
Abstract
Chronic kidney disease is a major global public health problem. The peptide hormones adropin and spexin modulate many physiological functions such as energy balance and glucose, lipid and protein metabolism. However, it is unclear whether these peptides may exert effects on renal damage, tissue remodeling, and inflammatory conditions. In view of the limited information, we aimed to investigate the effect of adropin and spexin on matrix metalloproteinase and inflammatory response genes a rat model of adenine-induced chronic kidney failure. Chronic kidney failure was induced in rats by administering adenine hemisulfate. Renal function was determined in an autoanalyzer. Histopathological modifications were assessed by H&E staining. mRNA expression levels of ALOX 15, COX 1, COX 2, IL-1β, IL-10, IL-17A, IL-18 IL-21, IL-33, KIM-1, MMP-1, MMP-2, MMP-3, MMP-7, MMP-9, MMP-13, NGAL, TGFβ1, TIMP-1, and TNFα in kidney tissue were measured by qPCR. Our results showed an increase of 24-h urine volume, serum creatinine, BUN, and urine protein levels in group with adenine-induced CKF. Adropin and spexin treatments decreased urine protein and 24-h urine volume. Renal damage, TIMP-1, IL-33, and MMP-2 increased after CKF induction, while COX 1, MMP-9, and MMP-13 levels were significantly reduced. Furthermore, KIM-1, TIMP-1, IL-33, and MMP-2 were downregulated by spexin treatment. Renal damage, NGAL, TIMP-1 IL-17A, IL-33, MMP-2, and MMP-3 decreased after adropin treatment, while MMP-13 levels were upregulated. Treatment with adropin+spexin decreased KIM-1, NGAL, TIMP-1, IL-1β, IL-17A, IL-18, IL-33, ALOX 15, COX 1, COX 2, TGFβ1, TNFα, MMP-2, MMP-3, and MMP-7, but increased MMP-13 levels. Our findings revealed that inflammatory response and MMP genes were modulated by adropin and spexin. These peptides may have protective effects on inflammation and chronic kidney damage progression.
Impact statement
The peptide hormones adropin and spexin modulate many physiological functions such as energy balance and glucose, lipid and protein metabolism. These hormones may play a protective role in the pathogenesis of many diseases such as diabetes, diabetic nephropathy, chronic kidney disease, endothelial dysfunction, heart failure, and cardiovascular disease. However, it is unclear whether these peptides may exert effects on renal damage, tissue remodeling, and inflammatory conditions. Within this context, we investigated the protective effect of adropin and spexin on inflammation and MMPs in a rat model of chronic kidney failure. Our findings revealed that inflammatory response and MMP genes were modulated by adropin and spexin. These peptides may have protective effects on inflammation and chronic kidney damage progression.
Introduction
Chronic kidney failure (CKF) provides a suitable environment for the occurrence of many major diseases in chronic kidney disease (CKD) patients. The condition mostly causes or is caused by fibrosis in the kidneys and blood vessels setting the stage for other diseases. Renal fibrosis has been shown to be the most reliable predictor of the progression of CKD to end‐stage renal disease (ESRD). 1 There are two important processes that determine the course of chronic kidney failure: inflammation and fibrosis. The kidneys of CKF patients produce many inflammatory mediators due to the chronic inflammatory status, including tumor necrosis factor alpha (TNF-α), interleukin 1β (IL-1β), IL-6, IL-10, IL-17A, IL-18, IL-21, and IL-33.2–5 In addition, cyclooxygenases (COX) and arachidonate lipoxygenases (ALOX) play an important role in the development of CKD by affecting both the production of inflammatory mediators and renal functions.6,7
Kidney failure can be detected by few biomarkers such as kidney injury molecule-1 (KIM-1) and neutrophil gelatinase-associated lipocalin (NGAL). KIM-1 indicates proximal tubule injury and NGAL is upregulated and secreted in response to kidney injury.8,9 These two biomarkers are also independent risk factors for progression of CKD. Other important biomarkers, which have an impact in CKF patients, are matrix metalloproteinases (MMPs) and their tissue inhibitors (TIMPs). The former are zinc-dependent endopeptidase. At least 28 MMPs have so far been identified. 10 The family of MMPs is classified based on sequence homology and substrate specificity into six groups: collagenases (MMP-1, MMP-13), gelatinases (MMP-2, MMP-9), stromelysins (MMP-3, MMP-10), matrilysins (MMP-7, MMP-26), membrane-type MMPs, and other MMPs. 11 MMPs cleave many extracellular matrix (ECM) components such as collagens and elastins, and play key roles in tissue homeostasis and remodeling. 12 Besides their tissue-remodeling function, MMPs regulate many non-ECM molecules such as growth factors, cell surface receptors, adhesion molecules, cytokines, and chemokines. 13 The activity of MMPs is tightly regulated by their endogenous inhibitors, known as TIMPs. 14 Imbalances in MMP/TIMP levels have been linked to the structural changes that occur in the development of renal scarring, fibrosis, and inflammation. 15 All of these biomarkers are important for determining the effects of CKF treatments and for disease prognosis.
Spexin, a member of the spexin/galanin/kisspeptin family, has recently been identified as a novel peptide hormone by bioinformatic tools.16,17 The spexin peptide consists of 14 amino acids and a molecular weight of approximately 1.6 kDa.17,18 Spexin is broadly expressed in numerous tissues such as ovary, testis, heart, skeletal muscle, kidney, lung, liver, pancreas, brain, thyroid, adrenal gland, spleen, adipose tissue, stomach, and GI tract. 19 Spexin activates galanin receptor types 2 (GALR2) and 3 (GALR3) in target cells. 20 Thus, this peptide modulates many physiological functions such as feeding, energy balance, lipid storage, arterial blood pressure, salt-water balance, and cardiorenal responses.21–23 Recent studies also indicate that this hormone may modulate inflammatory processes, but this mechanism is unclear. 24
Adropin, is a bioactive peptide hormone recently discovered by Kumar, which has a molecular weight of approximately 4.49 kDa.25,26 This peptide is extensively expressed in numerous tissues such as the brain, heart, kidney, liver, pancreas, and GI tract.27,28 Adropin regulates energy balance and glucose, lipid and protein metabolism by activation of GPR19 in target cells.29,30 Several studies reported that low serum adropin levels may play a role in the pathogenesis of many diseases such as cardiovascular disease, CKD, diabetes, and diabetic nephropathy.31–33 Moreover, adropin induces anti-inflammatory responses and may therefore exert a protective effect against the pathogenesis of these diseases.31,34 However, few studies have investigated the effects of adropin on inflammatory status and MMPs.
Within this context, we aimed to investigate the effect of adropin and spexin on MMP and inflammatory response genes in a rat model of adenine-induced CKF. Our findings revealed that inflammatory response and MMP genes were modulated by adropin and spexin. These peptides may have protective effects on inflammation and chronic renal damage progression.
Materials and methods
Animals
Twenty-five male Sprague Dawley rats (200–250 g), supplied by the Trakya University Animal Center, were housed in an air-conditioned room under a 12-h light/dark cycle, at a relative humidity of 65–70% and constant temperature (22 ± 2°C). The rats had free access to food and water. All experimental procedures were approved by the Trakya University Animal Research and Ethics Committee, Edirne, Turkey (Protocol number: TUHADYEK-2016/52).
Experimental design
The animals (n = 25) were randomly divided into a vehicle group (n = 5) and a CKF (n = 20) group. The vehicle group received daily 5% carboxymethyl cellulose (1 mL/kg) by oral gavage for 10 days, followed by saline injected intraperitoneally for four weeks. The CKF group received daily adenine hemisulfate solution at a dose of 600 mg/kg and 5% carboxymethyl cellulose, 1 mL/kg (Sigma, cat no: A9126) by oral gavage for 10 days to induce CKF.35,36
After the administration of adenine hemisulfate, the induction of kidney failure was confirmed by biochemical parameters (day 15 of the experiment). Following biochemical confirmation, the CKF group (n = 20) was randomly divided into four groups: CKF (saline 1 mL/kg); adropin (2.1 µg/kg/mL, Phoenix Pharmaceuticals, cat no: 032–35); 37 spexin (35 µg/kg/mL, PolyPeptide, cat no: SC1547); 22 adropin+spexin (adropin; 2.1 µg/kg/mL, spexin; 35 µg/kg/mL). The groups were treated daily for four weeks, five days per week. All rats were then transferred to a metabolic cage for 24-h urine sampling without food or water restriction. Following sacrifice under anesthesia (100 mg/kg ketamine, 12.5 mg/kg xylazine, i.p.), blood was collected by cardiac puncture and kidney tissues were removed. During the experiment, 24-h urine samples were collected and urine volume was recorded in a metabolic cage five days after the adenine hemisulfate treatment (day 15 of the experiment) and at the end of the adropin and/or spexin treatments (4th week). Blood samples were collected into a serum clot activator tube, centrifuged at 2000g (10 min at 4°C), and serum was taken on day 15 and at the end of the 4th week of treatment. Kidney tissues of each animal were removed, snap-frozen in liquid nitrogen, and stored at −80°C for the qPCR assays. In addition, the kidney tissue samples were fixed in formaldehyde for pathological assessment.
Renal function tests
Serum creatinine and BUN levels and creatinine and protein levels in 24-h urine samples were determined in a Clinical Chemistry Analyzer (Architect C16000, Abbott Laboratories). The urine protein/creatinine ratio, protein loss, and creatinine clearance were also calculated.
Light microscopy examination of kidney tissue
Kidney tissue samples were fixed in 10% buffered formaldehyde, dehydrated in ethanol, incubated in xylol for 2 h, embedded in liquid paraffin, and cut into 5 μm thick sections onto glass slides with an automatic slicing machine (Leica). The sections were stained with hematoxylin and eosin and examined under a light microscope (Leica) at 200× magnification to evaluate pathological changes. The severity of tubular necrosis was graded as follows: (1) <10% necrosis; (2) 10%–25% necrosis; (3) 25%–50% necrosis; (4) 50%–75% necrosis, and (5) more than 75% necrosis.
Gene expression analysis
Kidney tissues (200 mg) were homogenized (IKA) and total RNA was isolated with the GeneJET RNA Purification Kit (Thermo Scientific). The concentration of total RNA in the samples was determined in a spectrophotometer (Multiskan Go μDrop, Thermo). cDNA was synthesized from 100 ng total RNA using the Maxima First Strand cDNA Synthesis Kit (Thermo Scientific) according to the manufacturer’s instructions. The cDNA was used for quantitative PCR (qPCR) using the Maxima SYBR Green qPCR Master Mix (Thermo Scientific) and the threshold cycle (CT) was determined with the PikoReal™ Real-Time PCR System (Thermo Scientific). Relative gene expression levels of ALOX15, COX-1, COX-2, IL-1β, IL-10, IL-17A, IL-18 IL-21, IL-33, KIM-1, MMP-1, MMP-2, MMP-3, MMP-7, MMP-9, MMP-13, NGAL, TGF-β1, TIMP-1, and TNF-α were calculated as a fold change using the 2−ΔΔCT method, where ΔCT = CT (target gene)-CT (housekeeping gene, β-actin) and Δ(ΔCT) = ΔCT (vehicle)-ΔCT (treatment). The primers used for qPCR are listed in Table 1. Estimated PCR product sizes and target gene specificity were confirmed using NCBI Primer-BLAST (http://www.ncbi.nlm.nih.gov/tools/primer-blast/).
Nucleotide sequences of forward (F) and reverse (R) primers, size of the products and primer specific annealing temperatures (Ta) for the real-time PCR detection.

Statistics
Differences between the groups were evaluated statistically by ANOVA and the Mann–Whitney U test as a post hoc test using the GraphPad Prism 6.0 software. A p value <0.05 was accepted to be statistically significant. Data are presented as mean±SD in the figures.
Results
Adropin and spexin attenuate renal function impairment
Our results showed that adenine solution induced CKF, characterized especially by polyuria and proteinuria, in all rats. On day 15 of the experiment, serum creatinine (V: 0.20 ± 0 mg/mL; CKF:1.0 ± 0.32 mg/mL, p < 0.001) and BUN (V: 39.07 ± 3.10 mg/mL; CKF: 298.4 ± 66.91 mg/mL, p < 0.001) levels were increased in the CKF group compared to the vehicle group. In addition, adenine induction increased the 24-h urine volume (V: 9.35 ± 3.60 mL; CKF: 45.25 ± 10.2 mL, p < 0.001). Protein loss in 24-h urine was significantly increased by CKF induction (V: 2873 ± 1933 mg/mL; CKF: 6187 ± 1479 mg/mL, p < 0.05). We also calculated the protein/creatinine ratio to show the metabolic breakdown and renal damage and observed that this ratio increased significantly with renal damage (CKF: 21.16 ± 6.8; V: 3.90 ± 0.64; r = 0.688; p < 0.01). Urine creatinine levels were lower in the CKF groups probably due to polyuria. Adropin or spexin treatment did not cause any significant change in serum creatinine, BUN, or creatinine clearance levels, while adropin treatment reduced 24-h urine output (CKF: 50.17 ± 10.89; CKF+A: 27.0 ± 7.55, p < 0.05) compared to the CKF group. Spexin exerted lower effects on urine creatinine. Additionally, urine creatinine, urine protein, and daily protein loss were decreased by adropin+spexin treatment compared to CKF (Table 2).
Effects of adropin and spexin on kidney function in adenine-induced CKF.
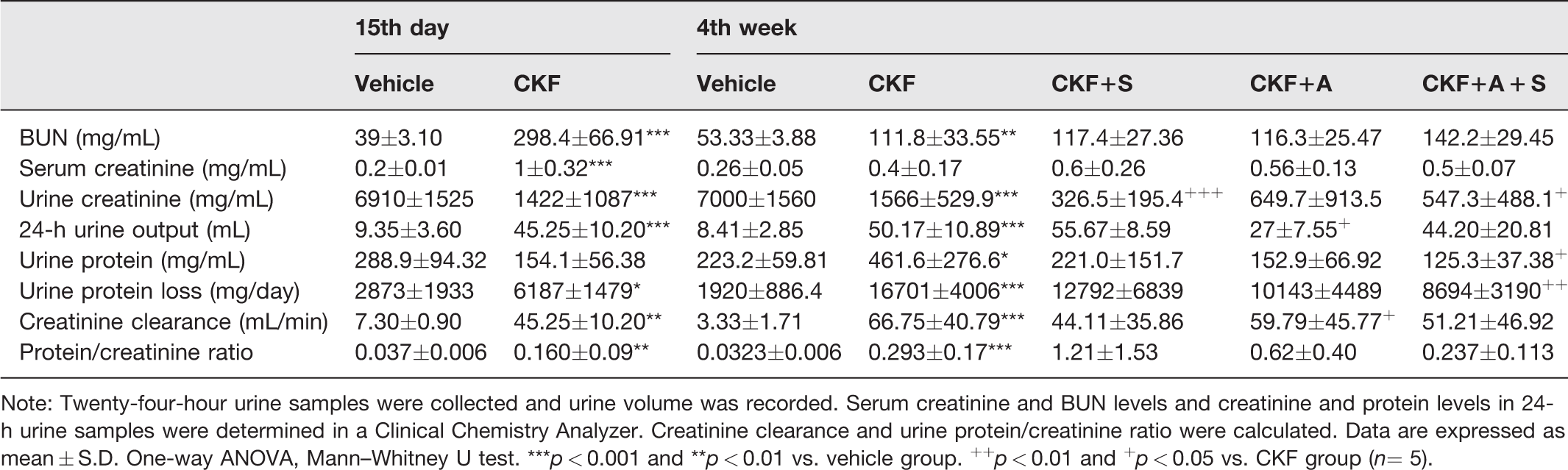
Note: Twenty-four-hour urine samples were collected and urine volume was recorded. Serum creatinine and BUN levels and creatinine and protein levels in 24-h urine samples were determined in a Clinical Chemistry Analyzer. Creatinine clearance and urine protein/creatinine ratio were calculated. Data are expressed as mean ± S.D. One-way ANOVA, Mann–Whitney U test. ***p < 0.001 and **p < 0.01 vs. vehicle group. ++p < 0.01 and +p < 0.05 vs. CKF group (n= 5).
Spexin and adropin have a protective effect on adenine-induced kidney damage
The macroscopic assessment of the kidney focused on surface color and shape. The macroscopic picture of the kidneys (Figure 1) showed a color change from red-brown to white-brown in the CKF groups compared to the vehicle groups. In addition, the surface of the kidney was not smooth, indicating an abnormal kidney.
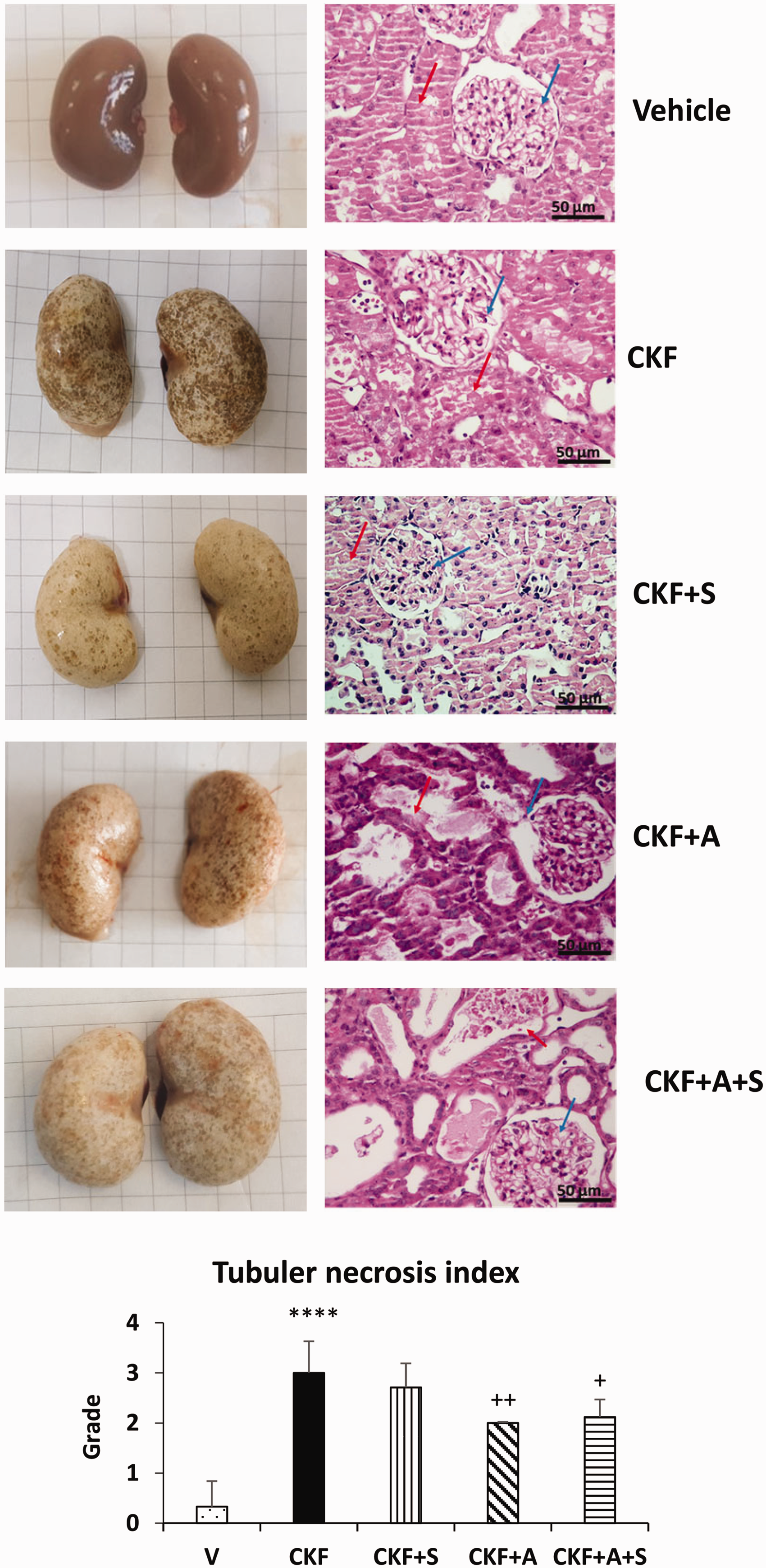
Effects of adropin and spexin on macroscopic appearance, histopathologic changes, and tubular necrosis in a model of adenine-induced CKF. Renal histopathological changes and tubular necrosis were examined by H&E staining. The red arrow indicates a glomerular area and the blue arrow indicates tubular necrosis. Photographs taken at 200× magnification (scale bar = 50 μm). Data are expressed as mean ± S.D. One-way ANOVA, Mann–Whitney U test. ****p < 0.0001 vs. vehicle group. ++P < 0.01 vs. CKF group (n= 5). (A color version of this figure is available in the online journal.)
Histopathological evaluation of the kidney tissues was performed by H&E staining (Figure 1). In the vehicle group, the kidneys displayed a normal renal tissue structure, uniform tubules lined with a single epithelial layer, and no obvious pathological changes. The kidney tissue in the CKF group exhibited degenerative changes such as glomerular bulging, glomerular hypertrophy, tubular dilatation, and tubular necrosis. Adropin and adropin+spexin treatments significantly attenuated the severity of kidney damage in the CKF group. Quantitative histopathology showed a significantly higher tubular necrotic index in rats of the CKF group compared to the vehicle group; conversely, adropin and combined adropin+spexin treatments significantly reduced this elevated tubular necrotic index in CKF rats. The pathological changes in kidney tissue of the CKF+S group were similar to the CKF group.
Additionally, for confirmation of the renal pathological condition, mRNA expression levels of the kidney injury markers KIM-1, NGAL, and TIMP-1 were assessed by qPCR (Figure 2). We observed that mRNA expression of TIMP-1 was significantly increased (7 fold), while mRNA expression levels of KIM-1 and NGAL remained unchanged in the CKF group compared to vehicle groups. Also, KIM-1 and TIMP-1 were downregulated by CKF+S compared to CKF, but NGAL was unchanged. In addition, NGAL and TIMP-1 were downregulated by CKF+A compared to CKF, while KIM-1 was unchanged. Moreover, mRNA expression levels of KIM-1, NGAL and TIMP-1 were downregulated by combined adropin+spexin treatment.
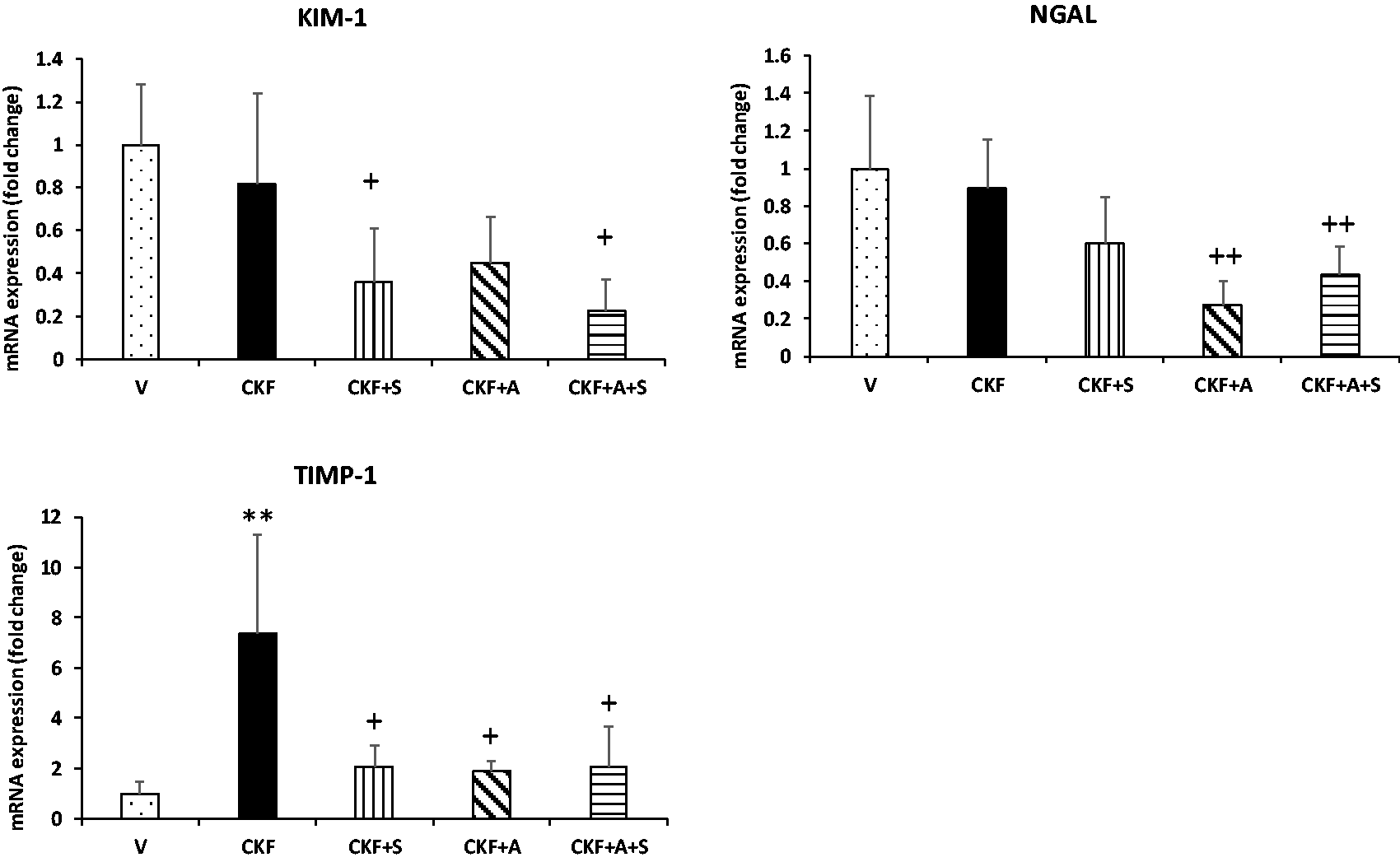
Effects of adropin and spexin on markers of kidney damage in CKF. mRNA expression levels of KIM-1, NGAL, and TIMP-1 in kidney tissue were measured by qRT-PCR and normalized to β-actin. Data are expressed as mean ± S.D. One-way ANOVA, Mann–Whitney U test. **p < 0.01 vs. vehicle group. ++p < 0.01 and +p < 0.05 vs. CKF group (n= 5).
Spexin and adropin promote an anti-inflammatory response
Following the determination of chronic kidney failure status, we evaluated alterations in the inflammatory response of kidney tissue. For this purpose, mRNA expression levels of interleukin, proinflammatory, and anti-inflammatory genes were determined by qPCR. As shown in Figures 3 and 4, IL-33 was significantly increased in the CKF group compared to the vehicle group, although COX-1 was significantly reduced. In addition, spexin treatment significantly decreased IL-33 gene expression compared to the CKF group. Adropin treatment also decreased IL-17A and IL-33 gene expression levels compared to the CKF groups. Additionally, mRNA expression levels of the IL-1β, IL-17A, IL-18, IL-33, ALOX15, COX-1, COX-2, TGF-β1, and TNF-α genes were significantly downregulated by combined adropin+spexin treatment compared to CKF.
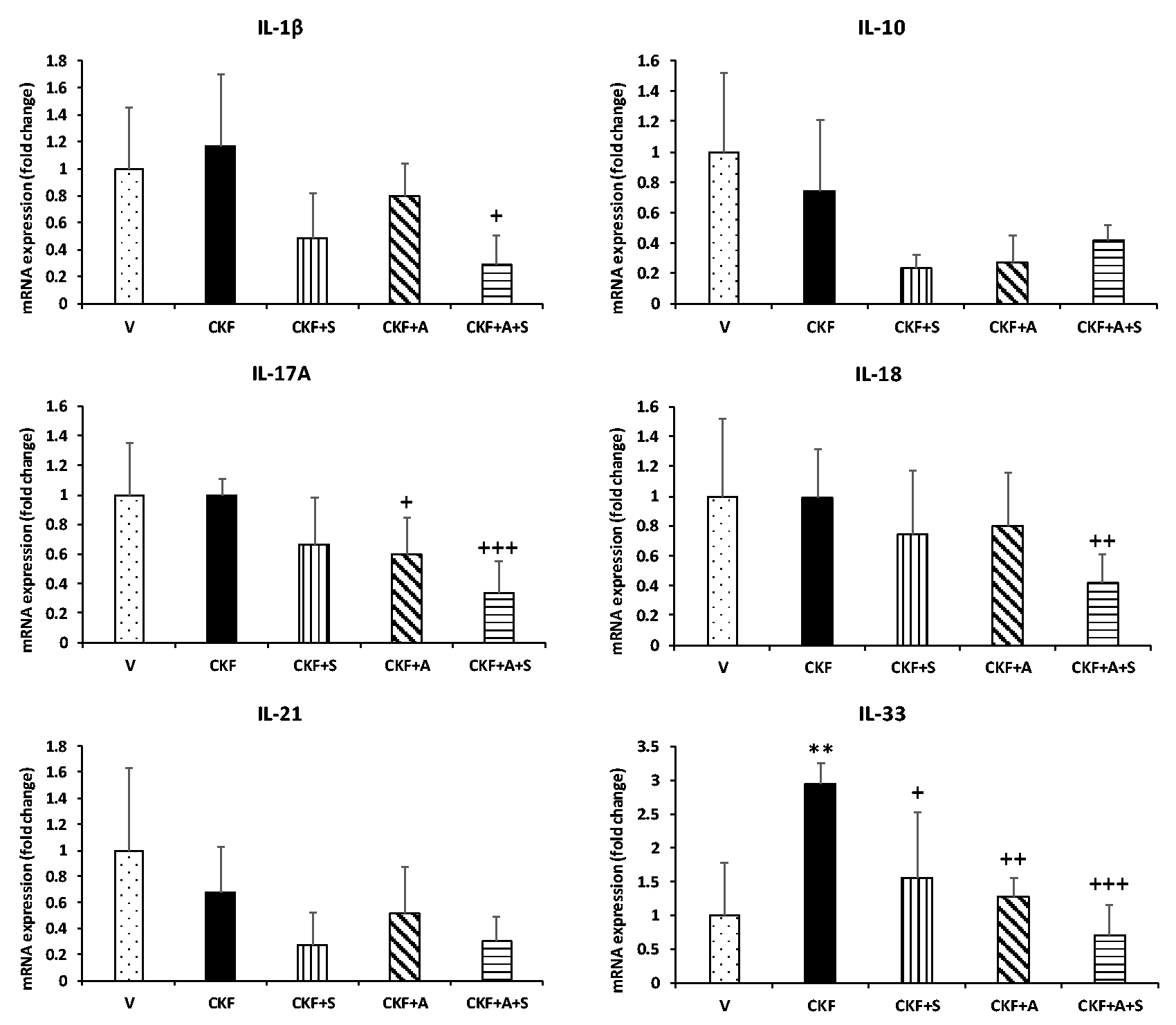
Effects of adropin and spexin on the expression of interleukin genes in CKF. mRNA expression levels of IL-1β, IL-10, IL-17A, IL-18, IL-21, and IL-33 in kidney tissue were measured by qRT-PCR and normalized to β-actin. Data are expressed as mean ± S.D. One-way ANOVA, Mann–Whitney U test. **p < 0.01 vs. vehicle group. +++p < 0.001, ++p < 0.01, and +p < 0.05 vs. CKF group (n= 5).
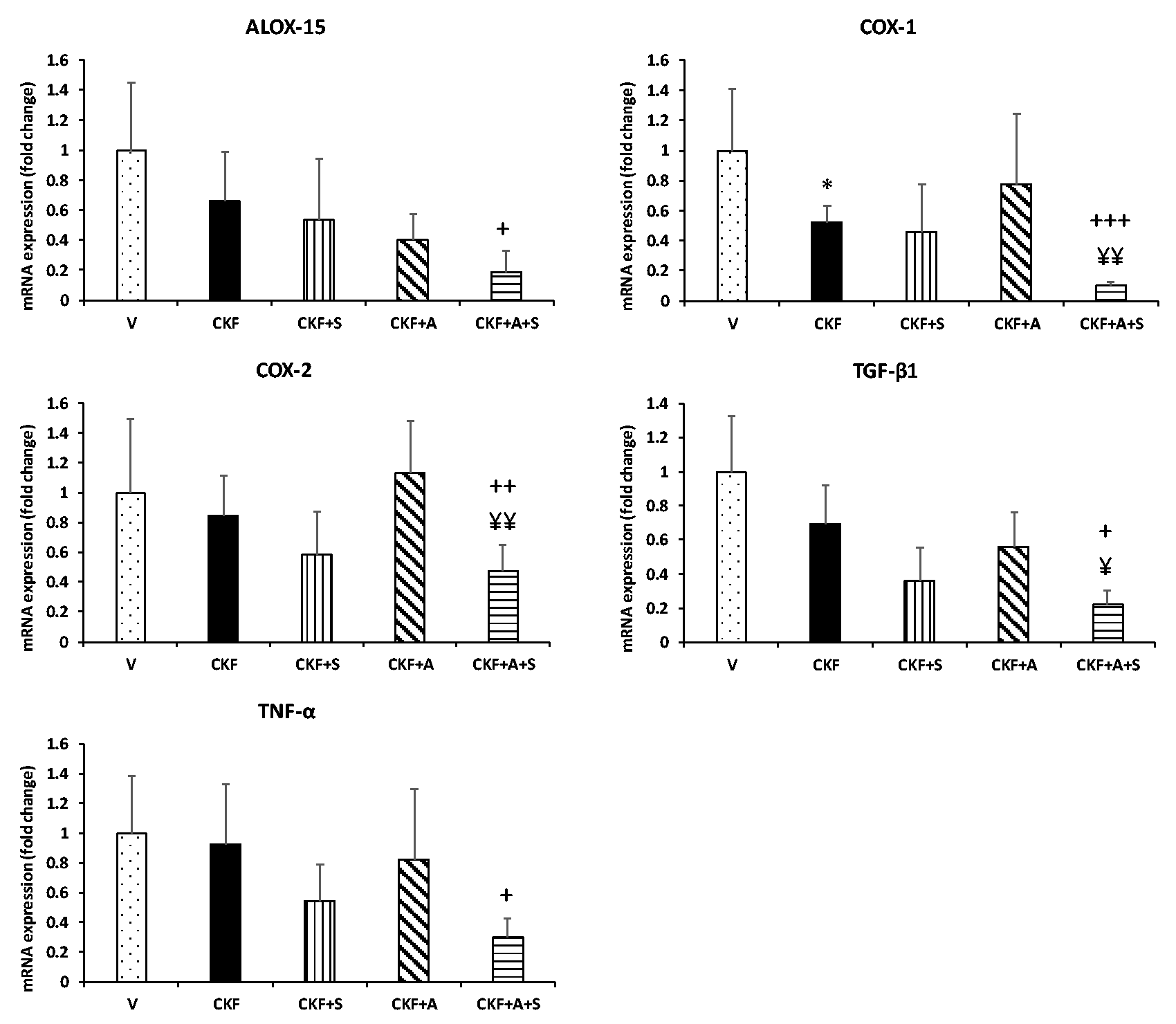
Adropin and spexin alter proinflammatory and anti-inflammatory genes in CKF. mRNA expression levels of ALOX15, COX-1, COX-2, TGF-β1, and TNF-α in kidney tissue were measured by qRT-PCR and normalized to β-actin. Data are expressed as mean ± S.D. One-way ANOVA, Mann–Whitney U test. *p < 0.05 vs. vehicle group. +++p < 0.001, ++p < 0.01, and +p < 0.05 vs. CKF group. ¥¥p < 0.01 and ¥p < 0.05 vs. adropin (n= 5).
Spexin and adropin treatments significantly modulate MMP gene expression levels
The effect of adenine-induced CKF development on the MMP-1, MMP-2, MMP-3, MMP-7, MMP-9, and MMP-13 genes that regulate tissue regeneration was determined by qPCR. As shown in Figure 5, our data indicate a marked increase in the mRNA expression of MMP-2 in the CKF group compared to the vehicle group; however, MMP-9 and MMP-13 levels were significantly reduced. On the other hand, no difference in the gene expression of MMP-1, MMP-3, or MMP-7 was observed between the CKF and vehicle groups. MMP-2 gene expression was significantly reduced by spexin treatment compared to the CKF group. Also, adropin treatment significantly downregulated MMP-2 and MMP-3 gene expression levels compared to the CKF group; conversely, MMP-13 levels were upregulated by adropin treatment. Moreover, combined adropin+spexin treatment significantly decreased MMP-1, MMP-2, MMP-3, and MMP-7 levels compared to CKF, but increased MMP-13 levels.
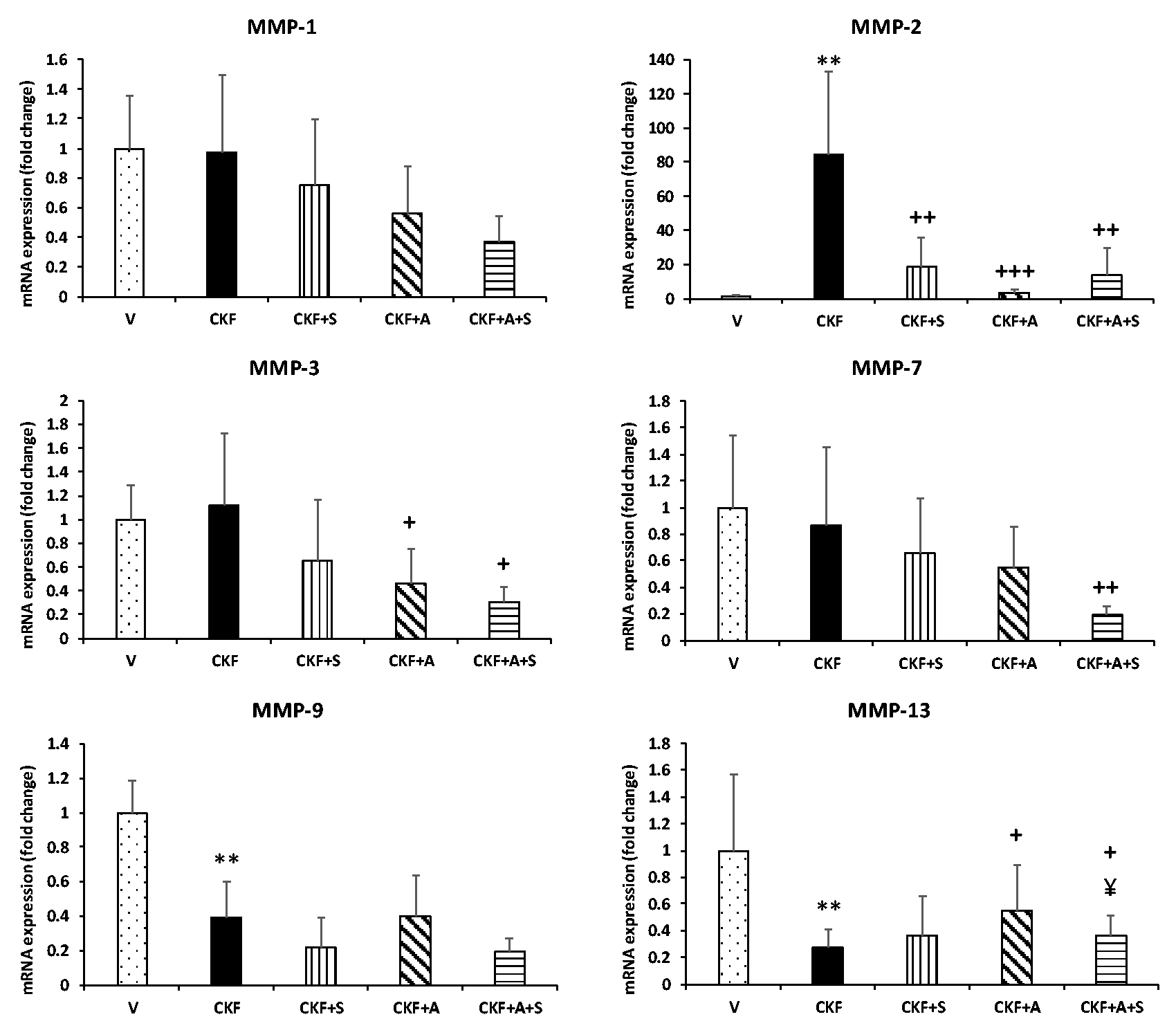
Adropin and spexin regulate MMP genes in CKF. mRNA expression levels of MMP-1, MMP-2, MMP-3, MMP-7, MMP-9, and MMP-13 in kidney tissue were measured by qRT-PCR and normalized to β-actin. Data are expressed as mean ± S.D. One-way ANOVA, Mann–Whitney U test. **p < 0.01 vs. vehicle group. +++p < 0.001, ++p < 0.01, and +p < 0.05 vs. CKF group. ¥p < 0.05 vs. adropin (n= 5).
Discussion
Chronic kidney failure is a chronic progressive renal loss of parenchymal function that is associated with a poor prognosis and high mortality. 38 According to WHO data from 2015, 1.2 million people have died from kidney failure. The disease poses a heavy economic burden on families and society worldwide. CKF including renal tubular cell damage and death is the most common cause of CKD in hospitalized patients.39,40 Studies using rat models of CKF have shown that adenine hemisulfate solution significantly increases BUN, serum creatinine, glomerular damage, tubular necrosis, interstitial fibrosis, and inflammatory mediators.35,41,42 In addition, many inflammatory markers such as IL-1β, IL-6, IL-18, IL-33, and TNF-α increase during the progression of CKF.43,44 Numerous studies involving CKD populations and disease-specific subgroups have shown associations between higher urinary and blood NGAL levels and the progression of kidney disease. 45 In both mice and rats, the increase in plasma KIM-1 levels directly reflected the degree of histologic and functional kidney injury. 8
Within this context, we aimed to investigate the effect of adropin and spexin hormones on MMP and inflammatory response genes in a rat model of CKF. Chronic kidney failure was induced in rats by administering adenine hemisulfate solution. Our kidney function test results (Table 2) showed that BUN and serum creatinine levels significantly increased after the administration of adenine solution, as demonstrated in previous studies on adenine-induced CKF. We also found that protein excretion significantly increased, while creatinine levels in 24-h urine samples significantly decreased in the adenine-induced kidney failure group. We calculated the protein/creatinine ratio to demonstrate the metabolic breakdown and renal damage and we observed a significant increase in this ratio with renal damage. In addition, macroscopic observation showed a change in kidney color from red-brown to white-brown in the CKF groups compared to the vehicle groups. Also, our light microscopy findings indicated degenerative changes such as glomerular bulging, glomerular hypertrophy, tubular dilatation, and tubular necrosis in the CKF group (Figure 1). Besides, the tubular necrotic index of rats in the CKF group was significantly greater compared to the vehicle group. For confirmation of the renal histopathological condition, we measured the gene expression levels of the kidney injury markers KIM-1, NGAL, and TIMP-1 (Figure 2). TIMP-1 levels were significantly increased (7 fold), while KIM-1 and NGAL levels remained unchanged in the CKF groups compared to the vehicle groups. Following the determination of CKF status, we evaluated alterations in the inflammatory response of kidney tissue (Figures 3 and 4). Our findings indicated that IL-33 was significantly increased in the CKF group compared to the vehicle group, although COX-1 was significantly decreased by CKF. No difference in the expression levels of the other genes was observed between the CKF and vehicle groups.
MMPs are zinc-dependent endopeptidase. 10 These enzymes cleave many ECM components such as collagens and elastins and play key roles in tissue homeostasis and remodeling. 12 Imbalances in MMP/TIMP levels have been linked to the structural changes that occur in the development of renal scarring, fibrosis, and inflammation. 15 Especially the expression of MMP-2 and MMP-9 is important in CKD.11,46 Many studies reported an association between progressive kidney fibrosis and increased serum MMP-2 concentrations in CKD patients.47–49 Altemtam et al. 50 showed that chronic nephropathy increases urinary MMP-2 excretion. Moreover, Tveitarås et al. 51 found that MMP-2 KO mice are protected from kidney fibrosis. MMP-9 plays an important role in development of CKD. According to some studies, the activity of MMP-9 is increased in the early stage of CKD, but decreases in the advanced stage of CKD. 11 Some studies observed that glomerular expression of MMP-1 and MMP-3 decreased in diabetic rats. MMP-1 expression was also reduced in alloxan induced-diabetic mice, whereas local application of MMP-1 into the kidneys of mice prevented diabetic renal fibrosis.52–54 Ren et al. 55 showed that Twist1 in infiltrating inflammatory macrophages enhances kidney fibrosis via downregulation of MMP-13; upregulation of MMP-13 also reduces fibrotic progression. However, another study on mice suggests that poricoic acid, which is an MMP-13 inhibitor, ameliorates renal fibrosis by suppressing MMP-13 activity. 56
Our results indicated a marked increase in the mRNA expression of MMP-2 in the CKF group compared to the vehicle group; however, MMP-9 and MMP-13 levels were significantly reduced by CKF. On the other hand, no difference in the gene expression of MMP-1 or MMP-3 was observed between the CKF and vehicle groups (Figure 5).
The peptide hormone spexin regulates many physiological pathways, including feeding, energy balance, lipid storage, arterial blood pressure, salt-water balance, and cardiorenal responses.21,22 Toll et al. 23 showed that intracerebroventricular injection of spexin peptide decreased urine flow rate. Spexin has recently been shown to influence the inflammatory response. Kumar et al. found that spexin levels decrease with obesity progression and that this peptide is negatively correlated with IL-6. 57 Similarly, in another study on obesity progression, spexin levels were found to be negatively correlated with hs-CRP. 58 Behrooz et al. showed that some serum inflammatory markers such as IL-10, IL-1β, and hs-CRP were closely associated with spexin levels in obese children. 59 Furthermore, Gambaro et al. 24 observed that the inflammatory markers IL-6, TNF-α, and IL-1β decreased after spexin administration in obese mice. However, the number of studies investigating the effects of spexin on kidney failure and inflammatory response is limited.
Our results revealed that spexin treatment exerts only reduced effects on urine creatinine in renal function tests. In addition, the kidney tissue in the CKF+S group showed a macroscopic appearance and pathological changes similar to those of the CKF group. Also, KIM-1 and TIMP-1 were downregulated by spexin treatment compared to CKF, but NGAL was unchanged. Moreover, IL-33 gene expression was significantly decreased by spexin treatment compared to the CKF group. Spexin treatment also significantly decreased MMP-2 gene expression compared to the CKF group.
Adropin regulates energy balance and glucose, lipid and protein metabolism pathways by activation of GPR19 receptors in target cells. Several studies suggested that this hormone may exert a protective effect against the pathogenesis of many diseases such as diabetes, diabetic nephropathy, CKD, and heart failure.29,31–33 Some studies have shown that serum adropin levels are negatively correlated with BUN and serum creatinine and a positive correlation with eGFR was reported.31,60 Besides, adropin may alter inflammatory responses.27,37,61 Sato et al. 62 showed that adropin treatment suppressed TNF-α induced THP1 monocyte adhesion to human umbilical vein endothelial cells (HUVECs), indicating the anti-inflammatory effect of this peptide. Ibrahim et al. 63 found that the inflammatory response markers IL-6 and TNF-α were negatively correlated with serum adropin levels in a diabetic nephropathic rat model. Furthermore, Gao et al. 64 showed that adropin deficiency in AdrKO mice increased the production of IL-1α, IL-1β, IL-6, IL-33, and TNF-α. In addition, adropin administration to HUVECs and human aortic smooth muscle cells (HASMCs) significantly increased the protein expression of fibronectin and elastin but not of collagen 1, collagen 3, MMP-2, or MMP-9. 62
Our findings indicate that adropin treatment reduced 24-h urine output compared to the CKF group. Besides, adropin treatment significantly reduced the elevated tubular necrotic index compared to CKF rats. NGAL and TIMP-1 were downregulated by CKF+A compared to CKF, while KIM-1 was unchanged. Our findings show that adropin treatment significantly attenuated the severity of renal injury in CKF. Furthermore, adropin treatment decreased IL-17A and IL-33 gene expression levels compared to the CKF groups. MMP-2 and MMP-3 gene expression levels were also significantly downregulated by adropin treatment compared to the CKF group; conversely, MMP-13 levels were upregulated by this treatment.
Combined adropin+spexin treatment reduced urine creatinine and urine protein loss compared to the CKF group. Similarly, adropin+spexin treatment significantly attenuated the severity of renal injury and tubular necrotic index in the CKF group. Also, mRNA expression levels of KIM-1, NGAL, and TIMP-1 were downregulated by combined adropin+spexin treatment. Moreover, combined adropin+spexin treatment significantly downregulated IL-1β, IL-17A, IL-18, IL-33, ALOX15, COX-1, COX-2, TGF-β1, and TNF-α expression levels compared to CKF. Furthermore, MMP-2, MMP-3, and MMP-7 levels were significantly decreased by combined adropin+spexin treatment compared to CKF; conversely, MMP-13 levels were increased by this treatment.
In conclusion, our results indicate a protective role of adropin and adropin+spexin in CKF, modulating renal damage, MMPs, and inflammatory responses. However, spexin exerts a limited effect on CKF. Further studies on these peptides are needed.
Footnotes
AUTHORS’ CONTRIBUTIONS
BY and GM designed the study, analyzed the data, and wrote the article; GM developed the model of kidney failure in rats; BY and FA performed the qPCR experiments; ET performed the histopathological assessments. All authors critically revised the manuscript and approved the final version of the submitted manuscript.
ACKNOWLEDGEMENTS
The authors would like to thank Amasya University Central Research Laboratory (AUMAULAB) for allowing the use of their facilities.
DECLARATION OF CONFLICTING INTERESTS
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
FUNDING
This work was supported by grants from Amasya University [grant numbers: FMB-BAP 19–0387 and FMB-BAP 20–0444] and Trakya University [grant number: 2018-118].