
Abstract
Cytoskeleton-associated protein 4 (CKAP4) acts as a key transmembrane protein that connects the endoplasmic reticulum (ER) to microtubule dynamics. Researchers have not examined the roles of CKAP4 in nasopharyngeal carcinoma (NPC). The study aimed at evaluating the prognostic value and metastasis-regulating effect of CKAP4 in NPC. CKAP4 protein could be observed in 86.36% of 557 NPC specimens but not in normal nasopharyngeal epithelial tissue. According to immunoblot assays, NPC cell lines presented high CKAP4 expression relative to NP69 immortalized nasopharyngeal epithelial cells. Moreover, CKAP4 was highly expressed at the NPC tumor front and in matched liver, lung, and lymph node metastasis samples. Furthermore, high CKAP4 expression reported poor overall survival (OS) and presented a positive relevance to tumor (T) classification, recurrence, and metastasis. According to multivariate analysis, CKAP4 could independently and negatively predict patients’ prognosis. Stable knockdown of CKAP4 expression in NPC cells inhibited cell migration, invasion and metastasis in vitro and in vivo. Moreover, CKAP4 promoted epithelial-mesenchymal transition (EMT) in NPC cells. CKAP4 knockdown was followed by the downregulation of the interstitial marker vimentin, and upregulation of the epithelial marker E-cadherin. In NPC tissues, high CKAP4 expression exhibited a positive relevance to vimentin expression and a negative relevance to E-cadherin expression. In conclusion, CKAP4 is an independent predictor of NPC, and CKAP4 might contribute NPC progression and metastasis, which may be involved in EMT with vimentin and E-cadherin.
Impact statement
Distant organ metastasis is the major factor resulting in treatment failure in nasopharyngeal carcinoma (NPC) patients. However, the molecular mechanism underlying metastasis is still not clearly understood. In this study, immunohistochemistry assisted in detecting the high expression of cytoskeleton-associated protein 4 (CKAP4) protein in a large sample of NPC tissues. According to western assays, NPC cell lines exhibited high CKAP4 expression. Moreover, a high level of CKAP4 related to NPC patients’ poor survival and exhibited a positive relevance to metastasis. As revealed by the in vivo and in vitro experiments, stable knockdown of CKAP4 expression suppressed cell migration, invasion, and metastasis and was correlated with vimentin and E-cadherin levels. Accordingly, CKAP4 may promote tumor development in progression and metastasis in NPC patients by influencing epithelial mesenchymal transformation of NPC, which indicated that CKAP4 could serve as a predictor or promising new therapeutic targeted for metastatic NPC.
Introduction
Nasopharyngeal carcinoma (NPC) refers to an epithelial carcinoma originated from the nasopharyngeal mucosal lining and patients with NPC are geographically concentrated in South China, Southeast Asia and North Africa. 1 Radiotherapy (RT) is the backbone of NPC treatment. Due to the widespread application of RT modulated by intensity, induction, and concurrent chemotherapy and the advancement of an accurate cancer staging system, the management of NPC has been greatly improved in recent decades. 2 The reported 5-year local control rates are outstanding, ranging from 70% to 90%. However, approximately about 20–30% of patients eventually develop distant metastasis or local recurrence resulting in treatment failure after the administration of radical chemoradiotherapy for locoregionally advanced NPC (stages III–IVB). 3 Local recurrence and distant metastasis are the main factors that lead to poor overall survival (OS) of NPC patients with advanced-stage disease. 4 Furthermore, the molecular mechanism of metastasis is still not clearly understood. Therefore, studying the molecular mechanism of NPC metastasis and developing new molecular targets are important for the optimization of treatment strategies.
Cytoskeleton-associated protein 4 (CKAP4), a 63-kDa palmitoylated type II transmembrane protein, was initially identified to anchor the endoplasmic reticulum (ER) by binding to microtubules in epithelial cells. 5 It remarkably impacts the formation and maintenance of microtubule network structure and the dynamic balance of the ER. It is also a plasma cell markers. However, in addition to being expressed in normal plasma cells and plasmacytoma cells, CKAP4 can also be observed in several cell types’ cell surface membrane, such as type II pneumocytes and vascular smooth muscle cells (VSMCs). 6 CKAP4 has been identified as a receptor for tissue plasminogen activator (TPA), 7 surfactant protein A (SP-A), 8 antiproliferative factor (APF), 9 alginate exopolysaccharides, 10 and Dickkopf (DKK) proteins. 11 Due to the binding to various ligands, CKAP4 protein exerts various effects. Moreover, CKAP4 palmitoylation and phosphorylation can critically help to regulate its translocation between the ER and plasma membrane or nucleus where it can exert different biological effects through its translocation between organelles. 12 Further research has demonstrated that phosphorylated CKAP4 is capable of translocating to the nucleus in Hela cells, and binds DNA through its outer domain, and subsequently alters the transcription of genes involved in regulating proliferation and tumogenesis.12,13 CKAP4 has been reported to be a molecule that promotes multiple cancers to progress, such as pancreatic cancer (PC), renal cancer, lung cancer, and esophageal cancer.14 –17 However, contrary studies have reported that CKAP4 can resist tumor development in different tumor types, such as hepatocellular carcinoma (HCC), intrahepatic cholangiocellular carcinoma (ICC), and glioma.18 –20 It is still necessary to determine whether CKAP4 can promote or resist cancer progression. Furthermore, up to now, researches fail to examine CKAP4 expression as well as its role in NPC.
In the study, immunohistochemistry (IHC) and immunoblot (Western blot) methods served for examining CKAP4 expression in primary and metastatic NPC tissues, thereby exploring the effect of CKAP4 on NPC. Second, analysis was conducted on the association of CKAP4 expression with clinical factors and NPC patients’ outcomes. At last, in vivo and in vitro experiments were carried out for evaluating how CKAP4 affected NPC cell metastasis and invasion.
Materials and methods
Patients and clinical tissue samples
The study included 557 NPC specimens from the Department of Pathology, Sun Yat-Sen University Cancer Center (SYSUCC), Guangzhou, China, from January 2001 to December 2003, adopting the same inclusion criteria as the previous literatuer. 21 All the patients underwent normal curative RT, accompanied or not accompanied by chemotherapy. A total of 23 lung, liver, or lymph node (LN) metastatic tissues from NPC patients and samples from the corresponding primary tumors were obtained from above department. The study had obtained the approval of the Institute Research Medical Ethics Committee of Sun Yat-Sen University.
IHC and evaluation
CKAP4 received IHC staining as previous description and vimentin and E-cadherin, paraffin-embedded tissues were cut into 5 mm sections. 21 In brief, the sections underwent one night of incubation in a rabbit anti-human CKAP4 polyclonal antibody (Cat. No.: HPA000792, Sigma-Aldrich; working dilution 1:500), mouse anti-human E-cadherin monoclonal antibody (Abcam, working dilution 1:500), or rabbit anti-human vimentin monoclonal antibody (Cat. No.: ab92547, Abcam; working dilution 1:500) at 4°C. An EnVision kit (DAKO) assisted in the detection of the primary antibodies, followed by the visualization of 3,3-diaminobenzidine substrate and hematoxylin counterstaining. The IHC results were evaluated and scored as described previously. 19 The pathologists’ evaluation results exhibited a concordance rate of 87.3%, demonstrating the high reproducibility of such scoring method.
Cell line and viral transduction
Cells of four NPC cell lines, namely, CNE2, SUNE1, 5-8F, and 6-10B, underwent culturing in RPMI 1640 with 10% fetal bovine serum (FBS). Cells of two additional NPC cell lines (S18 and S26) received culturing in DMEM (Dulbecco’s Modified Eagle Medium) added with 10% FBS. Cells of NP69 received culturing in keratinocyte serum-free medium with fetal calf serum (FCS) (5%), bovine pituitary extract (25 μg/mL), and recombinant epidermal growth factor (0.2 ng/mL), as per the instruction of the manufacturer. All cell lines underwent incubation in humidified atmosphere at 37°C with 5% CO2.
A BLOCK-iT Lentiviral Pol II miRNAi system served for establishing cell lines with stably expressed CKAP4 short hairpin RNA (shRNA) or a scrambled non-target shRNA, as per the instruction of manufacturer. The targets of human CKAP4 shRNAs were 5′-GGATAGGTTGTTTGTGAAA-3′ and 5′-GGAGCCAGAAGGAGATCAA- 3′.
Immunoblot analysis
RIPA buffer was adopted for harvesting and lysing cells. SDS-PAGE (Sodium Dodecyl Sulphate-Polyacrylamide Gel Electrophoresis) assisted in separating identical amounts of the denatured protein samples, which then were transferred to polyvinylidene difluoride (PVDF) membranes to receive immunoblot analysis. The immunoblot analysis adopted antibodies against CKAP4, E-cadherin, vimentin, N-cadherin, fibronectin, and β-catenin antibody, with anti-GAPDH mouse monoclonal antibody as a loading control. An improved chemiluminescent Western blotting kit served for the visualization of all protein bands.
ImageJ software (version 1.44) served for normalizing the gray value possessed by target bands under the detection of corresponding antibodies to those under the detection of GAPDH (loading control), thereby quantifying the protein level. We repeated each experiment no less than three times.
Wound-healing, Transwell migration, and invasion assays
Wound healing assays together with Transwell assays served for the evaluation of cell migration and invasion. For the Transwell assays, after 24 h, cells on the inner side of the membrane were cleared, and cells on the outer side received crystal violet dyeing and were counted microscopically. For the wound healing assay, cells were photographed at 0, 24, and 36 h after wounding, and the migrated areas were measured. We repeated all experiments three times.
Animals and spontaneous LN and lung metastasis assay
Five- to six-week-old male athymic BALB/c nu/nu mice were provided by Shanghai Institutes for Biological Sciences. All the animal studies conformed to the principles and procedures in the guidelines of the Institutional Animal Care and Use Committee of SYSUCC. Spontaneous LN metastasis analysis was carried out based on previous description. 22 In brief, each mouse was injected with 1 × 105 cells into the left hind footpad for generating a primary tumor. Seven to eight weeks later, we collected the popliteal LN of the left hind foot before experiment terminated for the regular tissue processing. In the lung metastasis assay, the lateral vein of athymic BALB/c nude mice was injected with 1 × 106 CKAP4 stable knockdown or scrambled cells in 100 μL phosphate buffered saline (PBS), and 7–8 weeks later, the mice received euthanization. The split lungs were photographed and weighed, followed by one night of fixation in 4% paraformaldehyde (PFA), cryoprotection in 30% sucrose in PBS, and freezing treatment in embedding media under optimal cutting temperature (OCT). Serial sections of the LN and lungs received H&E staining, and a light microscope served for counting the number of pulmonary colonies.
Statistical analysis
SPSS (version 25.0) served for statistical analysis. Monthly calculation of the survival time was conducted, and the calculation of OS started from the initial diagnosis date to the death date or the last follow-up. The χ2 test or Fisher’s exact probability method assisted in the comparisons between groups. The Kaplan–Meier and log-rank methods served for the analysis of the survival rate. The Cox regression model served for multivariate analysis. P < 0.05 reported statistical significance.
Results
Patient characteristics
Male patients occupied 74.5% (415) of the total number of patients (557), with an average age of 46 years (in the range of 19–78 years). Thereinto, 139 patients (25.0%) developed non-keratinizing differentiated carcinoma (NKDC), 402 patients (72.2%) developed non-keratinizing undifferentiated carcinoma (NKUC), and the rest 2.8% developed keratinizing squamous cell carcinoma (KSCC). The number of patients in stage I and stage II, and stage III and stage IV took up 26.4% (147) and 73.6% (410) of the total number of patients, respectively. The follow-up lasted for 58.63 months on average (in the range of 2–99 months). During the 5-year follow-up period, of all the patients, 184 (33.0%) died, and disease progressed in 208 (37.3%) patients. Supplemental Table 1 lists patients’ clinical information in detail.
CKAP4 is expressed at the invasive front of NPC tissues and is a poor prognostic indicator
For investigating the CKAP4 expression status in NPC biopsies, CKAP4 in a set of tissue microarrays that involved 557 NPC samples underwent IHC staining (Table 1; Figure 1(a)). IHC staining revealed that 484 of 557 (86.36%) paraffin-embedded samples showed positive staining of CKAP4 in a majority of tumor cells (Figure 1(a), red arrow); adjacent non-cancerous epithelial cells presented no positive staining (Figure 1(a), black arrow). As shown in Figure 1(b), CKAP4 mainly presented expression in the tumor cell cytolymph and were preferentially distributed at the tumor invasive front (Figure 1(b), red arrow).
Associations between CKAP4 expression and clinicopathological characteristics in NPC patients.
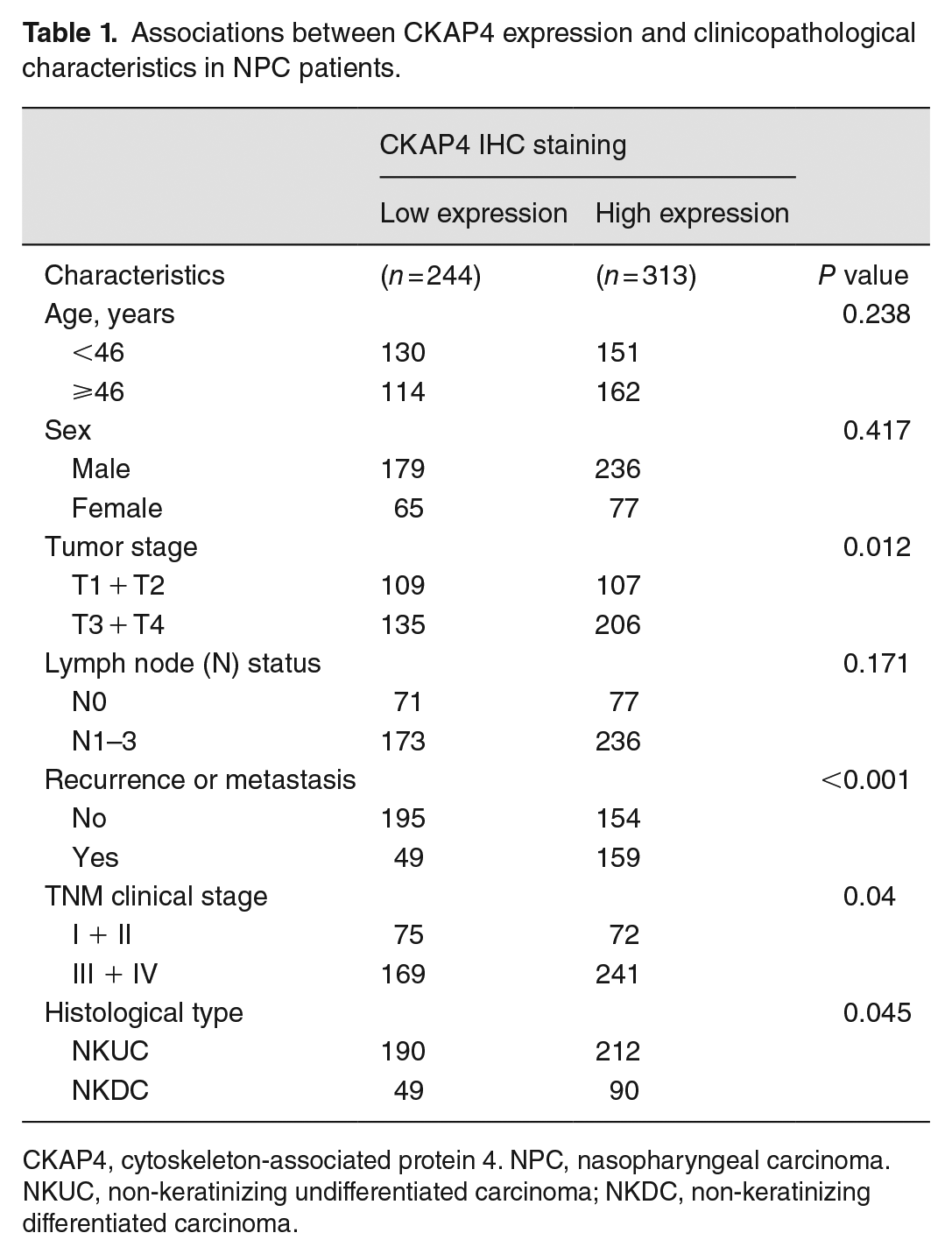
CKAP4, cytoskeleton-associated protein 4. NPC, nasopharyngeal carcinoma. NKUC, non-keratinizing undifferentiated carcinoma; NKDC, non-keratinizing differentiated carcinoma.
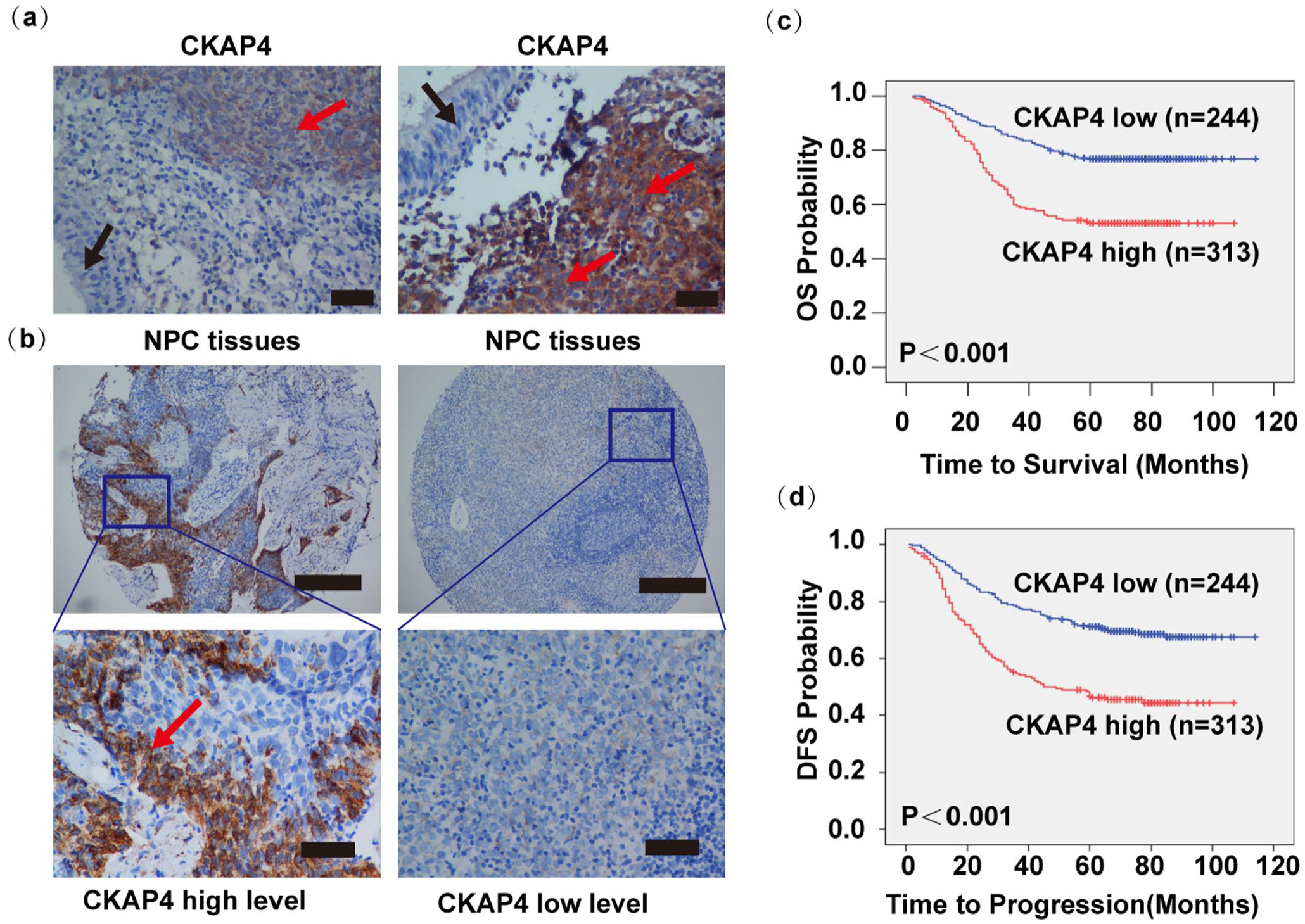
Elevated CKAP4 levels reported poor OS and DFS of NPC patients. (a) Immunohistochemical analysis demonstrated certain staining of CKAP4 in NPC tumor nests (red arrows) instead of in adjacent normal NPE cells (black arrow, scale bar is 100 μm). (b) High and low CKAP4 protein expression levels in the TMA are displayed with low and high magnifications under a light microscope (the top scale bar is 500 μm, and the bottom is 100 μm). (c, d) The low CKAP4 group presented remarkably higher OS rate and DFS rate.
The expressions and the cutoff value were taken into account for separating NPC samples into group with low CKAP4 expression and group with high CKAP4 expression. In all, 313 of 557 (56.2%) NPC tissues presented high CKAP4 expression. χ2 test analysis confirmed the positive relevance of CKAP4 expression to the tumor T classification (Pearson’s R = 0.107; P = 0.012), recurrence or metastasis (Pearson’s R = 0.314; P < 0.001), clinical stage (Pearson’s R = 0.087; P = 0.04), and histological type (Pearson’s R = 0.0085; P = 0.045) (Table 1); comparatively, CKAP4 expression was not obviously relevant to other clinicopathologic features (patient sex, age, and N classification) (P > 0.05, Table 1).
The cohort of 557 NPC patients presented a 5-year OS rate of 67.0% and disease-free survival (DFS) rate of 61.0%. According to Kaplan–Meier and log-rank test analyses, high CKAP4 expression reported poor OS (5-year survival rates, 55% vs 86.4%, P < 0.001, Figure 1(c)) and DFS (5-year survival rates, 49.2% vs 79.8%, P < 0.001, Figure 1(d)). According to multivariate analyses, a high CKAP4 expression could independently and negatively indicate patients’ OS and DFS or the distant metastasis-free survival (Table 2). Furthermore, the CKAP4 expression was positively associated with recurrence and metastasis (Table 2). Accordingly, a high CKAP4 expression in NPC presented an obvious relevance to unfavorable outcomes of patient, the localization at the invasive front indicated that it could promote tumor invasion, and CKAP4 protein levels might show a positive relevance to NPC metastasis.
Univariate and multivariate Cox proportional hazards regression analysis for CKAP4.
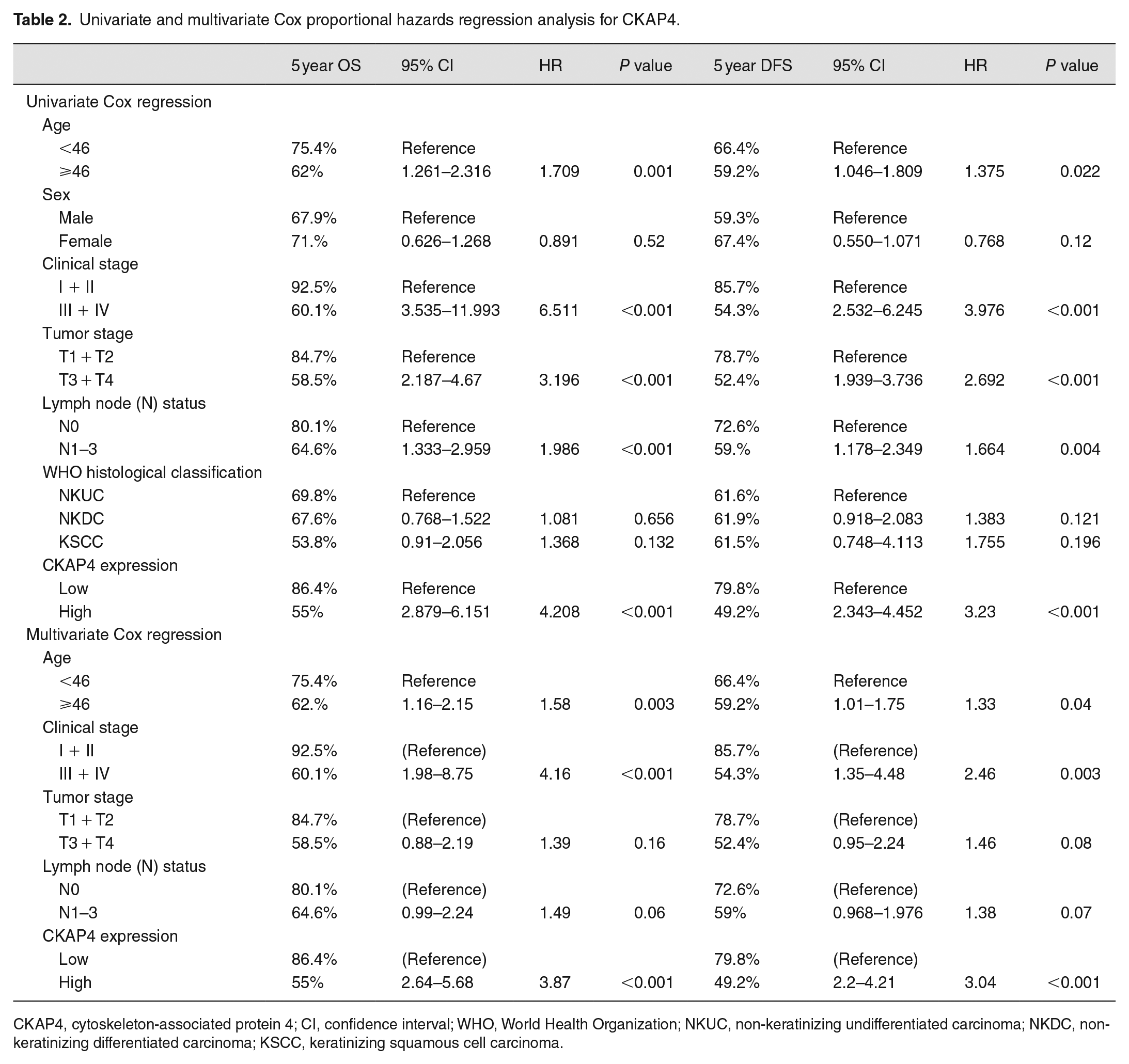
CKAP4, cytoskeleton-associated protein 4; CI, confidence interval; WHO, World Health Organization; NKUC, non-keratinizing undifferentiated carcinoma; NKDC, non-keratinizing differentiated carcinoma; KSCC, keratinizing squamous cell carcinoma.
CKAP4 is upregulated in liver, lung, and LN metastatic tissues from NPC patients and in highly migratory/invasive NPC cells
For evaluating the clinical relevance of the hypothesis in the study that CKAP4 was capable of regulating NPC metastasis, archival tissues of NPC patients’ liver, lung, and LN metastatic tissues were collected to be compared with primary NPC tissues (n = 23). IHC staining determined the obviously higher protein levels of CKAP4 in the liver, lung, and LN metastatic tissues relative to the primary tissues (Figure 2(a) and (b)). Relative to NP69 cells, NPC cell lines including 6-10B, 5-8 F, SUNE1, S26, S18, and CNE2 NPC cells presented remarkably increased CKAP4, according to the Western blot analysis (P < 0.001, Figure 2(c) and (d)). CKAP4 was also highly expressed in the highly metastatic S18 (P < 0.05) and 5-8F cells (P < 0.01), and lowly expressed in the metastatic potential S26 and 6-10B cells.23,24
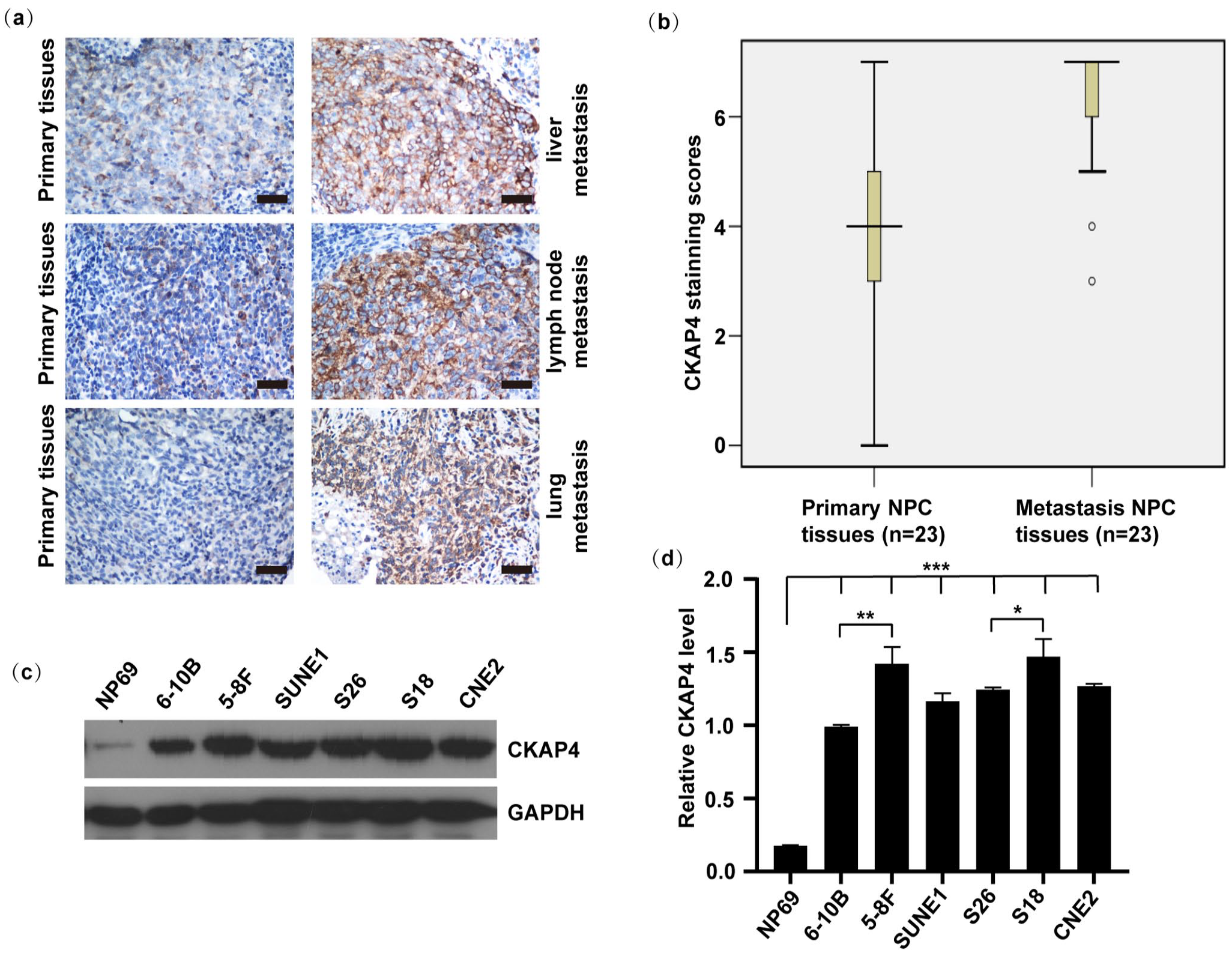
CKAP4 expression presented upregulation in liver, lung, and LN metastases and in highly metastatic NPC cell lines. (a) IHC staining served for evaluating the CKAP4 protein expression in primary NPC tissues and compare liver, lung, and LN metastases from NPC (scale bar is 100 μm). Metastatic lesions show obvious staining. (b) Statistical analysis (Mann–Whitney test) discovered remarkably increased CKAP4 expression in NPC metastatic tissues relative to primary NPC tissues (P < 0.001). (c and d) The Western blot examined the CKAP4 protein level. (c) ImageJ software served for normalizing the gray value possessed by target bands under the detection of corresponding antibodies to those under the detection of GAPDH (loading control), thereby quantifying the protein level. (d) We repeated each experiment at least three times. Data are in the form of the mean ± SEM. *, P < 0.05; **, P < 0.01; ***, P < 0.001. One-way analysis of variance.
CKAP4 knockdown suppresses NPC cell migration and invasion
For examining the causal effect of CKAP4 on NPC cell motility, we engineered S18 and 5-8 F cell lines stably expressing either shRNAs targeting CKAP4 (CKAP4 KD1 and CKAP4 KD2) or a scrambled non-target shRNA. Immunoblotting confirmed that CKAP4 expression was inhibited at the protein level (Figure 3(a)). The loss of CKAP4 led to an obvious suppression of S18 cell invasion and migration (Figure 3(b) to (d)). Similar results were observed in 5-8 F cells after we knocked down the expression of CKAP4. These results indicated that CKAP4 could facilitate NPC cell migration and invasion, in cells which were examined in culture.
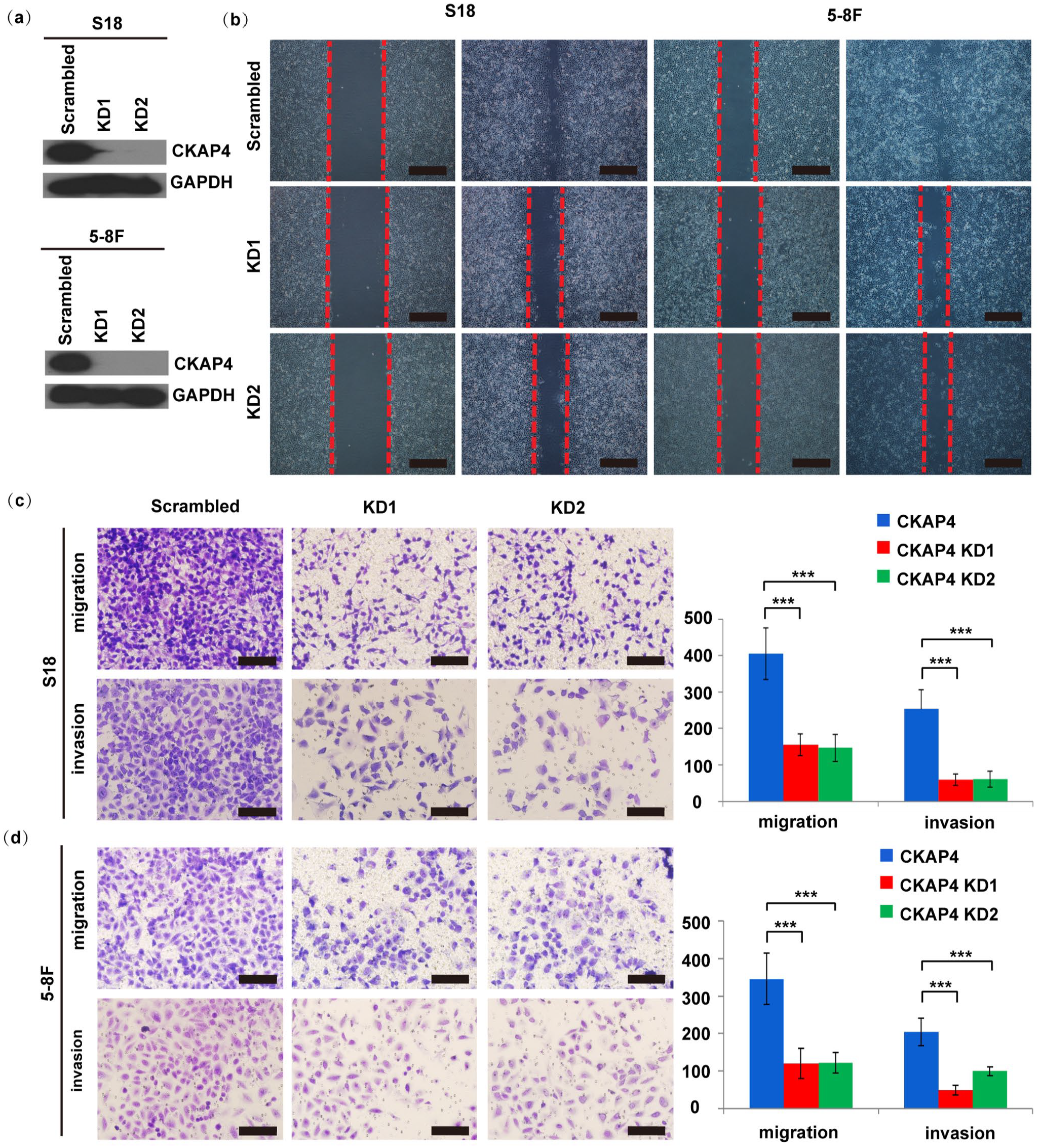
Suppression of CKAP4 inhibits S18 and 5-8 F cell migration and invasion. (a) Immunoblotting determined the CKAP4 protein levels. CKAP4 suppression in KD1 and KD2 cells led to a complete elimination of CKAP4 protein expression. (b) Suppression of CKAP4 resulted in remarkably decreased migration ability of S18 and 5-8F cells, as determined by a wound healing assay (scale bar is 100 μm). The red dashed lines represent the wound margins. (c, d) A two-chamber assay served for further evaluating S18 and 5-8F cell migration and invasion (scale bar is 100 μm). Columns, mean; bars, SE (from triplicates). Student’s t-test served for the statistical analyses. ***, P < 0.001 versus the scrambled shRNA group; ***, P < 0.001 for the KD1 and KD2 groups versus the scrambled shRNA group.
CKAP4 knockdown suppresses the NPC metastasis rate in vivo
LN and lung metastasis models assisted in determining the impact of CKAP4 in vivo. 20 CKAP4 knockdown cells presented an obviously lower metastasis rate (Figure 4(a) and (b)). Furthermore, CKAP4 silencing prevented lung metastasis in mice in a lateral vein lung metastasis tumor model (Figure 4(c) and (d)). Accordingly, CKAP4 could impact NPC metastasis in LN and lung metastasis models.
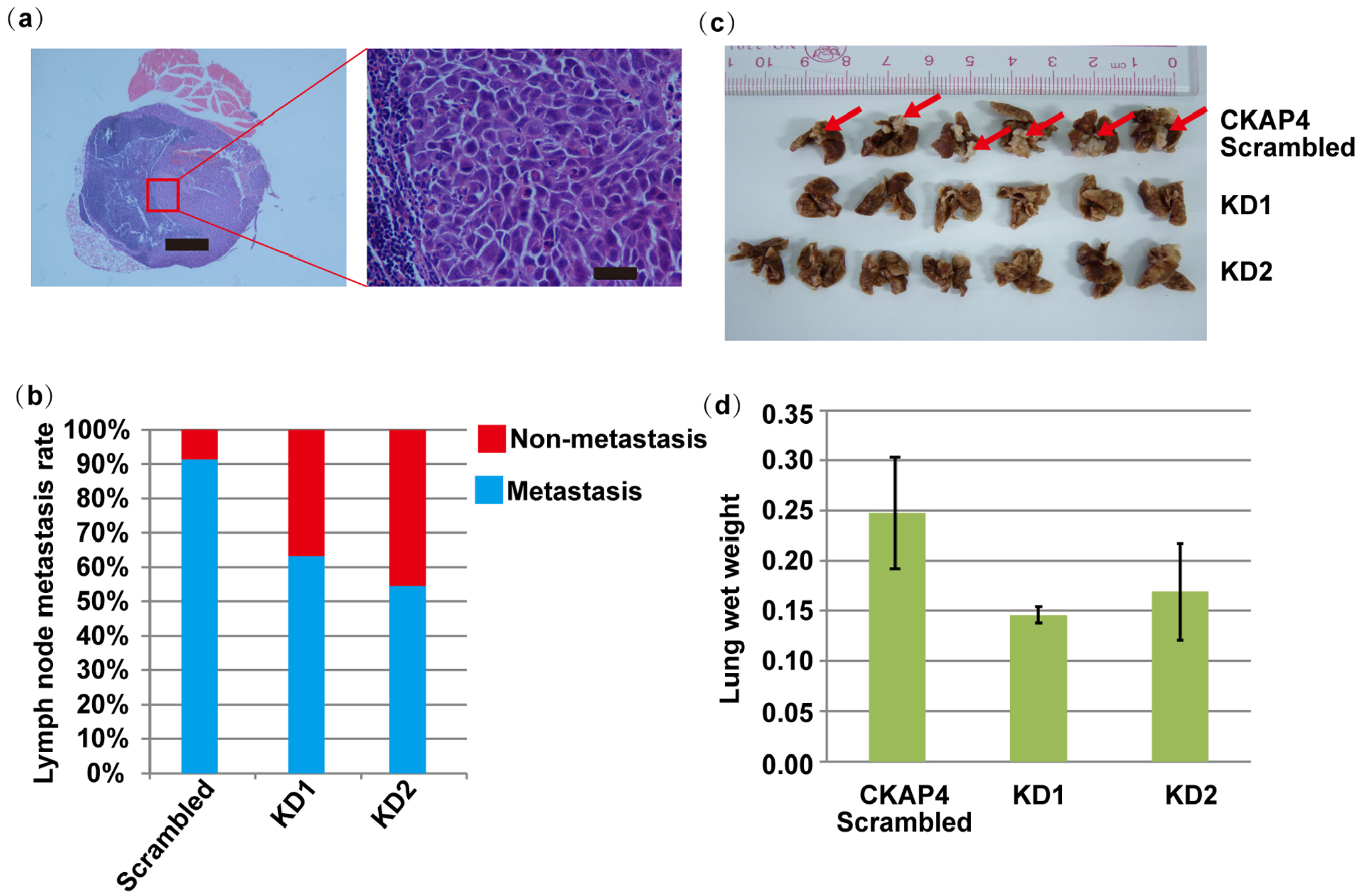
In vivo LN and lung metastasis rates of CKAP4 knockdown cells. (a) Image of hematoxylin-stained sections of popliteal LNs (left scale bar is 1000 μm, right scale bar is 100 μm). (b) The proportion of popliteal LN metastasis following the inoculation. CKAP4 knockdown in S18 cells considerably lowered the metastasis rate from 90.9% (10 of 11) to 63.6% (7 of 11) in the KD1 group and 54.5% (6 of 11) in the KD2 group. (c) Images of lungs harvested from scrambled and CKAP4 shRNA mice. The red arrows indicate metastatic foci. (d) The mean lung wet weight in scrambled shRNA (n = 6) and CKAP4 KD1 and KD2 shRNA group mice (n = 6 and 7, respectively). Bars represent the means and upper 95% confidence intervals; P < 0.001, Student’s t-test.
CKAP4 mediates NPC cell metastasis and is related to EMT
For more deeply exploring how CKAP4 promoted NPC metastasis, we evaluated the protein levels of many epithelial-mesenchymal transition (EMT) markers in S18 and S26 cells. The mesenchymal markers vimentin, fibronectin and β-catenin were more significantly expressed in the S18 cells relative to the S26 cells, while epithelial protein E-cadherin and N-cadherin presented lower expression in the S18 cells (P < 0.001, Figure 5(b) and (c)). High CKAP4 expression was accompanied by increased vimentin level and decreased E-cadherin level in the highly metastatic cells (Figure 5(a)). CKAP4 suppression led to lower vimentin levels and higher E-cadherin levels (Figure 5(a)). To confirm this finding, we examined the relationship between CKAP4 and EMT and carried out IHC staining (in 102 primary human NPC tissue samples) analysis of CKAP4, vimentin and E-cadherin. Accordingly, the CKAP4 expression level exhibited a positive relation to vimentin expression, and an inverse relation to E-cadherin expression (Figure 5(a), (d), (e), and (f)). These data suggest that CKAP4 modulates vimentin level, thereby impacting the EMT.
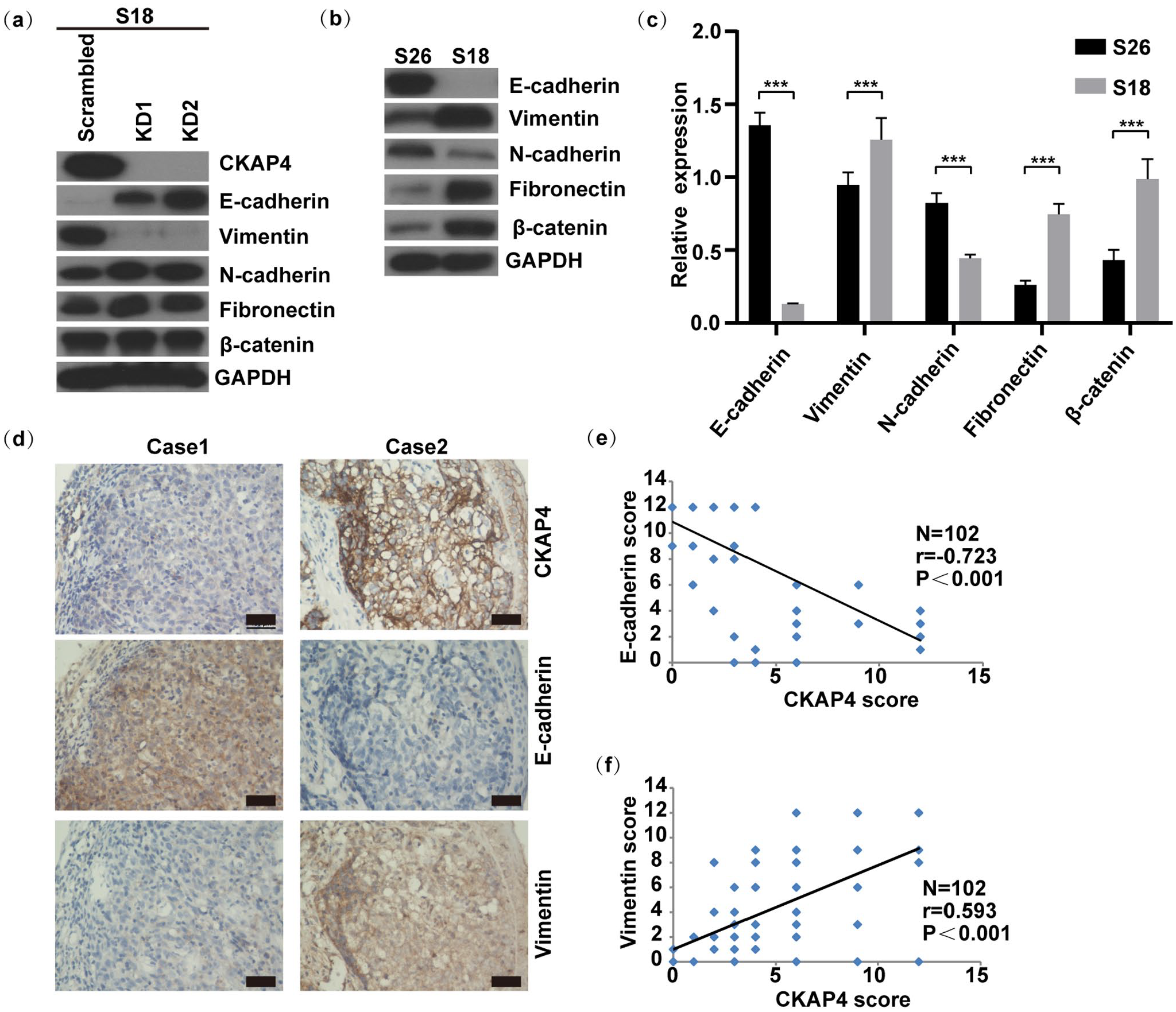
CKAP4 expression modulates mesenchymal marker expression in NPC cells. (a) Impact of stable CKAP4 knockdown on the EMT-associated markers. (b and c) Western blot served for examining the expression of EMT-associated proteins in S18 and S26 cells (b) ImageJ software served for normalizing the gray value possessed by target bands under the detection of corresponding antibodies to those under the detection of GAPDH (loading control), thereby quantifying the expression of these proteins (c). We repeated each experiment no less than three times. Data are in the form of the mean ± SEM. ***, P < 0.001. One-way analysis of variance. (d) Continuous sections of human NPC tissue received IHC staining using antibodies against CKAP4, vimentin and E-cadherin (scale bar is 100 μm). Low CKAP4 expression in the tumor tissue in case 1 resulted in lost vimentin and increased E-cadherin. Comparatively, high CKAP4 expression in case 2 resulted in increased vimentin levels and lost E-cadherin. (e) Same methods were adopted for scoring and plotting 102 primary NPC tissues that received the staining of antibodies against E-cadherin and CKAP4. A significant negative relevance of E-cadherin to CKAP4 levels was illustrated. (f) For vimentin staining, 102 NPC tissues underwent qualification for analysis. A markedly positive relevance of vimentin to CKAP4 levels was illustrated.
Discussion
Distant organ metastasis is currently the primary reason for treatment failure in NPC patients.2,25 Effective targeted therapy can improve the clinical treatment efficacy in patients with locally advanced and advanced NPC. 26 Hence, it is required to identify effective molecular targets for enhancing the NPC treatment. The present study conducted immunohistochemical staining, finding the high CKAP4 expression in NPC tumor tissues, instead of in normal NPE tissues. Moreover, we found that CKAP4 was specifically highly expressed at the invasion front of NPC tissues. It presented an obviously higher expression in liver, lung, and LN metastatic tissues than in matched primary NPC tissues. High expression of CKAP4 could increase the risk of metastasis, recurrence, and death for NPC patients. Hence, CKAP4 expression showed a close relevance to NPC metastasis. Cell function experiments showed that interference with CKAP4 expression could significantly reduce NPC cell migration and invasion ability in vitro. Furthermore, similar results were obtained in animal experiments: in plantar-popliteal LN and tail vein-lung metastasis mouse models, the ability of NPC cells metastasis to the LNs and lungs was significantly reduced after stable CKAP4 knockdown. These results strongly indicate that CKAP4 promotes NPC metastasis.
CKAP4 can significantly help to stabilize the ER structure, and is associated with tumorigenesis. 27 Many studies have found that CKAP4 can impact the development of a variety of tumors as a cancer-promoting factor.14,27 Kimura et al. 14 found that CKAP4 highly expressed on the plasma membrane can promote the cell proliferation regarding PC and lung cancers by activating the phosphoinositol 3-kinase-Akt pathway. Shinno et al. 17 reported that CKAP4 was upregulated in approximately 40% of patients who had esophageal squamous cell carcinoma (ESCC) and promoted ESCC cell proliferation by binding to the receptor DKK1 and activating the AKT pathway. CKAP4 overexpression promotes the proliferation, invasion, and migration of renal clear cell carcinoma through the cyclin B signaling pathway during the G2/M phase of the cell cycle. 15 Another study showed that CKAP4 are capable of mediating PC cell invasion and migration by binding endogenous β1 integrin in the cell surface and cytoplasm and recycling of α5β1 integrin. 6 On the other hand, CKAP4 plays an anticancer role in various tumors such as HCC, ICC, and glioma.18 –20 For example, CKAP4 suppresses the EGFR signaling, thereby restricting HCC growth and metastasis. 28 Therefore, CKAP4 exerts various impacts on tumor biology, which may be associated with various signaling pathways due to the binding of massive ligands in different microenvironments.
Epithelial cells are capable of obtaining mesenchymal features during the process of EMT. In cancer, during EMT, E-cadherin level is declined and vimentin level is increased, thus promoting metastasis and invasion of tumor cells and making cells more easily metastasized to target tissues or organs, which are related to tumor initiation, invasion, metastasis, and treatment resistance. 29 In NPC cell lines, knocking down endogenous CKAP4 significantly reduced vimentin expression and increased E-cadherin expression. The IHC results of 102 cases of NPC tissues also showed the positive relevance of CKAP4 expression to vimentin expression and the obviously negative relevance of CKAP4 expression to E-cadherin expression. Hence, interfering with CKAP4 can make NPC cell lose interstitial properties but acquire epithelial properties. The correlation between CKAP4 and the expression of vimentin and E-cadherin also suggests that CKAP4 may facilitate NPC cells in terms of the migration, invasion, and metastasis by inducing EMT. Additional studies on the specific mechanism by which CKAP4 promotes NPC metastasis need to be carried out.
In summary, this study revealed that the cytoskeleton protein CKAP4 is specifically expressed in NPC tumor tissues, especially in metastatic tissues and NPC cells, but is not expressed in normal NPE tissues. Furthermore, high CKAP4 expression indicates a higher risk of metastasis, recurrence, and death in NPC patients. After stable knockdown of CKAP4 expression, the invasion and migration ability of NPC cells and tumor LN and lung metastasis were significantly decreased in nude mice. Moreover, after CKAP4 knockdown, the EMT marker E-cadherin protein level presented an obvious upregulation, while the level of vimentin presented an obvious downregulation, which demonstrated that CKAP4 might promote NPC cell metastasis through inducing EMT.
Supplemental Material
sj-pdf-1-ebm-10.1177_15353702231167940 – Supplemental material for Expression of cytoskeleton-associated protein 4 is associated with poor prognosis and metastasis in nasopharyngeal carcinoma
Supplemental material, sj-pdf-1-ebm-10.1177_15353702231167940 for Expression of cytoskeleton-associated protein 4 is associated with poor prognosis and metastasis in nasopharyngeal carcinoma by Manbo Cai, Weijun Wu, Shengling Deng, Qiao Yang, Haibiao Wu, Haiyun Wang, Jiaxing Zhang, Qisheng Feng, Jianyong Shao, Yixin Zeng and Jianjun Li in Experimental Biology and Medicine
Footnotes
Acknowledgements
We thank Prof. Chao-Nan Qian from Sun Yat-Sen University Cancer Center for providing the N-cadherein, β-catenin, and fibronectin antibodies.
Authors’ Contributions
MB conducted all the experiments, wrote the initial draft, and revised the manuscript. WJ, SL, and QY helped to perform some of the molecular biology experiments. HB, HY, JX, and QS performed animal and IHC work. JY and YX participated in the study design and interpretation of the results. JJ contributed to the study coordination and revisions. All authors read and approved the final manuscript.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported partially by the Natural Science Foundation of Hunan Province, China (2020JJ4539 and 2021JJ30610); and the Key Guiding Project of Hunan Provincial Health Commission (202104101985); the Science and Technology Innovation Project of Hengyang City (202150064015); the University of South China Teaching Reform Research Project (2020YB-XJG127); and the National High Technology Research and Development Program of China (863 Program) (2012AA02A206).
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.