
Abstract
Interleukin (IL)-33 is a tissue-derived nuclear cytokine belonging to the IL-1 family. Stimulation-2 (ST2) is the only known IL-33 receptor. ST2 signals mostly on immune cells found within tissues, such as regulatory T cells (Treg cells), CD8+ T cells, and natural killer (NK) cells. Therefore, the IL-33/ST2 signaling pathway is important in the immune system. IL-33 deficiency impairs Treg cell function. ST2 signaling is also increased in active Treg cells, providing a new approach for Treg-related immunotherapy. The IL-33/ST2 signaling pathway regulates multiple immune-related cells by activating various intracellular kinases and factors in the tumor microenvironment (TME). Here, we review the latest studies on the role of the IL-33/ST2 signaling pathway in TME and Treg immunotherapy.
Impact Statement
This review will be helpful in understanding the importance of the interleukin (IL)-33/Stimulation-2 (ST2) signaling pathway in immunotherapy and its role in a wide range of diseases. Understanding the relationship between IL-33/ST2 signaling and the tumor microenvironment and Treg immunotherapy is important, as it is undoubtedly a potential target for immunotherapy and clinical diseases.
Introduction
Regulatory T cells (Treg cells) are a T cell subset with immunosuppressive functions. These cells are found in almost all tissues and regulate several cell types involved in innate and adaptive immunity. They inhibit the immune response in the body, maintain immune balance, and prevent autoimmune diseases. Treg cells are generated in the thymus and migrate to peripheral tissues and organs. Treg cells have been shown to inhibit the activation and proliferation of potential autoreactive T cells in normal environments, which influence immunoregulation capability. Treg cells are characterized by expression of forkhead box protein P3 (FOXP3). Since their discovery in early 1995, Treg cells are marked by CD4+ CD25+ FOXP3+ status.1,2 Treg cells can be divided into natural regulatory T cells (nTregs) and peripherally induced T cells (pTregs). Expression of helios is a marker of nTregs that distinguishes these cells from pTregs. 3
Treg cells function through a variety of mechanisms: direct inhibition or lysis of target cells; inhibition of antigen-presenting cell maturation; secretion of anti-inflammatory cytokines, such as tumor growth factor (TGF)-β and interleukin (IL)-10; and negative regulation of cell-surface receptors, such as cytotoxic T lymphocyte antigen 4 (CTLA-4), CD39, and CD73. Thus, they participate in the regulation of inflammation and various immune diseases. 4 Treg cells form a heterogeneous population, exhibiting different migratory and immunomodulatory properties. They have shown promise as therapeutic targets in the fields of inflammation, tumor escape mechanisms, and immunotherapy.5,6 Immune escape is one of the important causes of tumor development. Treg cell numbers and functions are upregulated in tumors and in the microenvironment, which further reinforce their immunosuppressive effect in promoting tumor development. 6 The use of immune checkpoint therapies has brought new possibilities for tumor therapy, as these agents regulate Treg cells in various ways to deplete or block Treg cells. 7
As a member of the interleukin (IL)-1 family, IL-33 is strongly expressed in endothelial cells, epithelial cells, and fibroblasts, activating immune cells expressing the Stimulation-2 (ST2) receptor, such as Treg cells, NK cells, and CD8+ T cells. 8 IL-33 exerts its activity through a heterodimeric receptor consisting of its primary receptor ST2 and an accessory receptor, the IL-1 receptor (IL1RAP). IL-33 amplifies Treg cells by inhibiting effector T cells, but also by inhibiting the production of interferon (IFN)-γ by CD8+ T cells. 9 As a proinflammatory factor, IL-33 mediates antitumor immune activity through Th1 and NK cells and participates in a tumor immune escape mechanisms through T helper (Th)2 and Treg cells. IL-33 also promotes the activation of myeloid suppressor cells (MDSCs), innate lymphoid cells (ILC)-2, CD8+ T cells, and dendritic cells (DCs).9–12
ST2 is expressed more frequently in Treg cells than in non-immune cells. 13 There are three main subtypes of ST2: ST2L (transmembrane type), sST2 (soluble type), and ST2V (suppression of tumorigenicity 2-variant).14,15 ST2L binds to the alarmin IL-33 to trigger downstream signaling. sST2 negatively regulates the IL-33/ST2 signaling pathway by acting as a decoy receptor to prevent IL-33/ST2L binding.16,17
In this review, we summarize the findings of recent studies on the role of the IL-33/ST2 signaling pathway in tumor microenvironment (TME) and Treg cell-related immunotherapy.
The role of IL-33/ST2 signaling in the tumor microenvironment
Effect of IL-33/ST2 signaling on Treg cells
As a proinflammatory factor, IL-33 expresses an antitumor immune response through Th1 cells and NK cells, and plays a role in the tumor immune escape mechanism through Th2 cells and Treg cells, promoting the activation of MDSCs, intervening in ILC-2 cells, CD8+ T cells, and DCs.18,19 IL-33 is involved in maintaining FOXP3+ Treg cell homeostasis at mucosal sites and directly induces the proliferation of FOXP3+ Treg cells and enhances their immunosuppressive capacity.20,21 At a steady state, ST2+ Treg cells represent the majority of ST2-expressing CD4+ T cells. IL-33 is dispensable for the production, maintenance, and tissue accumulation of ST2+ Treg cells. Conventional mouse CD11c+ DCs are stimulated by IL-33. These cells then secrete IL-2, leading to selective amplification of ST2+-inhibitory CD4+ FOXP3+ Treg. 9 ST2+ Treg cells inhibit CD4+ T cell proliferation in vitro, independently of IL-33, and this inhibitory capacity is mediated partly by the enhanced production and activation of the anti-inflammatory cytokines IL-10 and TGF-β. 22 Moreover, IL-33/ST2 signaling enhances the activity of GATA-3 and STAT5, and recruits GATA-3 and RNA polymerase II to the FOXP3 promoter and IL1RL1 sites. Thus, IL-33 not only indirectly promotes FOXP3 production but also enhances the expression of its receptors.
IFN-γ is secreted by CD4 Th1 and CD8 cytotoxic T cells, which have a direct antiviral replication effect and are important activators of macrophages. IFN-γ expression is downregulated in the TME. As a factor that affects Treg cell function, IFN-γ affects Treg cell function by making these cells fragile, leading to the weakening of Treg function and thus alleviating immune suppression and immune cell function in the TME. 23 As early as 2008, IL-33 was found to enhance antigen-dependent and antigen-independent T-cell responses, including IL-5, IL-13, and IFN-γ production. IL-33 is a pleiotropic cytokine that acts similar to IL-18 under certain conditions, promoting IFN-γ production and type 1 immunity.24,25
According to the literature, in colonic Treg cells, IL-33/ST2 signaling can promote the expression of FOXP3+ and promote the function of Treg cells by promoting TGF-β1-mediated differentiation. 26 At present, it remains unclear whether FOXP3+ promotes antitumor immune function and combination therapy. A more in-depth understanding of Treg cells is necessary.27,28
A major hallmark of T-cell exhaustion is the enhanced expression of multiple immune checkpoint proteins, including programmed cell death protein 1 (PD-1), CTLA-4, lymphocyte activation gene-3 (LAG-3), T cell immunoglobulin and ITIM domain (TIGIT), and T cell immunoglobulin-3 (TIM-3). 29 In the tumor-infiltrating environment, T cells play a tumor-killing role, and a large number of Treg cells infiltrate tumor tissues where they exert an immunosuppressive function. Tumor-infiltrating Treg cells are mostly activated and rapidly reproduce. These cells express high levels of cell surface molecules associated with T-cell activation, such as CD25, CTLA-4, PD-1, LAG3, TIGIT, ICOS, and TNF receptor superfamily members, including 4-1BB, OX-40, and GITR (GRAPH1). 30 Increasing evidence suggests that the removal of Treg cells can induce and enhance antitumor immune responses. However, the unselective removal of large numbers of Treg cells may simultaneously trigger harmful autoimmunity. 31
Effector Treg cells are the major cell type in tumor tissues. These effector Treg cells specifically express various cell surface molecules, including chemokine receptors, such as CCR4. Blocking of candidate molecules on effector Treg cells by using specific monoclonal antibodies is a potential approach to ameliorate tumor progression. Furthermore, CTLA-4 is strongly expressed on effector Treg cells, causing active proliferation and apoptosis, and is used to control their function specifically. Immune checkpoint therapy is a rising research direction in cancer immunotherapy recently. Targeting PD-1 and CTLA-4, which are expressed on Treg cells, has resulted in a better outcome in cancer patients. CTLA-4, which is expressed on tumor-infiltrating Treg cells, downregulates the expression of antigen-presenting cells and CD80/CD86 co-stimulatory molecules on DCs, thereby preventing the activation of conventional T cells. Blocking immune checkpoints, including CTLA-4 and PD-1, in vivo has been shown to promote antitumor immunity by inhibiting Treg cells and enhancing effector T-cell function. In the TME, PD-1 is expressed on Treg cells and binds to PD-L1 expressed by tumor cells. This suppresses T-cell function, aggravating immunosuppression and promoting cancer development (Figure 1). 32
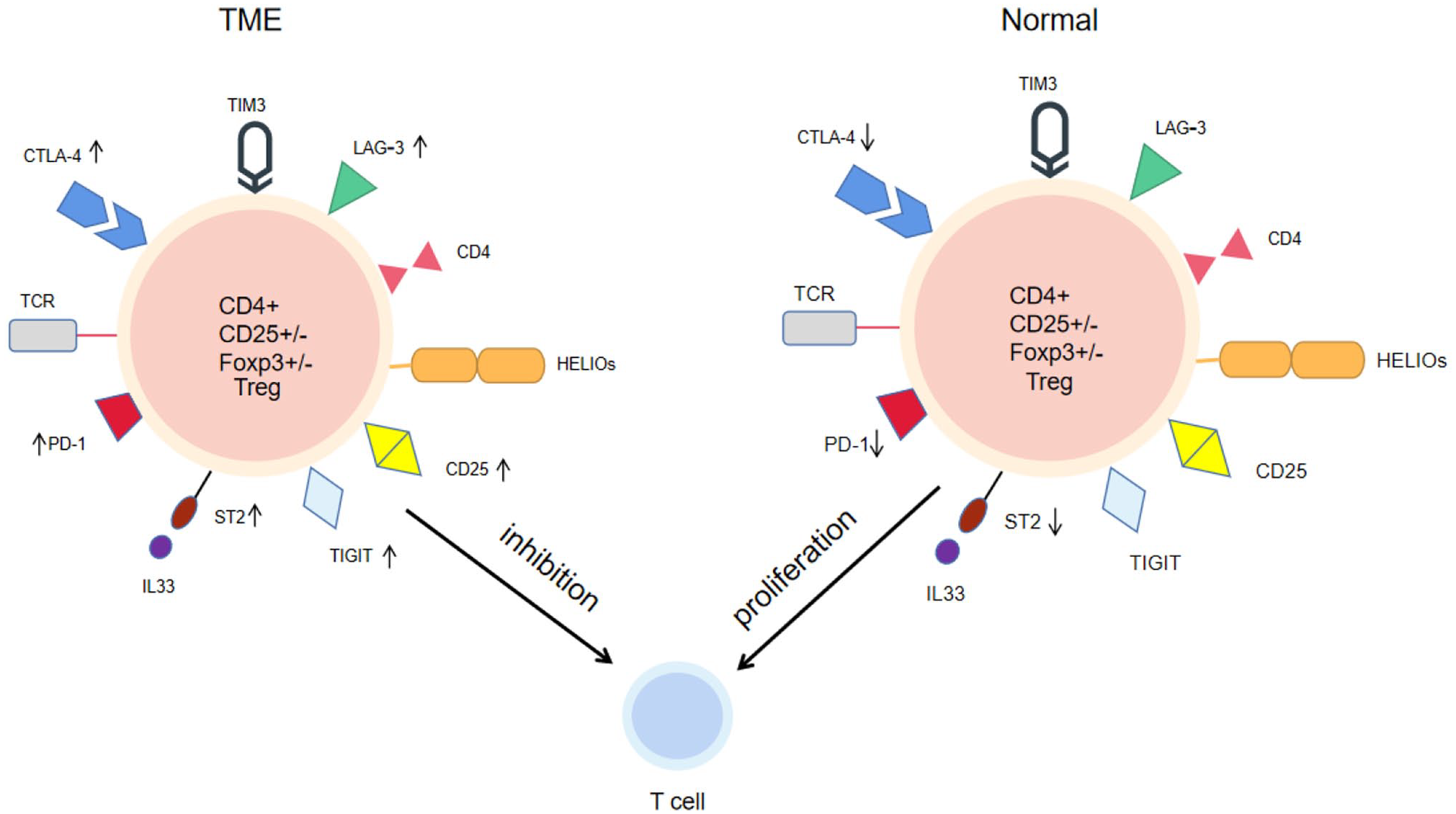
The figure shows the influence of IL-33/ST2 axis on Treg cells and surface molecules under normal physiological state and TME. In physiological state, IL-33/ST2 mainly maintains the balance of ST2+ Treg cells and maintains autoimmunity. In the TME, Treg cells surface molecular activity increased, such as CTLA-4 and PD-1. IL-33/ST2 may increase this risk and further enhance the number of Treg cells to inhibit T cell proliferation and activity (left). Inhibition of IL-33/ST2 axis and surface molecules could restore the number and activity of some T cells (right). (A color version of this figure is available in the online journal.)
Effect of IL-33/ST2 signaling on angiogenesis in the tumor microenvironment
Blood vessels are essential for tumor survival and development. During the occurrence and development of tumors, blood vessels supply large amounts of nutrients to the tumors and ensure the continuous existence of the TME. 33 The TME consists of tumor cells, surrounding cells, and secreted factors. Different TMEs have different backgrounds; therefore, IL-33/ST2 signaling may play various roles in tumor angiogenesis. IL-33 is a potent endothelial activator in blood vessels that promotes angiogenesis and vascular permeability. IL-33 regulates angiogenesis by promoting endothelial cell proliferation, differentiation, and morphological changes. These effects are inhibited by ST2 deficiency in vivo. In addition, IL-33/ST2 signaling increases endothelial permeability, downregulates occludin and VE-cadherin expression at mRNA level, and increases ICAM-1 expression. IL-33/ST2 signaling specifically disrupts the stability of the endothelial barrier, affects angiogenesis, and plays an important role in the TME.33–36
Effect of IL-33/ST2 signaling on natural killer cells in the tumor microenvironment
CD8+ T and natural killer (NK) cells are effective tumor-killing cells. In the TME, the number of these cells is reduced and their functions are inhibited, leading to aggressive tumor proliferation and progression. 37 IL-33/ST2 signaling can activate and recruit NK cells into the TME to clear tumor cells effectively and to reconstruct the microenvironment further. 38 IL-33/ST2 signaling is closely associated with NK cells and can induce NK cells to produce IFN-γ and release perforin and granzymes to kill tumor cells. IL-33/ST2 signaling can also enhance the function of CD8+ T cells and other immune-related cells, such as eosinophils and polarized T cells, induce the microenvironment, and support tumor regression.24,39–43 In IL-33-deficient mice, the number of NK cells is increased, indicating that disrupting the interaction between the IL-33/ST2 signaling pathway and Treg cells can significantly increase infiltration by NK cells. 44 Different amounts of IL-33 in the microenvironment have different effects. Low levels of IL-33 lead to immune tolerance, whereas high levels of IL-33 can enhance CD8+ T-cell functions. It can also be used as an immune adjuvant to enhance responses to vaccination.43,45
Treg cells in immunotherapy
Treg cells are considered important in tumor immune escape. These cells accumulate in large numbers in the TME. Different tumor types have different immunosuppressive microenvironments.31,46 Tregs are necessary for maintaining immune homeostasis and preventing autoimmune diseases in humans. Different subsets of Treg cells are involved in different responses and are necessary to understand the molecular pathways through which Treg cells operate in order to use these different pathways and responses to promote cancer immunotherapy.46–48
Immunotherapeutic effect of IL-10 on Treg cells
Treg cells alter the status of antitumor immune cells and affect tumor immune cell infiltration in the TME, which is a condition for tumor growth. 49 Many factors can affect the differentiation of Treg cells.48,50 IL-10 is a multifunctional factor that regulates the growth and differentiation of B, T, NK, and Treg cells. 51 Treg cells deficient in IL-10 receptors cannot maintain FOXP3 expression and have significantly downregulated expression of FOXP3-related factors, such as IFN-γ and other proinflammatory factors. These results indicate that IL-10 may act directly on Treg cells to promote their accumulation in the TME. 52 IL-10 is an important mediator of Treg cell-related immunity and regulates antitumor immune functions via IL-35. The loss of these two cytokines is restricted by Treg cells and has different effects on the TME. IL-10 is more important in limiting effector functions and proliferation, whereas IL-35 appears to limit memory T-cell differentiation. B lymphocyte-induced maturation protein 1 (BLIMP1) is a regulator of the terminal differentiation of T and B cells. IL-10 regulates BLIMP1 directly to affect the immune capacity of Treg cells. 53 In addition, IL-10 can regulate the effects of other targets of Treg immunotherapy, such as Th17 differentiation and STAT3 signaling.54,55
Therefore, IL-10 is a promising therapeutic target for Treg-based immunotherapy.
Effect of immune blockers on Treg cell immunotherapy
CTLA-4 plays a key role in T-cell-mediated immune tolerance. Blockade of CTLA-4 delays autoimmune diseases or downregulates the immunosuppressive effects of Treg cells.56–58 Traditionally, it has been thought that CTLA-4 and B7 interact to restrict the activation of naive T cells in lymphatic organs, leading to the failure of effector T cells in the TME. 59 However, this finding conflicts with recent studies claiming that binding of B7 to CTLA-4 may not be helpful in tumor immunotherapy. 60 As a co-inhibitory molecule, CTLA-4 negatively regulates T-cell activation. Blocking CTLA-4 can enhance activated T-cell signaling to achieve tumor immunotherapy.61,62 It seems that no single immune blocker can currently address the requirements for immunotherapy. At present, combined therapy is commonly used to control the development of cancer, such as the combination of PD-1 and CTLA-4 blockers, and the blockade of CTLA-4 and the IL15/IL15Rα complex can kill tumor cells by enhancing the activity of NK cells. 63 Combination therapy increases toxicity, including diarrhea, colitis, hepatitis, and endocrine diseases, and also improves therapeutic outcomes.64,65 In addition, Treg cells can develop resistance to immune checkpoint inhibitors, which may be due to changes in the TME that reduce the efficacy of immune checkpoint inhibitors. For example, some tumors produce more antigens that enhance Treg cell function and mediate immune escape.66,67 Thus, further improvement of potential combination therapies is needed.
Effect of Th17 cells on Treg cell immunotherapy
The balance between Th17 and Treg cells is an important factor in immune homeostasis. Th17 cells not only participate in the host defense against pathogens, but are also involved in autoimmune diseases and cancer.68,69 Th17 cells differentiate from naive CD4+ T cells and secrete IL-17. Th17 cells play multiple roles in inflammation, infection, and immune balance. 70 Both Treg and Th17 cells differentiate from naive CD4+ T cells, but have opposite functions. Th17 cells are involved in inflammation, autoimmunity, and cancer, whereas Treg cells inhibit immune responses. Previous studies have shown that TGF-β, IL-6, and IL-1β are important factors associated with Th17 cell differentiation. 71 Treg cells can express IL-10, TGF-β, and other factors. 72 The immunosuppressive ability of Tregs is regulated by promoting or inhibiting the differentiation of Th17 cells, which play an important role in autoimmune diseases, cancer immunotherapy, and immune homeostasis.68,73 Recent studies have shown that regulating the balance between Th17 cells and Treg cells can lead to effective treatments, such as using bone marrow mesenchymal stem cells (BMSCs) to balance Th17 and Treg cells through the FOXP3/RORγt pathway in chronic regenerative anemia. 74 An increasing number of clinical studies have focused on the effects of the Th17/Treg axis on inflammation, autoimmunity, and cancer therapy, and its role in conditions such as systemic lupus erythematosus disease. 75 The results of those studies indicated that Th17 cells are important factors in Treg immunotherapy.
Immune-associated role of IL-33/ST2 in the TME and Treg cells
Increasing evidence suggests that the IL-33/ST2 axis in Treg cells is one of the key pathways responsible for preferential accumulation of Treg cells in the TME. Proliferation-related, immunosuppression-related, and cytokine/chemokine receptor genes are upregulated in tumor-infiltrating Treg cells, suggesting that the IL-33/ST2 axis may be a potential therapeutic target for cancer immunotherapy. 76 As a negative regulator, soluble ST2 suppresses tumor growth and metastasis by inhibiting IL-33-induced angiogenesis, Th1 and Th2 immunoreactivity, macrophage infiltration, and M2 polarization. IL-33-deficient Treg cells exhibit attenuated inhibitory abilities in vivo and promote tumor regression in an ST2-independent manner. 5 In a murine pancreatic cancer model, cancer cell-derived sST2 enhanced tumor growth by upregulating CXCL3 via IL-33/ST2L signaling in the microenvironment of pancreatic cancer. These results suggest that sST2 and CXCL3/CXCR2 could be considered as therapeutic targets. 77 Studies on IL-33 in a murine breast cancer model showed that loss of ST2 in BALB/c mice decreases tumor growth and metastasis, accompanied by increased serum levels of cytokines such as IFN-γ, TNF-β, IL-17, and IL-4.78,79 The IL-33/ST2 signaling pathway controls complex cytokine-related pathways in the TME, with direct or indirect effects on Treg cells, such as enhancing blocking of ST2+ Treg cell proliferation by IFN-γ and IL-33. ST2 is constitutively expressed in Treg cells, depending on the tissue and disease type. IL-33/ST2 recruits Treg cells to the TME and activates their immunosuppressive function, supporting tumor immune escape and favoring tumor growth. 80 The interaction of other chemokine receptors on Treg cells with chemokines in tumors, such as CCR4 and CCL12, CCR4 and CCL17, CCR10 and CCL28, and CXCR4 and CXCL1, has been reported to be involved in the recruitment of Treg cells to tumors. 30 In esophageal squamous cell carcinoma, IL-33 stimulates CCL2 production by involving TGF-β for recruitment by Treg cells. 81 For antibody-mediated killing of tumor-infiltrating Treg cells, surface molecules that are specifically expressed on tumor Treg cells or are expressed at much higher levels in tumor Treg cells than on other T cells are good targets. These markers include the aforementioned T helper cell molecules, such as CD25, CTLA-4, GITR, 4-1BB, OX-40, LAG3, TIGIT, CCR4, and CCR8.
In short, there are similarities between targeting the IL-33/ST2 axis or Treg cells. Differential molecular expression and phenotypes play a unique role in cancer immunotherapy. Immunotherapy using targeted molecules should be further investigated.
Colorectal cancer
Epigenetic reprogramming of IL-33/ST2 in the TME is an important factor in maintaining the TME. Genetic defects in IL33 have shown benefits to immune efficacy. 5
Colorectal cancer (CRC) is a life-threatening condition. FOXP3+ Treg cells have been shown to play a limited and complex role in CRC through heterogeneity in both functional and genetic phenotypes. 82 In addition, the expression of IL-33/ST2 increases the activation of FOXP3+ Treg cells in CRC. Tumor-infiltrating Treg cells preferentially express increased levels of ST2. The IL-33/ST2 signaling pathway is proportional to the number of tumors. Pathological analysis has shown that the increase in ST2-positive cells is related to the accumulation of Treg cells in the CRC microenvironment.13,83,84 Interestingly, IL-33/ST2 signaling not only promotes the development of CRC but also inhibits its progression by inducing IFNG expression through NF-κB. 85 IL-33 has been shown to play dual roles in CRC; however, its exact mechanism of action remains unknown. 86
Acute myeloid leukemia
The IL-33/ST2 signaling pathway is also associated with hematologic malignancies. 87 The most common abnormality in acute myeloid leukemia (AML) is the inversion of chromosome 16, resulting in the fusion gene CBFB-MYH11.88,89 Recent studies have shown that IL-33/ST2 is dynamically increased in CBFB-MYH11+ leukemia cells, promoting their survival. IL1RL1, a key transcription factor for ST2, may be an important regulator of CBFB-MYH11+ LSCs. 90 AML exhibits a variety of immunosuppressive activities, such as the upregulation of Treg cells and of PD-1. IL-33/ST2 signaling promotes the effects of PD-1 blockers, further improving immunotherapeutic efficacy.91,92 PD-L1 in the TME increases FOXP3 expression and thus its immunosuppressive effects. 93 The combination of PD-L1 blockade and targeting of the IL-33/ST2 signaling pathway in the TME greatly reduces the infiltration of Treg cells and constitutes a potential immunotherapeutic strategy. 94 Targeting the IL-33/ST2 pathway has been explored in AML and has a robust rationale in related fields. 95
Gastric cancer
Gastric cancer (GC) is a serious disease with high morbidity and mortality rates. The IL-33/ST2 signaling pathway also plays an important role in gastric cancer progression.96,97 Recent studies have shown that the levels of ST2, soluble ST2, and the ST2V subtype are significantly reduced in cancer tissues as compared to noncancerous tissues in GC patients. The expression of ST2 in GC tissues was found to be significantly lower than that in adjacent tissues, suggesting that reduced expression of ST2 is related to the occurrence and development of GC. Thus, ST2 may be a potential molecular marker of GC development. 98 IL-33 stimulates mast cells in the GC TME to produce IL-11, which is necessary for the growth and development of GC. Inhibition of the IL-33/ST2 signaling pathway restricts tumor growth and reduces the production and release of the mast cell-dependent macrophage attractants CSF-2, CCL-3, and IL-6. Tumor-derived IL-33 triggers a mast cell‒macrophage-dependent signaling cascade, suggesting the potential value of IL-33 in GC treatment. 99 Treg cells are also involved in the complex TME of GC. Recent studies have shown that the number of CD4+ FOXP3+ Treg cells is positively correlated with the expression of FOXM-1 and Ki-67, which are related to immunosuppression in GC. 100 The formation of a complex TME, including CD4+/CD8+ T cells and FOXP3+ Treg cells, ultimately affects the progression and survival of GC. 101 In summary, the IL-33/ST2 signaling pathway triggers mast cells to produce IL-11, which induces the occurrence and development of GC. Gastric mast cells also produce PD-L1 to enhance the immunosuppressive ability of Treg cells. Inhibition of the IL-33/ST2 pathway may further enhance the clinical efficacy of immune checkpoint therapy for GC.
Discussion
The components of the microenvironments of different tumors are complex but closely related. The IL-33/ST2 pathway not only promotes the occurrence and development of tumors but also inhibits related activities. The regulatory effects of Treg cells are context-dependent. Different contexts are related to different regulatory functions, such as promotion of M2 macrophage differentiation, changed epigenetic reprogramming of IFN-γ, and other factors that can accelerate tumor growth. The IL-33/ST2 signaling pathway regulates Treg cell function and thus plays a role in tumor inhibition. Recent studies have shown that IL-33 induces the transformation of tumor cells into polyploid giant cells in the TME, directly or indirectly promoting tumor metastasis and development. These data suggest that IL-33 is a key driver of drug resistance in tumors. 102 The IL-33/ST2 signaling pathway not only regulates a variety of immune cells but also changes the diversity of Treg cells, which is one of the important pathways causing Treg cell enrichment in the TME.5,6,103,104 In addition to immunosuppressive Treg cells, IL-33/ST2 signaling can recruit other immune cells, such as NK and CD8+ T cells, to reconstruct the TME. This can block tumor angiogenesis to prevent tumors from absorbing nutrients.
Treg cells are important factors in maintaining the immune balance and accumulate in large quantities in the TME and relevant target tissues. The IL-33/ST2 signaling pathway can promote TGF-β1-mediated Treg differentiation, stimulate DCs to secrete IL-2, and selectively amplify ST2+ Treg cells.9,26 An increasing number of studies have focused on the genes, functions, and phenotypes of Treg cell per se. 105 Previous studies have shown that ILC-2 also regulate Treg cell activity through amphiregulin. IL-33 has been reported to promote the tumor-promoting activity of ILC-2.106,107 The functional expression of Treg cells is affected by various components of the TME, including IL-10 and Th17 cells. IL-10 regulates the proliferation and differentiation of immune-related cells and affects immune functions. IL-10 can directly regulate BLIMP1 to affect the immune function of Treg cells. The combination of IL-10 and IL-35 promotes tumor regression by affecting proliferation and differentiation. IL-10 mediates the differentiation of Th17 cells, which are important factors in immune homeostasis and are involved in immune regulation, cancer progression, and inflammation. Th17 cells enhance immunity and achieve therapeutic effects through the balance with Treg cell pathways. The functions of Th17 cells in autoimmune diseases, immune disorders, and cancers have been widely studied.
Many studies have focused on the inseparable relationship between the IL-33/ST2 signaling pathway and Treg cells. IL-33/ST2 signaling regulates various immune cells and inflammatory cytokines. It is associated with many immune system diseases by suppressing or promoting immune functions.
Blocking IL-33 and its pathway exerts antitumor effects. IL-33/ST2 signaling is not only a downstream signal for immunotherapy but also a target for tumor therapy. 108 There are many links between IL-33/ST2 signaling and Treg cells. Studies have focused on indirect inhibition of the TME by MDSCs or on protumor effects by increasing Treg cells by promoting IL-2 secretion.9,109,110 IL-33/ST2 signaling promotes FOXP3 expression and enhances Treg cell function in some contexts. The IL-33/ST2 signaling pathway can also regulate other genes to enhance or inhibit the expression of related genes. In CRC, IL-33/ST2 signaling increases the activated phenotypes of FOXP3+ Treg cells, increases Treg cell accumulation in the TME, and impairs immune functions. However, whether FOXP3 has antitumor effects requires further investigation. In GC, tumor-derived IL-33/ST2 signaling triggers intracellular signaling cascades in mast cells and macrophages. The IL-33/ST2 signaling pathway is dynamically expressed in leukemia cells and promotes their survival. Epigenetic reprogramming is a difficult aspect of treatment that warrants further study.
The development of different treatment modalities according to different tumor backgrounds, identification of common cancer treatment targets against common backgrounds, and investigation of signaling molecules or pathways associated with key factors are still ongoing. In future studies, more associations and roles of the IL-33/ST2 signaling pathway and Treg cells in the TME are likely to be explored.
Footnotes
Authors’ Contributions
SL, LZ, XZ, JJ, GQ, and JY devised and wrote the manuscript. All authors contributed to critical revisions of the manuscript. SL and JJ contributed equally to this paper.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by National Natural Science Foundation of China (grant nos 82060034, 82100234, 32060179), Guangxi Natural Science Foundation Project (grant no. 2020GXNSFBA297004), Guangxi University Middle-aged and Young Teachers’ Basic Research Ability Improvement Project (grant no. 2020KY12014), and Independent project of Guangxi Key Laboratory of Tumor Immunity and Microenvironment Regulation (grant no. 203030302007).