
Abstract
Sporulating bacteria such as Bacillus spp. have contributed to severity of opportunistic hospital acquired infections, including postoperative wounds and respiratory tract infections. This study determines the expression profiles of sporulation markers in multidrug-resistant Bacillus spp. isolated from Ghanaian hospital environments. Antimicrobial resistance (AMR) profiles of the bacteria were determined with disk diffusion and broth microdilution. Primer-specific polymerase chain reaction (PCR) amplification was used to profile the sporulation markers, and quantitative reverse transcription polymerase chain reaction (RT-qPCR) was used for the expression of the sporulation markers at different antibiotic concentrations. The strains are multidrug resistant (70–100%) to at least two of the eight classes of the antibiotics tested including cephalosporins, penicillin, aminoglycosides, and glycopeptide. The strains showed different resistance patterns to all the tested antibiotics, which might indicate diverse resistance mechanisms. Common (spoVK spoVE, spoJ, and sigF) and not commonly (sigJ, soJ, yrbC, and yjcE) reported sporulation markers were detected in the strains. The study showed an association of the sporulation markers with AMR as indicated by their expression profiles.
Impact statement
Sporulation genes have been limited to inducing sporulation or spore formation during extreme environmental conditions; however, these genes could also be implicated in antimicrobial resistance (AMR). They could serve as potential “resistance” markers and obstruct antibiotic action. Although, the phenotypic property of spores may not be evident during antibiotic treatment; however, the expression of the markers could facilitate strain survival in the presence of antibiotics. Generally, little emphasis has been placed on the role of sporulation markers mediating AMR, this study provided insights that can possibly bridge this gap.
Introduction
Bacteria have developed diverse survival mechanisms including spore formation, and this aids the bacteria to display resistance to antimicrobial agents, chemicals, and ultraviolet (UV) radiation. 1 Bacillus species particularly B. subtilis, B. cereus, and B. thuringiensis are known for rapid spore formation. 2 Bacillus subtilis as a model spore-forming bacterium differentiates into a dormant stress-resistant spore in harsh environmental conditions, such as high temperature and pressure, nutrient deprivation, and carbon or nitrogen starvation.3,4 Spore of various Bacillus species are resistant to conventional antibiotics, could cause food poisoning and human diseases, making sporulating bacteria a serious public health concern.4,5
In hospital environments where diverse bacteria are emerging and resistance to antibiotics is on the rise, 6 sporulation has been associated with hospital acquired infections including postoperative wound, sepsis, bacteremia, respiratory tract, and other opportunistic infections. 7 While sporulation helps bacterial reproduction, in enclosed hospital intensive care unit (ICU), sporulating bacteria takes advantage of environmental dispersal agents such as fomites for spore propagation and distribution. 8 Invasive devices and other medical handy-tools can harbor spores and spread infectious agents to immunocompromised and critically ill inpatients in the ICU. 8
Bacillus subtilis can survive for months on hospital flat surfaces, and its sporulation machinery has been traced to its genetic components. 9 Sporulation genes such as SpoIIIE, SpoVE, SpoVR, and SpoVK are turned-on when B. subtilis is exposed to environmental stresses including overuse of antibiotics, a common practice in the ICU. 10 These genes and a few others are involved in different spore formation stages upon expression which regulates sporulation and contributes to bacterial virulence and antimicrobial resistance (AMR).10 –12 In Ghanaian hospital environments where diverse bacterial pathogens are emerging including Bacillus species, this study aims to characterize sporulation markers and determine AMR expression profiles of Bacillus species isolated from hospital environments in Ghana.
Materials and methods
Bacterial strains information
Archived environmental B. subtilis and B. cereus were used in this study. The strains were obtained from AbiMosi™ Bacterial Culture Library at West African Centre for Cell Biology of Infectious Pathogens at the University of Ghana, where the study was also conducted. The strains, isolated from Ghanaian hospital fomites, were identified using MALDI-TOF and 16S rRNA amplification previously described by Schröttner et al. 13 All the strains were revived in Luria Bertani media (37°C for 24 h, 5% CO2) (Oxoid, England, CM0003) at the onset of the experiments.
AMR profiling
Antibiotic susceptibility test was performed using the standard Kirby–Bauer disk diffusion previously described Abiola et al. 14 Briefly, the test strains were grown overnight in sterile Mueller–Hinton broth (Sigma M7408-500G; PCode: 1001876334) and optical density was standardized to 0.5 at 600 nm. Sterile Mueller–Hinton agar plates (Oxoid, Basingstoke, UK) were seeded with the standardized culture and antibiotic disks (vancomycin [30 µg], trimethoprim [25 µg], penicillin [15 µg], amoxicillin [10 µg], ampicillin [10 µg], erythromycin [15 µg], chloramphenicol [30 µg], gentamicin [10 µg], cloxacillin [10 µg], tetracycline [30 µg], ceftriaxone [10 µg]) (Mast Diagnostics, Mast Group Ltd., Merseyside, UK) were aseptically placed on the culture plates (incubation, 37°C for 24 h). The zone of inhibition was measured with a meter rule (CLSI, 12th Edition) and data presented represents two independent experiments with three replicates. B. subtilis ATCC 23857 was used as a standard control. Percentage resistance was expressed as the ratio of antibiotics resisted (resistance to available antibiotics) by the test strains to the total number of antibiotics tested or available as treatment option multiplied by 100. Also, fractional resistance to the tested antibiotics was determined with a broth microdilution and spotting assay. Briefly, 100 µL of the overnight Mueller–Hinton cultured strains standardized to 0.5 OD (600 nm) was added to 100 µL of antibiotics in a 96-well plate at test concentrations previously stated and incubated overnight at 37°C. OD was determined relative to the strain in the control wells (without the antibiotics) to determine bacterial survival in the presence of the antibiotics. 5–10 µL of 0.1 v/v of the culture in the resistant wells was spotted on Mueller–Hinton agar; colonies were counted relative to the controls. The fractional resistance for each of the antibiotics was expressed as the percentage of colonies from the antibiotic resistant wells relative to the control without the antibiotics.
Sporulation genes profiling
Sporulation markers yrbC, sigF, sigA, spsE, spovT, spoVK, spo0J, spoIIIE, cotA, spsK, cotSA, spoVE, soJ, spoIIIAB, spo0M, spsJ, cgeE, and spo0A were profiled with polymerase chain reaction (PCR). Bacterial DNA extraction kits (Zymo Research, Irvine, CA, USA) and sporulation genes specific primers (S1) were synthesized by Inqaba Biotech (South Africa). The PCR amplification reaction mixture was a total volume of 25 μL, containing 5 μL of PCR buffer, 0.5 μL of deoxynucleoside triphosphates (dNTPs), 0.5 μL of forward and reverse primer each, 15.87 μL of distilled water, 0.13 μL of Taq polymerase, and 2.5 μL of template DNA. Negative control was the reaction buffer without the DNA template and positive control was the PCR amplicon from B. subtilis ATCC 23857. The PCR conditions included initial enzyme activation at 95°C for 10 min, followed by 40 cycles of amplicon denaturation at 95°C for 15 s, primer annealing at 60–65°C for 30 s to 1 min and elongation at 60°C for 1 min with final extension at 60°C for 1 min. 15 Amplification was performed in a thermocycler (Biometra, Gottingen, Germany). PCR products were separated on 1% agarose gel stained with 5 μL of Gel Red (Biotium – Glowing Products for Science, Fremont, CA, USA). The amplified PCR products were visualized under the UV transilluminator Imager (Amershan™ Imager 600, Tokyo, Japan).
Determination of minimum inhibitory concentrations and antibiotic stress assay
To determine the levels of AMR and if the sporulation markers contribute to resistance, minimum inhibitory concentrations of the strains at different antibiotic concentrations were determined with broth microdilution. Briefly, the strains were cultured in Mueller–Hinton broth overnight at 37°C and standardized to OD of 0.5 (600 nm). Following the CLSI standard as previously indicated, 100 µL of the culture was added to 100 µL of double concentrations of antibiotics at 10–160 µg/mL (cloxacillin, ceftriaxone, amoxicillin and ampicillin), 15–120 µg/mL (penicillin and erythromycin), 25–200 µg/mL (trimethoprim), 30–240 µg/mL (tetracycline, gentamicin, chloramphenicol, and vancomycin) in a 96-well plate (incubation at 37°C, 24 h, 120 r/min). Minimum inhibitory concentration was determined using the percentage of the OD (600 nm) of test strain to OD of control. The percentage OD < 10 was the assigned minimum inhibitory concentration (MIC) of the strains at the specific concentration. To determine if the sporulation genes are expressed and induced sporulation under antibiotic stress, the strains were exposed to high concentrations of antibiotics to which they displayed 100% levels of resistance and higher minimum inhibitory concentrations. The strains were exposed to 2× and 4× MIC in broth microdilution as previously stated. Subsequently, the stressed variants were harvested by spreading 10 µL of the culture on LB agar plate and incubated overnight at 37°C. Highly resistant colonies of the survived isolates were selected for RNA extraction.
RNA extraction and gene expression profile of sporulation markers
Total RNA was extracted from the multidrug-resistant B. cereus and B. subtilis with Trizol. Briefly, the cells were harvested in 1 mL of Trizol (TRIzol™, Invitrogen, Waltham, MA, USA) and incubated at room temperature for 5 min. 200 µL of chloroform was added and vigorously shaken for 15 s. Subsequently, the samples were incubated at room temperature for 2–5 min and centrifuged at 12,000g for 15 min at 4°C. The upper layer was transferred into 2 mL Eppendorf tube and 500 µL of absolute isopropyl alcohol was added, vortexed, and incubated at room temperature for 10 min to precipitate the RNA. The samples were centrifuged (12,000g, 10 min at 4°C); pellets washed in 1 mL of 75% ethanol and air dried (20 min). The pellets were suspended in RNase-free DEPC-treated H2O and stored at −20°C. Quantitative reverse transcription polymerase chain reaction (RT-qPCR) was used to determine the gene expression profiles of sporulation markers. The differential expression of each gene was determined using the Luna® Universal One-Step RT-qPCR Kit. 20 µL reaction mix for each sample includes 10 µL of reaction buffer, 1 µL of RT enzyme, 0.8 µL each of forward and reverse primers, 5.4 µL of nuclease-free water, and 2 µL of RNA. The QuantStudio (version 5) was used to analyze the gene expression profiles. Reverse transcription was performed at 55°C for 10 min, followed by initial denaturation at 95°C for 1 min, denaturation at 95°C for 15 s, extension at 60°C for 15 s, and melt curve at 60°C.
Statistical analysis
Descriptive statistics were used in this study (with SPSS 16.0 and GraphPad 6.0), and the data were presented in graphs. Two-way analysis of variance (ANOVA) and correlation analysis were done to compare the level of resistance of the bacterial strains to the tested antibiotics, resistance patterns, and distribution of the sporulation markers relative to the control. P < 0.05 was considered as significance level.
Results
Bacillus species are multidrug resistant
All the bacterial strains showed resistance to at least four of the ten antibiotics tested relative to the ATCC 23857 control strain. The highest level of resistance was to cloxacillin, amoxicillin, tetracycline, and ampicillin (Figure 1). The strains showed between 70% and 100% levels of resistance to all the tested antibiotics, with complete resistance (100%) to cloxacillin, amoxicillin, tetracycline, and ampicillin. The antibiotics showed considerable level of effectiveness against the ATCC 23857 as compared to the tested strains. Overall, the average percentage level of resistance to all the 11 antibiotics tested was 75% with no individual strain susceptible to all the antibiotics. The highest level of resistance was displayed by B. subtilis; however, all the strains were multidrug resistant to at least two of the classes of antibiotics tested with different resistance patterns. The minimum inhibitory concentrations vary across the tested antibiotics with 160–480 µg/mL for cloxacillin, amoxicillin, ampicillin, and tetracycline (Table 1). The rest of other antibiotics tested have an MIC range of 60–120 µg/ml with respect to the recommended CLSI standards relative to the ATCC control with 20 µg/ml as the least and 60 µg/ml as the highest (in vancomycin only).
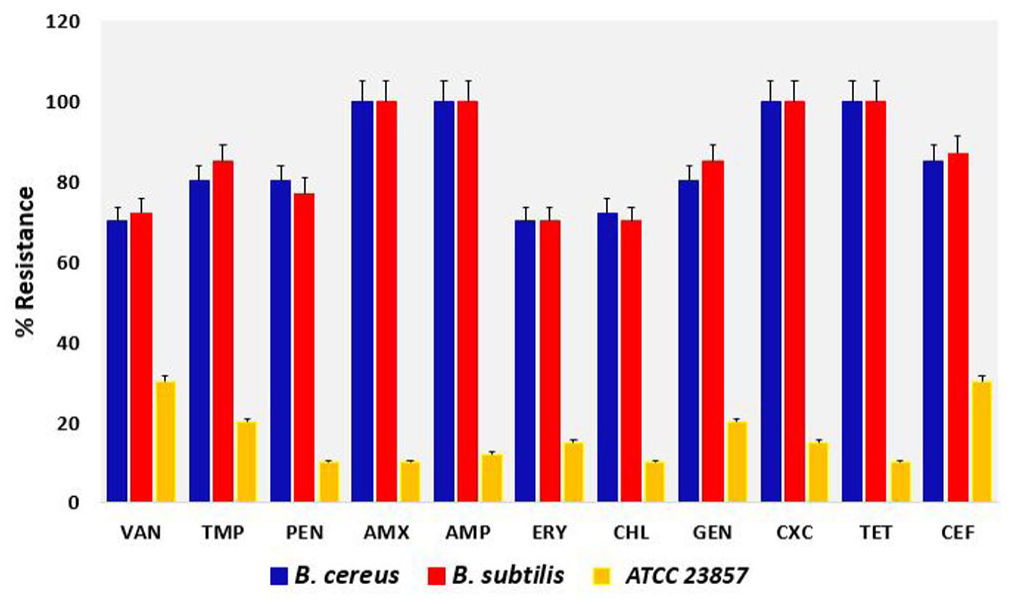
Resistance profiles of the bacterial strains to the seven antibiotics tested relative to the B. subtilis ATCC 23857 control with P = 0.002 using two-way ANOVA.
Minimum inhibitory concentrations of the strains.
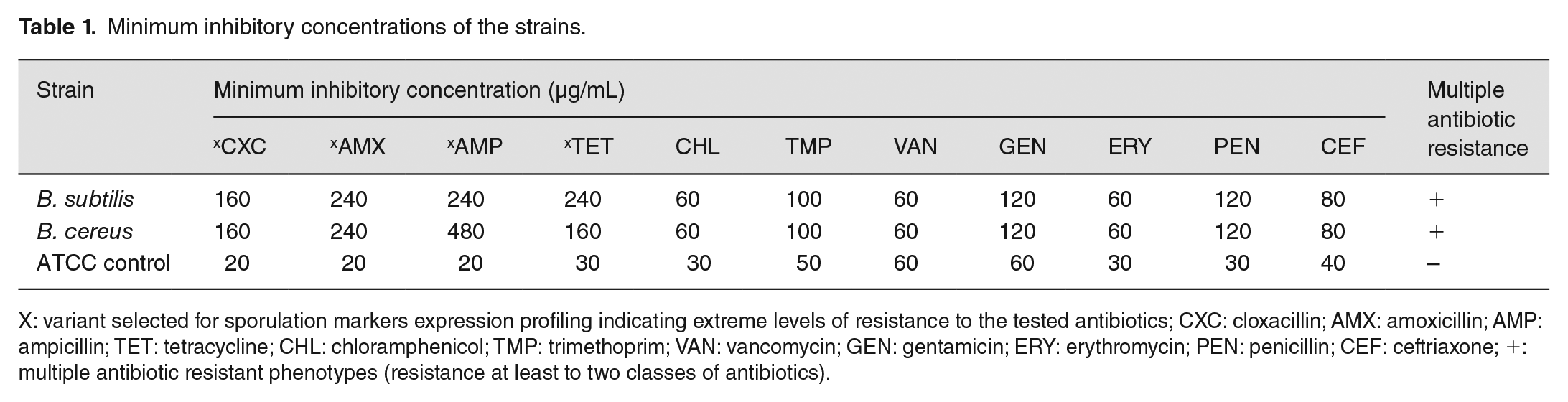
X: variant selected for sporulation markers expression profiling indicating extreme levels of resistance to the tested antibiotics; CXC: cloxacillin; AMX: amoxicillin; AMP: ampicillin; TET: tetracycline; CHL: chloramphenicol; TMP: trimethoprim; VAN: vancomycin; GEN: gentamicin; ERY: erythromycin; PEN: penicillin; CEF: ceftriaxone; +: multiple antibiotic resistant phenotypes (resistance at least to two classes of antibiotics).
Bacillus species have multiple sporulation genes
All the strains, B. subtilis and B. cereus were screened for the presence of sporulation markers yrbC, sigF, sigA, spsE, spovT, spoVK, spo0J, spoIIIE, cotA, cotD, spsK, cotSA, spoVE, soJ, spoIIIAB, spo0M, spsJ, cgeE, and spo0A with PCR-primer specific amplification. yrbC, sigA, sigF, spoVK, cotD, spo0J, soJ, spoIIIAB, spo0M, spsJ, cgeE, and spo0A (Table 2) were detected in all the strains. In addition to these genes, B. subtilis have spoVT, spoIIIE, cotA, spsK, and cotSA. Also, spoVE was not detected in all the strains including the control and spoVT, spoIIIE, and spsK only in B. subtilis. However, the strains harbors 14 (70%) of the 20 sporulation genes screened with yrbC, sigA, sigF, cotD, spo0J, soJ, spo0M, spsJ, cgeE, and spo0A equally distributed in all the strains analyzed (Table 2).
Detection of sporulation markers.
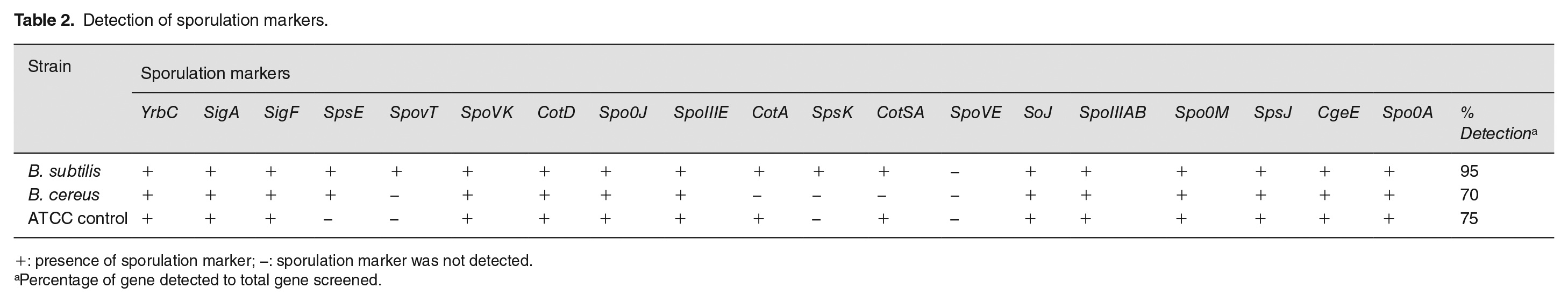
+: presence of sporulation marker; −: sporulation marker was not detected.
Percentage of gene detected to total gene screened.
Increase in the expression levels of sporulation markers in response to antibiotic stress
To determine the role of sporulation markers in bacterial resistance profile and associated sporulation, the expression profile of eight sporulation markers, including SigF, SpoVK, CotD, Spo0J, SoJ, SpsJ, CgeE, and Spo0A was monitored in B. subtilis and B. cereus. Relative to the control, seven of the sporulation markers were highly expressed in B. subtilis (Figure 2) in the presence of cloxacillin, amoxicillin, tetracycline (160–480 µg/mL, minimum inhibitory concentrations), and only one in ampicillin at the same concentration. However, the genes were differentially expressed in B. cereus (Figure 3). The expressions of all eight genes were increased in response to cloxacillin, while Spo0J, SoJ, SpsJ, and CgeE were moderately increased after exposure to amoxicillin, tetracycline, and ampicillin. Most of the markers in B. cereus showed a basal level of expression in response to amoxicillin, tetracycline, and ampicillin as compared to B. subtilis (Figure 4).
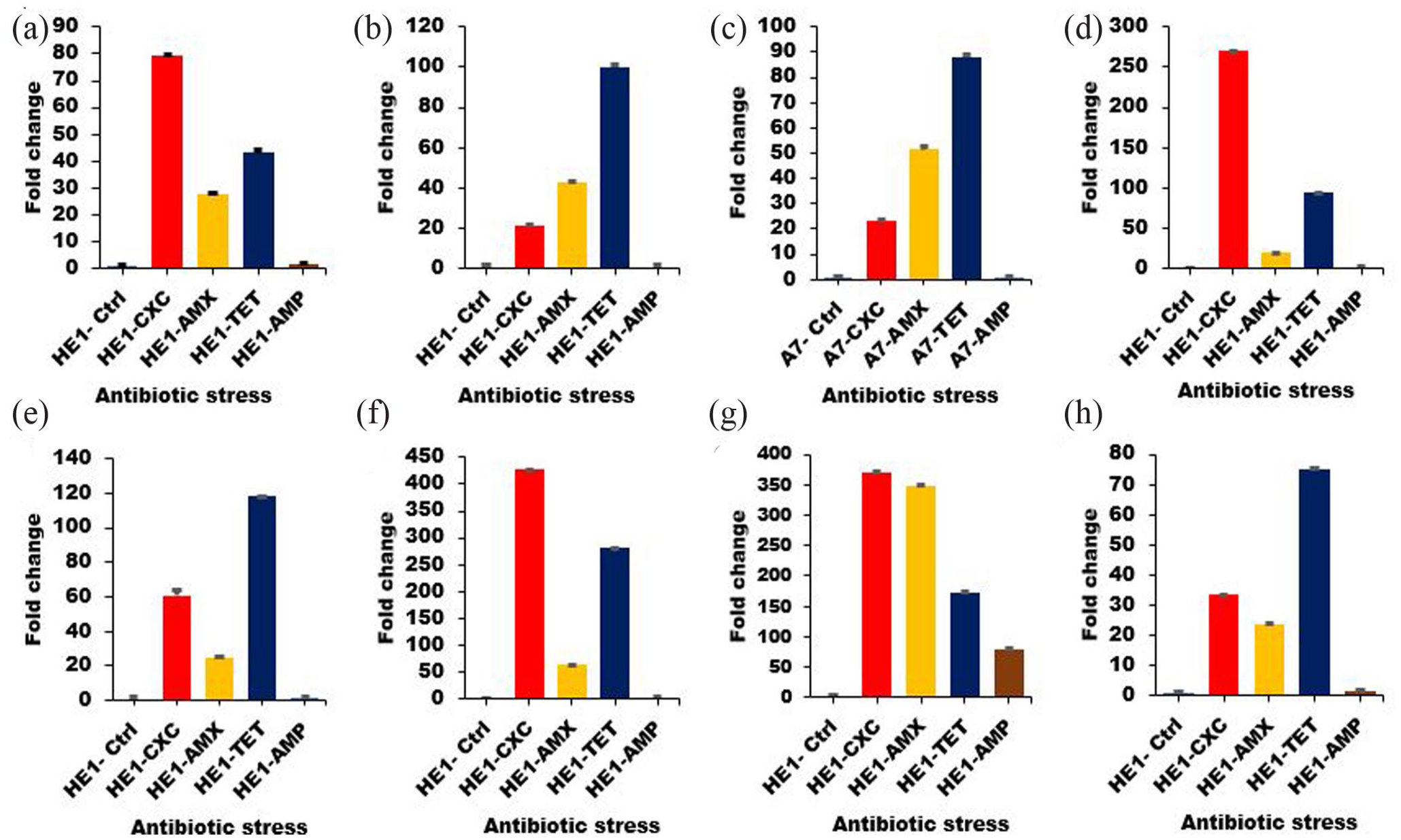
Expression profile of sporulation markers in B. subtilis (HE1) after exposure to antibiotics. The isolates were exposed to different antibiotics: cloxacillin (HE1-CXC), amoxicillin (HE1-AMX), tetracycline (HE1-TET), and ampicillin (HE1-AMP). The expression profile of eight genes was monitored following exposure to stress: (a) SigF, (b) SpoVK, (c) CotD, (d) Spo0J, (e) SoJ, (f) SpsJ, (g) CgeE, and (h) Spo0A. The samples were analyzed in triplicate using the delta-delta Ct method, and the experimental variations are shown as error bars.
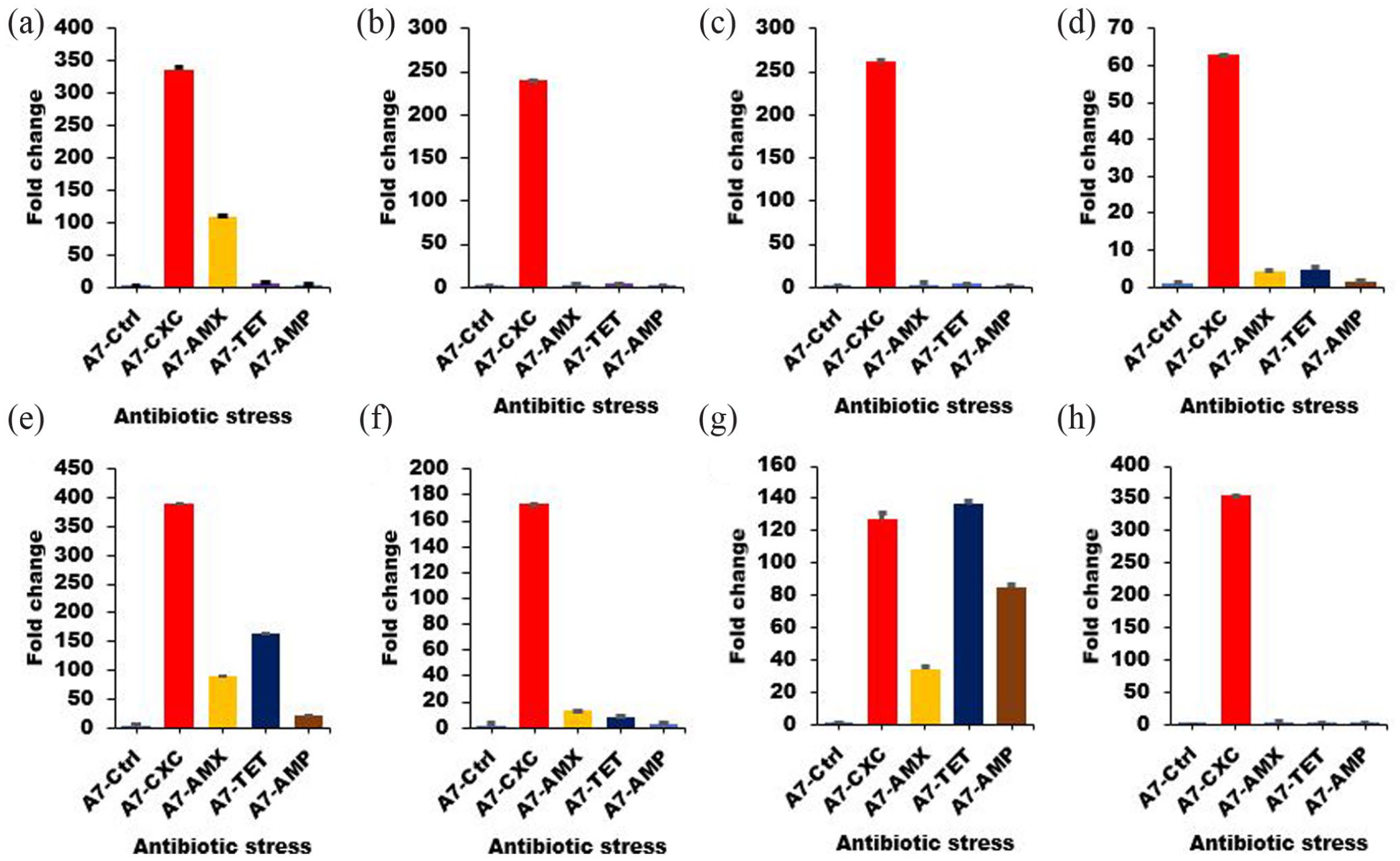
Expression profile of sporulation markers in B. cereus (A7) after exposure to antibiotics. The isolates were exposed to different antibiotics: cloxacillin (A7-CXC), amoxicillin (A7-AMX), tetracycline (A7-TET), and ampicillin (A7-AMP). The expression profile of eight genes was monitored following exposure to stress: (a) SigF, (b) SpoVK, (c) CotD, (d) Spo0J, (e) SoJ, (f) SpsJ, (g) CgeE, and (h) Spo0A. The samples were analyzed in triplicate using the delta-delta Ct method, and the experimental variations are shown as error bars.
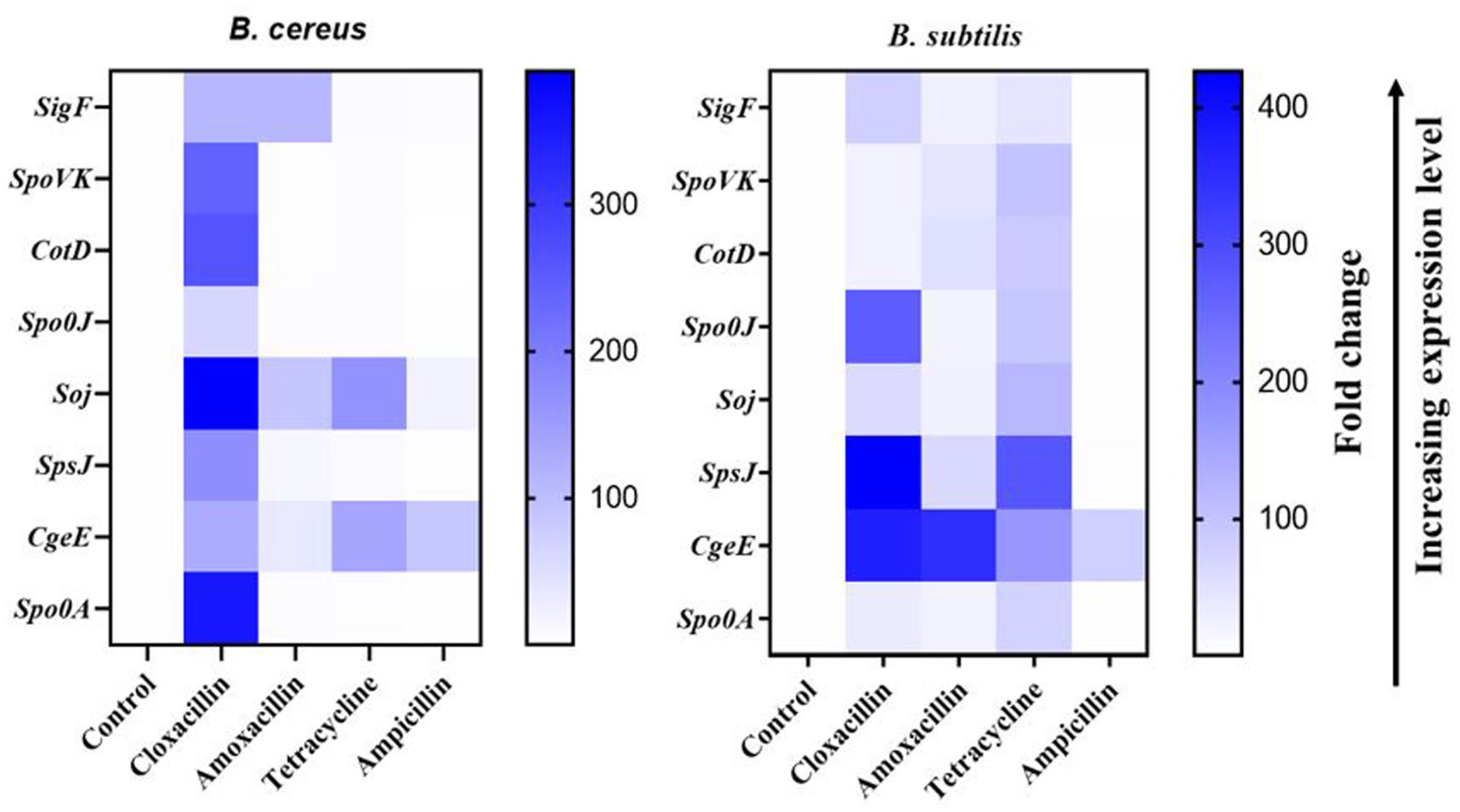
Comparative differential gene expression profile of sporulation markers between B. cereus and B. subtilis. Rows represent the sporulation markers and columns represent the fold change in the expression of each gene after exposure to the antibiotics (cloxacillin, amoxicillin, tetracycline, and ampicillin). The color intensity denotes the relative level of expression compared to their respective controls.
Discussion
Bacillus species are generally known for their ability to form endospores in stringent environmental and clinical conditions. 16 These stress factors may include naturally occurring antimicrobial substances (e.g. biocides – disinfectants, surfactants) or clinical antibiotics used to treat Bacillus-associated infections. 17 Beyond triggering the formation of endospores, these substances could select for bacteria subpopulations that are highly resistant to clinical antibiotics. 18 While spores are dispersed into the environments before they germinate and infect, evidences have highlighted their distribution in ICUs of hospitals as potential infectious agents.19,20 This study profiled the expression of sporulation markers in association with multidrug-resistant B. subtilis and B. cereus isolated from some Ghanaian hospital environments.
The Bacillus strains are multidrug resistant to 10 different antibiotics, including vancomycin, amoxicillin, cloxacillin, and penicillin, which are first-line treatment options for Bacillus-associated infections. This is similar to the studies conducted elsewhere in Africa and Asia, especially with spore-forming B. subtilis,5,21,22 confirming that spores contribute to bacterial resistance to antibiotics. A subsequent exposure of the strains to twofold (MIC ×2) and fourfold (MIC ×4) of the minimum inhibitory concentration showed that a subpopulation of the isolates were resistant at high antibiotic concentrations. Resistance to the selected antibiotic could be as a result of selection pressure or overuse of antibiotics which is a common practice in the ICU. This study suggests that the level of AMR displayed by environmental bacteria from hospitals is increasing, and this can pose a serious threat to public health.
Sporulation markers that have been commonly and uncommonly associated with spore formation in Bacillus species were identified. Nineteen of the twenty sporulation markers screened were detected in B. subtilis and fourteen in B. cereus. Some of these markers, for example, spoVK, spoVE, spoJ, and sigF are conserved in Clostridia and Bacillus species;23,24 while sigJ, soJ, yrbC, and yjcE are not commonly reported. In B. subtilis and B. cereus, spoVK and spoVE have been associated with morphogenesis25,26 encoding ATP-binding motif protein that makes endospores resist heat and antibiotics, 3 which might also account for the high level of resistance observed in this study. sigF, an alternative sigma factor, is essential for sporulation 18 and also regulate bacterial persistence and modulates AMR. 27 soJ and spo0J have been associated with Par-system (Par A/B) as possible proteins required for efficient plasmid partitioning from chromosome, 28 which is a molecular basis that drives spore formation and germination as well AMR.29,30 Although yrbC roles in sporulation is poorly understood, however, it has been associated with spore resistance in B. subtilis. 31
AMR and sporulation are stress response mechanisms that potentiate survival of environmental pathogens under harsh conditions. 32 Sporulation genes have been associated with AMR; however, the intersection of the mechanisms is unclear. In order to ascertain the regulatory roles of sporulation markers in bacterial resistance to antibiotics, expression profiles of eight genes (SigF, SpoVK, CotD, Spo0J, SoJ, SpsJ, CgeE, and Spo0A) present in all the strains were determined in B. subtilis and B. cereus (after exposure to cloxacillin, amoxicillin, tetracycline, and ampicillin antibiotics). The strain tested in this study was highly resistant to these antibiotics, even at high concentrations (MIC ×2/4). The data show increase in expression levels of all the genes in B. subtilis in the presence of cloxacillin, amoxicillin, and tetracycline. In B. cereus, the expression profile of all the genes increased in response to cloxacillin. Also, Spo0J, SoJ, SpsJ, and CgeE were moderately increased after exposure to amoxicillin, tetracycline, and ampicillin. This suggests that sporulation and its associated genetic factors could influence the resistance profiles observed in Bacillus spp. against tetracycline, cloxacillin, and other antibiotics. Differences in expression profiles and fold changes could be attributed to variation in resistance patterns to the tested antibiotics and also differences in the concentrations of antibiotics used for the stressed assay. Different bacteria have unique responses and survival mechanisms under stringent conditions, especially with fomite-resident strains as used in this study. Under these conditions, specific markers are triggered with functional relevance.
Overall, this study indicated that spore-forming Bacillus species isolated from hospital environments harboring sporulation markers might pose serious public health risks, as they are highly resistant to antibiotics. Also, the findings suggest a possible association between sporulation and AMR that can enhance the survival of Bacillus spp. in the environment or within host organisms. There is a need for appropriate antibiotic use and appropriate disinfection practices in the ICU to mitigate spread of spore-forming resistant Bacillus species.
Footnotes
Acknowledgements
The authors appreciate Mosi Research Lab and AMR Research Group led by Dr Abiola Isawumi at the Department of Biochemistry, Cell and Molecular Biology and West African Centre for Cell Biology of Infectious Pathogens (WACCBIP) at the University of Ghana for supports.
Authors’ Contributions
AI, EAA, and LM conceptualized and designed the study. AI, EAA, and MKA interpreted and analyzed the data. AI and EAA prepared the first draft of the manuscript. AI and MKA revised the draft for important intellectual content. LM made substantial contributions to the draft, critically reviewed the manuscript. All the authors approved the final draft of the manuscript for submission.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: MKA and EAA are supported by a WACCBIP-World Bank ACE PhD fellowship (ACE02-WACCBIP: Awandare) and a DELTAS Africa grant (DEL-15-007: Awandare). The authors have no other relevant affiliations or financial involvement with any organization or entity with a financial interest in or financial conflict with the subject matter or materials discussed in the manuscript apart from those disclosed.