
Abstract
Chinese native medicine Scutellaria baicalensis Georgi, also referred to as Chinese skullcap or Huang-Qin, is frequently used to treat cancer, viral infections, and seizures. This plant’s abundance of flavones (wogonoside) and their related aglycones (wogonin) is responsible for many of its pharmacologic effects. A significant ingredient in S. baicalensis that has been the subject of the most research is wogonin. Numerous preclinical investigations revealed that wogonin suppresses tumor growth by cell cycle arrest, stimulating cell death and preventing metastasis. This review focuses on a complete overview of published reports that suggest chemopreventive action of wogonin and the mechanistic insights behind these neoplastic activities. It also emphasizes the synergistic improvements made by wogonin in chemoprevention. The factual data in this mini-review stimulate additional research on chemistry and toxicological profile of wogonin to confirm its safety issues. This review will encourage researchers to generalize the merits of wogonin to be used as potential compound for cancer treatment.
Impact Statement
Numerous research experimental results have shown that wogonin’s anticancer effects are linked to its ability to induce apoptosis and stop the cell cycle. In addition, it prevents angiogenesis and the spread of malignant cells. Wogonin is a prospective candidate to be used in synergism with currently available anticancer medications since it has the ability to reverse cancer cells’ drug resistance. Future scientists and researchers can learn everything about wogonin’s cancer medication targets and how it affects much significant cancer hallmarks.
Introduction
It is estimated that by 2030, there will be 26 million new cancer cases and 17 million cancer deaths.1,2 Investigations suggested that dietary phytochemicals can help to prevent cancer and even have been found to improve oncopatients’ quality of life.3–5 The molecular basis of these bioactive compounds’ actions may provide new opportunities for the scientific community to create or enhance innovative treatment strategies to treat horrible illnesses, such as cancer, cardiovascular disease, and neurodegenerative disorders. A bioactive polyhydroxy flavone called wogonin (5,7-dihydroxy-8-methoxyflavone) was discovered in Scutellaria baicalensis Georgi. 6 In addition, it has been shown that the synthetic wogonin derivatives possess a variety of pharmacological activities, including antiviral, anti-inflammatory, antioxidant, neuroprotective, and antidiabetic associated with illness prevention.7,8 Wogonin has been investigated for its anticancer properties and targeted mechanistic insight concerning different cancers, such as breast, leukemia, and colorectal cancers (CRCs).9–14
The interaction of the signaling pathways for survival and apoptosis regulates tumor pathogenesis. It has been discovered that wogonin causes tumor cells to undergo apoptosis, which can be thought of as a technique to fight cancer.15,16 Similarly, different flavonoids activate cell cycle arrest and enhance tumor cell death in various cancerous cell lines to display powerful anticancer action.17,18 This review article focuses on the various anticancer activities of wogonin in various experimental model systems to better understand and draw attention to this ancient molecule. It describes the apoptosis function, cell cycle arrest, antiangiogenic, antimetastatic, and anti-inflammatory effects of wogonin. The co-effects of wogonin with traditional cancer medications are also considered, giving the most effective combinations for additional research. In addition, the use of contemporary nanotechnological techniques to address the low bioavailability problems typical of natural chemicals is explored. Therefore, the mini-review will offer various aspects of wogonin at a single platform for designing in vivo animal studies and human clinical trials, with the goal of eventually using it in therapeutic settings.
Chemistry of wogonin
Wogonin is the main component found in S. baicalensis, a medicinal plant belongs to the Lamiaceae family 19 and native of China. Chemically it is 5,7-dihydroxy-8-methoxyflavone (Figure 1) having molecular weight of 284.267 g/mol. It is crystalline solid with maximum solubility in organic solvents and limited aqua solubility. As far as concerned synthesis it can be extracted from plants and can also be prepared by chemical synthesis 20 in industry. Recently, Li et al. 21 reported an efficient approach using chrysin as starting material for the production of wogonin. The pharmacokinetic results indicated that wogonin has a half-life of 14 min (40 mg/kg) when administered intravenously to rat models. Wogonin (100 mg/kg) reached its Cmax value of 300 ng/mL after 28 min of intragastric administration. Rat urine, bile, and feces all had only 21% of wogonin eliminated unaltered.20,22,23 The biological availability of wogonin can be altered by substituting the hydroxyl groups present in its structure. It was found that the presence of methoxy group at position 8 is responsible for biological potential of wogonin, which is further evident from the reports of many researchers who synthesized wogonin derivatives (Figure 2), keeping the 8-methoxy group intact while replacing the hydroxyl groups. 24 A previous study reported that the non-toxic dose of wogonin in rats is 30 mg/kg when administered intravenously. However, the long period of wogonin treatment with high doses of around 120 mg/kg was toxic and could induce heart injury, as observed in rats (22).
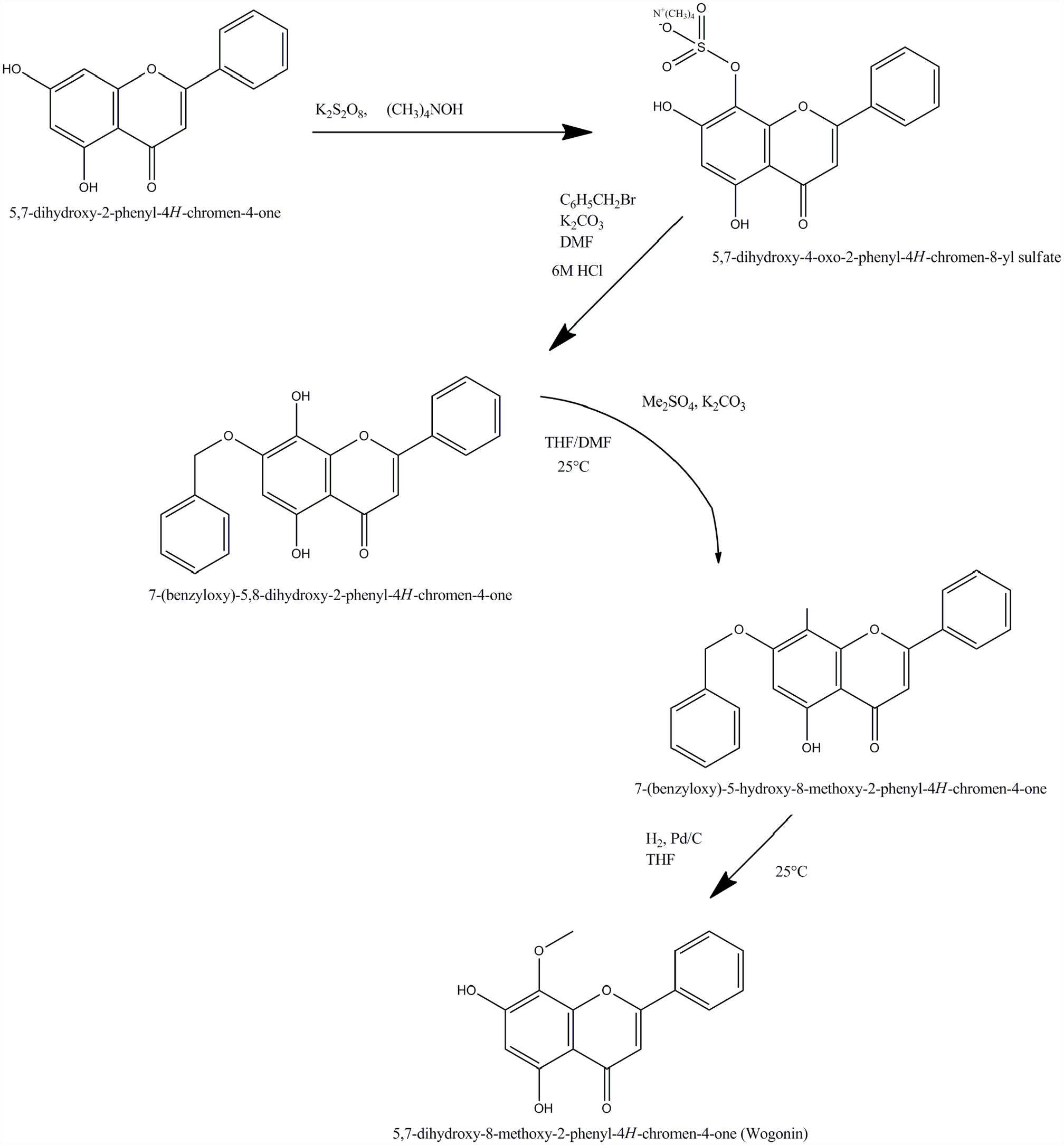
Synthesis of wogonin using chrysin as starting material.
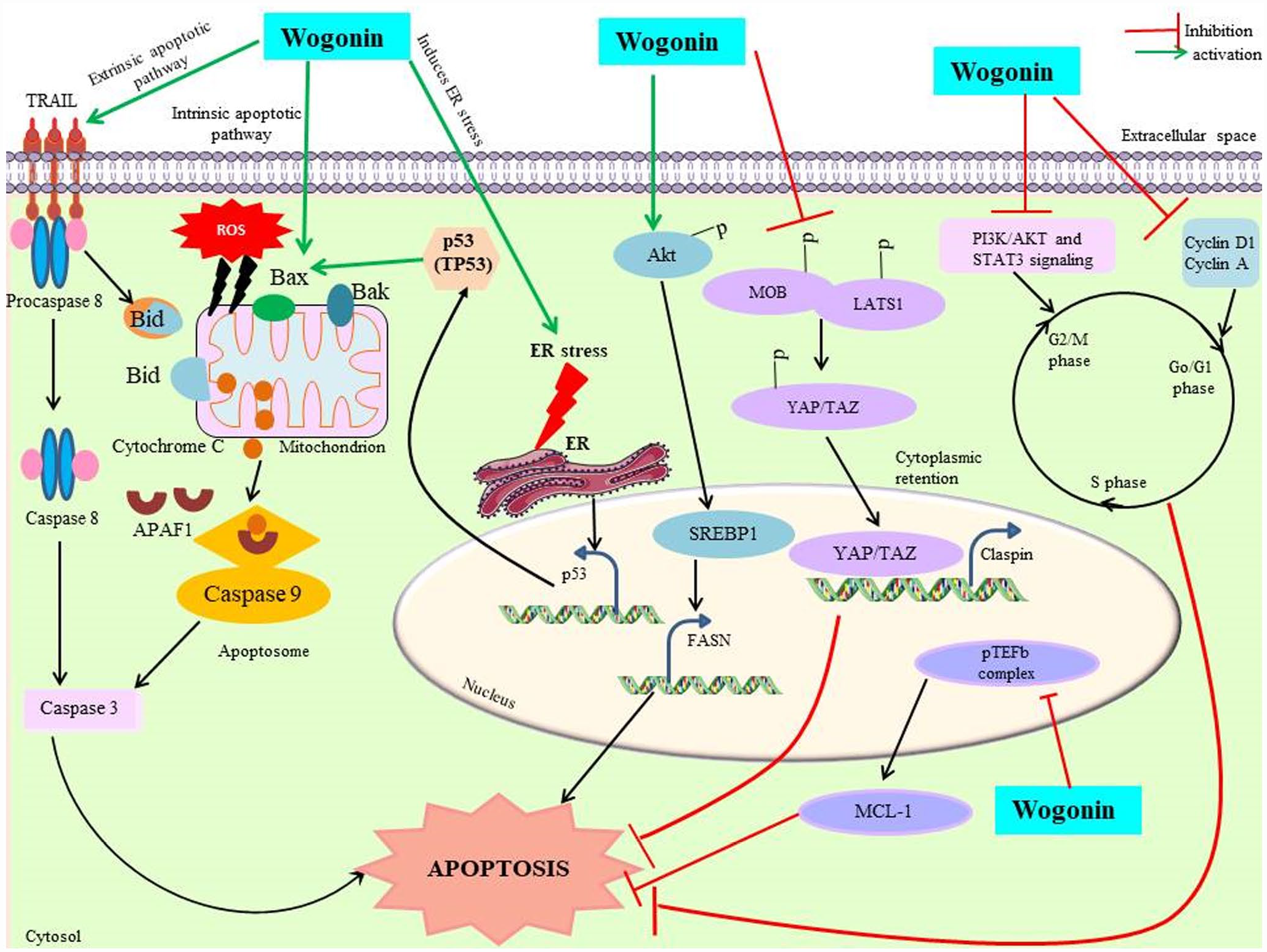
A pictorial representation of detailed underlying wogonin-mediated antiapoptotic effect.
Cellular interactions of wogonin in cancer
Apoptosis and cell cycle arrest
In apoptosis, cells are killed by controlled cell death, one of the key pathways which mediate anticancer effects of therapeutic molecules avoiding any inflammatory responses generated due to other pathways. Moreover, natural bioactive molecules are actively being studied for their potential to initiate this pathway for achieving eradication of cancer cells. Wogonin is a mono-flavonoid obtained from the extracts of S. baicalensis. Numerous reports suggest that wogonin exhibits anticancer potential by regulating cell proliferation, apoptosis (Figure 2), and metastasis. The mono-flavonoid was reported to activate tumor cell death in prostate carcinoma, in vitro and in vivo. In vitro experiments including wogonin treatment of DU145 and 22Rv1 cell lines at the concentration of 100 µM resulted in phosphorylation of AKT protein and increased nuclear localization of sterol regulatory element-binding protein 1 (SREBP1). Subsequently, apoptosis was stimulated due to the elevated levels of fatty acid synthase, which led to increased mitochondrial oxidation, free radical formation, and reduction of membrane potential of mitochondria mediated by carnitine palmitoyl-transferase 1A. Furthermore, wogonin was also able to repress tumor growth in prostate cancer xenograft at 100 mg/kg without any significant toxicity. The above mechanism suggests that wogonin induces cell death by inducing AKT-SREBP1-FASN signaling mechanism in prostate cancer. 25 However, in lung cancer cell lines A549 and H460, wogonin was found to stimulate tumor cell death by downregulating p-AKT and c-Myc mediated by MMP-1 levels. 26 Furthermore, wogonin also induces apoptosis at 50 µM through the formation of reactive oxygen species (ROS) and induction of caspase-3/8/9 axis in lung cancer as reported in A427 cell line. 27 Moreover, no cytotoxicity was reported at 50 µM in normal BEAS-2B cell line. Similarly, wogonin was shown to induce apoptosis in A2780 ovarian cancer cell line at 200 µM, which was supported by chromatin condensation and degradation of nuclear membrane. 18 Wogonin also regulates the activation of YAP and TAZ by inducing MOB1-LATS1 signaling, which downregulated Claspin, thus inducing apoptosis in SMMC-7721 and HCCLM3 hepatocellular carcinoma cell lines. 17 In, CRC HCT116 cells, wogonin was observed to generate endoplasmic reticulum (ER) stress which activates cytoplasmic localization of phosphorylated-p53 and caspase-mediated apoptosis. 28 Another study on a panel of DLD-1, SW48, HCT-15, LS-180, and SW1417 CRC cells reported apoptotic effect of wogonin induced by upregulation of caspase-3/8/9, Bax, and inhibition of PI3K/AKT and STAT3 pathways. 13 Aerobic glycolysis is the mainstay for to complete energy requirement by ATP formation in tumor cells. Anticancer potential of wogonin mediated by glycolytic pathway was studied across ovarian, colon, and hepatocellular cancer (HCC) cells. Wogonin showed p53-mediated increase in glycolysis and apoptosis regulator (TIGAR) and downregulation of glucose transporter 1 (GLUT1). Wogonin activates and stabilizes p53 by phosphorylation, acetylation, and inhibiting MDM2. However, these effects were restricted to p53-expressing cells and not in mutant p53-expressing cells. 29 Treatment with wogonin in chronic myeloid leukemia in K562 and KU-812 cells stimulates apoptosis by downregulating MCL-1, which is downstream to P-TEFb kinase and mediated by the inhibition of CDK9. 30
Another potential target of this therapeutic molecule to arrest cancer cells in specific phases is to allow control over indiscriminate proliferation and ultimately toward cell death (Figure 2). Wogonin reportedly arrests HCT116 CRC cells in G1 phase. The arrest in G1 phase occurs as a consequence of modulation of Wnt/β-catenin signaling pathway and involves downregulation of four cell cycle-related proteins, such as cyclin A, E, D1, and CDK2. 14 However, SW48 CRC cells were reported to be arrested in G2/M phase probably mediated by the suppression of PI3K/AKT and STAT3 signal transduction mechanisms. 13 Similarly, wogonin has been reported to arrest MCF-7 breast cancer and HL-60 leukemia cells in G1 phase by blocking the PI3K/AKT signal pathway.31,32 According to studies, wogonin inhibits A2780 ovarian cancerous cells in G1/G0 phase. 18 Wogonin arrests MHCC97L, HepG2 HCC cells and C6, U251 glioma cells in G0/G1 phase by inhibiting Cyclin D1 expression.33,34
Antiangiogenesis and antimetastasis action
During angiogenesis, new blood vessels are formed, controlled by certain biomolecules that help in providing oxygen and nutrition to cancerous cells for their growth, differentiation, and migration. 35 Targeting proteins associated with angiogenesis (Figure 3) have been summarized to affect tumor growth and metastasis and therefore have become therapeutic targets. 36 Wogonin obtained from the root extract of S. baicalensis has been studied to reduce tumor growth and metastasis in lung, liver, and kidney cancers in mice bearing LM8, at a dosage of 25 and 50 mg/kg, twice per day. Wogonin at 10–100 μM also suppressed phosphorylation of vascular endothelial growth factor receptor-3 (VEGFR-3) induced by vascular endothelial growth factor C (VEGF-C) in human lymphatic endothelial cells (HLECs) cells. 37 A study has shown that upregulation of metastasis suppressor protein called maspin was observed on administration of wogonin. It showed that wogonin at 1–10 μM inhibited ERK1/2, migration, and invasiveness in GBC-SD cancerous cell line. 38 The anticancerous mechanism has been studied to modulate signaling networks by regulating AKT protein, AMP-activated protein kinase (AMPK), cell death dependent or independent of p53, decreased DNA damage, growth and migration/metastasis of malignant cells, and inhibition of activity of telomerase. 39 Antimetastasis of hepatocarcinoma cell lines MHCC97L and PLC/PRF/5 was observed in the presence of wogonin. It was observed that binding of wogonin and MMP-9 was responsible for the inhibition of MMP-9 enzymatic function that subsequently leads to the inhibition of metastatic tumor growth. 40 Wogonin has also been studied to pharmaceutically target proteins, such as JAK/STAT, VEGF/VEGFR, Wnt/β-catenin, and tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)-mediated cell death in many cancerous cells. 15 Migration of human CRC cell lines, HCT116 and SW480, has been studied to show anticancerous activities on wogonin administration by suppressing IRF3 expression while upregulating p-YAP1. 41
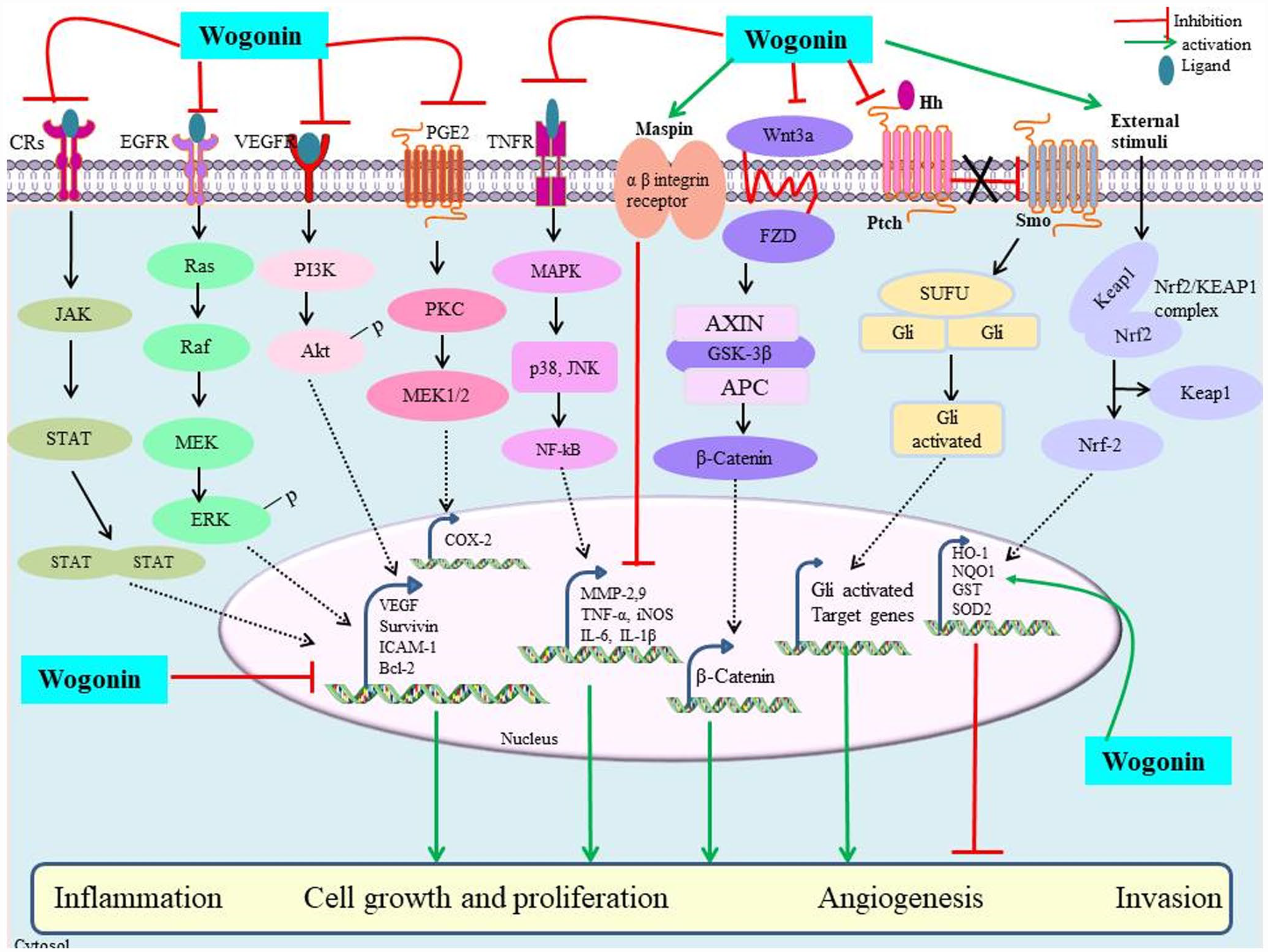
A detailed representation of antiangiogenesis and antimetastatic mechanism of wogonin.
Anti-inflammatory action
Inflammation arises during events, such as injuries, which leads to release chemotactic factors. It has been studied to be critical component of tumor progression, as many cancers have been studied to arise from infection sites, chronic irritation, and inflammation. Studies have accepted a relationship between inflammation, innate immunity, and cancer. 42 According to studies, tissue injuries release inflammatory proteins that lead to mitogenesis of neoplastic cells and reconstruction of normal tissues. These may potentiate tumor growth, stimulate angiogenesis, and migration. Studies are being focused on understanding anti-inflammation and anticancerous activities of compounds. 42 Studies have shown that wogonin was studied to show significant inhibition of inflammation in HT144 melanoma cell lines. It showed decrease in expression of inflammation-promoting factors led to the reduced levels of inflammatory cytokines; while increase in the levels of anti-inflammatory factors. It has been explained by inhibiting Hh signaling dependent control of inflammation in the presence of wogonin. 43 Wogonin dosed at 250–1000 μg/ear/three days was observed to lower mRNA transcripts of tumor necrosis factor alpha (TNF-α) and COX-2 in a intermediate skin inflammation. 44 A study showed that it reduced ear thickness and skin inflammation in dermatitis mouse models. It suppressed the levels of inflammation-mediating agents, such as TNF-α. 44 Wogonin lowers the expression of PMA-induced COX-2 by PKC and MEK1/2, resulting in the degradation of AP-1 and c-Jun proteins in human lung carcinoma A549 cells. 45 Similarly, a study showed that it reduced the formation of inflammation-promoting cytokines (i.e. IL-6, IL-1β) and also inhibited the phosphorylation levels of several proteins, such as ERK, p38, Nrf2, and NF-κB, in dextran sodium sulfate (DSS)-administered mice models having CRCs, and in THP-1 and HCT116 cancerous cells. 46 Recently, it has shown that wogonin, at a dose of 40 mg/kg, ameliorated nephrotoxicity, inflammation, free radical formation, and cell death in rat models with renal injury. It regulated different signaling cascades, such as PPAR-γ and Wnt/β-catenin. 47 It reduced the expression levels of cyclooxygenases-2 (COX-2) and prostaglandin E2 (PGE2) in murine skin fibroblasts. 48 It also reduced inflammatory response proteins, including inflammatory agents induced by IL-1β, IL-6, iNOS, and activated NAD(P)H quinone dehydrogenase 1 (NQO1), superoxide dismutase 2 (SOD2), and Nrf2/HO-1. 49 Wogonin at a concentration of 50 and 100 μM has also been observed to exhibit dose-dependent suppression of NO production, simultaneously reduced the proportions of S/G2 and induced apoptosis in YAC-1 cancer cells. 50
Synergistic effects of wogonin
Cancer, a global wreck, is really arduous to curb due to limited effective or targeted treatments, though numerous treatment regimens and surgeries are present. 51 The standard strategies used for chemotherapy or treatment of cancer offer minimal survival benefits with numerous adverse effects and drug resistance. 52 Naturally derived bioactive compounds or secondary metabolites have been reported to potentiate chemopreventive methods for treating cancer, and such combinatorial or synergistic approaches exhibit remarkable efficacy in cancer therapy due to their multitargeted actions, minimal side effects to the normal with little or no drug resistance, and lack of considerable toxicity.52,53 Wogonin was extensively studied and discovered to have a strong potential to combat the majority of malignancies (Table 1) through a variety of techniques. 51 In addition, studies indicate that wogonin may enhance the effectiveness of some medications that are used in cancer treatment and chemoprevention. According to research, it has been shown that this compound causes tumor cells to become more sensitive to chemotherapy drugs, such as etoposide, TNF, adriamycin, and TRAIL. It was observed that the enhanced caspase-3 activation led to the cleavage of its substrate PARP in cells administered with wogonin and cisplatin; the bioactive compound has been reported to sensitize tumor cell death induced by cisplatin in A549 and HeLa cell lines, thus providing significant evidence in favor of wogonin’s prospective usage to sensitize cisplatin in cancer treatment. 54 Likewise, studies revealed that in HL-60 and Jurkat cell lines, this flavone, wogonin dramatically increased the amount of etoposide-mediated cell death and enhanced lung cancer cell death caused by etoposide, as well.55,56 In addition, the bioactive molecule was reported to offer an imperative function in overcoming multidrug resistance by reducing the secretion of the chemopreventive drug mediated by P-glycoprotein and improving the therapeutic effectiveness of it in cancerous cells. 57 Moreover, the compound, wogonin has also been shown to potentiate navitoclax-mediated tumor cell death in various tumors, and in Acute myeloid leukemia (AML) and Acute lymphocytic leukemia (ALL) cancerous cells by reduced expression levels of Mcl-1 besides promoting the regression of tumors in living tissue in a human T cell leukemia mouse xenograft model. 58 Studies have demonstrated that wogonin in combination with cisplatin or paclitaxel suppressed the tumor proliferation in gastric cancerous cells under in vitro condition. 59 Furthermore, the wogonin study improved the anticancerous function of other therapeutic medications as well. There is evidence that the compound in combination with a first-generation protease inhibitor, Bortezomib (BTZ), in a clonal plasma cell tumor, multiple myeloma (MM) regulated by signaling cascades linked with tumor growth, medication resistance, and apoptosis. 60 Furthermore, it was investigated that wogonin potentiates TRAIL-induced growth inhibition and tumor cell death in A549 cancerous cells, as shown by the TUNEL assay. 61 In addition, it was also demonstrated that wogonin therapy reinstates the TRAIL-mediated apoptosis in the presence of ROS by upregulating p53 and PUMA in TRAIL-resistant cancer cells.62,63 Recently, a wogonin derivative (i.e. FV-429) was reported to be considered as a promising therapeutic candidate for metastatic non-small cell lung cancerous cells (NSCLC) by enhancing the susceptibility of NSCLC cells to exposure to paclitaxel through inhibition of cell cycle arrest at G2/M phase and by deactivating the Wnt signaling mechanism 64 and also reversing paclitaxel aversion in human epithelial ovarian carcinoma by blocking c-Src/Stat3/HIF-1α signaling cascade and inhibition of cell cycle arrest at G2/M phase. 65 Moreover, the combinatorial therapy of wogonin and 5-fluorouracil has resulted in reduced survival and substantial combined inhibitory function on elevated levels of COX-2 in SMMC-7721 HCC cells. 66
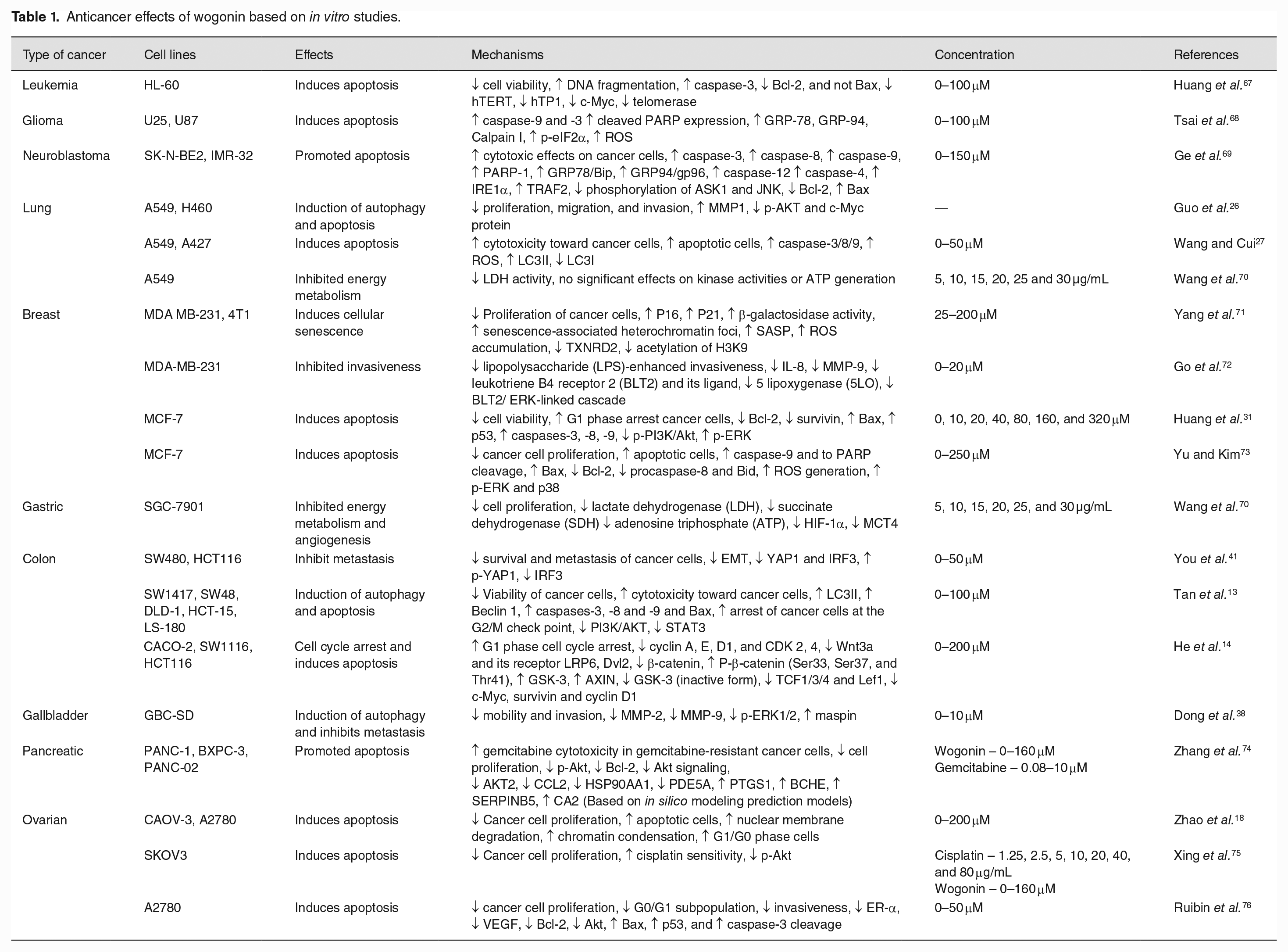
Anticancer effects of wogonin based on in vitro studies.
Conclusions
A potential flavonoid produced from S. baicalensis is called wogonin. The review summarized research on wogonin’s chemistry, anticancer, and synergistic activity. The key underlying mechanisms for wogonin’s biological actions are, as was already mentioned, its anti-inflammatory and antioxidant properties. However, no research has been conducted addressing the DNA-damaging activity and potential toxic effects of wogonin, over the long term and in the short term. Furthermore, toxicological profile of wogonin will be warranted to elucidate its translational potential and could be used as a promising therapeutic medication for cancer therapy. Therefore, wogonin’s safety must first be clarified, along with their metabolic conversion and potential bioactivities of different metabolites. Second, it is necessary to elucidate on the appropriate dosage schedules. In this sense, the current review offers a solid foundation for moving forward with these following steps to ultimately discovering unique, secure, and more effective anticancer agents.
Footnotes
Authors’ Contributions
HST, PR, AC, GP, HJ, and NCP did the conceptualization, methodology, validation, and writing. Formal analysis and data curation were performed by IR, SR, DA, MK, and RR. All authors have read and agreed to the published version of the manuscript.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.