
Abstract
As an alternative to pharmacological treatment to diseases, lifestyle interventions, such as dietary changes and physical activities, can help maintain healthy metabolic conditions. Recently, the emerging analyses of volatile organic compounds (VOCs) from breath and short-chain fatty acids (SCFAs) from plasma/feces have been considered as useful tools for the diagnosis and mechanistic understanding of metabolic diseases. Furthermore, diet-induced changes of SCFAs in individuals with diagnosed metabolic abnormalities have been correlated with the composition changes of the gut microbiome. More interestingly, the analysis of exhaled breath (breathomics) has gained attention as a useful technique to measure the human VOC profile altered as a result of dietary interventions. In this mini-review, we examined recent clinical trials that performed promising dietary interventions, SCFAs analysis in plasma/feces, and VOC profile analysis in exhaling breath to understand the relationship between dietary intervention and metabolic health.
Impact statement
The prevalence of metabolic diseases such as obesity and diabetes has been rapidly increasing in the 21st century, and it has become a major global health problem. Recently, changes in lifestyle, such as dietary modifications, were found useful in preventing metabolic diseases. Meanwhile, the positive impact of dietary interventions on human health has been linked with the production of short-chain fatty acids (SCFAs) by gut microbiota and exhaled breath volatile metabolites through the complicated metabolic process. Thus, monitoring SCFAs and exhaled breath volatiles as biomarkers can be used to evaluate the effectiveness of dietary interventions on metabolic diseases in clinical studies. The dietary intervention-induced alteration of SCFAs and exhaled volatiles can also be useful for the development of novel prevention approaches and drug-free treatments for metabolic diseases.
Introduction
Normal metabolism is the key to good health. On the other hand, altered metabolic processes generally lead to the development of metabolic diseases such as obesity, diabetes mellitus, dyslipidemia, and osteoporosis. 1 In our modern age, the rapidly elevated prevalence of metabolic diseases is considered as a major health problem. 2 It has been reported that more than a third of adults in the United States are diagnosed with metabolic abnormalities, most likely due to an unhealthy lifestyle. 3 As preferred alternatives to therapeutics or surgical treatments, lifestyle changes such as dietary modifications and adopting a physically more active lifestyle can be beneficial to people with metabolic dysregulations. 4 Considering this, various clinical studies have been undertaken to assess the impact of dietary intervention on different metabolic diseases.4–6
The diagnosis of metabolic diseases is generally cumbersome, and causes, risks, or consequences of metabolic diseases have been assessed by measuring altered levels of key physiological metabolite biomarkers as an important diagnostic approach. 7 These biomarkers are used as an indicator of a given biological state or diseased condition. 8 However, very few biomarkers related to the nutritional effects on the development and progression of various metabolic diseases have been identified so far. The complex pathophysiology of metabolic diseases underlines the limitation of known routine markers such as lipid profiles, glucose, and insulin in their diagnostic accuracy, and nutritional interventions add additional layers of complexities on top of these factors. In recent years, metabolomics, a systematic study of metabolites, has been considered as a powerful approach in biomarkers discovery with the promise to be eventually applied to clinical settings. 9
Thus far, mass spectrometry (MS)-based metabolomics is considered as a useful tool in clinical studies to investigate the effect of dietary interventions in improving metabolic disorders, including discovering potential biomarkers of route-cause pathophysiology of these syndromes. 10 An ultra-performance liquid chromatography coupled with tandem mass spectrometry (LC-MS/MS) and a gas chromatography equipped to mass spectrometry (GC-MS) are the most popular analytical techniques for metabolomics analyses.11–14 Nowadays, the use of these analytical tools in dietary interventions associated with human metabolic disorders has been growing, especially when it comes to identifying and quantifying small molecular weight compounds, volatile organic compounds (VOCs) and short-chain fatty acids (SCFAs), in human biofluids such as plasma and feces.11,15–17 SCFAs are produced as a result of fermentation of indigestible foods by gut microbiota and are normally required for maintaining optimal gut health. 18 These SCFAs act as signaling molecules that can enter the systemic circulation and directly affect metabolic or peripheral tissue functions. 19 They were also found to have a considerable role in the control of body weight and insulin sensitivity. 19 The production of SCFAs is influenced by the diet-mediated changes and physiological activities in the gut. 20 Thus, critical assessment of SCFAs levels can be crucial in understanding healthy and diseased conditions. Recently, the exhaled VOC analysis, called breathomics, has also been considered as a promising tool for investigating metabolic responses in a non-invasive manner. However, the significance of breathomics in clinical trials of dietary interventions, especially in biomarker quantification, remained to be fully explored. In this mini-review, we summarize the research progress of SCFAs production, breathomics, and their potentials as real-time biomarkers in the context of human dietary interventions to human metabolic health.
Short-chain fatty acids and their connection with diet, gut microbes, and human health
It is well-known that humans cannot digest the bulk of dietary fibers due to the lack of related enzymes, and the non-digestible carbohydrates are fermented in the large intestine by the colonic/gut microbiota.20,21 SCFAs, saturated aliphatic organic acids, become the major group of metabolites that are generated by the gut microbial community as a consequence of fermentation of dietary fibers (Figure 1).22,23 SCFAs consist of one to six carbons, and acetate (C2), propionate (C3), and butyrate (C4) have been reported as the most abundant ones by representing 90–95% of the SCFA present in the colon.20,24 Previous studies have identified that the formation of SCFAs is influenced by various factors, including the pattern of food intake, antibiotics treatment, and microbial populations.20,25–27 In particular, carbohydrates are the main source of SCFAs production; meanwhile, amino acids, such as valine, leucine, and isoleucine from the protein breakdown are also involved in the production of branched SCFAs such as isobutyrate, isovalerate, and 2-methyl butyrate. 20
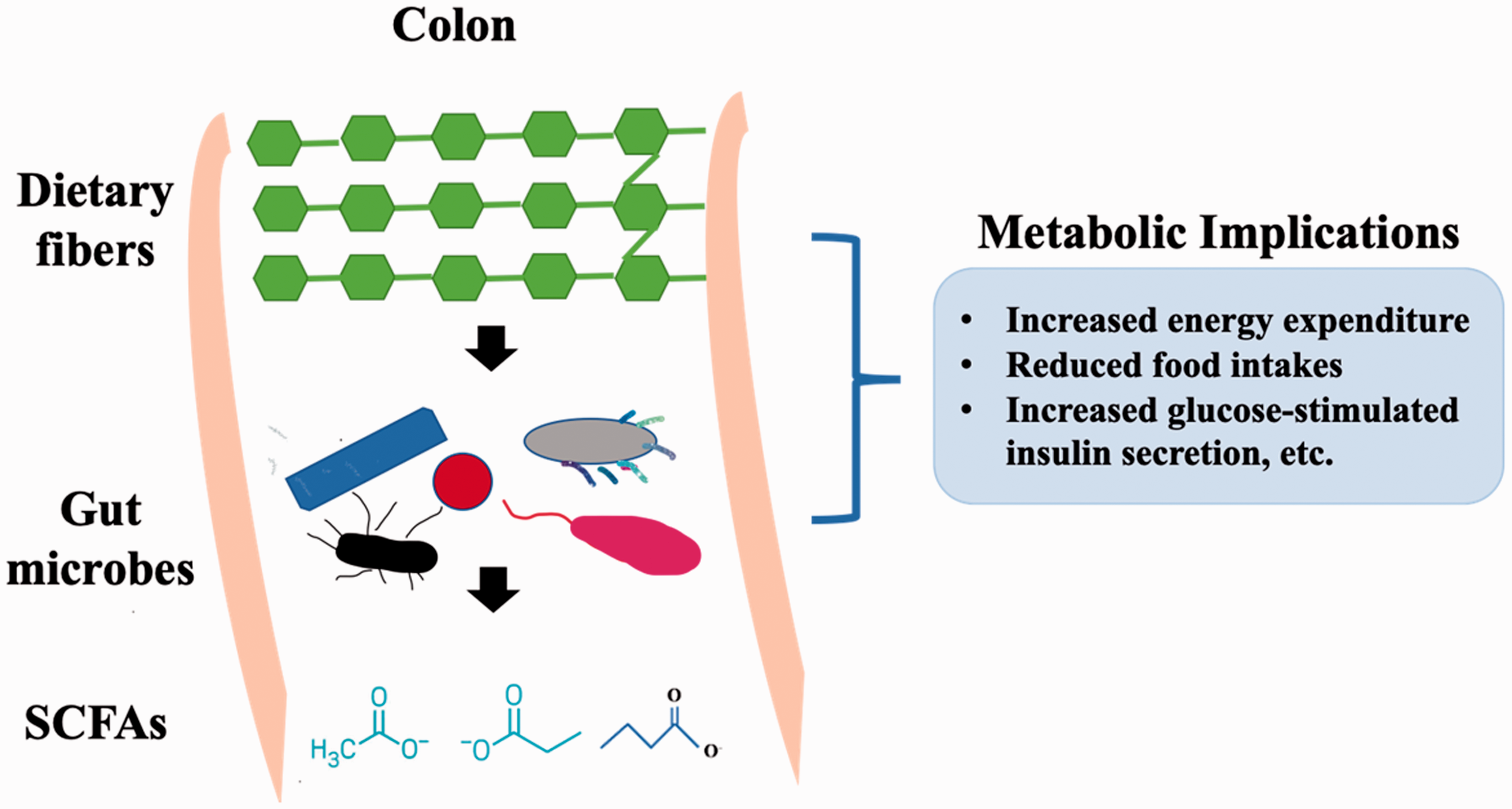
A diagram demonstrates the relationships of dietary fibers, gut microbes, their SCFA metabolites, and their implications to host metabolic health. (A color version of this figure is available in the online journal.)
Earlier studies have described the metabolic routes of several SCFAs. For example, two major paths for synthesizing acetate are initiated from formic acid through the Wood–Ljungdahl pathway or from hydrogen and carbon dioxide by acetogenic bacteria.28,29 In the case of propionate generation, three pathways, involved in succinate, acrylate, and propanediol synthesis, are considered as major routes. 30 Among them, propionate production via succinate pathway is prevailing, and several Firmicutes and Bacteroidetes were found to be involved in succinate pathways, and their richness was also related to the level of propionate in fecal samples. 31 Another pathway involving the butyryl-CoA:acetate CoA-transferase gene for butyrate formation has been elucidated with the participation of butyrate-producing bacteria, 20 which can convert butyryl-CoA to butyrate with a single step enzymatic reaction in the presence of butyrate-producing Firmicutes, such as Faecalibacterium, Eubacterium, and Roseburia. 32 The connections between these elucidated pathways of SCFA productions and the involvement of multiple members of the gut microbial community highlight the need to study the chemical fingerprints profiles in conjunction with the microbial fingerprints in metabolic studies on the evaluation of dietary intervention effects to patients with metabolic dysregulations.
Besides, epidemiological studies have demonstrated that the gut microbiome may participate in host-signaling mechanisms via microbial metabolites, as well as use SCFAs as energy sources to feed host cells. 20 SCFAs were also found to influence host physiology and potentially modulate the development of metabolic syndrome by incorporating into glucose and lipid metabolism (Figure 1).33–35 Also, SCFAs play vital roles in the maintenance of the gut barrier function as fuels for the intestinal epithelial cells which may lead to the increase of mucin productions to prevent occurrence and development of gut diseases.20,36,37 Mainly, butyric acids have been reported to prevent bacterial adhesion and improve the tight-junctions integrity.38,39 Furthermore, the immune-modulating function of the microbial community that produces SCFAs has been described during the host inflammatory processes. 40 For instance, it has been reported that butyrate and propionate may promote the differentiation of T-regulatory cells into effector and regulatory T cells via suppression of histone deacetylase. 41 Consequently, the reduced incidence of intestinal inflammation may have a beneficial impact on improving host immune tolerance and reducing the risk of inflammatory bowel disease and colorectal cancer. 20
Human breath volatile analysis: The new frontier in human health research
Thousands of VOCs are emitted from the human body, and their components can reflect a person’s metabolic condition. Therefore, metabolic diseases often result in a change in body smell. 42 These VOCs can be originated from breath, urine, feces, saliva, sweat, and skin. Among them, exhaled breath contains hundreds of reported VOCs that can be attributed to either exogenous or endogenous sources. 43 For example, exogenous VOCs include compounds inhaled or derived from the external environment, such as smoking a cigarette or ingesting food. On the other hand, endogenous VOCs consist of circulating bloodstream compounds that were produced by many metabolic processes at different organs/tissues including ones from colonic microbial metabolism and released through the lungs. The concentrations of VOCs in breath vary from picomolar to nanomolar levels, which make it challenging to distinguish endogenous VOCs from the exogenous compounds. 44 Nonetheless, breath sampling has gained attention for its non-invasive, extremely simple, and painless manners. Therefore, the breath analysis has become the top choice for some clinical studies of which clinicians and researchers have managed to successfully identify disease-specific VOCs. For example, trimethylamine was detected in the breath of patients with trimethylaminuria as a potential diagnostic marker; and in the breath of patients with fetor hepaticus, methyl-mercaptan was identified as a useful biomarker for the diagnosis of this disease.45,46
Metabolic diseases often result in the production and accumulation of certain metabolites in body fluids due to impairment in normal biochemical pathways. Therefore, the detection of increased concentrations of these metabolites can be used for the diagnosis of these diseases. 47 For example, in the case of diabetes, cells utilize glucose as the main source of energy in the body, and the pancreas produces insulin to signals cells for glucose uptake. 48 Glucose then accumulates in the blood of patients with type 1 diabetes who cannot synthesize insulin, and eventually excreted in the urine. Given these unusual changes, fatty acid catabolism will be promoted as cells tend to use fatty acids rather than glucose as an energy source. As a result, this change creates ketone compounds, such as acetone, acetoacetate, and 3-hydroxybutyrate as by-products, and these ketone bodies cause blood acidity, leading to ketoacidosis of the blood. These excess ketones are secreted in the urine and breath. 48 As a result, the patient’s urine and breath could have a fruity odor due to the presence of acetone. Sequentially, it is reasonable to expect that when interventions such as dietary patterns’ changes were applied to these patients, their breath patterns could change accordingly (Figure 2).
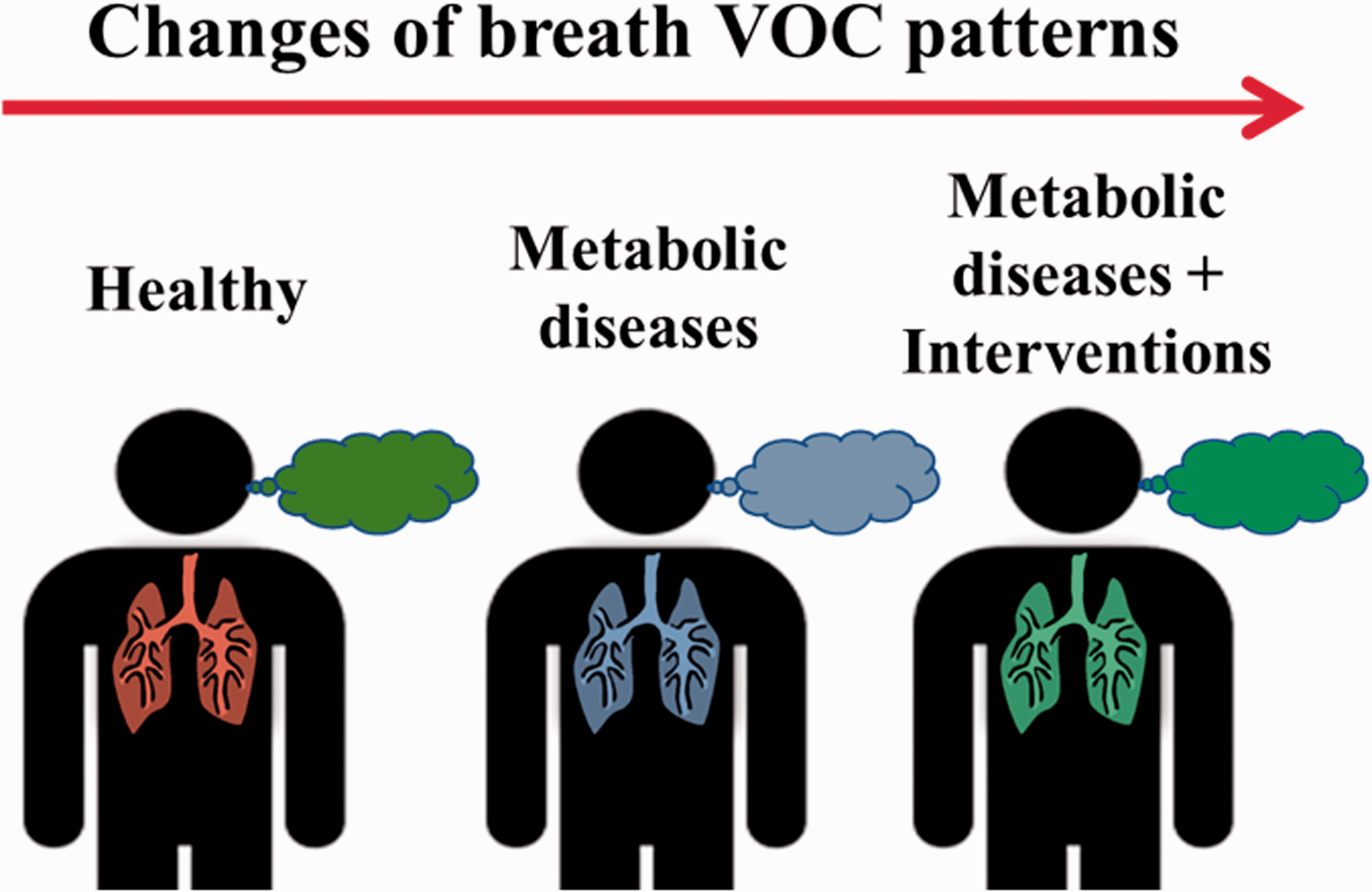
A schematic demonstrates the possible breath VOC pattern changes from a healthy individual to a patient with metabolic diseases, and to the patient who went through the dietary interventions of the metabolic diseases. Different colors of the breath indicate different breath VOC patterns. (A color version of this figure is available in the online journal.)
Overview of clinical trials of dietary interventions with constant monitoring of SCFAs
It is well known that individuals with prediabetes have increased risks of the development of type 2 diabetes (T2D) and multiple vascular complications. 49 Numerous randomized controlled trials (RCTs) have demonstrated the feasibility of preventing progression from prediabetes to T2D, using lifestyle or pharmacological interventions.50,51 studies reported in the past decade on the detections of SCFAs associated with dietary interventions were summarized in Table 1. The RCTs reviewed here provide compelling evidence that lifestyle modification can decrease the risk of progression to T2D in people with prediabetes with a maximum effect size of ∼60% risk reduction compared with placebo.52,53 Out of the various lifestyle modifications, precision nutrition has the potential to provide personalized dietary guidance for more effective prevention or management of T2D. 54 A majority of these clinical studies have investigated the effectiveness of dietary interventions for preventing or delaying metabolic abnormality in individuals with MetS or at high risk for MetS (e.g. obese or overweight) by leveraging SCFAs as potential biomarkers.
Summary of dietary interventions induced changes in SCFAs productions in populations with metabolic abnormalities.
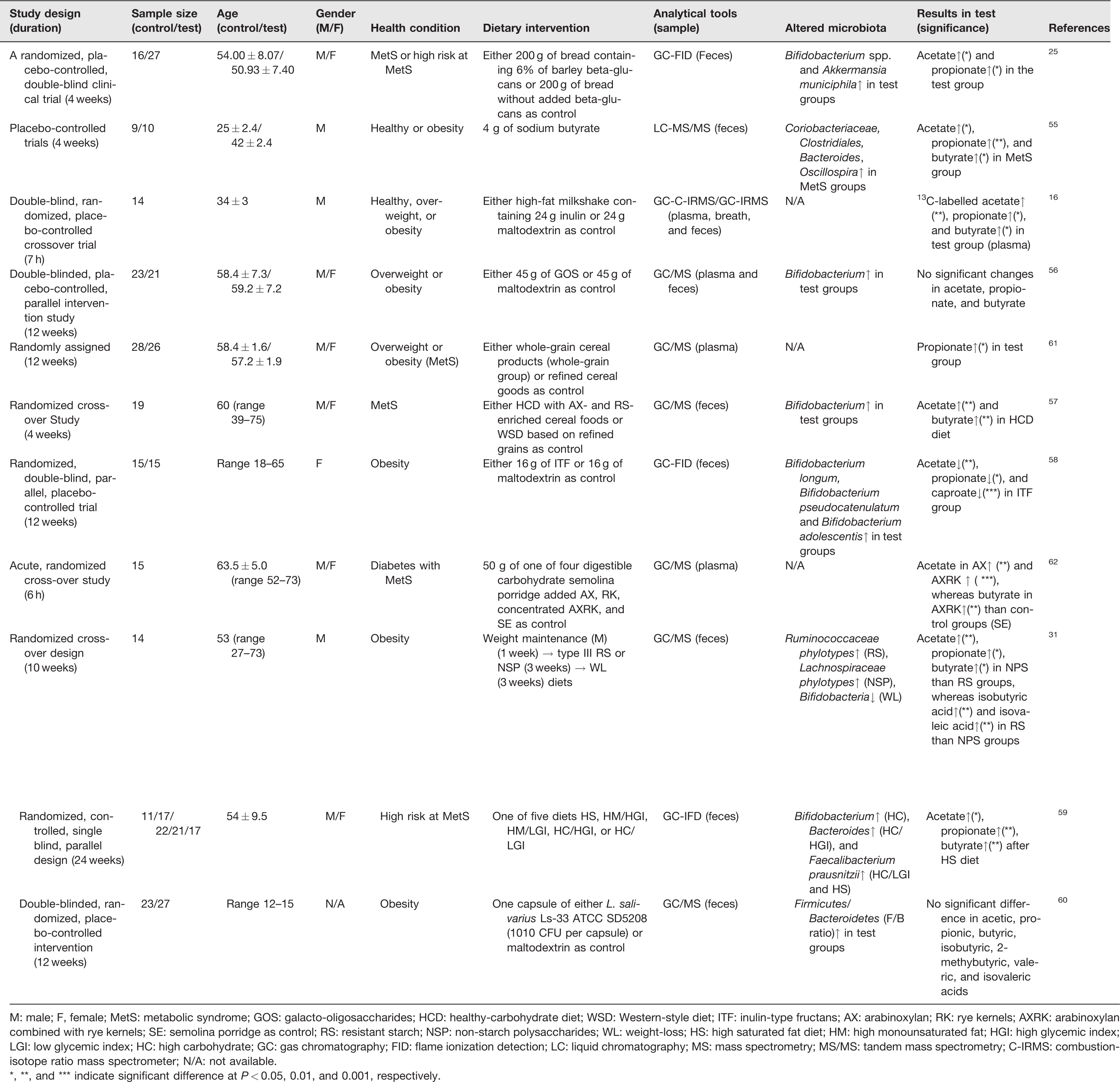
M: male; F, female; MetS: metabolic syndrome; GOS: galacto-oligosaccharides; HCD: healthy-carbohydrate diet; WSD: Western-style diet; ITF: inulin-type fructans; AX: arabinoxylan; RK: rye kernels; AXRK: arabinoxylan combined with rye kernels; SE: semolina porridge as control; RS: resistant starch; NSP: non-starch polysaccharides; WL: weight-loss; HS: high saturated fat diet; HM: high monounsaturated fat; HGI: high glycemic index; LGI: low glycemic index; HC: high carbohydrate; GC: gas chromatography; FID: flame ionization detection; LC: liquid chromatography; MS: mass spectrometry; MS/MS: tandem mass spectrometry; C-IRMS: combustion-isotope ratio mass spectrometer; N/A: not available.
*, **, and *** indicate significant difference at P
In Table 1,2 out of 11 clinical trials used healthy individuals as control groups, in which the same diet was assigned to both healthy and obese/overweighted individuals as test groups.16,55 The sample sizes of these studies ranged from 14 to 88 subjects, and most of these clinical trials were designed in a randomized and blind manner. The duration of the dietary intervention varied from 7 h to 24 weeks. Mainly, plasma or fecal samples were chosen for metabolic analysis, and particularly, eight cases not only investigated fecal SCFAs, but also reported the diet-induced alteration of fecal microbial composition using the 16S ribosomal RNA analysis.25,31,55–60 As summarized in Table 1, liquid chromatography 55 and gas chromatography31,56,57,60–62 were generally combined with derivatization methods and MS, which have been most commonly used as effective analytical techniques. In other cases, gas chromatography was coupled with isotope ratio MS 16 or flame injection detectors25,58,59 for the determination of small and polar SCFAs.
Summary of dietary interventions induced changes of exhaled breath VOCs in selected clinical trials.
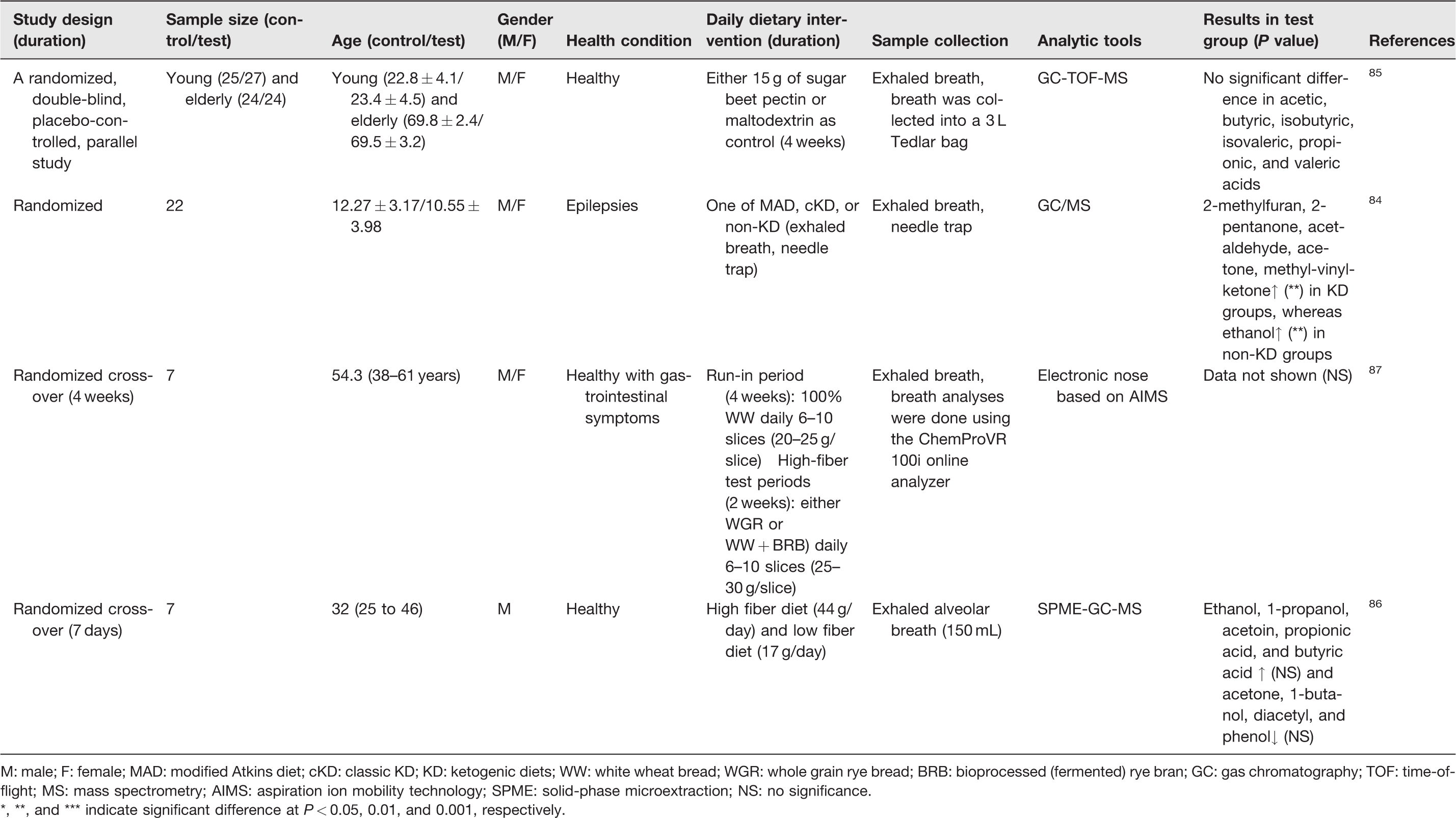
M: male; F: female; MAD: modified Atkins diet; cKD: classic KD; KD: ketogenic diets; WW: white wheat bread; WGR: whole grain rye bread; BRB: bioprocessed (fermented) rye bran; GC: gas chromatography; TOF: time-of-flight; MS: mass spectrometry; AIMS: aspiration ion mobility technology; SPME: solid-phase microextraction; NS: no significance.
*, **, and *** indicate significant difference at P
Overall, several RCTs demonstrated the beneficial effects of dietary components on preventing the risk of developing metabolic derangements according to the diet-induced change in metabolic and anthropometric parameters.16,25,56,58–60 Thus, the results of these RCTs may suggest a variety of effective dietary intervention strategies that last from days to months, and successful metabolic risk management can be achieved. Meanwhile, small-molecule metabolite biomarkers, especially SCFAs, have been considered as important indicators related to biochemical changes that underlying the physiological changes after dietary intervention. Epidemiologic studies have encouraged dietary fiber intakes in diabetic and metabolic disease patients based on its health benefits such as improving insulin sensitivity and SCFAs producing intestinal microflora.63,64 Here, we focused on previous RCTs which mostly tested dietary fiber-based ingredients, including barley beta-glucans, inulin, galactooligosaccharides, whole-grain cereal products, healthy-carbohydrate diet containing rich arabinoxylan and resistant starch, inulin-type fructans, rye kernels, concentrated arabinoxylan combined with rye kernels (AXRK), wheat bran non-starch polysaccharides, or non-dietary fiber ingredients such as sodium butyrate and Lactobacillus salivarius to examine diet-induced changes of SCFAs (Table 1).
The diet-induced changes of SCFAs in subjects diagnosed with MetS or at a high risk of metabolic diseases varied depending on the types of experimental ingredients. For instance, studies on diets containing rich dietary fibers, such as barley beta-glucans, inulin, whole-grain cereal products, a healthy-carbohydrate diet with rich arabinoxylan and resistant starch resulted in increased amounts of SCFAs (acetate, propionate, and butyrate), compared with control groups.16,25,57,61 Meanwhile, galactooligosaccharides and inulin-type fructans were not found to lead significant influences by presenting no considerable changes or decrease of SCFAs, respectively.56,58 In some cases, diverse patterns of SCFAs alteration were observed according to different types of foods. For example, a study by Hartvigsen et al. examined the effect of 50 g of semolina porridge as control, or 50 g of control diet supplemented with one of three digestible carbohydrates, namely arabinoxylan, rye kernels, or concentrated AXRK, on individuals with diabetes. As a result, significantly increased acetic acid abundance was found in the group that ingested arabinoxylan diets. Whereas higher levels of acetic acid and butyric acid were detected from the AXRK-intaking group, compared with control groups. 62 Overall, these clinical trials have shown that a dietary fiber-enriched diet can affect the level of plasma or fecal SCFA in a fiber type-specific manner.
Regulations of SCFA productions and their related gut microbes
Previous studies have shown that the diet is one of the key regulatory factors, in terms of modulation of the symbiosis or dysbiosis of gut microbiota, and ultimately affects human health.65,66 The food rich in dietary fibers has been reported to prevent and mitigate T2D mellitus, cardiovascular disease, and colon cancer.67,68 Furthermore, SCFAs are produced by the fermentation of fibers and counteract obesity by reducing fat accumulation in adipose tissues and accelerating energy expenditure.69–71 Commonly, clinical studies perform in-depth gut microbiota analysis while examining metabolic changes in humans in response to diet. Most of the studies reviewed in Table 1 were conducted with the evaluation of the effects of specific dietary ingredients on changes in the gut microbial population along with the SCFA production. Mainly, gut microbes from four bacterial phylum, including Actinobacteria (Bifidobacterium and Coriobacteriaceae), Verrucomicrobia (Akkermansia municiphila), Firmicutes (Clostridiales, Oscillospira, Ruminococcaceae, Lachnospiraceae, and Faecalibacterium), and Bacteroidetes (Bacteroides), were identified at genus, species, family, or class levels as described in Table 1. In general, Actinobacteria is known as a minority group of commensal bacteria, and it plays a pivotal role in the development and maintenance of gut homeostasis, involved in the modulation of gut permeability and the immune system. 72 Moreover, health beneficial Actinobacteria species, such as Bifidobacterium and Coriobacteriaceae, have been reported to reduce obesity, blood lipids, and cardiovascular risk in patients with MetS and to improve T2D conditions.73,74 Akkermansia municiphila (Phylum Verrucomicrobia), a mucin-degrading bacteria, is also described as SCFA-producing (such as propionate and acetate).75,76 Firmicutes and Bacteroidetes are the major population in healthy gut microbiota. 77 Consumption of plant-based diets may regulate the growth of Firmicutes and Bacteroidetes, and the reduced ratio of abundance of Firmicutes to Bacteroidetes (F/B) has been reported to indicate healthier gut microbiota. 78 In addition, changed F/B ratios have also been found to be associated with various pathologies. For instance, the decreased F/B ratio has been linked to colonic inflammation in patients with inflammatory bowel disease, and the altered F/B also has been observed in obese and T2D.79–82
In most cases in Table 1, increased abundances of bacteria were observed along with elevated levels of SCFAs after consuming dietary fibers (barley beta-glucans, resistant starch) or carbohydrate-based diets.25,31,55–57,59 In a study by Hald et al., a healthy-carbohydrate diet with arabinoxylan- and resistant starch-enriched cereal foods, or a Western diet based on refined grains was given to participants diagnosed with MetS in a randomized and crossover fashion. 57 The substantially enhanced values of acetic acid and butyric acid were observed along with the increased Bifidobacterium abundance in healthy-carbohydrate diet groups. 57 In another study, either 200 g of bread containing 6% of barley beta-glucans (beta-glucans bread) as a test diet or 200 g of bread without added beta-glucans as control were provided to a group of individuals either have MetS or have a high risk of developing one. After the dietary intervention, significantly changed levels of SCFAs in feces were observed with elevated propionic acid in the test group (43.2%, P = 0.045). 25 Meanwhile, significantly increased Bifidobacterium spp. and Akkermansia municiphila were detected in a subgroup of participants who have strong cholesterol reduction after the beta-glucans bread intervention. 25 These results showed that even though a large cohort study may generate a statistically meaningful dataset, a more personalized and precise nutritional intervention that relying on multiple layers of information needs to be tailored to ensure effectiveness.
In other cases, interventions with sodium butyrate and inulin-type fructans that resulted in the decreased SCFAs contents were observed, despite the increased values of some detected bacteria.55,58 For instance, Salazar et al. reported that the significantly decreased levels of acetic acid, propionic acid, and caproic acid were detected in obese people after ingesting 16 g of inulin-type fructans than the observed levels in those who served with 16 g of maltodextrin as controls. 58 Interestingly, however, they did detect a richer abundance of Bifidobacterium longum, Bifidobacterium pseudocatenulatum, and Bifidobacterium adolescentis in the intervention groups. 58
More complicated, in another study by Fava et al., different combinations among high saturated fat (HS), high monounsaturated fat (HM), high glycemic index (HGI), low glycemic index (LGI), and high carbohydrate (HC) were prepared as HS, HM/HGI, HM/LGI, HC/HGI, or HC/LGI diets, and one of these five was served to those who are in high risk of developing MetS. 59 The fecal SCFA analysis revealed significantly increased acetic acid, propionic acid, and butyric acid levels after the HS diet. Moreover, changes in microbial population were closely connected to the types of diet, for example, considerably richer levels of Bifidobacterium and Bacteroides were detected in HC and HC/HGI groups, respectively, as well as increased abundance of Faecalibacterium prausnitzii was observed in both HC/LGI and HS diets. 59
Overall, most of these changes in gut bacteria in Table 1 were considered as results of experimental diet intakes, and the production of SCFAs, such as acetate, butyrate, and propionate, were also observed at relatively higher levels compared with control groups. However, exceptions that did not fall into such a pattern also exist. For example, a galactooligosaccharides intervention rendered an increased number of Bifidobacterium, but no significant difference of SCFAs was reported among the detected compounds. 56 Similarly, enhanced levels of B. longum, B. pseudocatenulatum, and B. adolescentis were found in a group of participants after the inulin-type fructans-based dietary intervention, with substantially reduced contents of acetate, propionate, and caproate were detected in the same group. 58 Taken together, it is clear that many further studies are needed to deeply understand the interaction between the gut microbiome and the SCFA productions, and their implications in the prevention and treatment of several systemic metabolic abnormalities related to human health. 72
The impact of dietary intervention on human volatile
Despite extensive researches have focused on blood metabolite biomarkers, their clinical translations remain challenging, therefore research efforts have been continued by many scientists and physicians in this area. In this context, interests in the metabolic analysis have been shifted to breathomics studies, which are defined as a special branch of metabolomics to detect VOCs in the breath of individuals as potential biomarkers for screening and detecting diseases. 83 Specific profiles of volatile compounds in exhaled breath, and metabolites in EBC (breathprints) have been proposed to be potentially useful surrogate markers of inflammatory respiratory diseases. 15 In this mini-review, we identified several studies of exhaled breath analysis that focused on whether and how various types of diets served to those who are healthy or with disorders, such as epilepsies and gastrointestinal symptoms, can change the metabolic patterns in these groups of people (Table 2).
A study by Ruzsanyi et al. examined the ketogenic diets (KDs), which generally comprise high-fat, low-carbohydrate, and moderate-protein intakes, for their potential beneficial effects on seizures of children who were diagnosed with epilepsies. 84 To analyze VOCs’ profiles, the breath samples were collected after the participants were served with a modified Atkins diet (MAD), a classic KD, or a non-KD, and these breath VOCs were analyzed by GC-MS using a needle trap. In total, 18 VOCs were identified and 6 exhaled VOCs were considerably altered, including elevated levels of 2-methylfuran, 2-pentanone, acetaldehyde, acetone, and methyl-vinyl-ketone in KD groups, whereas the increased concentration of ethanol was observed in non-KD groups. 84 This clinical trial also found that KD-induced breath VOC changes, such as acetone, may be involved in β-oxidation. Taken together, this study suggested that the analysis of exhaled VOCs can be a promising technique for exploring a variety of potential biomarkers for dietary interventions. However, as summarized in Table 2, some studies which analyzed breath samples post food ingestions, such as sugar beet pectin and high fiber diets (HFDs), did not result in statistically significant changes in VOC profiles.84–86 For example, the impact of sugar beet pectin intervention on young and elderly adults in a randomized, double-blind, placebo-controlled parallel design was assessed to understand its role in healthy aging. 85 In total, 100 individuals randomly received either 15 g of sugar beet pectin (young [n = 25] and elder [n = 24]) or maltodextrin as placebo (young [n = 27] and elder [n = 24]) twice daily for four weeks. Then, exhaled breath samples were collected into a 3-L Tedlar bag to analyze VOC profiles using a GC-MS. 85 While eventually, no significant difference due to tested diets was reported, this study represented the effort of attempting to reduce aging-related comorbidities via dietary intervention and non-invasive, potentially high-compliance breath analysis. Similarly, seven healthy individuals with gastrointestinal symptoms were provided with 6–10 slices (20–25 g/slice) of 100% white wheat bread daily for a run-in period (4 weeks). Successively, they were served with 6–10 slices (25–30 g/slice) of sourdough whole grain rye bread and wheat bread with bioprocessed, fermented, rye bran daily for two weeks of each diet (a total of four weeks). The alveolar exhaled breath samples (150 mL) were collected and used for detecting signals of aspiration ion mobility spectrometry after whole-grain rye bread diet intervention in comparison with other diets. In contrast to the expected outcome, there was no significantly different effect of tested diets; however, an individual variation on the breath signals was observed even they were offered with the same diets. 85
In another study, Raninen et al. performed a pilot clinical trial that HFD (44 g/day) and low fiber diet (LFD; 17 g/day) were randomly assigned to seven healthy male subjects for a week. 86 At the end of the diet periods, alveolar exhaled breath samples were taken at fasting state and postprandially 30, 60, and 120 min. In total, 15 VOCs, including acetone, ethanol, 1-propanol, 2-propanol, 1-butanol, acetic acid, propionic acid, butyric acid, valeric acid, isovaleric acid, 2-methylbutyric acid, hexanoic acid, acetoin, diacetyl, and phenol were detected using solid-phase microextraction and GC-MS. Among the screened VOCs, relatively enhanced levels of ethanol, 1-propanol, acetoin, propionic acid, and butyric acid were found in HFD interventional trials as compared to LFD, but without statistical significance. 86 These pilot studies imply that exhaled breath analysis can be a suitable technique for examining diet-induced VOC profiles in fasting and postprandial state simply and efficiently.86,87 Despite a wide range of VOCs can be screened from exhaled breath samples, a significant difference between tested diets was not always observed, which could be due to various influencing factors, such as sample size, high inter-individual variation, or diet-linked metabolic background. Therefore, standardization of procedures, longitudinal studies, as well as external validation are needed to further develop breathomics into clinical tools with improved sensitivity, accuracy, and specificity.79,87,88 On the positive note, these exhaled breath analysis studies, while still at their early stage for potential clinic applications, underscored the promising potential of MS-based breathprints analysis as an early screening tool in studying physiological functionality of dietary components to human metabolism. 87
Conclusions and future directions
Analyses of plasma/fecal SCFAs and breath VOC profiles have been considered as promising and minimum-invasive/non-invasive tools for evaluating changes in human metabolism in response to a variety of dietary interventions in randomized clinical trials. The biological implications of these small molecular metabolites to the gut microbial metabolism and human metabolic network are a fascinating area of research, which highlighted the need for more comprehensive studies in the future. In this review, we investigated the association between diets and fecal SCFAs in patients with MetS or at high risk of MetS, and diet-induced changes of the gut microbiome and their production of SCFAs are also discussed. Some interesting clinical evidence that proved the usefulness of these metabolite biomarkers in reflecting the effectiveness of investigated dietary interventions was discovered in these microbe-gut-host health nexus studies. In addition, we highlighted the possibility of breath VOCs analysis to examine the dietary intervention effect on human metabolism. However, several factors, including the impact of food components, time, and dose-response of these dietary interventions to these metabolite biomarkers, and the response stability need to be further investigated between individuals with diverse genetic profiles.
Moreover, the use of many emerging MS-based analytical platforms, such as secondary electrospray ionization (SESI),26,89,90 proton transfer reaction (PTR), 91 or selected ion flow tube mass spectrometry. 92 have become popular recently. These breath analysis techniques have been reported to have various advantages in facilitating rapid data acquisition based on real-time and online analysis of VOCs. 93 Notably, PTR-MS and SESI-MS have gained attention based on the high sensitivity at trace levels of part per billion (ppb) or a detection limit of 0.2 parts per trillion (ppt), respectively.89,90,94 This merit enables the determination of potential biomarkers by fast detection and identification of microbial VOCs in the headspace of in vitro culture in non-invasive and spontaneously responsive manners.84,85 Therefore, these recently invented MS-based tools can be well-suited for breathomics analysis, and can be further considered for robust and reproducible analytical measurements in the nutrient intervention trials. Moving forward, we believe plasma/fecal SCFA analysis and human exhaled VOC analysis can be one of the most effective and powerful tools for exploring the impact of dietary intervention on responsive metabolic alteration towards the development of personalized nutrition.
Footnotes
AUTHORS’ CONTRIBUTIONS
Conceptualization, JZ; methodology, JZ and JHJL; formal analysis, JHJL; investigation, JZ and JHJL; resources, JZ; data curation, JHJL; writing—original draft preparation, JHJL; writing—review and editing, JZ; project administration, JZ; funding acquisition, JZ. All authors have read and agreed to the submitted version of the manuscript.
DECLARATION OF CONFLICTING INTERESTS
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
FUNDING
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Research reported in this publication was supported by the National Institute of General Medical Sciences of the National Institutes of Health under Award Number R35GM133510. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.