vs. N: 0.59 ± 0.12 μ2) (Figure 2(c)). In addition, we observed a significant increment in hypertensive animals in collagen deposition in the pulmonary arteries when compared to normotensive animals (H 26 ± 1.41 vs. N: 2.6 ± 0.5%) (Figure 2(d)).
Lung elastance
The total respiratory system of H rats presented significantly higher static elastance (Est) and dynamic elastance (Edny) values compared to control animals (H: 5.07 ± 0.06;N:3.75 ± 0.11 and H:6.33 ± 0.18; N 4.65 ± 0.11 cmH2O·mL−1, respectively).
Lung elastance provided after opening the mice chest wall was higher in H rats when compared to control for both Est and Edny (H: 4.12 ± 0.16; N: 3.02 ± 0.08 and H: 5 ± 0.16; N: 3.84 ± 0.14 cmH2O·mL−1, respectively). The chest wall, which is calculated by the difference between respiratory system and lung, was the same for both groups (Figure 3).
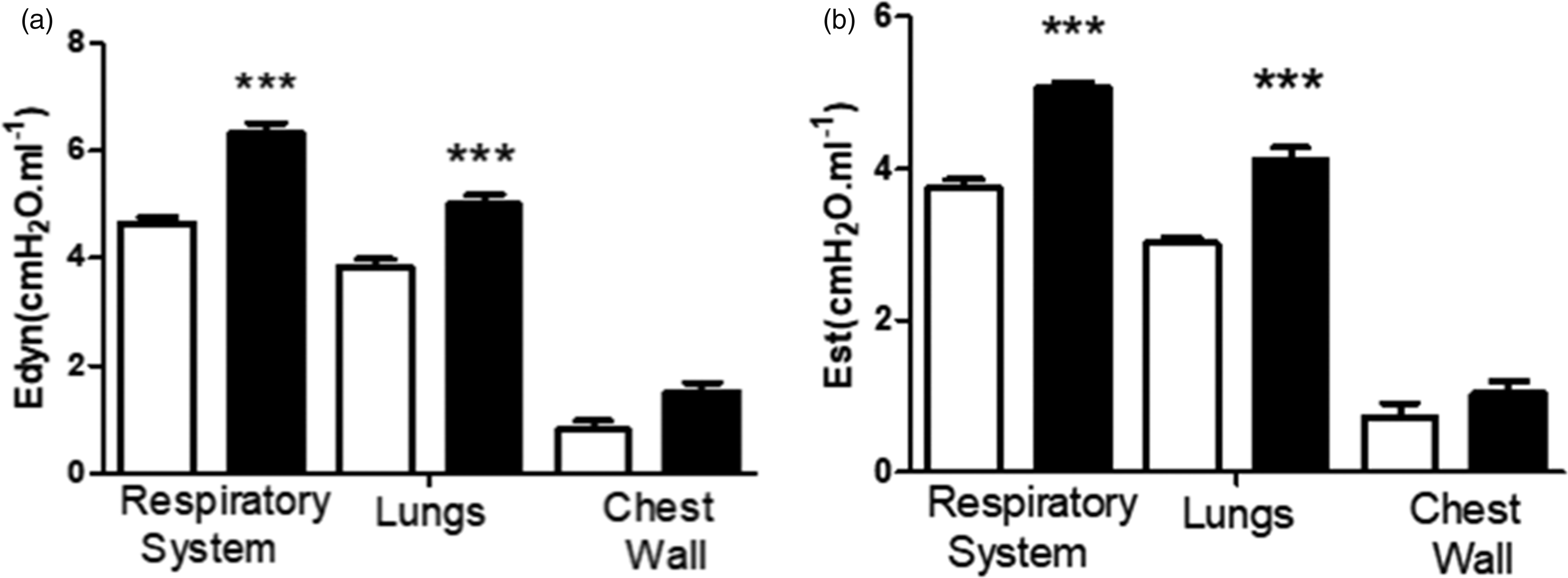
(a) Dynamic (Edyn) and (b) Static (Est) elastances in White column: normotensive group; Black column: hypertensive group determined by the end-inspiratory airway occlusion method. ***P < 0.05 when comparing NT and H.
Diaphragm evaluations
In H group, the cross-sectional area from type I fiber was higher in relation to normotensive rats. The IIa fibers were not different between the groups. However, the diameter of type IIb fiber was significantly reduced in hypertensive group (Figure 4).
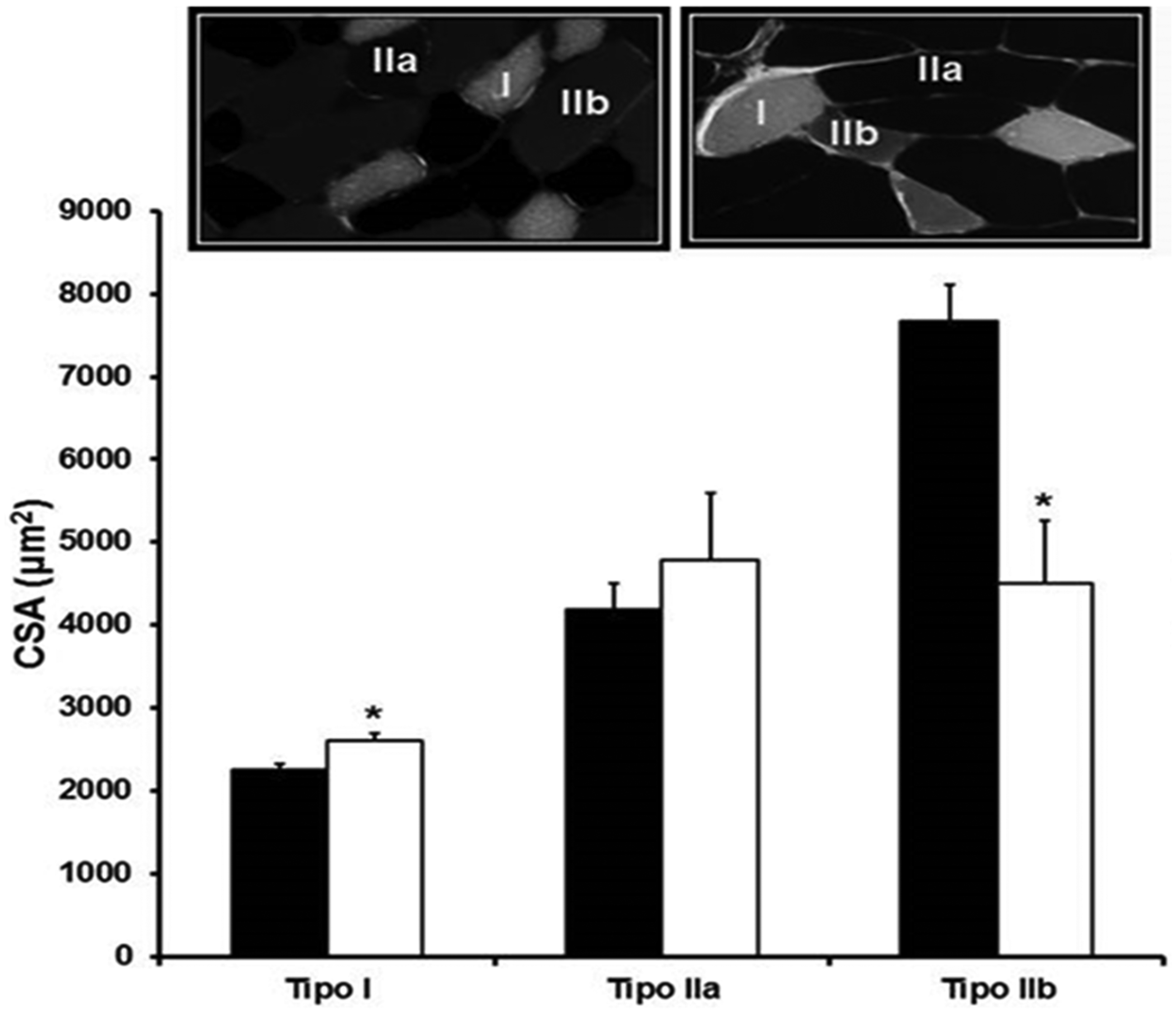
Representative ATPase staining at pH 10.3 in diaphragm muscle (20×) from normotensive (top left) and hypertensive group (top right). Type I (white), IIa (Black), and IIb (gray) fibers are indicated. Cross-sectional areas (CSA) of different types of diaphragm muscle fibers of groups of normotensive and hypertensive groups. Data are reported as mean ± SEM. Number by groups 5–6. N: normotensive group; H: hypertensive group.*P < 0.05 vs. N. Student t test.
26S proteasome activity assay
We measured the chymotrypsin-like proteasome activity and observed two-fold increase in H (Figure 5(a)).We assessed the lipid peroxidation levels as marker of oxidative stress and nearly two-fold changes were observed in the lipid peroxides levels in H (Figure 5(b)), and these increases in proteasome activity and lipid peroxidation were not accompanied by the accumulation of ubiquitinated and misfolded proteins (Figure 5(c) and (d), respectively)
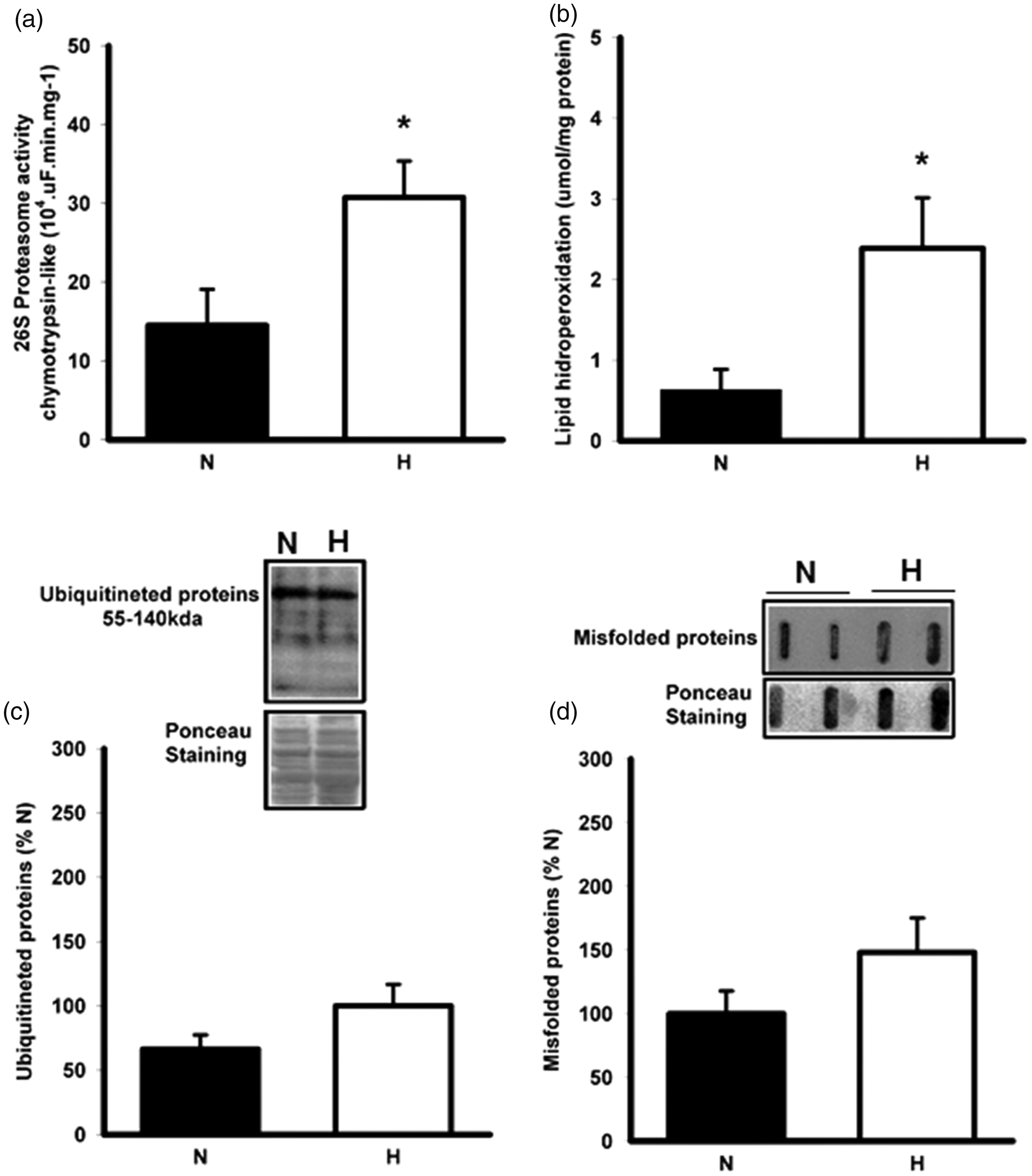
26S Proteasome activity (a), lipid hydroperoxidation (b), ubiquitinated proteins (c), misfolded proteins (d) and correlation between lipid hidroperoxidation vs. proteasome activity in normotensive and hypertensive groups. Data are reported as mean ± SEM. Number by groups 7–8. N: normotensive group; H: hypertensive group.*P < 0.05 vs. N. Student t test.
Discussion
This is the remarkable study showing the impact of hypertension in ventilatory mechanic and their relations with diaphragm muscle protein degradation. Regarding these, we demonstrated that, even in early stages of arterial hypertension, the ventilatory pattern alters in the fibers type I of diaphragm cross-sectional area and atrophy in type IIb fibers. Parallel to this, there is an increased ubiquitin-proteasome activity associated with increased lipid peroxidation in the diaphragm muscle, but not in the accumulation of ubiquitinated and misfolded proteins.
We used the experimental model of hypertension (spontaneously hypertensive rats, SHR) developed by Okamoto and Aoki, 25 since this model mimics the human arterial hypertension impairment, such as autonomic, hemodynamic, vascular, and target organ damage.
In our study, the analysis of arterial blood gases showed that SHR presents the trend of change in breathing patterns with hyperventilation. Likewise, human studies have shown that the development of hypertension and aggravation of the disease is related to the modification of the pattern and breathing rhythm, with and without development of obstructive sleep apnea. 26 This change in breathing pattern, tending to hyperventilation interspersed with hypopnea periods modifies the muscle demand, which may be related to the development of abnormal morphology of the diaphragm, as described in heart failure. Here we demonstrated for the first time the relationship between hypertension and diaphragm adaptation. In addition, changes in breathing pattern in hypertension present a relationship with cardiovascular components changes autonomic control (chemoreflex sensitivity and sympathetic modulation of BP) and cardiopulmonary interaction. Indeed, these modifications contributed to the increase and the maintenance of BP levels, as well as to an increased risk of target organ damage, as related in chronically hypertensive individuals.
In the present study, SHR showed decreased PO2 and increased PCO2, suggesting a hyperventilatory pattern. This behavior may be associated with diaphragm morphofunctional adjustments. In this sense, we investigated the occurrence of diaphragmatic muscle adaptation in hypertension. Studies have shown that some dyspnea disorder with cardiovascular diseases is associated with diaphragm myopathy. 27 Since this muscle is the most relevant in inspiration process in mammals, perhaps it could be also the most damaged due to its role in the improvement of ventilatory endurance. Diaphragm mass muscle loss has been demonstrated under experimental conditions such as mechanical ventilation 28 and monocrotaline-induced pulmonary hypertension. 29 However, in these models mentioned before, atrophy occurs in both oxidative and glycolytic fibers in opposition to what we observed in our study.
The finding of reduced lung compliance in SHR rats is suggested to have a major clinical impact on hypertension. This reduced lung compliance in SHR rats, demonstrated by higher elastance (Est and Edny), probably inducing an increase in respiratory rate which affected the arterial blood gases (pCO2 and SpO2). This hypocapnia indicates higher demands on the inspiratory muscles, here demonstrated by the diaphragm. The diaphragm overactivity induced a fiber type transformation shown in Figure 4 (fiber type IIb in fiber type I) leads to reduced inspiratory muscle strength. 30 The increase of collagen in pulmonary arteries (Figure 2) may be an important factor of this higher lung elastance in this model of hypertension. The loss of the normal distensibility of these arteries reduces pulmonary vasculature compliance consequently decreasing the lung expansion.
We speculate that the largest cross-sectional area in slow-twitch oxidative fibers (type I) in detriment to reduction in glycolytic-fibers (IIB) in SHR (Figure 2) highlights a possible adaptation to changes in ventilatory pattern, probably as an attempt to improve the ventilator endurance and prevent the development of fatigue and muscle weakness which contrasts with changes observed in appendicular skeletal muscle of SHR.31,32 This suggests that the diaphragm muscle weakness in hypertension results from a local insult and is not part of a generalized process of muscle weakness and atrophy.
It is known that muscle trophicity is dependent on the balance between the rate of protein synthesis and proteolysis and UPS.UPS is a major proteolytic pathway responsible for the disposal of damaged proteins and protein turnover in muscle. 33 Diaphragmatic myopathy occurs in parallel to increased proteasome activity.33,34 Our results indicate that oxidative stress may be related to activation of UPS process since we observed in SHR a redox imbalance which can modulate UPS directly for oxidizing proteins which are substrates for the UPS.35–37 Moreover, in the present study, we observed a positive correlation between chymotrypsin-like proteasome activity and lipid peroxidation levels (r = 0.6, P < 0.05).
In fact, oxidative stress has been implicated in the diaphragmatic atrophy and contractile dysfunction in rats.37,38 We observed no increase in ubiquitinated and misfolded proteins, despite the UPS increase activity and lipid peroxidation level in diaphragm. Therefore, our results showed that, in early stages, hypertension may carry a hyperactivation of the UPS in the diaphragm muscle, in order to minimize damage that could be caused by the accumulation of damaged proteins. We do not discard the possibility that in more advanced stages, the extent of oxidative damage may yet be larger.
Conclusions
We suggest in this study that hypertension induces modifications in the respiratory profile associated with diaphragm fiber type changes (increase in the cross-sectional area of slow-fibers (type I) and reduced the cross-sectional area of type II fibers) and increase of UPS activity and oxidative stress. In conclusion, we believe there is an adaptation in ventilatory pattern in regarding to prevent the development of fatigue and muscle weakness and improve ventilatory endurance.
Footnotes
Authors’ contributions
PRMS and FMC planned and conceived the whole project. PRMS and RKP wrote the manuscript. FS, WMAM, AM, MKK and FMA conducted experiments. PRMS,KDA and RPK analyzed and interpreted the data. RKP, MCI and FMC revise the manuscript. All authors checked and approved the submission.
DECLARATION OF CONFLICTING INTERESTS
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
FUNDING
This study was supported by a fellowship from the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), FAPESP, CNPq and Fundação Zerbini.