
Abstract
Aldehyde dehydrogenase 2 plays a pivotal role in detoxifying aldehydes, and our previous study revealed that aldehyde dehydrogenase 2 could alleviate diabetic retinopathy-associated damage. We aimed to characterize the potential role of aldehyde dehydrogenase 2 in diabetic keratopathy. Twenty-four rats with streptozotocin-induced (60 mg/kg, single intraperitoneal injection) type 1 diabetes mellitus (T1DM) were divided the T1DM group and the T1DM + Alda1 (an activator of aldehyde dehydrogenase 2) group (5 mg/kg/d, intraperitoneal injection, 1/2/3 months), while an additional 12 healthy rats served as the control group. Corneal morphology was examined in vivo and in vitro at one, two, and three months after T1DM induction. Additionally, serum inflammatory factors were measured by ELISA, and the expression of corneal vascular endothelial growth factor A (VEGF-A) and aldehyde dehydrogenase 2 was measured by immunofluorescence staining. Corneal cell death was evaluated by terminal-deoxynucleotidyl transferase-mediated nick end labeling (TUNEL) staining. Slit lamp analysis showed that the area of corneal epithelial cell injury in the T1DM + Alda1 group was significantly smaller than that in the T1DM group at one and two months after T1DM induction (all P < 0.05). OCT analysis and HE staining showed that the central corneal thickness (indication of corneal edema) and the epithelial keratinization level in the T1DM + Alda1 group was evidently decreased compared with those in the T1DM group (all P < 0.05). The serum inflammatory factors interleukin-1 and interleukin-6 were significantly upregulated in the T1DM group compared with the T1DM + Alda1 group at three months after T1DM induction (all P < 0.05), while there were no differences in SOD or TNF-α levels among all groups. Furthermore, corneal VEGF-A expression and corneal cell death in the T1DM + Alda1 group were dramatically reduced compared to those in the T1DM group (all P < 0.05). In conclusion, the aldehyde dehydrogenase 2 agonist Alda1 attenuated rat corneal dysfunction induced by T1DM by alleviating corneal edema, decreasing corneal cell death, and downregulating corneal VEGF-A expression.
Impact statement
The present study showed that ALDH2 could protect against diabetic keratopathy (DK). This work could be essential, as DK is one of the most common sight-threatening ophthalmological complications associated with diabetes mellitus (DM), and current clinical therapies for DK show limited effectiveness. STZ-induced type I diabetes mellitus caused various pathological changes in all rat corneal layers, including the epithelial layer, matrix layer, and endothelial layer. Remarkably, activating ALDH2 with the specific activator Alda1 could preserve corneal structural integrity, alleviate corneal edema, and delay DK progression. Intriguingly, the results further showed that the underlying molecular mechanism of ALDH2 involved alleviating the serum inflammatory response, reducing corneal VEGF-A expression, and decreasing apoptosis (or cell death). Our research findings are the first to support a protective role of ALDH2 in T1DM-induced corneal injury, suggesting that ALDH2 may be a good therapeutic candidate/target for DK treatment.
Introduction
Diabetes mellitus (DM) is a common chronic metabolic disorder that has drawn increasing global health attention. According to data released by the International Diabetes Federation (IDF), nearly 463 million people suffer from DM worldwide. 1 Although great progress in DM prevention and treatment has been achieved, the excessive risk of complications among diabetic patients remains a difficult problem for clinicians. 2
Diabetic retinopathy (DR), glaucoma, cataracts, and keratopathy are the most frequent sight-threatening DM-related ocular comorbidities. While much attention has focused on DR, the impact of diabetic corneal injury has seldom been explored. In fact, epidemiological investigations showed that approximately 70% of patients suffer from diabetic keratopathy (DK) during the process of DM.3–5 Recently, studies have demonstrated that corneal disorders could have predictive value for the systemic progression of DM.6–8
The cornea is covered with unmyelinated nerve endings at the epithelium and is sensitive to the environment. Interestingly, the cornea is avascular and acquires oxygen mainly from the air and mineral substances from the aqueous humor. 9 This context contributes to the role of the cornea as a smooth optical surface and a superb diffusion barrier to prevent water-soluble substance penetration. However, the unique corneal structure is a major site of damage in chronic hyperglycemia conditions. Microstructural disorders in DK include epithelial basement membrane (EBM) thickening, hemidesmosome decreases, and advanced glycation end product (AGE) accumulation, which lead to epithelial cell proliferation, recurrent corneal erosion, and corneal basal cell degeneration.10,11 Therefore, hyperglycemia significantly lowers epithelial function, resulting in the breakdown of barrier function, decreased sensitivity and abnormal wound healing. Furthermore, clinical observations indicated that surgical procedures for diabetic eyes were accompanied by long-lasting epithelial erosion with poor healing of the epithelial defects.12–14
Accumulating evidence has shown that DK is closely associated with a prolonged hyperglycemic state. Specifically, chronic hyperglycemia induces the overexpression of various cytokines, chemokines, and other proinflammatory proteins and proapoptotic genes, ultimately leading to the initiation and progression of DK. 8 Inflammatory factors, such as tumor necrosis factor-α (TNF-α), interleukin (IL), and vascular endothelial growth factor (VEGF), can lead to poor corneal functional and morphological changes. Oxidative stress induced by mitochondrial dysfunction and reactive oxygen species (ROS) accumulation also contributes to the decline in corneal epithelial cell density and the loss of epithelial function, which are involved in the pathogenesis of DK.10,15,16
Mitochondrial aldehyde dehydrogenase 2 (ALDH2) is an enzyme that is responsible for detoxifying noxious reactive aldehydes and lipid peroxidation-produced alkanols that plays a unique role in pathophysiological processes. 17 Numerous clinical and experimental findings have proven the favorable effects of ALDH2 on cancer, metabolic diseases, neurodegenerative diseases, and cardiovascular diseases.18–26 Our previous work revealed that ALDH2 functioned as a potent protector against DM-induced retinal disorders by decreasing inflammatory factor expression and activating the antioxidant stress response. 27 Moreover, our recent study showed that ALDH2 could protect retinal function and morphology in rats with MNU-induced retinitis pigmentosa. 28 This evidence supports a beneficial role of ALDH2 in endothelial dysfunction, angiogenesis, neurodegeneration, and energy metabolism disturbance.
Accordingly, it is worthwhile to investigate whether activating ALDH2 with the specific activator Alda1 (which stimulates acetaldehyde oxidation of ALDH2 by enhancing NAD binding) could improve diabetic corneal disorders caused by streptozotocin (STZ)-induced T1DM and further elucidate the role of ALDH2 in mediating inflammation, oxidation, and apoptosis (or cell death).
Materials and methods
Animals
Thirty-six male Sprague-Dawley rats (two to three months old, 180–220 g) were obtained from the Animal Center of the Fourth Military Medical University (license no. 2014270138S). The rats were analyzed and housed in accordance with a previous study. 29 All experimental protocols were approved by the Ethics Committee of the Animal Care and Experimental Committee of the Fourth Military Medical University. All experiments were performed in accordance with the Association for Research in Vision and Ophthalmology (ARVO) statements for ophthalmic research animal use. Lethal sodium pentobarbital (Sigma, St Louis, MO, USA) was used to euthanize the animals at one, two, and three months after DM induction.
Diabetic rat model
A diabetic rat model was established after a 10-day acclimatization period. Briefly, 24 rats were intraperitoneally injected with a single dose of streptozotocin (STZ, 60 mg/kg body weight) (Sigma, USA) to establish a type I diabetes mellitus (T1DM) rat model, and high-fat and high-calorie chow 27 was provided during the study. Then, the blood glucose levels were measured as previously described 72 h after STZ injection. 27 All of the rats (24/24) exhibited a random blood glucose level above 16.7 mmol/L, which was considered T1DM, and these rats were used in the study. Then, diabetic rats were divided into two groups: the DM + Alda1 group (5 mg/kg body weight Alda1, once daily, 50% DMSO/50% H2O) (MedChemExpress, USA) was administered an intraperitoneal injection of Alda1 for one, two, and three months after DM induction (n = 12); and the DM group received an equal volume of the vehicle (50% DMSO/50% H2O) at the same times (n = 12).
Slit lamp and OCT detection
Slit lamp detection and OCT tests were used at one, two, and three months after T1DM induction (n = 12, 8 and 4 rats). Slit lamp examinations were performed under dim conditions as previously described. 29 OCT examinations were performed in compliance with the operator manual as described previously. 29 OCT image analysis software was used to calculate corneal thickness.
HE staining
The morphological alterations in diabetic corneas were visualized by hematoxylin eosin (HE) staining at one, two, and three months after T1DM induction (n = 4 rats). Briefly, rats were euthanized by a lethal administration of sodium pentobarbital, and the eyeballs were quickly removed and enucleated. Corneal samples were fixed in 4% paraformaldehyde, embedded in paraffin, and sectioned along with the central cornea at a thickness of 4 μm. Then, routine HE staining was conducted as previously described.30,31
Proinflammatory parameter detection
Rat blood was collected via the abdominal aortic method at three months after T1DM induction (n = 4 rats). The serum concentrations of IL-1, IL-6, SOD, and TNF-α were measured with enzyme-linked immunosorbent assay (ELISA) kits from Sangong (Shanghai, China) according to the manufacturer’s instructions.
Immunofluorescence staining
Immunofluorescence staining was performed at one, two, and three months after T1DM induction (n = 4 rats) as previously described. 31 Anti-ALDH2 (Abcam, ab108306, USA) and anti-VEGF-A (GeneTex, GTX102643, USA) primary antibodies diluted 1:100 were used. Then, FitC-conjugated IgG (H + L fluorescent secondary antibodies (Zhuangzhi, EK023, China) were used at a dilution of 1:200. Finally, 4ʹ,6-diamidino-2-phenylindole (DAPI) was used to stain the nuclei. Images were obtained with a fluorescence microscope. The percentage of positive fluorescence signals (Area %) was measured using ImageJ software and normalized to the value of the control (CON) group.
TUNEL staining
Apoptosis levels were examined under different conditions using the TdT-mediated dUTP nick-end labeling (TUNEL) technique (cell death detection kit; Roche), which labeled the cut ends of DNA fragments in the nuclei of apoptotic (or necrotic) cells. The nuclei of corneal cells were stained with DAPI (100 ng/mL). The fluorescence intensity of the apoptotic signal was quantified by ImageJ software.
Statistical analysis
The differences among groups were analyzed with one-way analysis of variance (ANOVA) followed by Bonferroni’s post hoc analysis unless otherwise specified. The values are shown as the mean ± standard deviation (SD), and the level of significance was defined as a P-value ≤ 0.05.
Results
DM rat model
Diabetes was induced in rats by STZ and a high-fat and high-calorie diet. Twelve diabetic rats were treated with (T1DM + Alda1 group) or without (T1DM group) Alda1. Another 12 age- and weight-matched healthy male rats served as the control (CON) group. In the final experiment (three months after T1DM induction), the levels of blood glucose and insulin were measured. As shown in Figure 1, the levels of blood glucose in the T1DM and T1DM + Alda1 groups were 25.14 ± 1.50 mmol/L and 23.70 ± 1.71 mmol/L, respectively, which were significantly higher than those in the CON group (6.63 ± 0.58 mmol/L). Moreover, the levels of blood insulin in the T1DM and T1DM + Alda1 groups were 0.20 ± 0.02 ng/mL and 0.22 ± 0.05 ng/mL, respectively, which were notably lower than those in the CON group (0.33 ± 0.08 ng/mL).
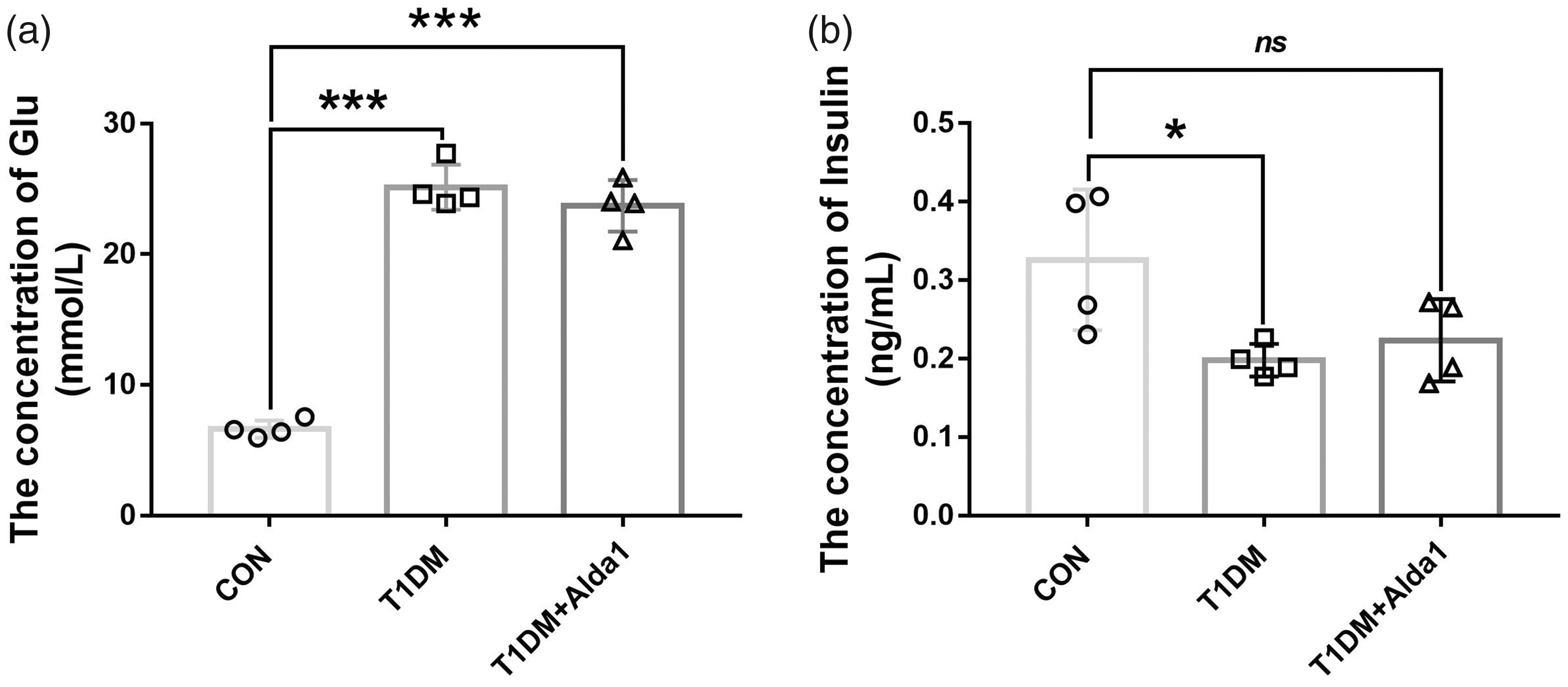
The levels of rats’ blood glucose and insulin at three months after T1DM induction. (a) The level of rats’ blood glucose. (b) The level of rats’ blood insulin. All analysis was performed in duplicates. The data were expressed as mean ± standard deviation (SD), n = 4 rats per group. *P < 0.05, T1DM group and T1DM+Alda1 group vs. CON group; ***P < 0.001, T1DM group and T1DM+Alda1 group vs. CON group; nsP > 0.05, T1DM group and T1DM+Alda1 group vs. CON group.
Effects of ALDH2 on corneal structure
To assess the severity of damage to diabetic corneas, slit lamp examination was performed at one, two, and three months after T1DM induction. Then, 1% fluorescein sodium (1 μL) was added to the conjunctival sac, after which the cornea was observed with a slit lamp under cobalt blue light conditions. As shown in Figure 2, the area of corneal fluorescein sodium staining in the T1DM + Alda1 group was less than that in the DM group at one and two months after T1DM induction, and there was no corneal fluorescein sodium staining in the CON group. At three months after T1DM induction, the T1DM group developed cataracts, and the crystalline lens in the T1DM + Alda1 group were relatively normal.
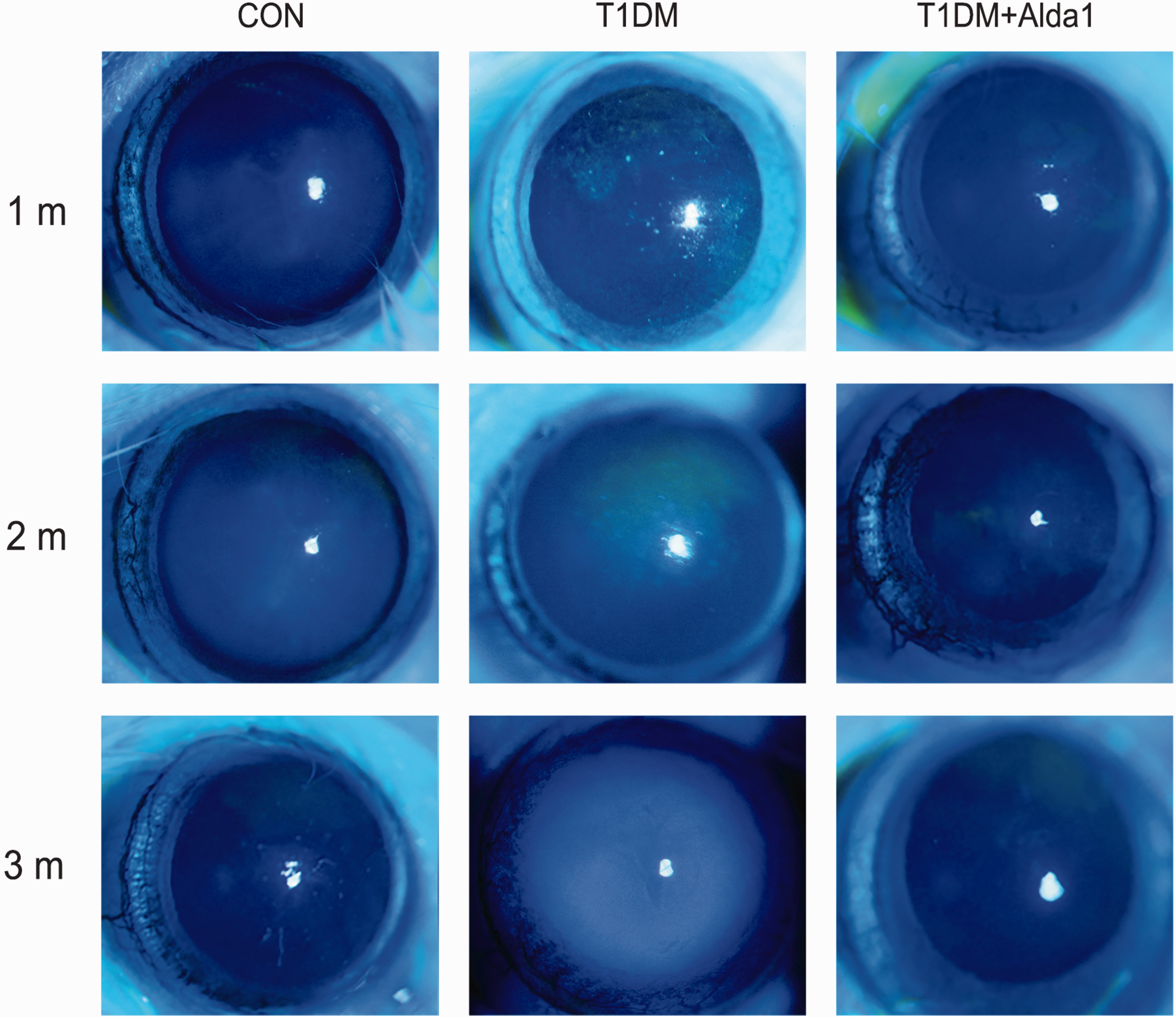
The typical rats’ corneal pictures with slit lamp at one, two, and three months after T1DM induction. (A color version of this figure is available in the online journal.)
Additionally, corneal morphology in diabetic rats was evaluated by HE staining and corneal optical coherence tomography (OCT) analysis. The HE staining results showed that the epithelial tissue in the CON group was arranged neatly, and the collagen fibers in the stroma were arranged parallel and were regular and compact. However, epithelial edema and structural disorders were both observed in the T1DM group. Interestingly, as shown in Figure 3(a), the corneal structure in the T1DM + Alda1 group was basically intact, and epithelial edema was alleviated. Moreover, the findings suggested that the corneal epithelial cells in the T1DM group were in an active proliferation state at one month after T1DM induction. At two months after T1DM induction, the corneal epithelial cells in the T1DM group were in an inactive proliferation state, while those in the T1DM + Alda1 group were more active than those in the T1DM group. Furthermore, the corneal epithelial cell layer in both the T1DM group and T1DM + Alda1 group was larger, covered with fewer corneal epithelial cells, and exhibited more epithelial keratinization than that in the CON group. As shown in Figure 3, the thickness of the central cornea in both the T1DM group (381.4 ± 70.8 μm) and T1DM + Alda1 group (260.2 ± 30.7 μm) was increased compared with that in the CON group (198.0 ± 9.9 μm) at three months after T1DM induction (all P < 0.05). Interestingly, the cornea in the T1DM + Alda1 group was less edematous than that in the T1DM group, but there was no significant difference (P < 0.05).
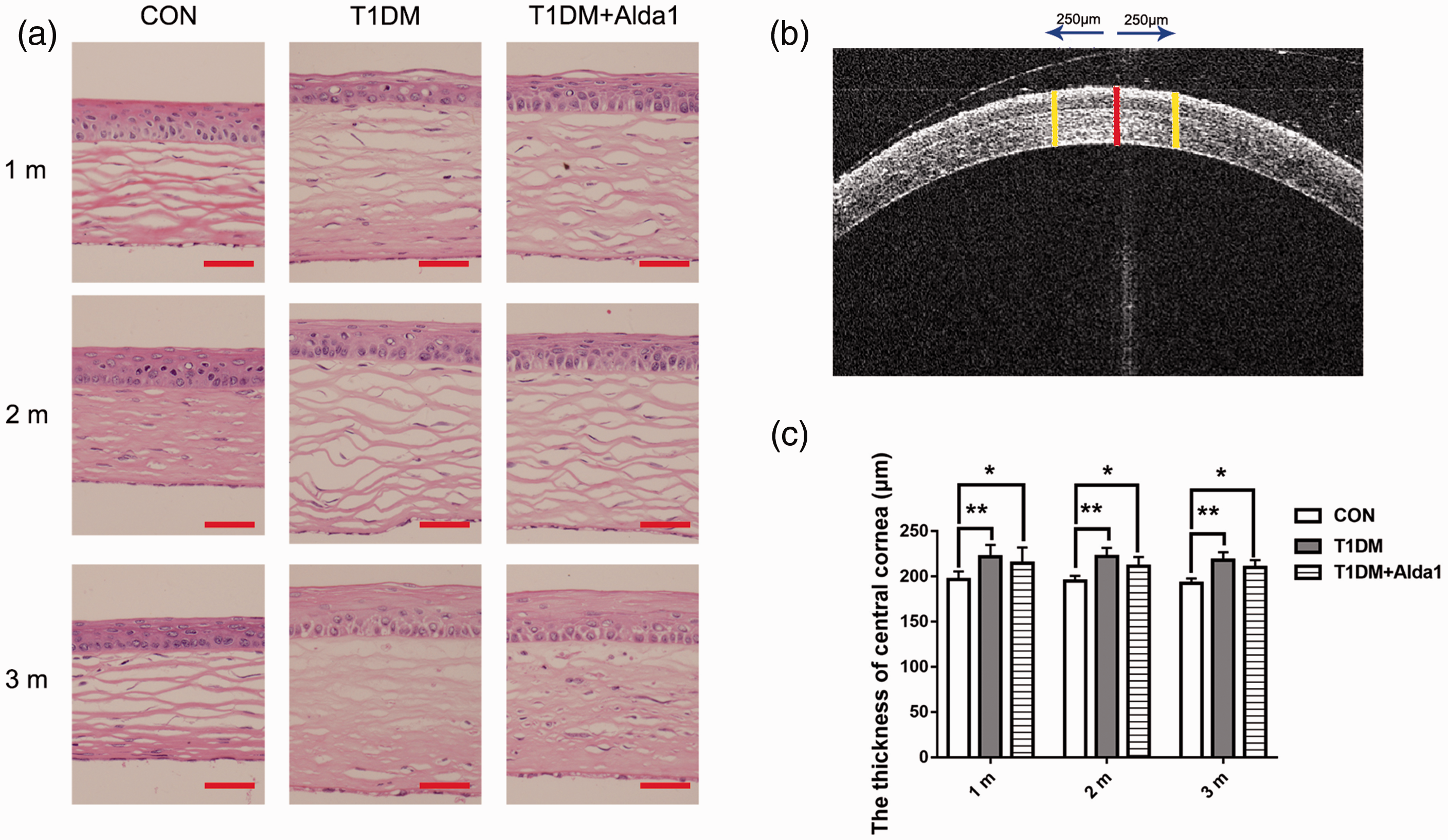
ALDH2 alleviated the diabetic rats’ corneal structure injury at one, two, and three months after T1DM induction. (a) The typical corneal HE staining picture at one, two, and three months after T1DM induction; (b) The typical corneal OCT picture at one month after T1DM induction; (c) The thickness of rats’ cornea at one, two, and three months after T1DM induction. All analysis was performed in duplicates. The data were expressed as mean ± standard deviation (SD), n = 4–12 rats per group. Scale bar: 50 μm. *P < 0.05, T1DM group and T1DM +Alda1 group vs. CON group; **P < 0.01, T1DM group and T1DM +Alda1 group vs. CON group. (A color version of this figure is available in the online journal.)
Effects of ALDH2 on systemic inflammation
To estimate corneal inflammation in diabetic rats, serum inflammatory factors and antioxidative factors were measured. As shown in Figure 4, the ELISA results revealed that the serum IL-1 and IL-6 levels were increased in diabetic rats to some extent compared to CON rats at three months after T1DM induction. Interestingly, those factors were significantly reduced in the DM + Alda1 group compared with the T1DM group (all P < 0.05), and there were no differences between the CON group and the T1DM + Alda1 group. However, the levels of SOD and TNF-α were not significantly different among the CON group, T1DM group, and T1DM + Alda1 group.
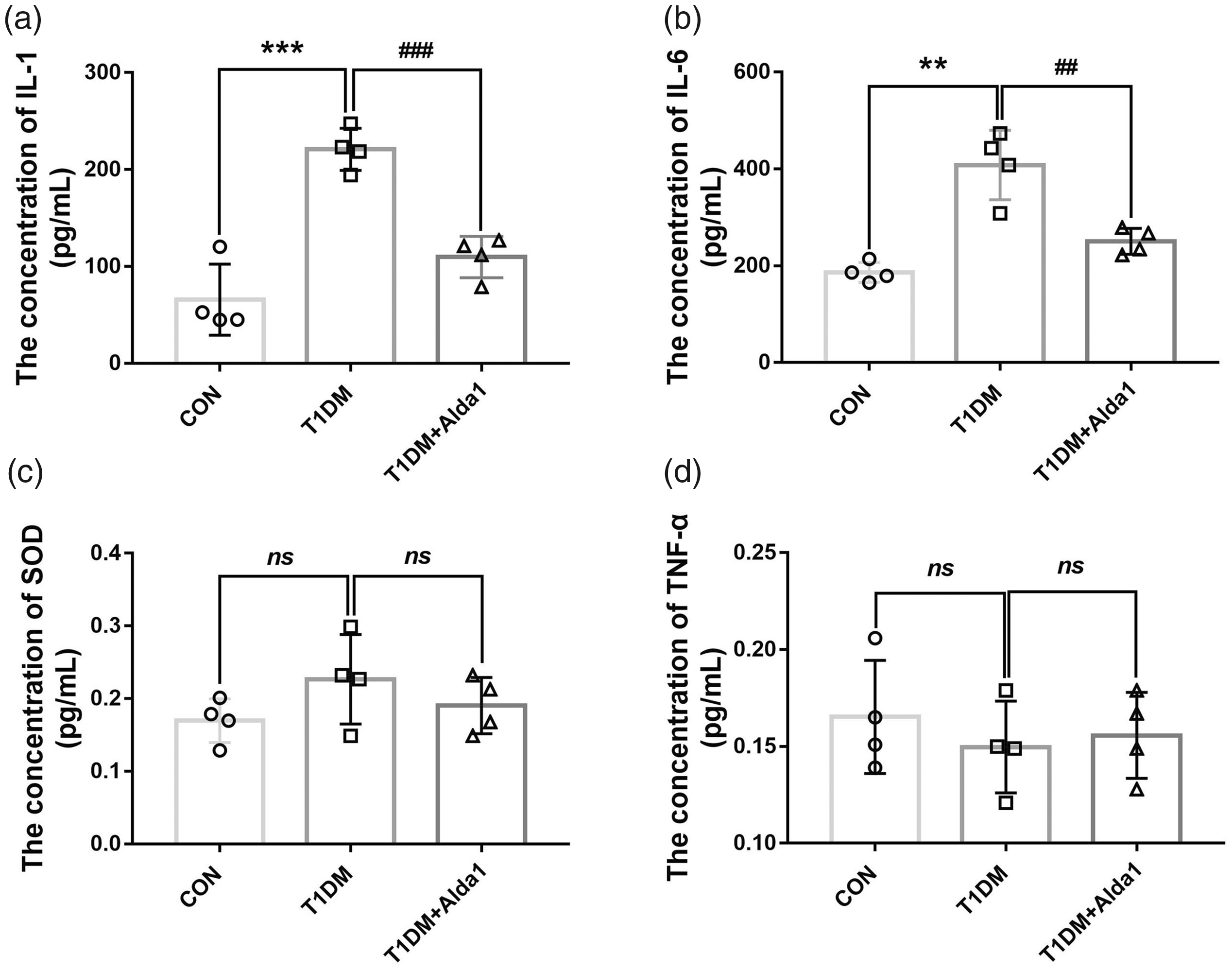
ALDH2 decreased serum levels of IL-1 and IL-6 at three months after T1DM induction. (a) The serum level of IL-1 at three months after T1DM induction; (b) The serum level of IL-6 at three months after T1DM induction; (c) The serum level of SOD at three months after T1DM induction; (d) The serum level of TNF-α at three months after T1DM induction. All analysis was performed in duplicates. The data were expressed as mean ± standard deviation (SD), n = 4 rats per group. **P < 0.01, T1DM group and T1DM +Alda1 group vs. CON group; ***P < 0.001, T1DM group and T1DM +Alda1 group vs. CON group; nsP > 0.05, T1DM group and T1DM +Alda1 group vs. CON group or T1DM +Alda1 group vs. T1DM group; ##P < 0.01, T1DM +Alda1 group vs. T1DM group; ###P < 0.001, T1DM +Alda1 group vs. T1DM group.
Effects of ALDH2 on the expression of corneal VEGF-A
To further measure the expression of VEGF-A, a crucial factor in angiogenesis and inflammation, an immunofluorescence assay was used. Figure 5 clearly shows that ALDH2 levels in diabetic rat corneas were lower than those in the CON group at one, two, and three months after T1DM induction (all P < 0.05). However, the expression of ALDH2 was obviously upregulated in Alda1-treated diabetic corneas compared to those in the T1DM group at two and three months after T1DM induction (all P < 0.05). As shown in Figure 6, VEGF-A was robustly expressed in the corneal epithelial layer. The expression of VEGF-A in the T1DM group at one, two, and three months after T1DM induction was dramatically increased compared with that in the CON group (all P < 0.05), while the expression of VEGF-A in the T1DM + Alda1 group, which had activated corneal ALDH2, was lower than that in the T1DM group at two and three months after T1DM induction (all P < 0.05).
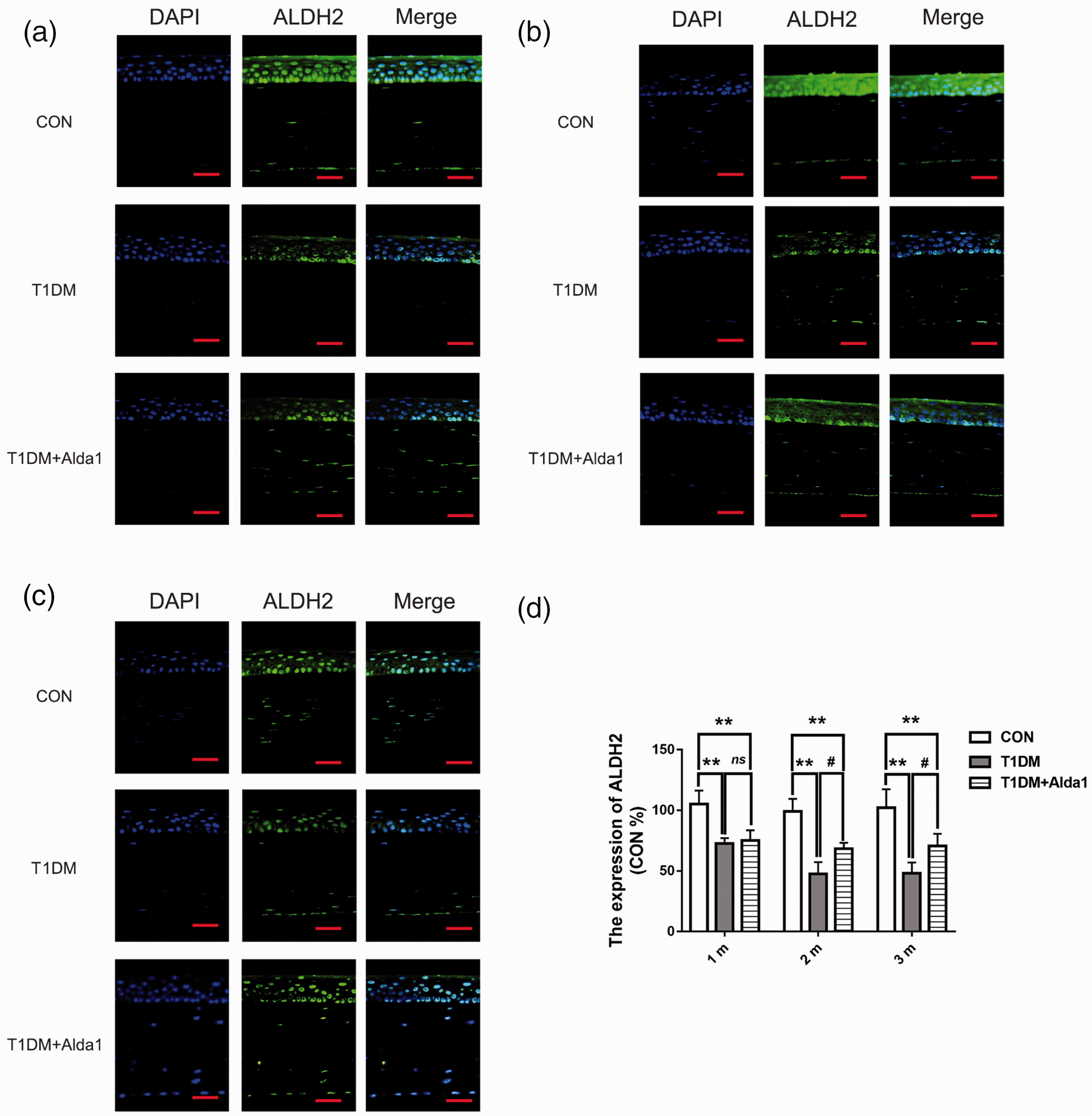
Alda1 upregulated the expression of ALDH2 at two and three months after T1DM induction. (a–c) The typical ALDH2 immunofluorescence staining pictures at one, two, and three months after T1DM induction. (d) The expression of ALDH2 at one, two, and three months after T1DM induction. All analysis was performed in duplicates. The data were expressed as mean ± standard deviation (SD), n = 4 rats per group. Scale bar: 50 μm. **P < 0.01, T1DM group and T1DM +Alda1 group vs. CON group; #P < 0.05, T1DM +Alda1 group vs. T1DM group; nsP > 0.05, T1DM +Alda1 group vs. T1DM group. (A color version of this figure is available in the online journal.)
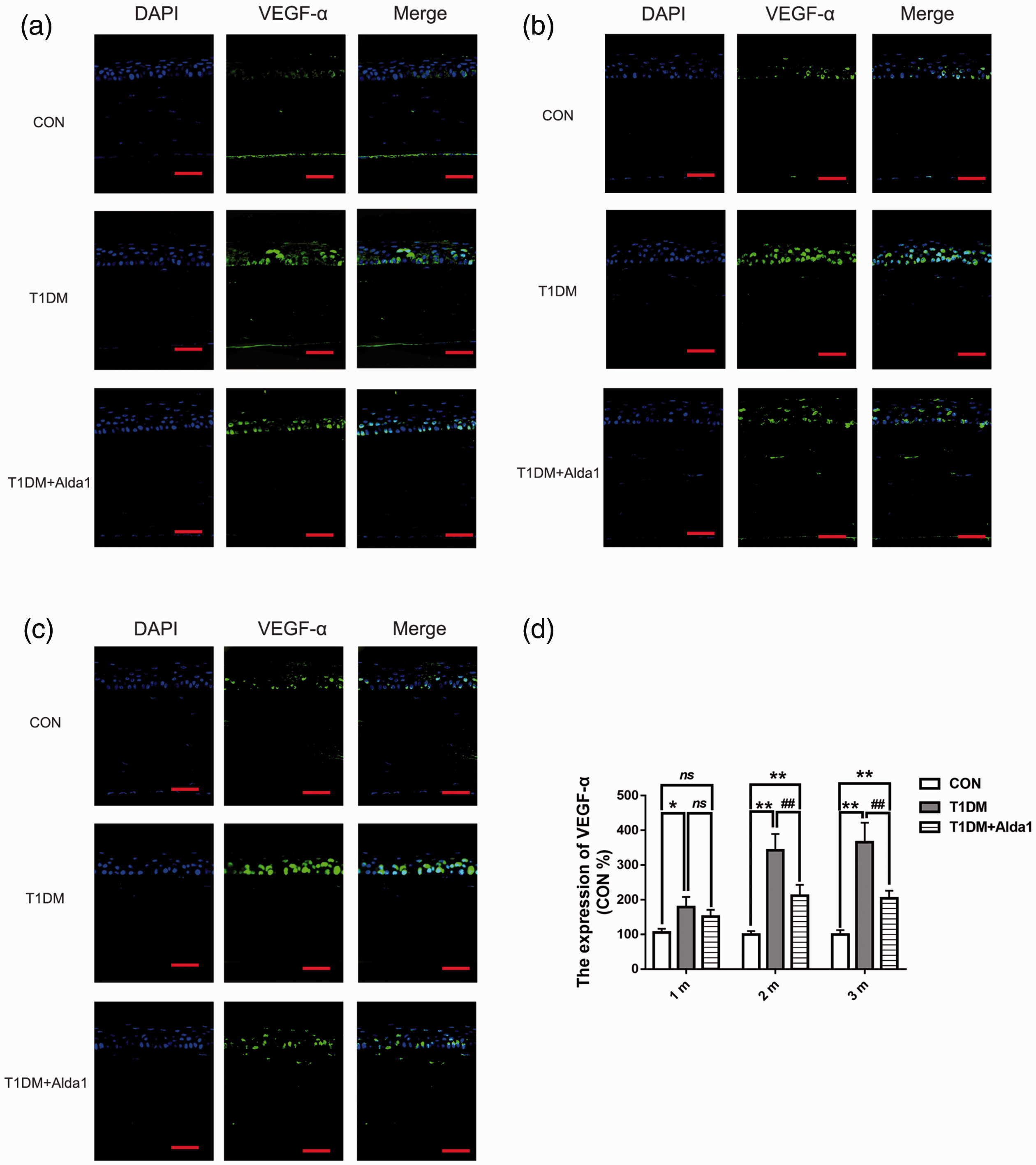
ALDH2 decreased the expression of VEGF-A at two and three months after T1DM induction. (a–c) The typical VEGF-A immunofluorescence staining pictures at one, two, and three months after T1DM induction. (d) The expression of VEGF-A at one, two, and three months after T1DM induction. All analysis was performed in duplicates. The data were expressed as mean ± standard deviation (SD), n = 4 rats per group. Scale bar: 50 μm. *P < 0.05, T1DM group and T1DM +Alda1 group vs. CON group; **P < 0.01, T1DM group and T1DM +Alda1 group vs. CON group; nsP > 0.05, T1DM group and T1DM +Alda1 group vs. CON group or T1DM +Alda1 group vs. T1DM group; ##P < 0.01, T1DM +Alda1 group vs. T1DM group. (A color version of this figure is available in the online journal.)
Effects of ALDH2 on the level of corneal cell death
To estimate corneal apoptosis (or cell death) in cells treated with or without Alda1, a TUNEL staining assay was performed. As shown in Figure 7, corneal apoptosis mainly occurred in the corneal epithelial layer. Furthermore, the level of TUNEL staining in the T1DM group and T1DM + Alda1 group at one, two, and three months after T1DM induction was dramatically increased compared with that in the CON group (all P < 0.05), while the level of TUNEL staining in the T1DM + Alda1 group was less than that in the T1DM group at one and two months after DM induction (all P < 0.05).
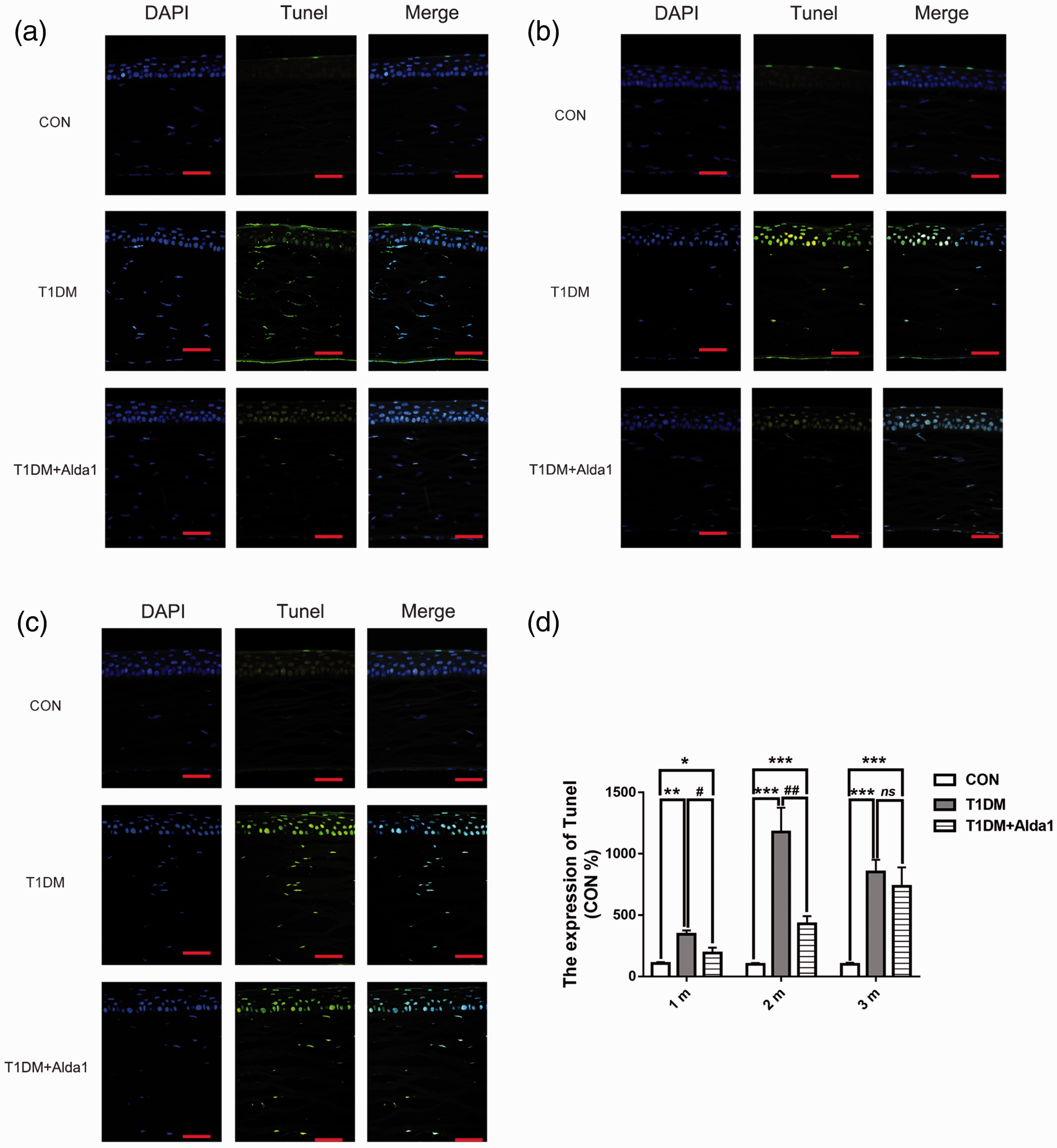
ALDH2 decreased the corneal cell death at one, two, and three months after T1DM induction. (a–c) The typical corneal TUNEL staining pictures at one, two, and three months after T1DM induction. (d) The expression of corneal TUNEL at one, two, and three months after T1DM induction. All analysis was performed in duplicates. The data were expressed as mean ± standard deviation (SD), n = 4 rats per group. Scale bar: 50 μm. *P < 0.05, T1DM group and T1DM +Alda1 group vs. CON group; **P < 0.05, T1DM group and T1DM +Alda1 group vs. CON group; ***P < 0.001, T1DM group and T1DM +Alda1 group vs. CON group; nsP > 0.05, T1DM +Alda1 group vs. T1DM group. #P < 0.05, T1DM + Alda1 group vs. T1DM group. (A color version of this figure is available in the online journal.)
Discussion
Currently, with the increasing prevalence of diabetes mellitus (DM), ophthalmological complications have emerged as key causes of vision loss worldwide. 32 In addition to retinal disorders (diabetic retinopathy) and lens lesions (diabetes-related cataracts), varying types of corneal injuries are also reasonably common in DM patients.33,34 In the present study, we found that STZ-induced T1DM could result in corneal epithelial keratinization, stromal structure disorders, and systemic inflammation. Remarkably, the activation of ALDH2 alleviated corneal stromal cell edema, maintained corneal epithelial cell integrity, improved corneal oxygen deficits, and reduced systemic inflammation.
In this study, corneal structure disorders were observed in T1DM rats. According to the corneal fluorescein staining results, T1DM rats exhibited enhanced corneal fluorescence, showing prominent corneal epithelial injury and indicating progressive impairment of epithelial barrier function. Additionally, the dry eye-like symptom of corneal keratinization was observed in T1DM rats. Epithelial proliferation may participate in corneal structural repair and functional reconstruction as a stress reaction to DM. 35 We considered corneal keratinization to be associated with epithelial cell hyperproliferation and hypertrophy. Moreover, this pathological keratinization is an important reason for severe visual loss. 36 Interestingly, ALDH2 alleviated pathological corneal epithelial cell keratinization, which maintained a relatively intact corneal structure.
While the pathogenesis of diabetic corneal abnormalities is incompletely understood, persistent hyperglycemia remains the principal instigator. Although DK partially results from epithelium-basement membrane adhesion disorders and corneal neuropathy, complicated epithelial abnormalities may also be associated with specific variations in cell signaling.37,38 The underlying molecular pathways sustaining cellular homeostatic processes are diminished in DM patients. Disadvantages, such as hypoxia, inflammation, and oxidative stress, affect corneal structure and function, which leads to deferred epithelial wound healing in diabetic corneas.39–41 Consistent with these findings, our results demonstrated significantly upregulated levels of inflammatory factors, such as IL-1 and IL-6, in T1DM rat serum, indicating a prominent systemic inflammatory response. Other researchers have reported that hyperglycemia-induced dysfunction in intracellular antioxidative abilities may cause severe corneal epithelial cell damage in diabetic patients.39,42 However, in our study, serum SOD and TNF-α levels showed no statistically significant difference between diabetic and nondiabetic rats. It is hypothesized that the regulation of systemic oxidative and inflammatory homeostasis was not compromised entirely during the three-month duration of diabetes in this study. This finding is consistent with the evidence that limited chronic low-grade inflammation exists during the development of diabetic complications. 33
It was reported that epithelial disorder in diabetic corneas was related to epithelial cell loss. 43 The occurrence of corneal epithelial apoptosis (or cell death) was convincingly demonstrated by the TUNEL assay in the present study. This result further showed that apoptosis (or cell death) occurred at the corneal epithelial layer in T1DM rats. Therefore, we supposed that a vicious cycle of apoptosis (or cell death) endangered epithelial cell function, leading to the pathologic progression of DK. Our findings were consistent with those of other studies that identified apoptosis as one of the most pivotal pathological reactions in DK.40,44 Notably, ALDH2 inhibited corneal epithelium apoptosis (or cell death) to some extent. This effect may be associated with the ability of ALDH2 to improve the systemic inflammatory environment.
In many pathological conditions, inflammation is the key driver of corneal angiogenesis.45,46 Proinflammatory cytokines (IL-1 and IL-6) can promote corneal neovascularization by enhancing the production of the proangiogenic molecules basic fibroblast growth factor (bFGF) and VEGF by corneal epithelial cells.47,48 This study showed that VEGF-A was overexpressed in the cornea in T1DM and was strongly expressed in the epithelium and in some keratocytes. This finding conforms to the fact that ocular tissues in DM, including retinas and vitreous and aqueous humor, often have elevated VEGF levels. 49 Indeed, the increased protein level of VEGF-A in the diabetic cornea was similar to that in the diabetic retina in previous studies by our group and others. 47 Furthermore, corneal VEGF-A binds to soluble VEGF receptor 1 and is sequestered by the basement membrane to maintain corneal physiological avascularity under normal conditions. 50 We hypothesize that corneal VEGF-A plays a role in inducing an angiogenic response during injury and pathological conditions such as DK. It was also reported that in the hypoxic cornea, enhanced VEGF expression was a mechanism to reinforce the supply of oxygen to the cornea. 51 Significantly, our results indicated that ALDH2 activation could suppress the expression of corneal VEGF-A in T1DM rats, which prevented pathological neovascularization and could protect the cornea from inflammation and hypoxia.
One important point to note is that the molecular mechanism of ALDH2 investigated in the present study focused on DK, which, to our knowledge, has not been explored before. However, because this was an initial attempt to investigate the potential role of ALDH2 in diabetic corneas, the study period was limited, and the specific mechanisms, especially downstream signal transduction, of ALDH2 remain to be further elucidated in the future. Ultimately, it is certain that the activation of ALDH2, which is known to detoxify noxious aldehydes, can protect against the development of DK. ALDH2 can protect corneal structure, reduce corneal apoptosis, and attenuate systemic inflammation, and these effects are associated with the VEGF pathway. Therefore, the present study sheds light on a novel mechanism of ALDH2 in corneal abnormalities in T1DM rats.
Conclusions
In summary, DK can cause a variety of pathological corneal alterations. Moreover, ALDH2 protected corneal structural integrity by alleviating corneal edema, improving inflammatory reactions, and attenuating VEGF-A expression and apoptosis. Overall, this study provided effective evidence that ALDH2 may be a good therapeutic candidate/target for DK treatment.
Footnotes
AUTHORS' CONTRIBUTIONS
PL and MSH contributed equally to the study. YHH and FH generated the conception and experimental design; PL and MSH conducted the experiments, made the data analysis, and wrote and revised the manuscript. XZ, TL, and YS collected data, analyzed data, and prepared results. HL and WJ provided intellectual support and edited the manuscript.
ACKNOWLEDGEMENTS
The authors thank Master Dongyu Wei for helping establish rat model, who are from Center of Clinical Aerospace Medicine, Fourth Military Medical University.
Declaration OF CONFLICTING INTRESTS
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
FUNDING
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Nature Science Foundation of China (82001484). The funding body had no influence on the design, collection, analysis or interpretation of the data, or in writing the manuscript.