
Abstract
Sepsis is characterized by persistent systemic inflammation, which can cause multi-organ dysfunction. The poly polymerase-1 inhibitor olaparib possesses anti-inflammatory properties. This study aimed to assess the effects of olaparib (pre- and post-treatments) on sepsis, and to investigate whether it could suppress CD14 expression via the ERK pathway in polymicrobial sepsis and peritoneal macrophages models. Sepsis was induced by cecal ligation and puncture in C57BL/6 male mice. Fifty mice were randomly divided into five groups: The sham group was treated with vehicle or olaparib, the cecal ligation and puncture group with vehicle or with olaparib (5 mg/kg i.p.) 1 h before or 2 h after surgery. Olaparib pretreatment significantly improved the survival of septic mice (P < 0.001). Pre- and post-treatment of mice with olaparib partly alleviated cecal ligation and puncture-induced organ injury by decreasing the amounts of the pro-inflammatory mediators TNF-α and IL-6 as well as bacterial burden in the serum, peritoneal lavage fluid, and organs (P < 0.05). The protective effect of olaparib was associated with CD14 suppression via inhibition of ERK activation. Olaparib facilitated negative regulation of ERK-mediated CD14 expression, which may contribute to multi-organ injury in sepsis.
Keywords
Impact statement
Sepsis is accompanied by numerous counter-regulatory mechanisms, resulting in widespread organ injury and even death. So far, no specific pharmacological compound treats this condition, despite intense research and clinical trials targeting immune inflammatory and coagulation abnormalities. The PARP1 inhibitor, olaparib possesses anti-inflammatory properties. Our present study found that olaparib pretreatment increases survival rates and attenuates CLP-induced organ injury by decreasing the levels of TNF-α and IL-6 and of the bacterial burden in the serum, peritoneal lavage fluid, and organs. The protective effect of olaparib treatment was associated with suppression of the expression of CD14 via the inhibition of ERK activation, which may contribute to multi-organ injury in sepsis.
Introduction
Sepsis is considered as a life-threatening organ dysfunction resulting from dysregulated host immune response to infection, 1 and still has an unacceptably high fatality rate in intensive unit (ICU). 2 This dysfunction results in elevated systemic release of pro- and anti-inflammatory molecules. 3 Dysregulated host immune response is characterized as overwhelming cytokine production. Despite numerous researches and clinical trials had been conducted on immune inflammatory responses, seldom breakthroughs had been accepted in clinical practice. Therefore, the treatment of sepsis still requires continuous and in-depth research.
Recent works have demonstrated that Poly (ADP-ribose) polymerase-1 (PARP-1) plays a key role in cell necrosis and organ failure in a variety of inflammation-related diseases.4,5 PARP-1 activation is a central mechanism of LPS-induced acute lung injury (ALI). 6 PARP inhibitors, INO-1001 treatment significantly attenuated smoke inhalation-induced ALI. 7 Olaparib is a clinically approved PARP-1 inhibitor that possesses anti-inflammatory properties. Kapoor K et al. 8 demonstrated that olaparib exerts therapeutic effects in mice exposure to acute lung injury and acute kidney injury. Recent work of Choudhuri et al. 9 indicated that PARP1 deletion significantly induced pro-inflammatory macrophage activation in Chagas mice.
Although olaparib has the potential benefit effect for the treatment of tissue injury following inflammatory condition, the mechanism of olaparib in sepsis remains unclear. Mitogen-activated protein kinases (MAPKs) involved in several signal transduction, transmitting signals from the cell surface to the nuclear compartment. 10 Early studies indicated that MAPK activation triggered the production of pro-inflammatory cytokines and other mediator releases under LPS stimuli.11,12 Recent study found that inhibition of ERK1/2 prevents PARP-1 activation in neuronal cell death and inflammation. 13 Some studies also reported that ERK signaling is an alternative pathway responsible for PARP-1 activation. 14 Works of Chowdhury et al. 15 and Hocsak et al. 16 show that PARP-1 inhibition reduced the phosphorylation of ERK, resulting reduction of cell proliferation and ROS production separately. Inhibited the activation of ERK1/2 can significantly decreased inflammatory response, 17 but the relevant downstream targets remain poorly defined.
CD14 represents an important pattern recognition receptor in innate immunity, mediating local and systemic host responses to components released by Gram-negative bacteria, e.g. LPS.18–23 As an endotoxin receptor, CD14 is found as the soluble type in the serum or on the surface of macrophages. 23 Upregulation of CD14 increases LPS-induced inflammatory reactions in monocytes/macrophages. 12 CD14 blockade provides therapeutic potential in endotoxin-challenged monkeys 20 and rabbits. 22 Previously, our team revealed that decreased expression of CD14 is associated with markedly reduced inflammation and improved survival in mice with sepsis. 24 Furthermore, ERK pathway suppression results in a significant decrease of CD14 expression in monocytes treated with 27-hydroxychalesteral. 25
In this study, we have investigated whether olaparib protects from organ inflammation and damage related to sepsis via ERK-mediated CD14 expression.
Materials and methods
Animals
Male wildtype mice (C57BL/6; 8–12 weeks) were purchased from Shanghai Laboratory Animal Co. Ltd. (SLAC, Shanghai, China), and underwent housing in specific pathogen-free (SPF) conditions in the vivarium of Shanghai Tongji University under a 12-h–-12-h light/dark cycle. Assays involving animals had approval from the Ethics Committee of Tongji University, and strictly followed the National Institutes of Health Guidelines for the Use of Laboratory Animals (Ethic Number: TJ00121101).
CLP-induced sepsis
After anesthesia with intraperitoneal (i.p.) xylazine (10 mg/kg) and ketamine (100 mg/kg), the abdomen of mice was shaved, and followed by a 2 cm midline incision. The cecum isolation and ligation with a 4–0 silk ligature at 75% of the distance between cecal distal pole and base were performed. 26 The cecum was punctured with a 21-G needle, from the mesentery to the antimesenteric side, and followed by ligation. After returning the cecum to the peritoneum, incision closure was performed immediately by using 4–0 sterile synthetic absorbable suture. Fifty animals were randomly divided into five groups: (1) Control group: sham mice treated with vehicle, samples were taken after surviving 24 h; (2) Ola group: sham animals treated with olaparib (AZD2281, Ku-0059436) (5 mg/kg i.p.), samples were taken after surviving 24 h; (3) CLP group: CLP animals administered vehicle, samples were taken after CLP 24 h; (4) CLP+ola-pre group: CLP animals administered olaparib 1 h prior to surgery, samples were taken after CLP 24 h; (5) CLP+ola-post group: CLP animals administered olaparib 2 h following surgery, samples were taken after CLP 24 h. Groups were cervical dislocation for euthanized after surviving or CLP 24 h and samples were collected immediately for subsequent analysis.
Mortality assessment
Based on previous research, 26 buprenorphine (0.1 mg/kg) is injected subcutaneously every 12 h for postoperative analgesia. The survival study was performed by monitoring the mice every 12 h for a maximum period of seven days. The surviving mice underwent euthanasia by cervical dislocation.
Histological assays
Appropriate lung and liver lobe samples were used to perform histological staining by hematoxylin and eosin (H&E). 24 For assessing injury level, histological changes were quantitatively scored via neutrophil migration, edema and tissue infiltration, hyperemia and congestion, hemorrhage and debris, and cell hyperplasia. These parameters underwent semi-quantitative analysis as no, minimal, light, moderate, and severe, with scores 0, 1, 2, 3 and 4, respectively.
Total protein samples
The extracted alveolar lavage fluid of mice was centrifuged at 1500 r/min for 10 min at 4°C, and the supernatant was taken out. The BCA method was utilized to assess the supernatant, referring to the literature for specific steps. According to the obtained OD value and the standard curve, the sample’s protein concentration was calculated.
Bacterial load assessment
Following 24 h of incubation after CLP, serum, PLF, and tissue (spleen, lung, liver and kidney) specimens from mice were collected into sterile 1.5 mL EP tubes, respectively. The tissues were homogenized in 0.5 mL of sterile DPBS. The principle of aseptic operation was used, and serum samples, PLF specimens, and the homogenates of each tissue were diluted at a ratio of 1:20. Subsequently, 100 µL of the suspension were evenly spread on tryptic soy agars (BD, Franklin Lakes, NJ) for incubation at 37°C for 24 h. Bacterial amounts were expressed as colony-forming units (CFUs). Finally, log transformation was carried out for statistical analysis.
Cytokine level assessment
Circulating and PLF amounts of TNF-α and IL-6 were obtained with specific enzyme-linked immunosorbent assay (ELISA) kits (Raybiotech), as directed by the manufacturer.
Quantitative PCR
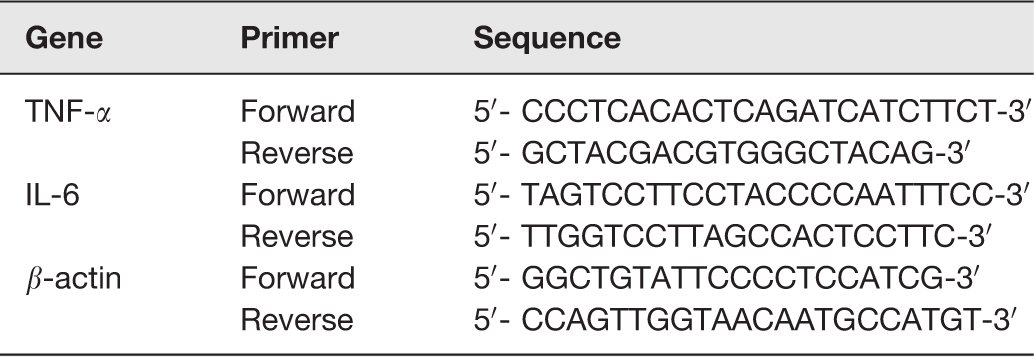
Total RNA extraction was carried out from tissues (lung, liver and kidney) with TRIzol reagent (Life Technologies, Grand Island, NY). First-strand cDNA was synthesized with the Reverse Transcription System (Takara) as directed by the manufacturer. Primers (Sangon Biotech) are displayed in Table 1.
Primers for quantitative polymerase chain reaction.
Immunoblot
Immunoblot for the detection of CD14 and ERK in tissues (lung, liver and kidney) and macrophages was performed according to standard protocols. 17 After blocking (5% skim milk), the membranes underwent overnight incubation with primary antibody against CD14 (BD Pharmingen, 557896, 1:1000) and phosphorylated ERK (p-ERK) (CST 4695 T, 1:1000), respectively. Next, the membranes were washed and incubated at ambient (2 h) with secondary antibodies (1:10,000). Subsequently, the Odyssey Infrared Imaging System (LI-COR) was utilized for image acquisition and analysis.
Cell cultures
Peritoneal macrophages (PMs) were obtained from wild-type mice (three days) following i.p. administration of 4% thioglycollate (2 mL) and placed in RPMI1640 with 10% FBS, 50 U/mL penicillin and streptomycin. 17 The cells were stored overnight. The floating containers were washed with PBS, and cells underwent incubation in freshly prepared medium overnight. Then, adherent macrophages underwent further incubation in serum-free RPMI1640 at 37°C for 2 h and subsequent treatment with LPS (List Biological Laboratories, Escherichia coli 055:B5), Olaparib, or their combination.
Flow cytometry
Cells from mouse PLF specimens underwent labeling with fluorochrome-linked primary antibodies targeting F4/80 (eBioscience, 299805) and Ly-6G (Gr-1) (eBioscience, E00737-1633) for 30 min. Cell sorting was carried out flow-cytometrically after washing. F4/80- and Gr-1-positive cells were considered macrophages and neutrophils, respectively. Those positive for both F4/80 and Gr-1 were considered macrophages that had phagocytozed neutrophils. An LSR II flow cytometer (BD Biosciences) was utilized for data acquisition; data analysis used the DIVA software (BD Biosciences).
Statistical analysis
Data are mean ± standard error of the mean (SEM) from assays repeated three times or more. The survival curve was analyzed by Kaplan–Meier curve. Nonparametric Mann–Whitney U statistics was used to analyze the difference in the values of bacterial colony-forming units between groups. One-way ANOVA was used for comparison between multiple groups and Student’s t-test was used for comparison between two groups. P < 0.05 indicated statistical significance. Graph Pad Prism 6.0 was employed for data analysis.
Results
Olaparib improves survival in septic mice
The present study performed an initial investigation to assess whether olaparib (5 mg/kg, i.p.) reduces mortality when administered 1 h prior to CLP in intact mice. As expected, a significant decrease in mortality was noted following CLP with 5 mg/kg olaparib pretreatment. As shown in Figure 1(a), the survival rates in the CLP + olaparib group were markedly elevated versus CLP group values (P < 0.001). Collectively, these results indicated that olaparib pretreatment effectively decreased CLP-induced mortality in mice and played a protective role in sepsis.
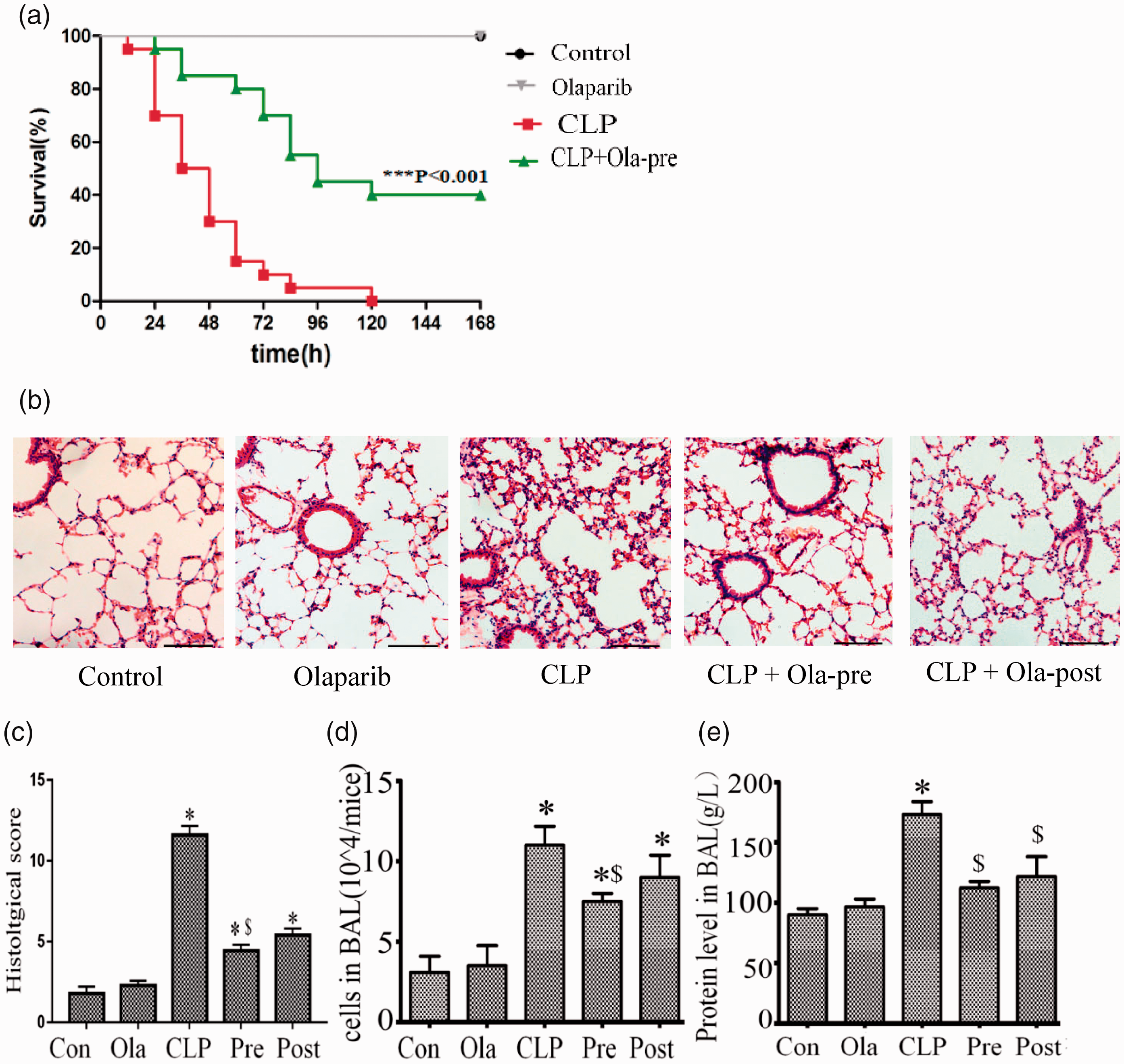
Olaparib improves survival and attenuates lung injury in septic mice. Mice were treated with i.p. olaparib (5 mg/kg) at 1 h prior to CLP induction or 2 h following CLP. (a) Survival rates (n = 12 per group). Kaplan–Meier curve was performed for survival analysis. *P < 0.001 versus CLP group. (b) Hematoxylin and eosin (H&E) stained lung samples from the five groups. Scale bar represents 100 µm. (c) Lung injury scores were measured. (d) Cellular counts. (e) Protein levels in BALF were detected from mice (24 h after Sham or CLP) administered olaparib (5 mg/kg) or not. Statistical difference was tested using Student’s t-test; *P < 0.05 versus control group; $P < 0.05 versus CLP group. (A color version of this figure is available in the online journal.)
Olaparib ameliorates multiple organ injury induced by CLP
Tissues (lung and liver) from mice that survived 24 h following CLP were examined by H&E staining. CLP resulted in ALI as determined by histopathological analysis (Figure 1(b)), and olaparib markedly alleviated ALI as reflected by decreased lung injury scores (Figure 1(c)), cellular counts (Figure 1(d)), and total protein amounts (Figure 1(e)) in the bronchoalveolar lavage fluid. Moreover, the improvement effect was more pronounced in the pre-treatment group in comparison with the post-treatment group.
Figure 2(a) depicts additional data demonstrating CLP-associated liver injury (and the alleviating effect of olaparib). CLP induced severe hepatocellular necrosis, which was partly suppressed by olaparib, as determined by liver injury scores (Figure 2(b)). The serum amounts of ALT and AST induced by CLP increased verifying the occurrence of live injury (Figure 2(c) and (d)), which were also partially suppressed by olaparib pre-treatment, but not by post-treatment. Therefore, olaparib pre-treatment exhibited a protective effect on the liver in mice with sepsis. Additionally, CLP-reduced increases in blood biomarkers of renal dysfunction (i.e. elevated serum BUN and creatinine amounts) were alleviated by both olaparib pre-treatment and post-treatment (Figure 2(e)).
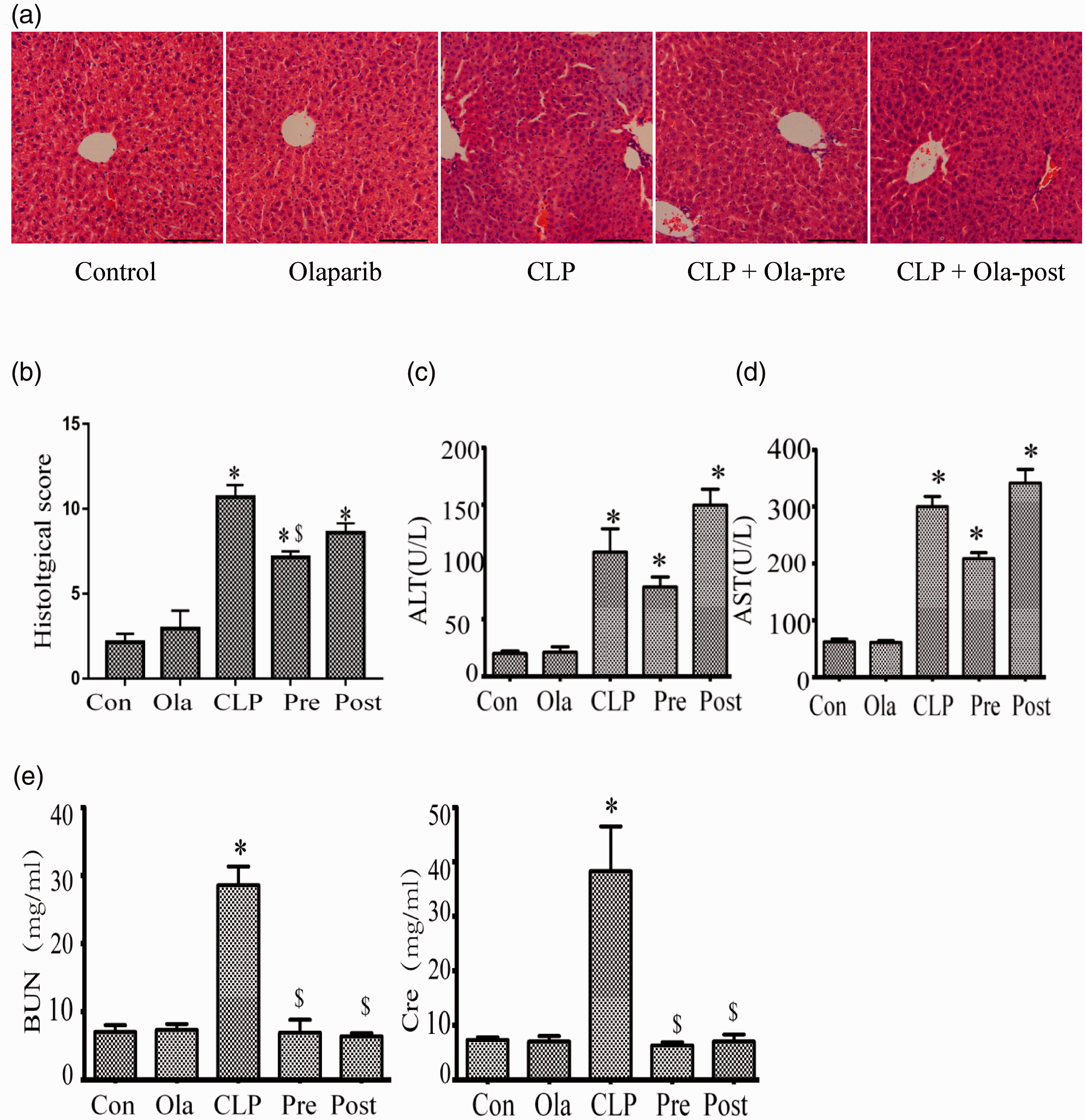
Olaparib ameliorates multiple organ injury induced by CLP. Mice were treated with i.p. olaparib (5 mg/kg) at 1 h prior to CLP induction or 2 h following CLP. (a) Hematoxylin and eosin (H&E) stained liver samples from four groups. Scale bar represents 100 µm. (b) Liver injury scores. (c, d) Serum ALT and AST levels determined for hepatocellular injury assessment. The serum levels of (e) BUN and creatinine were detected at 24 h post-CLP challenge. Statistical difference was tested using Student’s t-test; *P < 0.05 versus control group; $P < 0.05 versus CLP group. (A color version of this figure is available in the online journal.)
Olaparib suppresses pro-inflammatory cytokine expression in the tissue of intact mice with CLP
The expressions of TNF-α and IL-6 in serum (Figure 3(a) and (b)) were increased in the CLP group compared with those of the sham and olaparib groups. Although olaparib pre-treatment slightly inhibited TNF-α secretion compared with CLP group, the difference was not statistically significant. It is important to note that olaparib exhibited no significant effects on IL-6, either during the pre-treatment or the post-treatment periods. Compared with CLP group or CLP + post-treatment group, olaparib pre-treatment markedly decreased the secretions of TNF-α and IL-6 (Figure 3(c) and (d)) in PLF of septic mice, whereas the inhibition of TNF-α expression was more pronounced than that noted for IL-6.
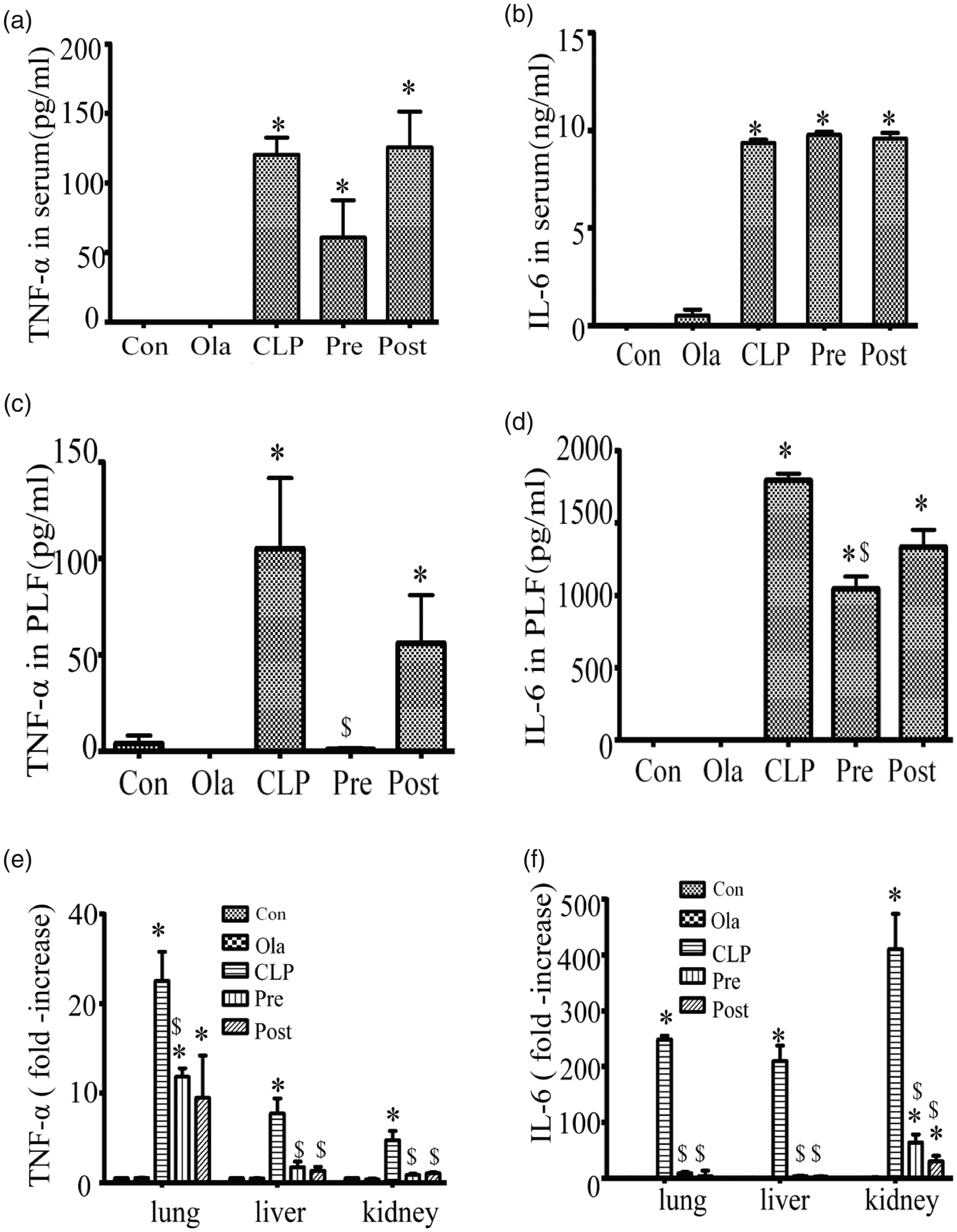
Olaparib suppresses pro-inflammatory cytokine secretion in intact mice with CLP. Mice were treated with i.p. olaparib (5 mg/kg) at 1 h prior to CLP induction or 2 h following CLP. TNF- and IL-6 amounts in (a, b) serum, (c, d) PLF, and (e, f) tissues (lung, liver, and kidney) in mice (24 h after Control or CLP operation) after treatment with olaparib (5 mg/kg) or not. Statistical difference was tested using Student’s t-test; * P < 0.05, **P < 0.01 versus control group; $P < 0.05 versus CLP group.
Moreover, both pre-treatment and post-treatment with olaparib significantly downregulated the expression of TNF-α and IL-6 induced by CLP in the tissues (lung, liver, and kidney) (Figure 3(e) and (f)), which was more notable compared to the changes observed for these inflammatory factors in serum and PLF. The above findings suggested olaparib reduced mainly the expression levels of TNF-α and IL-6 in local organs.
Olaparib regulates the functions of peritoneal macrophages and neutrophils in mice with sepsis
CLP was induced and following 24 h of cell incubation, the total number of inflammatory cells in PLF was clearly increased, whereas following olaparib treatment it was decreased (Figure 4(a)). It was shown that olaparib regulated the function of peritoneal macrophages/neutrophils in septic mice. Besides, the levels of F4/80 and Gr-1 were detected by flow cytometry (Figure 4(b)). CLP caused a decrease in the percentage of macrophage-phagocytic neutrophils, whereas olaparib treatment significantly increased the proportions of macrophages and phagocytic neutrophils, notably during the pre-treatment period (Figure 4(c)). Olaparib pre-treatment exhibited a more pronounced effect on immune cell function compared with the post-treatment group.
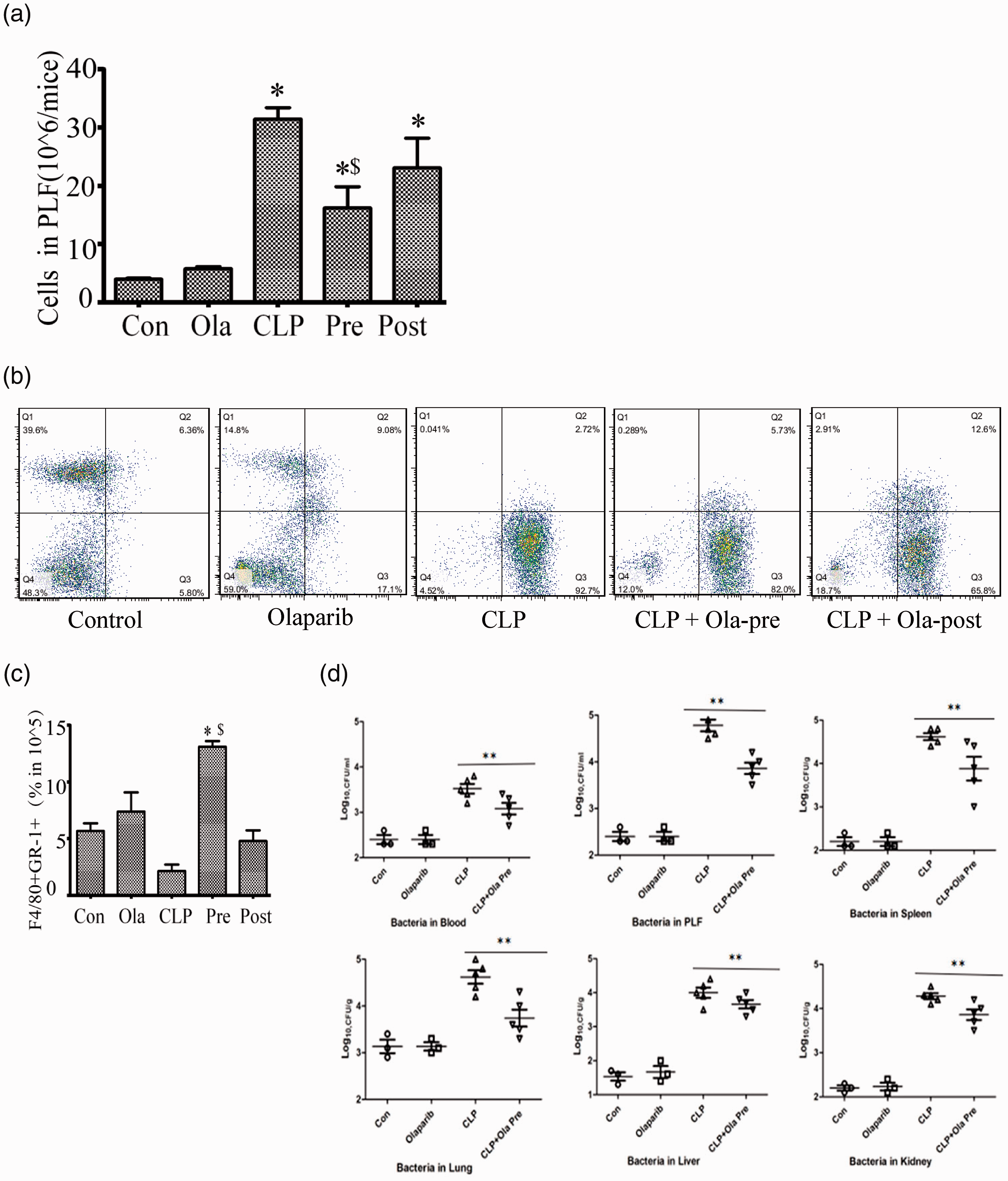
Olaparib regulates the function of peritoneal macrophages/neutrophils in septic mice. Mice were treated with i.p. olaparib (5 mg/kg) at 1 h prior to CLP induction or 2 h following CLP. (a) Cell counts in PLF of mice (24 h of Sham or CLP operation) after treatment with olaparib (5 mg/kg) or not. (b) F4/80 or Gr-1 positive cells detected flow-cytometrically. (c) Cells expressing both F4/80 and Gr-1. (d) Bacterial loads in serum, PLF, and tissues (spleen, lung, liver and kidney). Nonparametric Mann–Whitney U statistics was used to analyze the difference. Statistical difference was tested using Student’s t-test; *P < 0.05 versus control group; $P < 0.05 versus CLP group. (A color version of this figure is available in the online journal.)
Although CLP caused local infection in the abdominal cavity of experimental mice, the inflammatory response was systematic, which was also confirmed by bacterial load analysis. As shown in Figure 4(d), bacterial loads in the blood, PLF and tissues (lung, liver, kidney and spleen) were significantly increased in mice with sepsis, which could exacerbate the existing systemic infections. Olaparib pre-treatment inhibited CLP-induced bacterial loads in the serum, PLF, and tissues. This may be associated with the anti-inflammatory and enhanced bactericidal effects of the macrophages following phagocytosis of neutrophils.
Olaparib downregulates CD14 and inhibits the activation of ERK in tissues of intact mice with CLP
The expression characteristics of CD14 and p-ERK were detected using a relevant polymicrobial sepsis model. The tissues (lung, liver and kidney) were obtained at 24 h following the CLP surgery.
As shown in Figure 5, the synthesis of CD14 and p-ERK in tissues (lung, liver and kidney) was markedly elevated following CLP. Particularly, CD14 and p-ERK expression levels were greatly reduced regardless of olaparib pre- or post-treatment. Overall, these results implied that olaparib could suppress CD14 and p-ERK expression to alleviate multi-organ injury following CLP, whereas the association between CD14 and p-ERK requires further investigation.
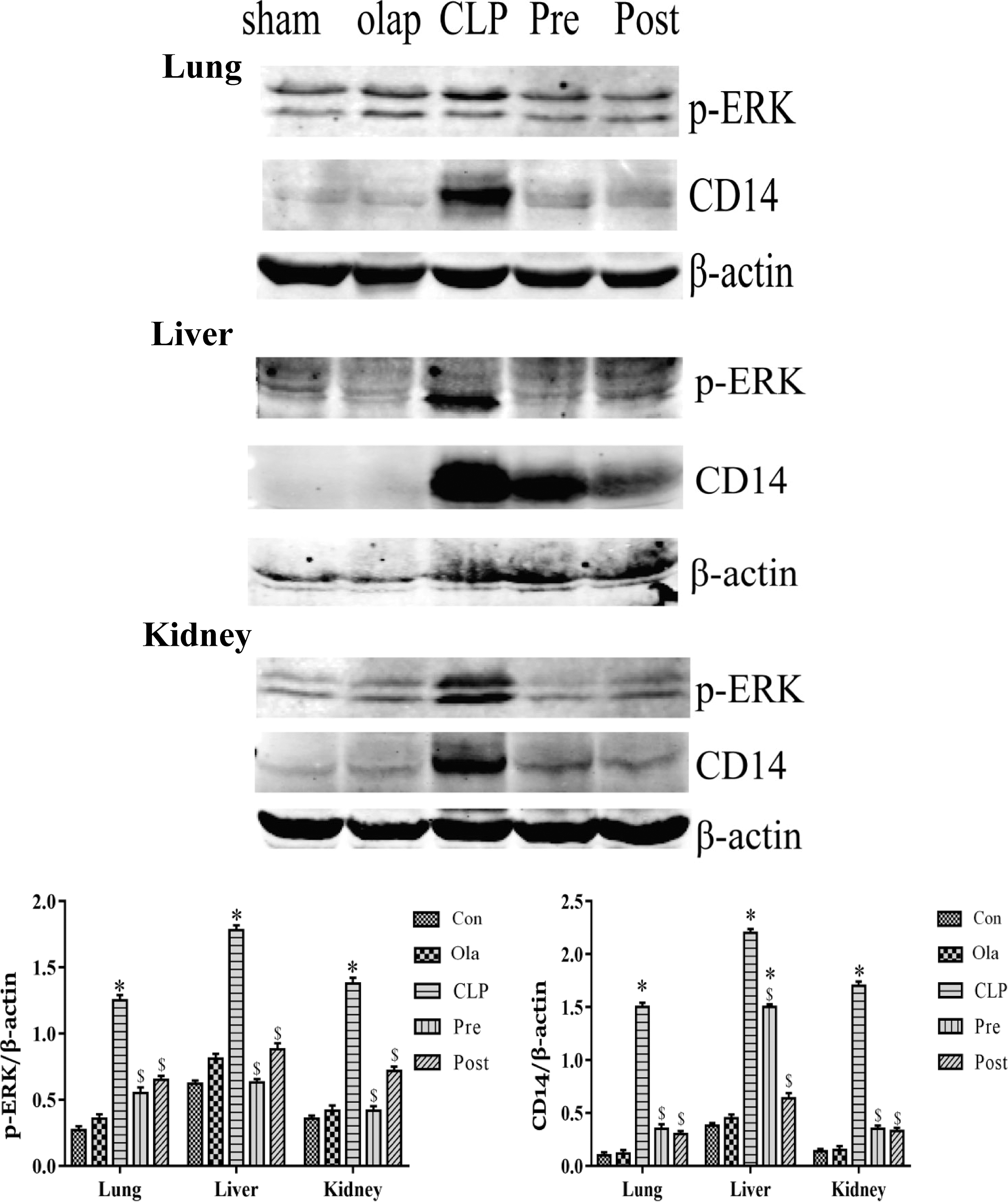
Olaparib downregulates the expression of CD14 and inhibits the activation of ERK in mice. Tissues (lung, liver, and kidney) were removed from the mice (24 h of Sham or CLP operation) after treatment with olaparib (5 mg/kg) or not. CD14 and p-ERK protein amounts assessed by immunoblot in intact mice treaded with CLP. Statistical difference was tested using Student’s t-test; *P < 0.05 versus control group; $P < 0.05 versus CLP group.
Olaparib attenuates CD14 expression by inhibition of the ERK activation in macrophages stimulated with LPS
To detect the function of olaparib on CD14 expression in PMs stimulated with LPS, this compound was administered at different dosages (0, 100, and 200 nM) and during different time periods of treatment (1 h prior to, and 0, 1 and 2 h following LPS). Olaparib treatment markedly inhibited LPS-induced CD14 expression at the indicated treatment periods (Figure 6(a)), and its effects were dose-dependent (Figure 6(b)). However, the mechanism by which olaparib inhibited CD14 expression remains unclear.
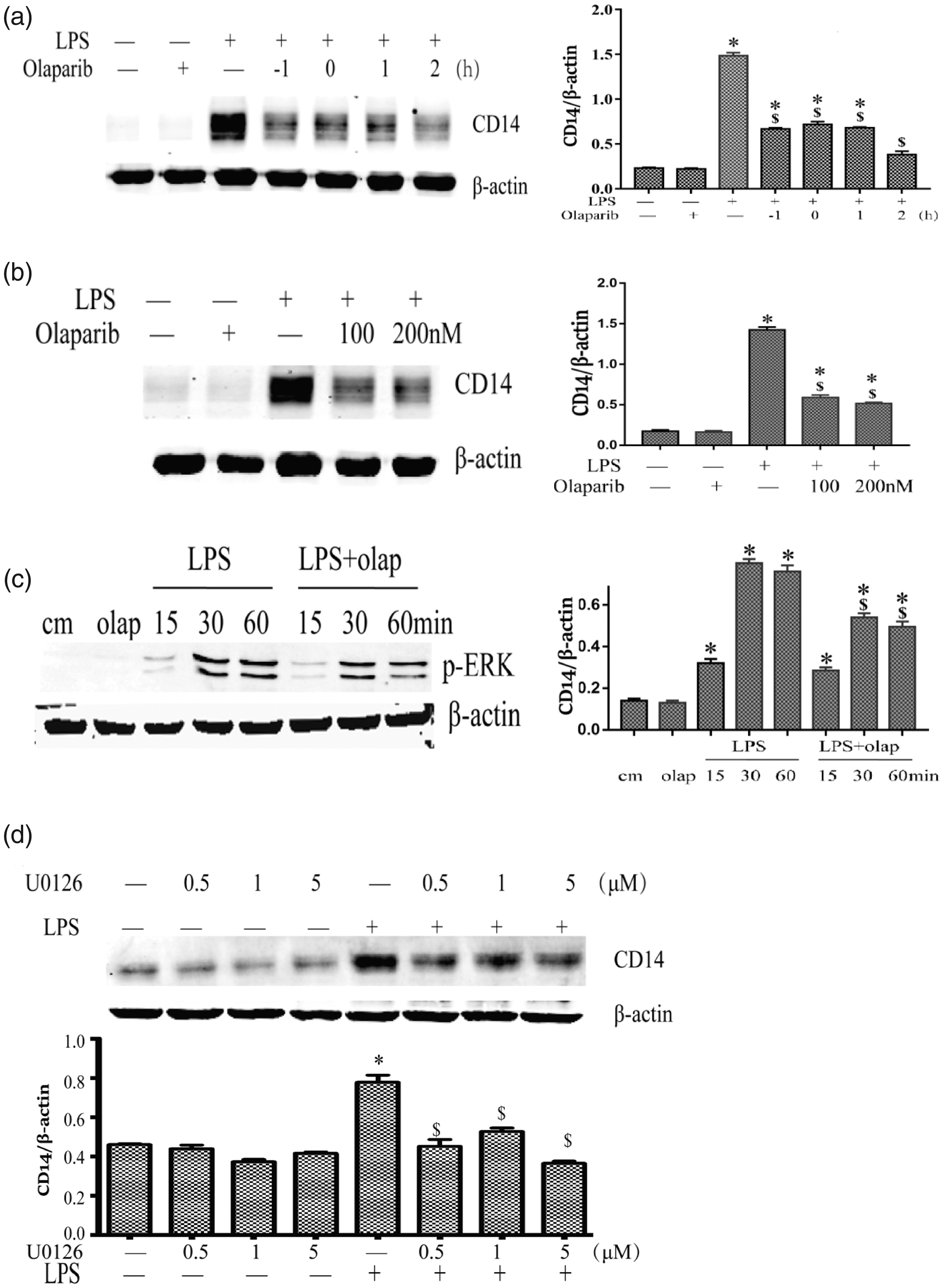
Olaparib attenuates CD14 expression by inhibition of the ERK pathway in macrophages. PMs were incubated with olaparib (0, 100, and 200 nM) 1 h prior to, and 0, 1, and 2 h following LPS stimulation for 12 h. (a–b) Immunoblot analysis of CD14 protein amounts. PMs were stimulated with LPS, with or without olaparib (200 nM) at different times (15, 30, and 60 min), and subsequently assessed for CD14 protein amounts by immunoblot (c). PMs were induced with LPS, with or without the ERK inhibitor U0126 (0.5, 1, and 5 µM), and subsequently examined for CD14 protein amounts by immunoblot (d). PMs were stimulated with LPS (100 ng/mL). Statistical difference was tested using Student’s t-test; *P < 0.05 versus control group; $P < 0.05 versus CLP group.
The results demonstrated that olaparib could effectively inhibit CLP-induced p-ERK expression in macrophages (Figure 6(c)). Moreover, the role of ERK in CD14 expression was assessed by pharmacological inhibition in present study. PMs were pretreated with an ERK inhibitor (U0126) for 1 h and subsequently administered LPS for 24 h. The results showed that U0126 inhibited LPS-induced CD14 protein expression (Figure 6(d)). In addition, the inhibitor itself did not affect CD14 expression in macrophages. Collectively, these findings indicated that olaparib could decrease CD14 expression by inhibiting the activation of ERK that partially contributed in the reduction of sepsis-associated inflammation.
Discussion
Sepsis represents a complicated disease that generally evolving into the host immune response to infectious agents. 2 The course of sepsis is often accompanied by acute multi-organ injury.3,27,28 It has been shown that early organ injury in sepsis results from excessive inflammatory reactions, and uncontrolled inflammation is a key step in the progression of this disorder. 3
CD14 is involved in binding of LPS to TLR4 as well as signal transduction that activates immune cells.29,30 Inhibition of CD14 was successful in improving survival by attenuating thrombo-inflammation and cardiopulmonary dysfunction in a baboon model. 31 Furthermore, anti-rabbit CD14 monoclonal antibody exhibits protective effects on organ dysfunction and cell death in rabbits treated with endotoxemia. 22 CD14-deficient mice are more effective in inhibiting the spread of E. coli intraperitoneally inoculated than wildtype mice. 19 The above findings indicate that CD14-related LPS responses are meaningful targets for inhibiting inflammation in order to prevent tissue damage and organ dysfunction in septic shock. This work aimed to assess whether olaparib, a potent and selective PARP-1 inhibitor, could inhibit CD14 signaling to prevent multiple organ inflammation induced by sepsis.
Although olaparib was initially used in clinical practice as an anti-ovarian cancer drug, 32 it was subsequently shown to act as an on-oncological agent and a regulator of inflammation. 33 The distinction between the outcomes of preventive and therapeutic treatments is a good way to evaluate the potential of a drug. We examined whether olaparib treatment could protect mice against CLP-induced multi-organ injury and increase survival rate in septic animal models. As shown above, olaparib pre-treatment caused a higher inhibition of liver and lung injury compared with post-treatment, although no apparent differences were noted between the remarkable protective effects on renal injury conferred by pre- and post-treatment of olaparib. Olaparib treatment overtly restrained the expression of pro-inflammatory cytokines (IL-6 and TNF-α) in the tissues (lung, liver, and kidney) of septic mice, whereas such effect was not observed in serum. Unsynchronized detection times may lead to difference in IL-6 and TNF-α amounts between the tissues and serum samples. In addition, pre-treatment olaparib markedly decreased TNF- and IL-6 amounts in PLF of CLP mice, whereas post-treatment did not exhibit significant changes. Since neutrophils induce immunopathology during inflammation, olaparib’s profitable effects on different indicators or organ damage might be, directly or indirectly, related to neutrophil efferocytosis by macrophages.
Phagocytosis of neutrophils by macrophages exhibits certain anti-inflammatory effects, and uptake of chemotactic or pathogen-activated neutrophils can promote the anti-bacterial activity of macrophages. 34 Efferocytosis of neutrophils by macrophages boosts anti-inflammatory signaling, restrains neutrophil lysis, and dampens the immune response. 35 Overexpression of PARP-1 leads to enhanced cell death. Cytolysis products promote an inflammatory response. 5 The present study indicated that olaparib could promote efferocytosis of neutrophils by macrophages in septic mice, which may suppress excessive inflammation. The levels of macrophage phagocytosis influence their activation, differentiation, and death, as well as the inflammasome activation and the inflammatory factors release, which not completely control the progression and outcome of sepsis.34,36,37 Olaparib treatment decreased the total number of inflammatory cells in PLF, which was more consistent with the changes noted in inflammatory factors detected in PLF and local tissues. Persistent pathogens lead to amplify an inflammatory response, which is considered the pathophysiological basis of sepsis.3,38 Olaparib pre-treatment could decrease bacterial load in serum, PLF, and tissue (lung, liver, kidney, and spleen) samples from mice treated with CLP. The effects of olaparib may be associated with bacterial clearance caused by the efferocytosis of neutrophil by macrophages. Therefore, olaparib could alleviate inflammation, which is a critical consideration for ultimate clinical application.
In accordance with previous studies,19–22,27 we found that the expression of CD14 was enhanced in the organs (lung, liver, and kidney) of mice with sepsis as well as PMs. Olaparib treatment downregulated CD14 both in mice with sepsis and macrophages treated with LPS. The present study demonstrated for the first time that olaparib exerted a protective effect in sepsis by inhibiting CD14-mediated inflammatory response. The alternative mechanism of PARP-1 induction is mediated by direct interaction with phosphorylated ERK2,which in turn regulates epigenetic mechanisms that promote cell growth, proliferation, and differentiation. 14 It has also been confirmed that ERK inhibitors effectively block the interaction between PARP1 and ERK1/2. 39 Olaparib enhances the inhibition of the ERK pathway in non-small cell lung carcinoma. 15 The above data indicated that phosphorylated ERK levels were starkly reduced by olaparib in tissues (lung, liver, and kidney) after CLP. To further assess the mechanism of the CD14-ERK pathway in sepsis, the ERK inhibitor U0126 was administered in macrophages to examine the changes in CD14 expression. The results indicated that CD14 expression was significantly decreased by U0126 treatment. Olaparib exerted anti-inflammatory effects by inhibiting the CD14-ERK signaling pathway. The above findings provided novel insights into the mechanism by which olaparib exerts anti-inflammation, suggesting the potential use of olaparib for the treatment of sepsis.
PARP suppression may modulate sepsis severity by reducing the inflammatory response.6,7,33 In addition, olaparib had no side effects in the mice treated. Currently, PARP inhibitors are being assessed in clinical studies for specific conditions, including ovarian and non-small lung cancers.15,32 The present study demonstrated that post-treatment with olaparib inhibited organ damage in sepsis by reducing the release of inflammatory factors and promoting efferocytosis of apoptotic neutrophils. Furthermore, olaparib reduced the secretion of CD14 via ERK in sepsis, which provides a new perspective for its clinical application.
The CLP model was assessed vigorously in young healthy mice, with significantly higher mortality than noted in serious sepsis in humans. In preclinical and clinical studies, the efficacy of the anti-inflammatory agents in sepsis is compromised by the risk of death. 40 Differences in various parameters, including the type of factors that initiate sepsis, age and co-morbidities, have prevented the animal experiments from fully modeling human sepsis. 28 Therefore, a certain deviation exists in predicting human trial data for new agents based on animal models. Regarding the choice of olaparib dosing, many studies are not consistent, but are not very different. In the mouse model of intratracheal endotoxin (i.e.) administration, i.p. injection of olaparib at 5 mg/kg protects lung and kidney functions. 8 Olaparib can produce a clocking effect, i.e. olaparib exerts its maximum therapeutic effect at 5 mg/kg, and shows partial toxicity at 10 mg/kg, destroying the blood–brain barrier. 41 Our study lacked higher dose data, and follow-up studies should be performed.
Overall, the present study confirmed a new role of olaparib in facilitating negative regulation of ERK-mediated CD14 expression, which may contribute to the efficacy of olaparib in inhibiting multi-organ injury in sepsis.
Footnotes
AUTHORS’ CONTRIBUTIONS
ZXC, YHC, ZZS, and QL participated in the design. All authors participated in the experimental design, analysis of the data, interpretation of the results, and review of the manuscript. ZXC, YHC, ZZS, and QL wrote and revised the article.
DECLARATION OF CONFLICTING INTERESTS
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
FUNDING
The authors are appreciative of the financial support from the National Natural Science Foundation of China (grant numbers: 81772114 and 81971882).