
Abstract
Alternative splicing of RNA is an essential mechanism that increases proteomic diversity in eukaryotic cells. Aberrant alternative splicing is often associated with various human diseases, including cancer. We conducted whole-transcriptome analysis of 18 osteosarcoma bone samples (paired normal—tumor biopsies). Using RNA-seq, we identified statistically significant (FDR <0.05) 26 differentially expressed transcript variants of leptin receptor overlapping transcript (LEPROT) gene. Some of the transcripts were overexpressed in normal cells, whereas others were overexpressed in tumor cells. The function of LEPROT is not completely understood. Herein, we highlight a possible association between OS and aberrant alternative splicing events and its interaction with the expression of LEPROT. We also discuss the role of LEPROT in regulating growth hormone and its receptor, and the relationship with initiation and progression of OS. This research study may help to understand the association of alternative splicing mechanism in OS and in tumorigenesis more generally. Further, LEPROT gene can also be considered as a potential biomarker of osteosarcoma.
Impact statement
Osteosarcoma (OS, also known as osteogenic sarcoma) is the most common primary malignancy of bone in children and adolescents. The molecular mechanisms of OS are extremely complicated and its molecular mediators remain to be elucidated. We sequenced total RNA from 18 OS bone samples (paired normal—tumor biopsies). We found statistically significant (FDR <0.05) 26 differentially expressed transcript variants of LEPROT gene with different expressions in normal and tumor samples. These findings contribute to the understanding of molecular mechanisms of OS development and provide encouragement to pursue further research.
Introduction
Osteosarcoma (OS) is the most common primary tumor of bone in children and adolescents, with a second peak in incidence in people over the age of 50.1,2 OS is characterized by the presence of malignant mesenchymal stem cells (MSCs) producing immature bone matrix or osteoid. 3 OS commonly develops in long bones such as the distal femur, proximal tibia, or proximal humerus. 4 Patients usually present with pain and swelling. The diagnosis is confirmed by histology and staging studies such as MRI and systemic radionuclide scans.5–7 Current treatment of OS, a combination of surgery and chemotherapy (doxorubicin, methotrexate, and cisplatin), has improved outcomes significantly, although this regime is still only successful in 90% of patients. 8 The precise underlying mechanism of OS remains obscure in most individuals; however, the risk of developing OS is influenced by bone turnover, age, environment (drug or radiation therapy), other bone diseases (such as Paget’s disease), genetic alterations and hormonal regulation of puberty (such as growth hormone).9–13
RNA sequencing (RNA-Seq) technology is a powerful tool to analyze the transcriptome of a cell. 14 Gene expression studies have been traditionally carried out by northern blot and quantitative polymerase chain reaction (qPCR), which are limited to single transcript expressions.15,16 The RNA-seq technique provides higher resolution of the dynamic nature of the transcriptome.15,17 Beyond measuring gene expression, this technique facilitates the discovery of novel transcripts, identification of alternative (differential) splicing (AS) events in genes, and detection of allele-specific expression on a genome-wide scale.15–18 Consequently, RNA-seq is a useful method for interpreting the functional elements of the genome and also understanding the underlying mechanisms of complex diseases in a species.19,20 AS is a crucial mechanism that generates proteomic diversity in eukaryotes. 19 It is mainly regulated by cis-acting elements and trans-acting factors. 21 Growing evidence suggests that more than 90% of multi-exon genes undergo AS in humans.22,23 Deregulation of AS programming can produce a variety of transcript isoforms with unique protein-coding and possibly different or antagonistic biological functions from the same gene.24,25 Predictably, abnormal splicing can disturb normal cellular physiology and eventually lead to diseases, including cancer.23–26 The involvement of AS in each of the widely recognized and accepted hallmarks of cancer has been investigated. 27 In particular, apoptosis and metastasis are directly affected by AS. 28 Moreover, it can also play an essential role in invasiveness, angiogenesis, and chemo/radio-resistance to therapy.28,29
Little is known about the underlying molecular mechanism of AS in various cancer types, especially in OS. To improve our understanding of the genetic mechanisms involved in developing OS, we performed RNA-Seq transcriptome analysis from 18 normal—tumor pairs of bone biopsies by computational bioinformatics. Differential exon usage was studied to highlight AS and transcription events present between normal and tumor samples using the DEXSeq package. Another aim of this study was to identify potential molecular biomarkers and novel therapeutic target candidates for the early detection and treatment of OS.
Materials and methods
Samples collection
The study was investigated and approved by the Ethics Review Committee on Biomedical Research of Hue University of Medicine and Pharmacy. The participants and patient representatives signed and dated the informed consent forms before surgery.
Normal and cancerous bone samples were collected directly after surgery (limb sparing or amputation) from 18 Vietnamese patients who, previously, had histological confirmation of OS. The collected biopsies were replaced on dry ice and stored at −80°C until processed for RNA extraction.
Total RNA isolation from bone biopsies
Approximately 50 mg of bone sample was ground with liquid nitrogen in a pestle and mortar and pre-TRIzol treatment was performed (Invitrogen Corp., Carlsbad, CA, USA). RNeasy Fibrous Tissue Mini Kit (Qiagen Inc., Valencia CA, USA) was used to isolate total RNA from bone tissue following the manufacturer’s protocol. Isolated RNA was completely dissolved in RNase-free water and stored at −80°C. The quality of total RNA was determined by Agilent 2100 Bioanalyzer system and the RNA 6000 Nano Kit (Agilent Technologies Inc., CA, USA).
Fifty nanograms of total RNA was amplified using Ovation RNA-Seq System V2 (NuGen, Emeryville, CA, USA) and the resulting cDNAs were pooled in equal amounts. The pooled cDNA was used to prepare the DNA fragment library using SOLiD System chemistry (Life Technologies Corp, Carlsbad, CA, USA). Sequencing was done by SOLiD 5500 W platform and DNA sequencing chemistry (Life Technologies Corp., Carlsbad, CA, USA). Raw data (75 bp) were color-space mapped to the human genome hg38 (GRCH38) reference through the mapping algorithm implemented in the Lifescope™ Genomic Analysis software (Life Technologies Corp.). The mapping confidence had been noted as higher than 90 due to the quality threshold was set to 10. The reads scores below than 10 were filtered out. Mapping quality was at average of 30. Annotation of the reads based on the gencode v22 files. 30
Statistical data analyses were performed through the DEXseq package for R. 31 DEXseq is a Bioconductor package specifically designed to find differential exon usage (DEU) based on RNA-seq data. DEU describes the relative usage of exons induced by the experimental conditions. 32
More details on data pre-processing can be found here (https://bioconductor.org/packages/release/bioc/vignettes/DEXSeq/inst/doc/DEXSeq.html). The DEXseq package also performs sample comparison and adjusts the P-value to overcome multiple testing problems.33–35 Through DEXseq package, the Benjamini–Hochberg procedure used to control the false discovery rate (FDR). 36
Results
Patient characteristics
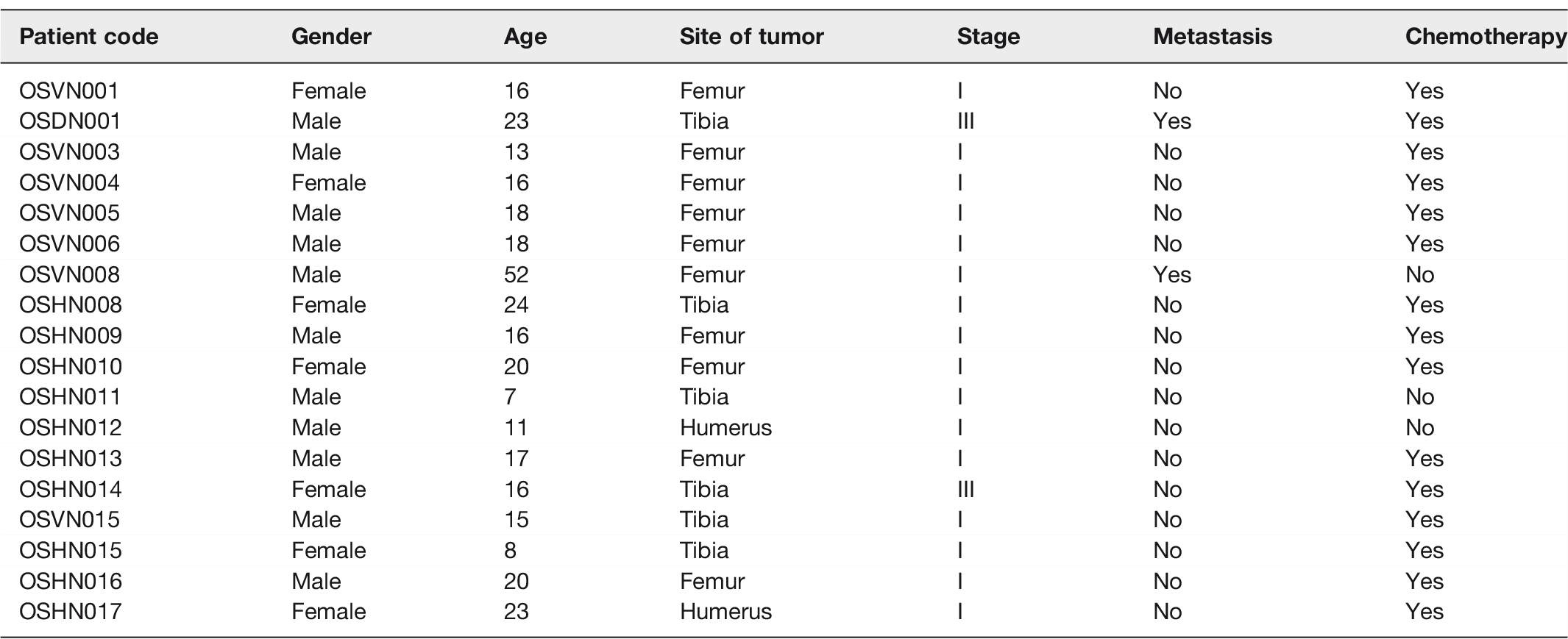
Paired samples (normal—tumor) of surgical biopsies of bone were collected from 18 Vietnamese OS patients. Diagnosis was confirmed by a pathologist by examination of the biopsies. Tumor sites were located in the femur (55.56%), tibia (33.33%), and humerus (11.11%). Patient age ranged from 7 to 52 years (mean = 18 years) and there were 7 females and 11 males. Patients’ demographic and clinical characteristics are listed in Table 1.
Characteristics of osteosarcoma patients in the present study (n= 18).
Validation of differential exon usage of LEPROT in patient samples
A mean-average (MA)-plot of the log2 fold change of all transcripts (over 4075 transcripts) expressed in normal and tumor samples is presented in Figure 1. Red points indicate the log fold change versus average normalized count per exon (FDR < 0.05), whereas red triangles represent exons expressed with log2-fold change greater than 2 or less than −2.
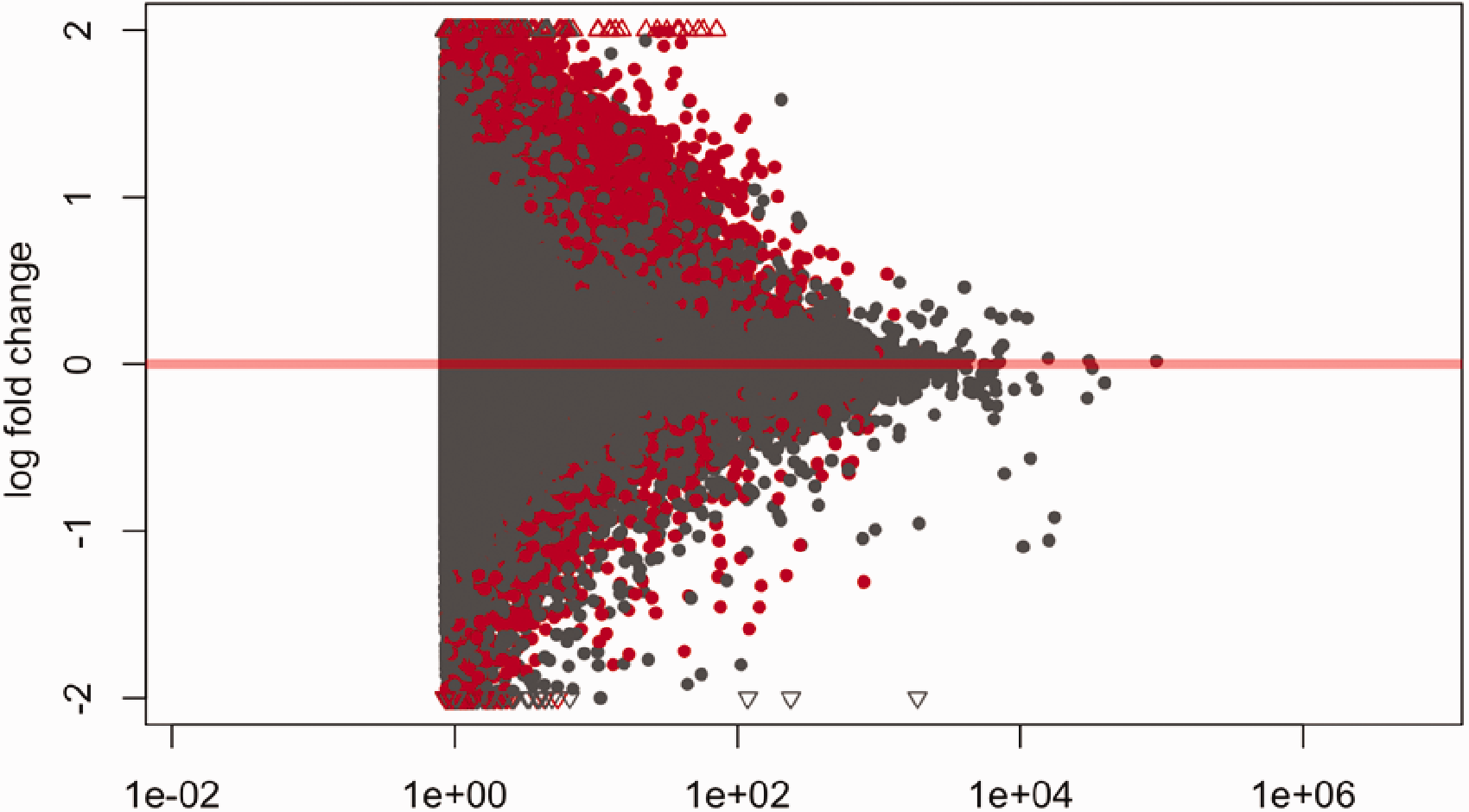
Mean-average (MA)-plot of normalized mean expression versus log2-fold change. The red marks represent the log of fold change versus average normalized count per exon that are significant (FDR <0.05). The red triangles represent exons with log2-fold change greater than 2 or less than −2. The grey marks and triangles represent non-significant results. (A color version of this figure is available in the online journal.)
The purpose of the study was to investigate AS events between 18 paired normal-tumor OS samples by RNA-seq. As shown in Figure 2(a), statistically significant (FDR < 0.05) changes in expression levels of leptin receptor overlapping transcript (LEPROT) exons were observed between normal and tumor samples. Most of the exons are expressed in normal tissue, while expression in the tumor sample remained low. However, DEXseq removes the exon level changes in expression to highlight differential exon usage of the gene. The exons shown in purple presented with significant differential exon usage (bottom panel, bin E014, E040, and E042), suggesting potential AS events (Figure 2(b)). The comparisons of each exon expression between tumor and normal samples were performed by normalization counts, as shown in Figure 2(c).
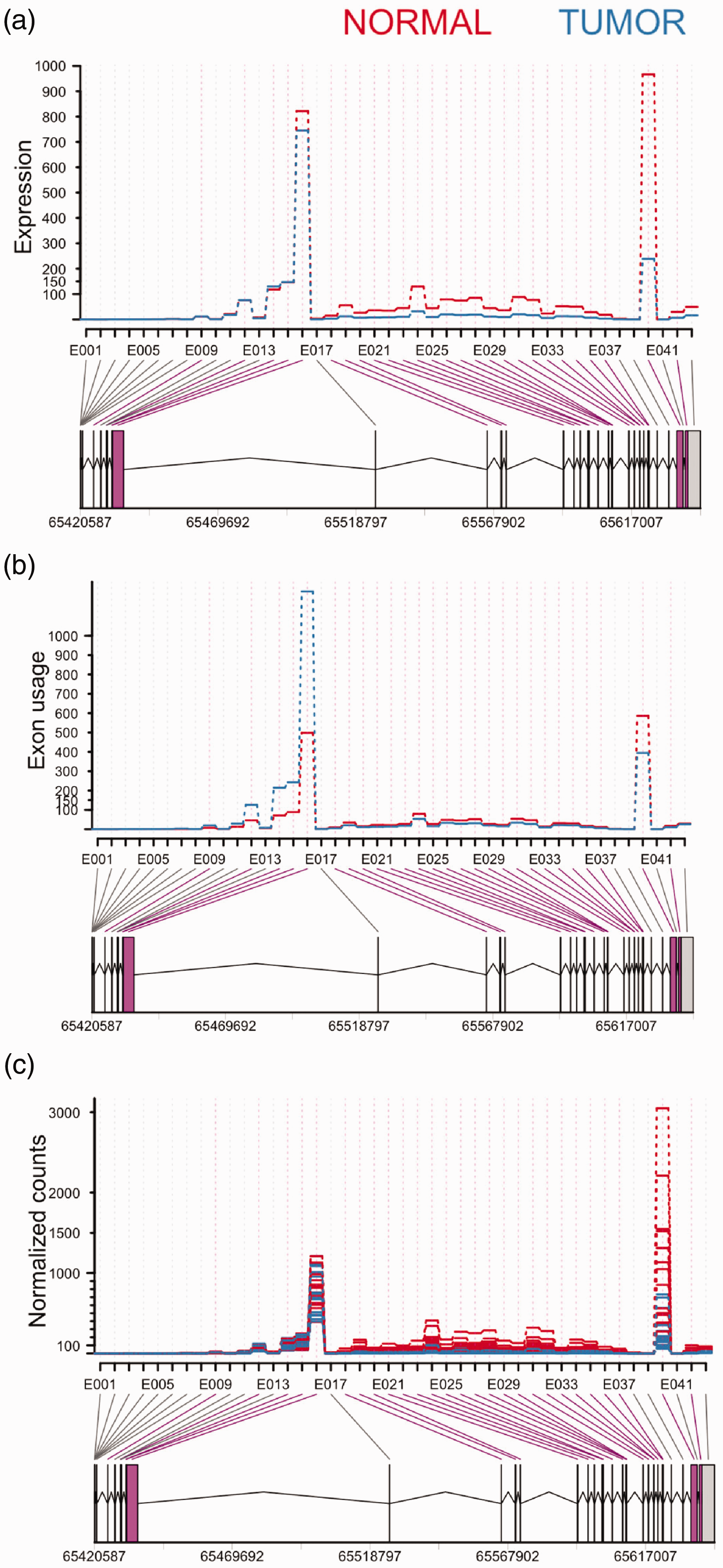
DEXseq analysis on RNA-Seq data. The x-axis shows individual exons within LEPROT gene, whereas the y-axis represents exon expression (a), exon usage (b), and normalization counts (c). The bars below the x-axis represent exons. The lines between the exons are introns. The numbers at the bottom are genomic locations of LEPROT. A highlights the fitted expression values of each of the exons of LEPROT gene for normal (red) and tumor (blue) samples. B visualizes exon usage in the gene. The exon in purple (E014, E040 and E042) and 26 purple lines between the bars and exon names indicate a significant differential exon usage (FDR < 0.05), whereas E037 revealed insignificant splicing (FDR > 0.05). C shows the comparison of exon expression between the samples. (A color version of this figure is available in the online journal.)
We identified 27 novel transcript variants of the LEPROT gene (Table 2), 26 of those transcripts have statistically significant (FDR < 0.05) different exon usage. This suggests that different isoforms use the exons differently. One transcript (E037) identified as having different exon usage was subsequently shown to be non-significant (FDR > 0.05, highlighted in red in Table 2).
Differently spliced transcript variants of LEPROT.
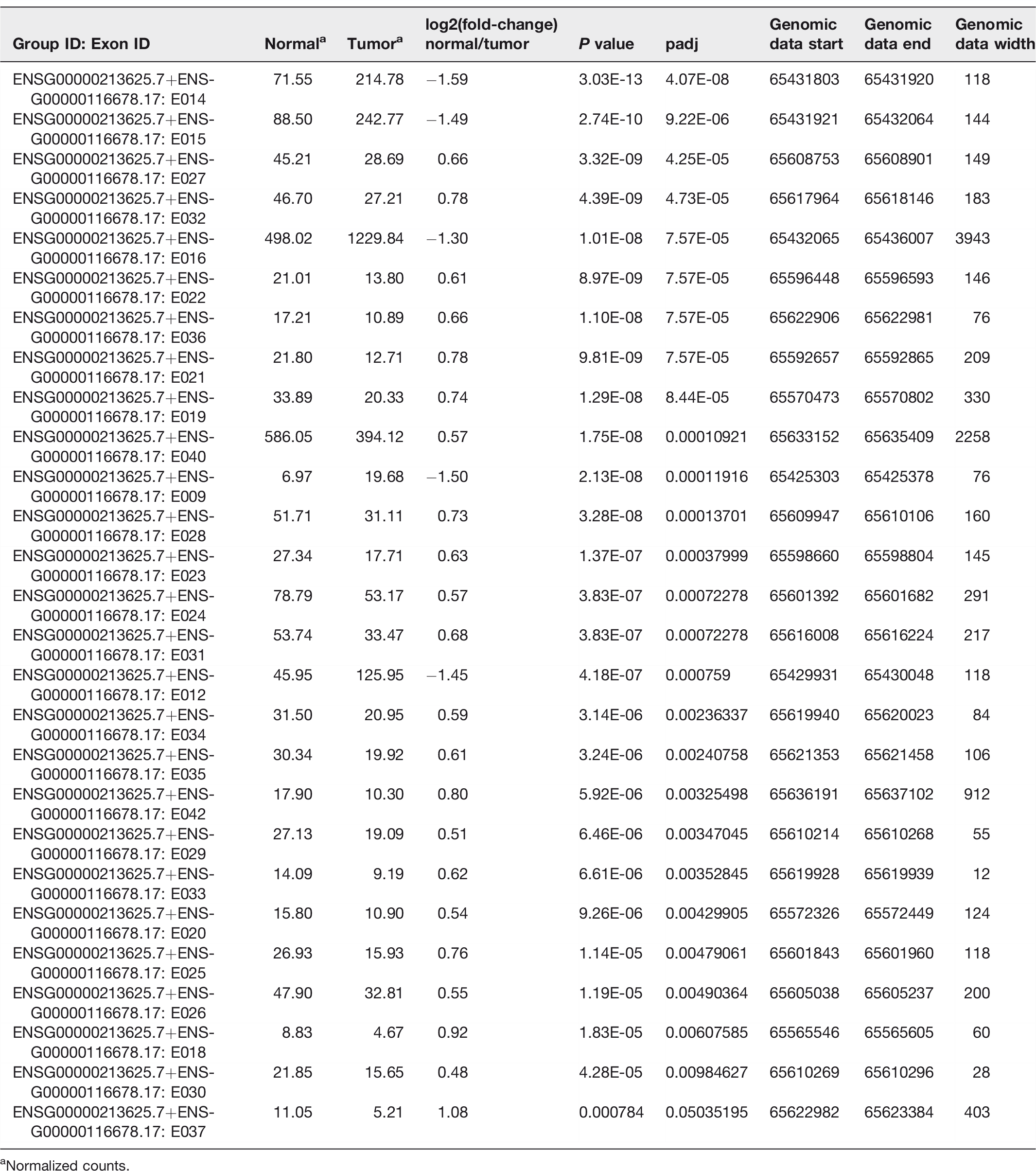
aNormalized counts.
As visualized in Figure 2(a) and listed in Table 2, 5 transcripts of LEPROT demonstrated overexpression in the tumor samples, while 22 transcripts (21 significant/1 non-significant) presented with overexpression in the normal samples. Decrease expression of LEPROT transcripts in tumor samples eventually downregulates general LEPROT expression in tumor samples.
Discussion
The limiting factor in improving treatment for OS is a lack of understanding of the molecular mechanism of the disease, which leads to an inability to diagnose and treat OS at an earlier stage, or to develop more effective therapies. RNA-seq can be a potentially powerful method to identify differential exon usage in OS and allow identification of potential biomarkers of the disease. 27 Differential usage of exons produces different transcripts from the same gene. 18 Cancerous cells can have alternate patterns of exon usage within individual genes compared to normal cells, suggesting that AS may play a key role in shaping the phenotype of the tumor. 23
In this study, we have investigated AS events in 18 paired normal – tumor OS samples. Interestingly, our results showed that the LEPROT gene underwent AS and identified 26 statistically significant (FDR < 0.05) novel transcripts. These transcripts showed different expression levels between tumor and normal tissues, suggesting there is a potential functional association between AS of LEPROT and cancer progression.
A genome-wide expression analysis has suggested an association between LEPROT regulation and apoptosis pathway. 37 Unfortunately, the study did not explain the association further. However, the manifestations of LEPROT gene mutations have not been extensively studied, resulting in an incomplete understanding of the function or dysfunction of this gene in cancer metabolism. 38
LEPROT is expressed widely in many human tissues, but does not appear to be expressed in tumors. 39 Therefore, loss or frequent downregulation of LEPROT expression could be associated with tumor formation. The cause of downregulation of the LEPROT gene in cancer cells remains unknown. One study has suggested that AS can downregulate overall gene expression by tagging specific transcripts for degradation through the nonsense-mediated mRNA decay (NMD) surveillance pathway. 40 NMD frequently controls the translation stage and degrades the transcripts if they contain a premature termination codon (PTC), which can arise from genetic mutations or AS.41,42 Truncated proteins cannot function properly or could even be toxic to the body; therefore, the synthesis of truncated proteins can contribute to various cancer types. 43 We therefore encourage further investigation of the regulation of LEPROT gene expression in normal and tumor cells. Our analysis highlighted LEPROT transcripts as being significantly under-expressed in 21 out of 26 tumor samples. Decrease expression of LEPROT transcripts in tumor samples, eventually can lead to reduction of total LEPROT expression level. The five overexpressed transcripts (absolute value of log2 fold-change > 1) can be considered as the noise from overall downregulation of LEPROT gene in OS. Therefore, it can be argued that they are not specific to OS initiation and progression. LEPROT is encoded by LEP receptor (LEPR) gene and they share the first two 5ʹ-UTR exons. 44 Unsurprisingly, we previously identified significant downregulation of LEPR in OS. 30
Several studies have discussed the association between LEPROT expression level and growth hormone (GH) activity. 44 GH stimulates the development of bone and cartilage in children and adolescents. 45 It also has been associated with induction of insulin-like growth factor 1 (IGF-1) and IGF-1 receptor (IGF-1R) gene expression, alterations in glucose metabolism, and modulation of cell proliferation genes.46–48 IGF-1 is produced in the liver, stimulated by GH, and plays a main role in tissue growth and development.49–52 Higher circulating levels of IGF-1 have a profound impact on cell proliferation, differentiation, promotion of cellular longevity, and inhibition of apoptosis.53,54 Consequently, overexpression of IGF-1/IGF-1R has been implicated in tumor formation, angiogenesis, and metastasis in various human cancer types, including OS.46–48,53 According to one study, LEPROT is directly involved in a receptor-mediated cell signaling pathway by regulating cell surface expression of growth hormone receptor (GHR) at the molecular level. The same study also suggested that silencing of LEPROT increases cell-surface expression of GHR in a mouse model. 55 Thus, increasing GHR expression level will lead to higher circulating levels of IGF-1 and may eventually result in tumorigenesis.47–49,53,55
Dysregulation of LEPROT is associated with various bone inflammation diseases in humans through the key inflammatory cytokines such as tumor necrosis factor alpha (TNF α) and interleukin 6 (IL-6). Several studies have highlighted that bone inflammation is a frequent precursor to initiation and progression of OS. 56 Further, OS cells promote local inflammation and this leads to activation of local immune responses. 57 Not surprisingly, upregulation of IL-6 and TNF-α has been observed in human OS cells.58,59 In addition, IL-6 mediates tumor-host interactions that facilitate lung colonization of metastasis-initiating by OS cells.60,61 Consequently, LEPROT may be implicated in OS initiation and metastasis through upregulating expression of IL-6 and TNF-α.
In conclusion, our data provide strong evidence that AS events of the LEPROT gene may be a risk factor for OS, and downregulation of LEPROT can be associated with tumorigenesis in bone. These results may facilitate a better understanding of the underlying molecular mechanism related to the initiation and progression of OS, thereby allowing identification of effective novel molecular target candidates for the treatment of the disease.
Footnotes
Authors’ contributions
ER wrote the manuscript. XDH contributed in samples collection and laboratory works. JX provided valuable opinions related to the study. AM and KM participated in design of the experiments. DW and SK supervised the experiments. All authors revised the manuscript.
DECLARATION OF CONFLICTING INTERESTS
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical approval
The Ethics Review Committee on Biomedical Research of Hue University of Medicine and Pharmacy approved the protocols and informed consent form elaborated in the study. The informed consent forms signed and dated by the participants and representatives of patients.
FUNDING
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The study has been supported by institutional research funding (IUT20-46) from the Estonian Research Agency and by the European Union’s Seventh Framework Programme (FP7/2007–2013) under a grant agreement [grant number 602398] (HypOrth).