
Abstract
Defects in migration and invasion caused by dysregulation of trophoblast epithelial-mesenchymal transformation (EMT) are one of the key factors in the pathogenesis of preeclampsia (PE). RNA-binding motif protein 25 (RBM25) is an RNA-binding protein involved in a variety of cellular processes, including cell proliferation, apoptosis, cell migration and invasion, and EMT. However, the expression and function of RBM25 in placental of PE remain unclear. In this study, we reveal that the expression of RBM25 is significantly elevated in PE placental tissue. RBM25 depletion and over-expression in trophoblast cells increase and decrease, respectively, cell migration and invasion by regulating EMT marker E-cadherin and Vimentin expression. Mechanistically, Grhl2 is involved in RBM25-regulated trophoblast cell migration, invasion, and EMT through RBM25-facilitated mRNA stabilization. Furthermore, the upregulation of Grhl2 enhances the expression of RBM25 through transcription and forms a positive feedback regulation in the progression of PE. These findings suggest that upregulation of RBM25 induces dysregulation of trophoblast EMT by enhancing positive feedback regulation of Grhl2 and RBM25, leading to defects in cell migration and invasion. Targeting this newly identified regulatory axis may provide benefits in the prevention and treatment of PE.
Impact statement
Preeclampsia (PE) is a pregnancy-specific disorder characterized by proteinuria, hypertension, or renal insufficiency, which is the leading cause of maternal and perinatal mortality. However, the definitive pathogenesis of PE is not fully understood. Emerging studies confirm that defects in migration and invasion caused by dysregulation of epithelial-mesenchymal transformation (EMT) are one of the key factors in the pathogenesis of PE. In this study, our findings suggest that upregulation of RBM25 induces dysregulation of trophoblast EMT by enhancing positive feedback regulation of Grhl2 and RBM25, leading to defects in cell migration and invasion. Targeting this newly identified regulatory axis may provide benefits in the prevention and treatment of PE.
Introduction
Preeclampsia (PE) is a pregnancy-specific disorder characterized by proteinuria, hypertension, or renal insufficiency, 1 which is the leading cause of maternal and perinatal mortality with the incidence of 5–7%.2,3 In the first trimester, invasive extravillous trophoblasts (EVTs) are responsible for remodeling the spiral arteries of the uterus to meet the nutritional supply required by the fetus. 4 Although the definitive pathogenesis of PE is not fully understood, impaired spiral artery remodeling due to insufficient invasion of trophoblast cells is considered one of the causative factors of PE. 5 Epithelial-mesenchymal transformation (EMT) is the transformation of resting epithelioid cells into cells with mesenchymal cell characteristics after a series of gene expressions. 6 EMT is a normal physiological process in human cells that gives cells greater ability to invade and migrate.7,8 Emerging studies confirm that defects in migration and invasion due to EMT dysregulation of trophoblast cells may be one of the important causes of obstruction of spiral artery remodeling. 9 Therefore, revealing the molecular mechanism of EMT in trophoblasts will provide new methods and targets for the prevention and treatment of PE.
RNA-binding motif protein 25 (RBM25) is mainly localized in the nuclear speckles. 10 It has been reported that RBM25 contains a common arginine glutamate enrichment core region, which is conducive to its binding to splicing factor enriching nucleosomes on one hand, and to the binding of its C-terminal PWI region to target RNA on the other hand. 11 Therefore, RBM25 is generally considered to be a splicing factor for the RNA-binding protein (RBP) family. For example, RBM25 regulates selective splicing of Bcl-x pre-mRNA by binding to the CGGGCA sequence of the second exon on Bcl-x RNA. RBM25 controls splicing of key genes such as BCL-X and the MYC inhibitor BIN1. 12 In addition, RBM25 also regulates a variety of intracellular signaling pathways such as AKT and EPO signaling pathways. 13 In addition, RBM25 functions much more than that as an RBP. A recent study found that RBM25 is involved in cellular EMT processes and tumor cell migration by binding and promoting circAMOTL1L biogenesis. 14 Fascinatingly, Jacobo-Baca et al. 15 used proteomic profile to find a significant increase in RBM25 expression in serum of PE pregnant women. However, the expression and function of RBM25 in PE placental are unclear.
In this study, we revealed that the expression of RBM25 is significantly elevated in PE placental tissue. The upregulation of RBM25 inhibited trophoblast cells migration and invasion by regulating EMT marker genes expression. Our findings reveal, for the first time, that Grhl2 is involved in RBM25-regulated trophoblast cell migration, invasion, and EMT through RBM25-facilitated mRNA stabilization. Further exploration found that Grhl2 promotes RBM25 expression through transcription, and positive feedback regulation between them promotes the occurrence and development of PE.
Materials and methods
Clinical samples
The placental tissue of 30 PE patients and 30 normal maternal were collected from the obstetrics department of Shijiazhuang Fourth Hospital, from June 2019 to December 2018. Tissue on the surface of the placenta was collected according to the ACOG 2013 PE diagnostic criteria. The study protocol was approved by the Ethics Committee of Shijiazhuang Fourth Hospital and with the written consent of each patient.
Cell culture and transfection
Human villous extratrophoblast cell lines HTR8/SVneo, CM-H051 and human placental cell line 3A-subE were preserved in our laboratory. Human normal placental cell line Hs 795.Pl (ATCC CRL-7526™) was purchased from ATCC® (MD, USA). The first three cells used RPMI 1640 medium (Gibco, Beijing, China) and the latter used Dulbecco’s modified eagle medium (Gibco). All cell culture systems were supplemented with 10% FBS (Gibco), 100 units/mL penicillin, and 100 μg/mL streptomycin (Gibco) and maintained at 37°C in a humidified atmosphere containing 5% CO2. Transfection of cells is performed using the operating manual of Lipofectamine 2000 (Invitrogen, CA, USA). Briefly, 16 HTR8/SVneo cells (1 × 105 cells/mL) are seeded into a growth dish until the cells grow to approximately 80 confluence. The cells are washed twice with PBS and then the mixture of the vector with liposomes is added to the cells in serum-free medium and mixed well with gentle shaking. The complete medium is replaced after 6 h of incubation, and incubation is continued for another 48 h.
QRT-PCR analysis
Total RNA is extracted from tissue and cultured cells according to the RNA kit II (Omega, #R6934) manufacturer’s operating manual. The concentration and quality of total RNA are checked using the NanoDrop 2000 spectrophotometer. The first-strand cDNA is synthesized using MonScript™ RTIII Super Mix with dsDNase (Two-Step) and oligonucleotide (dT) primers. qRT-PCR analysis of mRNA using the following primers was performed on the ABI 7500 FAST system using MonAmp™ ChemoHS qPCR Mix (Mona, China). GAPDH is used as a reference gene for total RNA, calculated using the 2-ddCt formula described earlier [26,27]. All primers used in this study are as follows: RBM25F: CGTGCACTCAGATTATTACATGACCTGC; RBM25-R: GGGCATTTAGCTCACTGGAGTATTCACG; ZEB1F: CTGCTGGGAGGATGACACAG; ZEB1-R: ATGACCACTGGCTTCTGGTG; Grhl2F: ACCTGTAGC-TC-TTGTTCTGCCAT; Grhl2-R: TACAGTTGCCATCAGC-CCAAA; GAPDH-F: AATGGGCAGCCGTTAGGAAA; and GAPDH-R: GCGCCCAATACGACCAAATC.
Western blotting
Proteins are extracted from cells and frozen placental tissues using radioimmunoprecipitation assay (RIPA) lysis buffer and protease inhibitor mixture. 17 Protein concentration was quantified using the modified Bradford method. An equal amount of protein is then loaded onto the gel. After separation by SDS-PAGE, proteins are electrically transferred to a polyvinylidene fluoride (PVDF) membrane (Miripore, IPVH00010) and blocked with 5% skim milk. Finally, the membrane bound to the protein is incubated with different primary antibodies overnight at 4°C. The antibodies used were as follows: RBM25 (Abcam, 1:1000, ab72237), Grhl2 (Abcam, 1:1000, ab271023), E-cadherin (Abcam, 1:500, ab231303), Vimentin (Abcam, 1:1000, ab8978), ZEB1 (Abcam, 1:1000, ab203829), and β-actin (Proteintech, 1:1000, 66009-1-Ig). The next day, the PVDF membrane was washed and reacted with HRP-labeled secondary antibodies (1:10000; Rockland, PA, USA) for 30 min. The membrane was treated with Immobilon Western Chemiluminescence HRP substrate and photographs were acquired at Fuazon™ Fx (Vilber Lourmat, PR, FR) by enhanced chemiluminescence method.
Hematoxylin and eosin, immunohistochemistry staining
Fresh PE and normal placental tissue are fixed in formalin and sliced 4 μm thick. Tissue sections are preformed to hematoxylin and eosin and immunohistochemical staining. Immunohistochemical staining analysis was performed as described previously. 18 Before antigen retrieval, tissue sections are dewaxed and immersed in water. After treatment with H2O2, tissue slides are blocked with 5% goat serum for 30 min. Then, these sections are incubated with the primary antibody overnight at 4°C. The next day, HRP-conjugated secondary antibodies are incubated with tissue slides and visualized using DAB treatment.
Immunofluorescence staining
Cells cultured on glass slides are fixed and permeabilized with 0.5% Triton X-100 for 5 min each three times. And then 5% goat serum was used to block the slides as previously described. 19 The slides were incubated with primary antibody overnight at 4°C. The fluorescein-labeled secondary antibody reacts with a glass slide and then the nucleus is colored with DAPI. Images were acquired with a Leica microscope (DM6000B; Leica, Switzerland) and digitally analyzed using LAS V.4.4 (Leica).
RNA immunoprecipitation assay
Interaction between RNA and protein is examined by RNA immunoprecipitation (RIP) as described previously. 14 Briefly, cells are harvested and lysed using NETN buffer. The manufacturer’s instructions were followed for RIP experiments using anti-labeled antibodies or IgG and Dynabeads™ Protein G Immunoprecipitation Kits (10007D; Thermo Fisher). Next, the Dynabeads bead-RNA complex is washed with NETN buffer for three times. RNA isolation and purification was performed using the RNA kit II (#R6934; Omega, GA, USA) according to the manufacturer’s protocol. Finally, RNA was detected and quantified using NanoDrop 2000 (Thermo Fisher, MA, USA) for subsequent RT-qPCR analysis.
Co‑immunoprecipitation assay
Co‑immunoprecipitation (CoIP) was performed by a Pierce™ Classic Magnetic IP/Co-IP Kit (88804; Thermo Scientific™, MA, USA). 20 Briefly, cultured cells were lysed by lysis/wash buffer in ice and then lysates were immunoprecipitated with anti-RBM25 or IgG for 1 h at room temperature. The prewashed pierce protein A/G magnetic beads were added to the lysates for incubating 1 h at room temperature. The bead–antigen–antibody complex is collected through the magnetic rack and washed with elution buffer. Western blotting is used to detect bound proteins.
ChIP assay
HTR8/SVneo cells were treated with formaldehyde for ChIP assay as previously described. 21 The cross-linked chromatin was sonicated to an average size of 300–500 nt. After dilution, the sample is incubated with salmon sperm DNA-treated protein A-agarose. DNA fragments were immunoprecipitated with anti-GRHL2 or anti-IgG antibodies. After the cross-linking was reversed, the Grhl2 occupancy on the RBM25 promoter was detected by qRT-PCR. The ChIP primer used are as follows: Proximal-F: GGTTACAGGCGTGAGCCACCG; Proximal-R: GCACTCATAACCATCTATCATAACATAC; Distal-F: CAACCTGGCCAACATGGTGAAACC; Distal-R: GAGGAGGTGTCTCTGTTTCCCAGGC.
Transwell assays
After the cultured cells were treated accordingly, the cells were implanted into the top compartment of the insert (3422; Corning, NYS, USA) and allowed to migrate. 14 After 24 h, the lower surface cells were fixed with 100% ice methanol and stained with 0.05% crystal violet.
Three-dimensional Matrigel drop invasion assays
Cells were performed 3D Matrigel drop invasion assay as described previously. 22 HTR8/SVneo cells were transfected as indicated and then suspended in 10 μL Matrigel. The 5 × 104 cells were mixed with the matrix and quickly dropped into the 12-well plate. The medium was then added slowly after waiting 15 min for matrix drops to form. The radial distance between the cells and the edge of the matrix was measured after seven days of cell culture.
Luciferase assays
RBM25 2000 bp promoter was amplified and constructed into pGL3-basic plasmid and then used Sanger sequencing to confirm the sequence. HTR8/SVneo cells were cultured in a 24-well plate and co-transfected with indicated vectors. Luciferase activity was measured as previously described 23 by Dual-Glo Luciferase Assay System (Promega, WI, USA) with a Flash and Glow reader (LB955; Germany). The activity of RMB25 promoter was expressed as the relative activity ratio of firefly luciferase to Renilla luciferase.
Statistical analysis
All data are expressed as the mean ± SEM of three independent experiments. Student’s t-test was used to analyze differences between two groups. Values of P < 0.05 were considered statistically significant. GraphPad Prism 8.0 software was used for the statistical analysis (GraphPad Software).
Results
RBM25 is upregulated in PE placental tissue
Previous studies reported upregulated expression of RBM25 in plasma samples of preeclampsia (PE) patients. 15 In order to investigate the expression and function of RBM25 in placental tissues of PE pregnant women, we collected placental tissues of normal and PE pregnant women, respectively. First, hematoxylin and eosin (HE) staining was used to identify these tissues (Figure 1(A)). We then randomly selected normal and PE placental tissues and detected RBM25 expression in these samples. The results showed that the protein and mRNA level of RBM25 were significantly increased in PE tissues compared with normal placental tissue (Figure 1(B) to (D)). The same results were obtained in immunohistochemical staining of placental tissue (Supplementary Figure 1). Next, we analyzed clinicopathologic features in normal and PE pregnant women. As shown in Supplementary Table 1, there were statistically significant differences in systolic blood pressure, diastolic blood pressure, 24 h urinary protein, gestational age at delivery and neonatal birth weight, but not with age and prepregnancy body mass index (BMI). Subsequently, we measured RBM25 expression in different placental cell lines and found that it was significantly upregulated in HTR-8/SVneo trophoblast cell lines (Figure 1(E)). In the following experiments, we will investigate the function of RBM25 in HTR-8/SVneo cell lines. These results revealed that RBM25 also showed high expression in PE placental tissues.
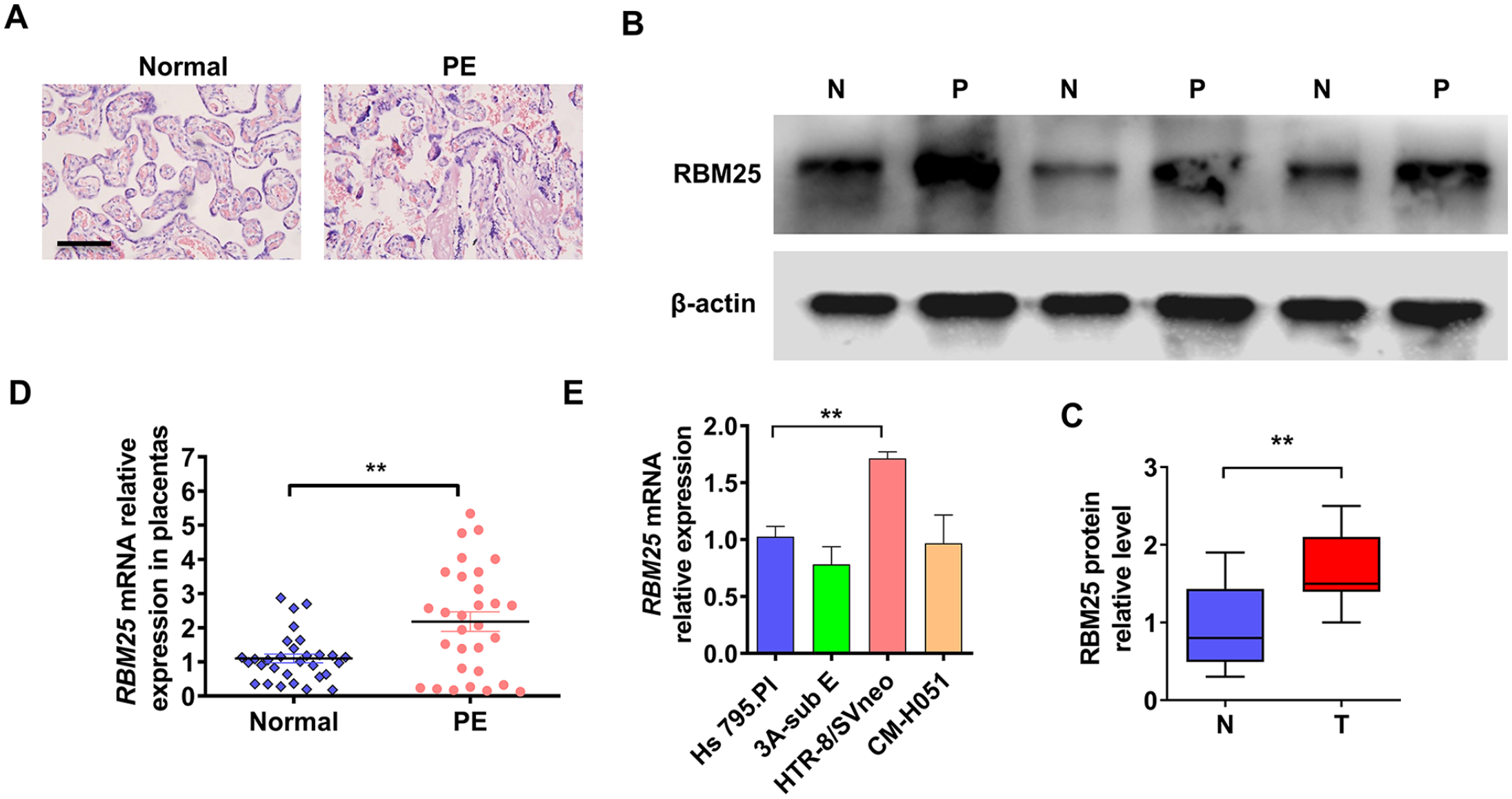
The expression of RBM25 was upregulated in PE tissues. (A) Hematoxylin and eosin (HE) staining was used to identify normal and preeclampsia (PE) placental tissues. Scar bar = 100 μm. (B) Placental tissues from three normal and three PE cases were randomly selected for Western blot analysis of RBM25 protein expression. (C) Quantitative analysis of Western blot results in (B). (D) RT-qPCR was used to examine RBM25 mRNA expression in normal (n = 30) and PE (n = 30) placental tissues. (E) RBM25 expression was measured in cell lines derived from placental tissue by using RT-qPCR. All data were from three independent experiments and were presented as a mean ± standard error.
Depletion of RBM25 promotes trophoblast cells migration and invasion
Studies reported that decreased trophoblast cells migration and invasion is one of the key factors promoting the development of PE during pregnancy. 24 To explore whether RBM25 is involved in trophoblast cell migration and invasion, we first constructed two shRNAs targeting RBM25. As shown in Figure 2(A) to (C), the mRNA and protein level of RBM25 were obviously decreased in both shRNAs transfected-HTR-8/SVneo cell. Next, HTR-8/SVneo cell was transfected with both RBM25 shRNAs and a transwell assay was performed to examine the cells migration. The results showed that knocking down RBM25 significantly promoted the migration of trophoblast cells (Figure 2(D) and (E)). Subsequently, the 3D Matrigel droplet assay was used to explore the invasion of HTR-8/SVneo cells. As shown in Figure 2(F) and (G), depletion of RBM25 markedly promoted the invasion of HTR-8/SVneo cells. These data support a role for RBM25 in trophoblast cells migration and invasion regulation.
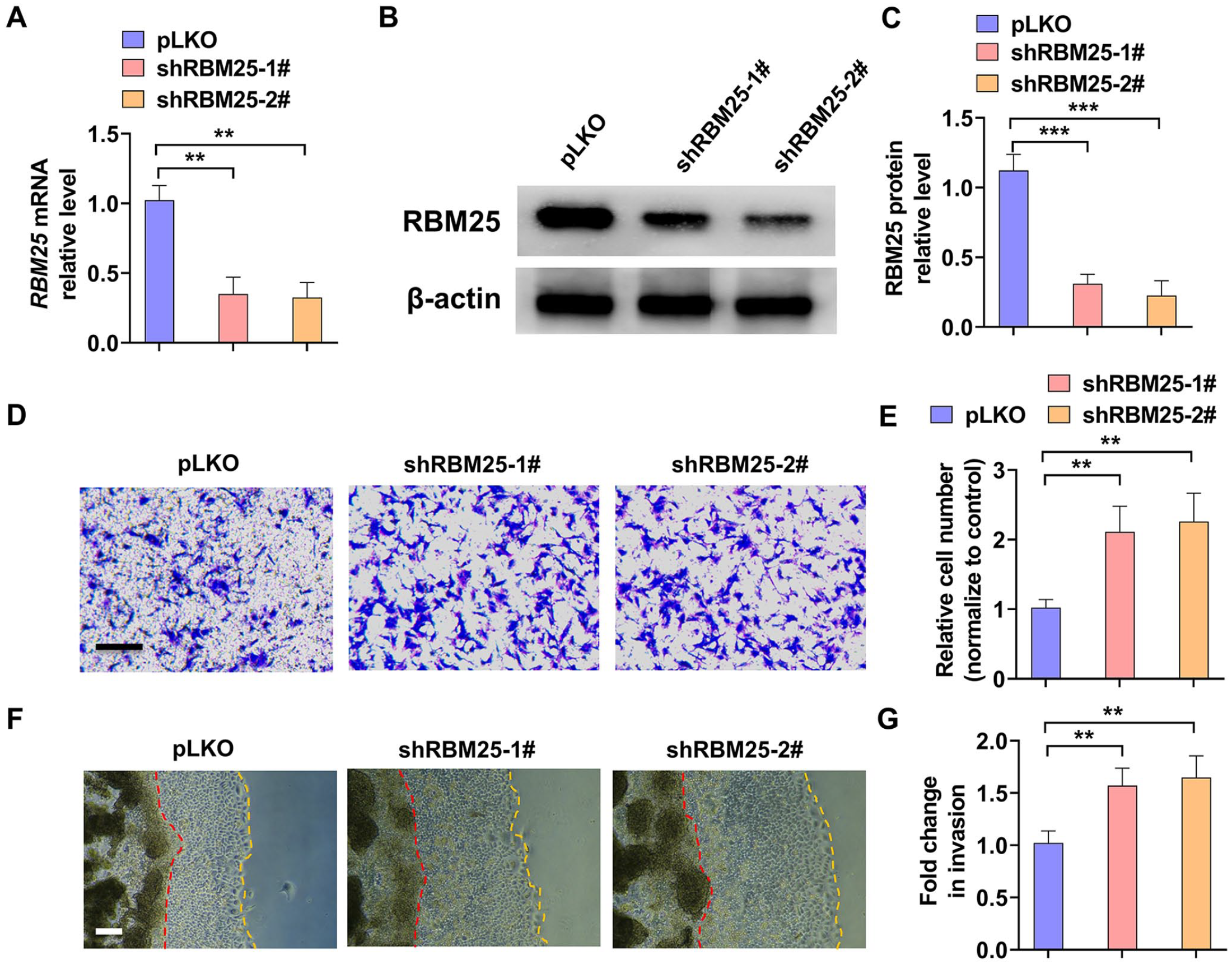
Knockdown of RBM25 promoted trophoblast cells migration and invasion. (A) HTR-8/SVneo trophoblast cells were transfected with RBM25 shRNAs (shRBM25-1# and shRBM25-2#) or control vector (pLKO) and then RT-qPCR was used to detect RBM25 mRNA expression. (B) HTR-8/SVneo cells were transfected as in (A) and Western blot was used to examine RBM25 protein level. (C) Quantitative analysis of Western blot results in (B). (D) HTR-8/SVneo cells were transfected as in (A), and then a transwell assay was used to explore cells migration. Scar bar = 50 μm. (E) Quantitative analysis of relative cell number of migration in (D). (F) A 3D Matrigel droplet assay was used to explore the invasion of HTR-8/SVneo cells after transfected as in (A). Scar bar = 100 μm. (G) Quantitative analysis of fold change of cell invasion in (F). All data were collected from three independent experiments and were presented as a mean ± standard error.
Over-expression of RBM25 depresses trophoblast cells migration and invasion
To further confirm the role of RBM25 in trophoblast cell migration and invasion, we constructed a vector for RBM25 over-expression. As shown in Figure 3(A) to (C), transfection of oeRBM25 over-expression vector significantly elevated the mRNA and protein level of RBM25 in HTR-8/SVneo cells. Then, a transwell assay was used to detect the cells migration. The results showed that over-expression of RBM25 markedly decreased the migration of HTR-8/SVneo cells (Figure 3(D) and (E)). In parallel, we used the 3D Matrigel droplet assay to examine the cells invasion. As shown in Figure 3(F) and (G), oeRBM25 transfection significantly depressed HTR-8/SVneo cells invasion from the Matrigel droplet. Together, these data establish a function for RBM25 in regulating trophoblast cells migration and invasion.
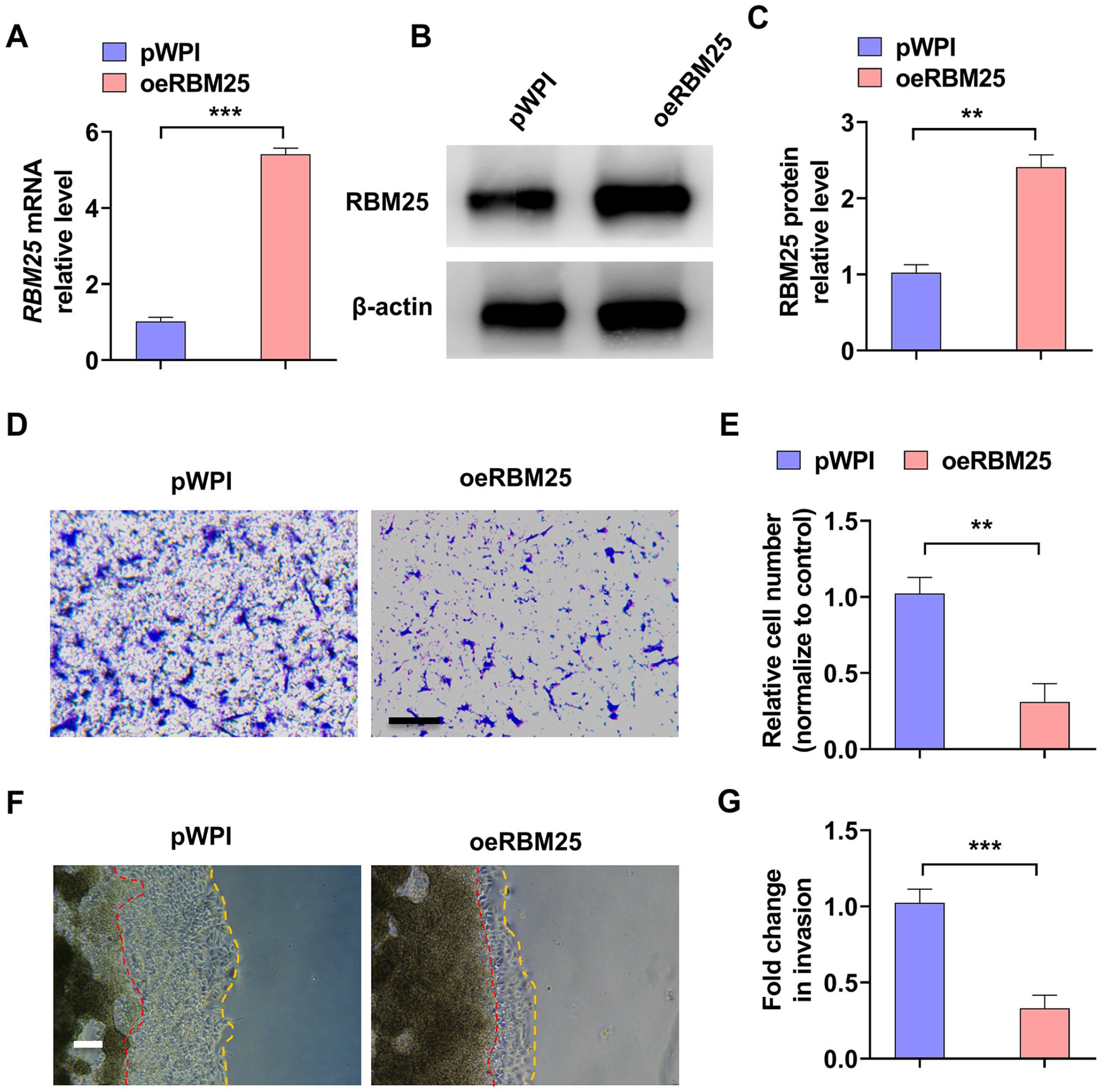
Over-expression of RBM25 inhibited trophoblast cells migration and invasion. (A) HTR-8/SVneo cells were transfected with RBM25-over-expressed vector (oeRBM25) or control vector (pWPI), and then RT-qPCR was used to measure RBM25 mRNA level. (B) Western blot was used to examine RBM25 protein level in oeRBM25 or pWPI vector transfected-HTR-8/SVneo cells. (C) Quantitative analysis of Western blot results in (B). (D) HTR-8/SVneo cells were treated as in (A), and then a transwell assay was used to examine cells migration. Scar bar = 50 μm. (E) Quantitative analysis of relative cell number of migration in (D). (F) The 3D Matrigel droplet assay was used to detect the invasion of HTR-8/SVneo cells after oeRBM25 or pWPI vector transfection. Scar bar = 100 μm. (G) Quantitative analysis of fold change of cell invasion in (F). All data were collected from three independent experiments and were presented as a mean ± standard error.
RBM25 negatively regulates EMT of trophoblast cells
Previous studies have shown that cellular EMT is a prerequisite for cell migration and invasion. 14 To investigate whether RBM25 promotes cell migration and invasion by regulating cellular EMT, we first examined the expression of EMT-related genes. As shown in Figure 4(A) and (B), knocking down RBM25 obviously reduced the epithelial cell marker E-cadherin expression while increased mesenchymal cell marker Vimentin protein level. Conversely, over-expression of RBM25 promoted E-cadherin while inhibited Vimentin proteins level (Figure 4(C) and (D)). In addition, the results of double immunofluorescence staining also confirmed that knocking down RBM25 significantly promoted the expression of Vimentin in the cytoplasm and inhibited the expression of E-cadherin at the intercellular junction. However, over-expression of RBM25 obtained the opposite results (Figure 4(E) and (F)). Subsequently, we detected these EMT relative genes expression in the clinical samples using immunohistochemical staining. As shown in Figure 4(G) and (F), the epithelial cell marker E-cadherin was significantly upregulated in PE tissues compared with normal placental tissues, while mesenchymal cell marker Vimentin was downregulated in PE tissues. These results suggest that RBM25 inhibits the process of EMT in trophoblast cells by regulating EMT marker gene expression.
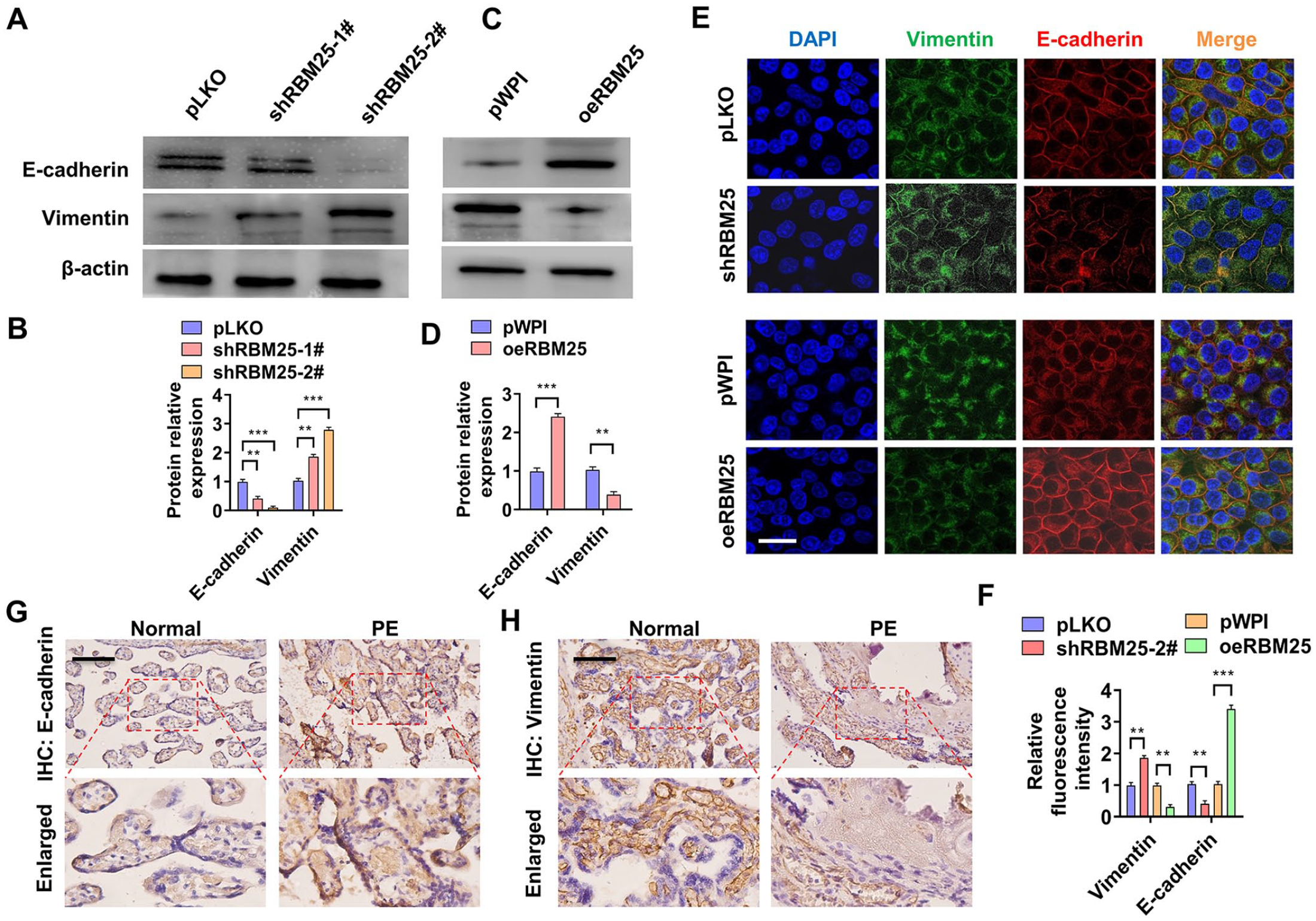
RBM25 is involved in EMT of trophoblast cells by regulating EMT marker gene expression. (A) HTR-8/SVneo cells were transfected with shRBM25-1# or shRBM25-2# or pLKO and then Western blotting was used to detect E-cadherin and Vimentin protein expression. (B) Quantitative analysis of Western blot results in (A). (C) Western blotting was used to examine E-cadherin and Vimentin protein expression in oeRBM25 or pWPI-transfected HTR-8/SVneo cells. (D) Quantitative analysis of Western blot results in (C). (E) HTR-8/SVneo cells were transfected with shRBM25-2# or oeRBM25 or their corresponding control vectors, and then double immunofluorescence staining was used to explore E-cadherin and Vimentin protein expression. DAPI (blue), Vimentin (green), and E-cadherin (red). Scar bar = 25 μm. (F) Quantitative analysis of relative fluorescence intensity in (E). (G and H) Immunohistochemical staining detects E-cadherin or Vimentin expression in normal (n = 30) and PE (n = 30) placental tissues. All data were collected from three independent experiments and were presented as a mean ± standard error.
RBM25 promotes Grhl2 expression by stabilizing mRNA
Emerging studies have shown that Grhl2 plays a vital role in the EMT process.16,25 To study whether Grhl2 is involved in RBM25-regulated cells migration and invasion, HTR-8/SVneo cells were over-expressed or knocked down with RBM25 and then detected Grhl2 and ZEB1 expression. As shown in Figure 5(A) to (C), over-expression of RBM25 significantly increased Grhl2, not ZEB1, mRNA, and protein level, while depletion of RBM25 reduced Grhl2 expression. As an RBP, not a transcription factor, we wanted to investigate how RBM25 promotes the mRNA and the protein expression of RBM25. HTR-8/SVneo cells were transfected with oeRBM25 or shRBM25 or their control vectors, and then cells were treated with actinomycin D to inhibit gene transcription. Next, we used RT-qPCR to detect the maintenance of Grhl2 mRNA at different processing times. As shown in Figure 5(D), over-expression of RBM25 promoted and knockdown of RBM25 inhibited Grhl2 mRNA maintenance. To detect whether RBM25 promotes Grhl2 mRNA stabilization by interacting with mRNA, HTR-8/SVneo cells were transfected with oeRBM25 or control vector (pWPI) and then a CoIP was used to confirm pulldown effect by RBM25. The results showed that over-expression of RBM25 markedly facilitated RBM25 precipitation (Figure 5(E)). Subsequently, a RIP assay was used to explore RBM25 and Grhl2 mRNA interaction. As shown in Figure 5(F), the RBM25, not RBM5, antibody significantly enriched the Grhl2 mRNA. Conversely, we used the mRNA probes to pulldown RBM25 and the results revealed that Grhl2, not ZEB1, mRNA probes efficiently pulled down RBM25 protein molecules (Figure 5(G)).
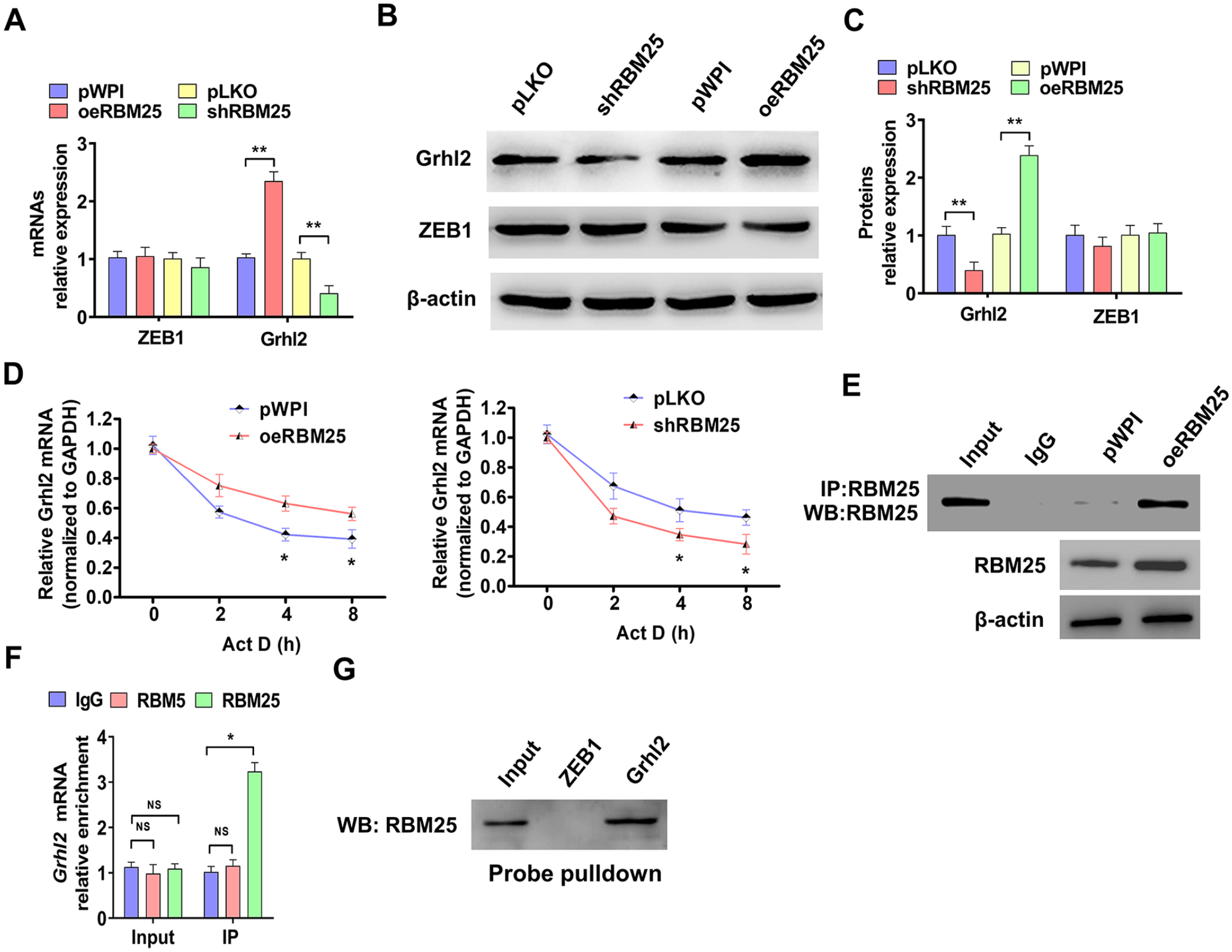
RBM25 promotes Grhl2 expression by stabilizing mRNA. (A) HTR-8/SVneo cells were transfected with oeRBM25 or shRBM25 or their control vectors, and then RT-qPCR was used to detect ZEB1 and Grhl2 mRNAs expression. (B) HTR-8/SVneo cells were transfected as in (A), and Western blot was performed to measure Grhl2 and ZEB1 protein level. (C) Quantitative analysis of Western blot results in (B). (D) HTR-8/SVneo cells were transfected as in (A) and treated with actinomycin D (Act D) for 0, 2, 4 and 8 h. RT-qPCR was used to examine Grhl2 mRNA. (E) HTR-8/SVneo cells were transfected with oeRBM25 or pWPI vector, and the CoIP-Western blot was used to confirm RBM25 protein precipitation by a RBM25 antibody. (F) RNA binding protein immunoprecipitation assay (RIP) was used to examine proteins and Grhl2 mRNA interaction. RT-qPCR was used to detect the Grhl2 mRNA enrichment by using RBM25, RBM5, and IgG antibodies. (G) ZEB1 and Grhl2 mRNA probes were designed and synthesized, then a Pulldown-Western blot assay was used to explore RNA interaction with RBM25. All data were collected from three independent experiments and were presented as a mean ± standard error.
Grhl2 is involved in RBM25-regulated trophoblast cells migration, invasion and EMT
In order to explore whether Grhl2 is involved in the EMT of trophoblast cells regulated by RBM25, we performed rescue experiments. HTR-8/SVneo cells were transfected with shRBM25 or oeGrhl2 alone or both together, and Western blotting analysis was used to examine EMT marker genes expression. As shown in Figure 6(A) and (B), transfection of oeGrhl2 significantly promoted E-cadherin while reduced Vimentin expression. However, co-transfection of shRBM25 partially counteracts the regulatory effect on the EMT marker gene by over-expression of Grhl2. In contrast, over-expression of RBM25 increased the expression of E-cadherin and decreased the expression of Vimentin, while depletion of Grhl2 together reversed the expression of these proteins regulated by over-expression of RBM25 (Supplementary Figure 2). Subsequently, we examined the effects of Grhl2 in RBM25-regulated EMT by double immunofluorescence staining. The results confirmed that over-expression of Grhl2 significantly promoted the expression of E-cadherin at the intercellular junction while inhibited the expression of Vimentin in the cytoplasm. However, knockdown of RBM25 partially reversed the expression of these genes regulated by over-expression of Grhl2 (Figure 6(C) and (D)). Subsequent functional analysis showed that knockdown of RBM25 significantly promoted cell migration. However, the simultaneous co-transfection of oeGrhl2 counteracted the cell migration effect promoted by knockdown of RBM25 (Figure 6(E) and (F)). Consistent with this, the same results were obtained by 3D Matrigel droplet assay (Figure 6(G) and (H)). Collectively, these data establish that Grhl2 is involved in the RBM25 regulation of trophoblast migration, invasion, and EMT.
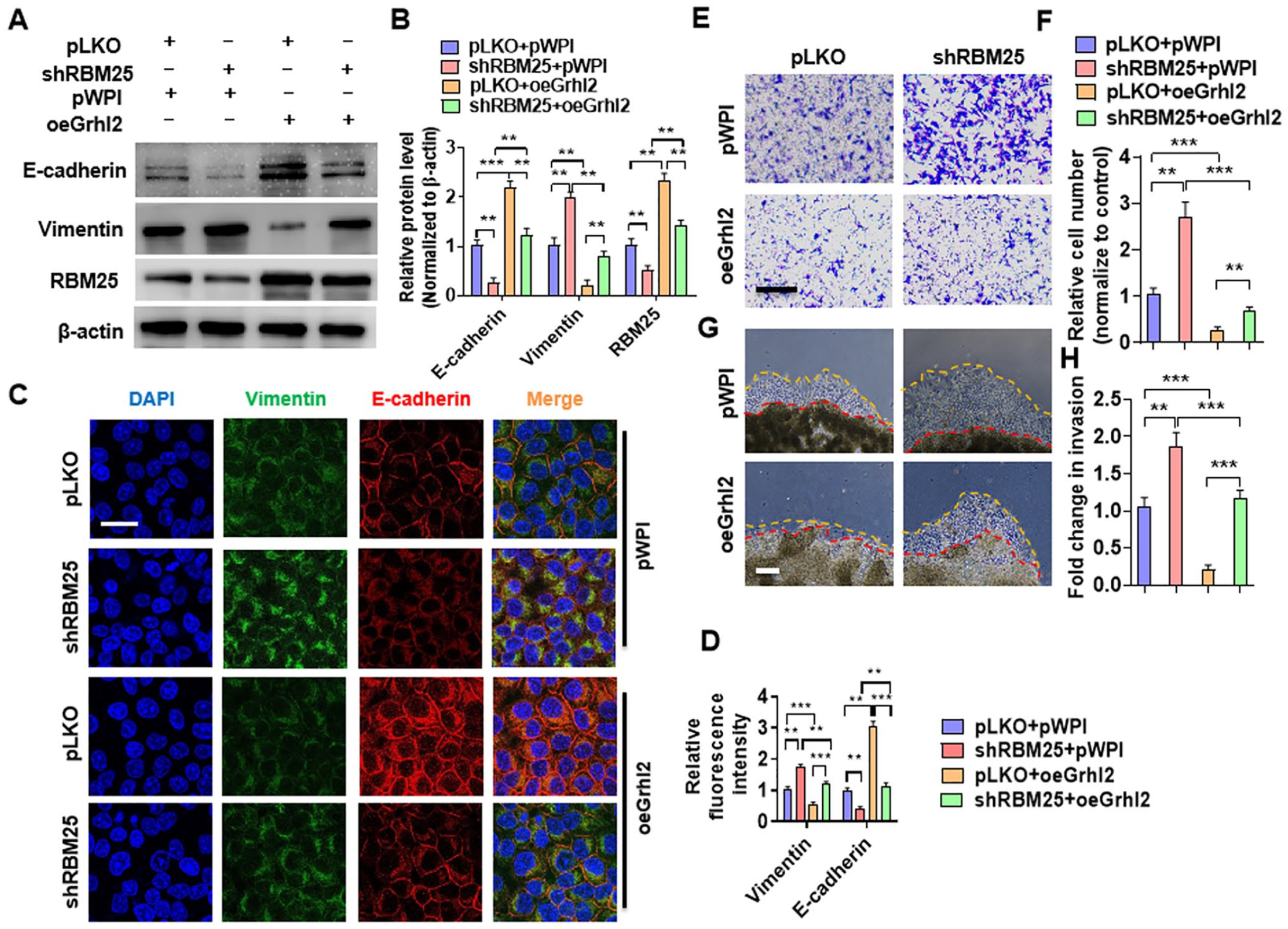
Grhl2 is involved in RBM25-regulated trophoblast cells migration, invasion, and EMT. (A) HTR-8/SVneo cells were transfected with shRBM25 or oeGrhl2 alone or together, and then Western blotting was used to measure E-cadherin, Vimentin, and RBM25 expression. (B) Quantitative analysis of Western blot results in (A). (C) HTR-8/SVneo cells were transfected as in (A), and double immunofluorescence staining was used to explore E-cadherin and Vimentin protein expression. DAPI (blue), Vimentin (green), and E-cadherin (red). Scar bar = 25 μm. (D) Quantitative analysis of relative fluorescence intensity in (C). (E) Transwell assay was used to explore cell migration in HTR-8/SVneo cells after transfection with shRBM25 or oeGrhl2 alone or together. Scar bar = 50 μm. (F) Quantitative analysis of relative cell number of migration in (D). (G) The 3D Matrigel droplet assay was used to explore the invasion of HTR-8/SVneo cells after transfected as in (A). Scar bar = 100 μm. (H) Quantitative analysis of fold change of cell invasion in (G). All data were collected from three independent experiments and were presented as a mean ± standard error.
Grhl2 and RBM25 form a feedback loop to regulate the progress of PE
Previous results showed that over-expression of Grhl2 elevated RBM25 protein level (Figure 6(A)). Intriguingly, we wondered if Grhl2, as a transcription factor, could in turn promote transcription of RBM25. Then, HTR-8/SVneo cells were transfected with oeGrhl2 or shGrhl2 or their control vectors, RT-qPCR was used to examine RBM25 expression. As shown in Figure 7(A), over-expression of Grhl2 promoted while knockdown of Grhl2 inhibited RBM25, not ZEB1, mRNA expression. Subsequently, HTR-8/SVneo cells were transfected with oeGrhl2 and simultaneously treated with Act D, and then Western blot was used to analyze RBM25 expression. The results showed that over-expression of Grhl2 elevated RBM25, not ZEB1, protein level. However, simultaneous treatment with Act D eliminated the promoting effect of Grhl2 on RBM25 expression (Figure 7(B) and (C)). To identify whether Grhl2 promotes RBM25 transcription, we analyzed the Grhl2 binding sites in promoter of RBM25 by JASPAR website (https://jaspar.genereg.net/). 23 And we found that there are two potential bound motifs in this region (Figure 7(D)). Next, a ChIP-PCR was used to confirm that Grhl2 bound to the RBM25 promoter (–993~–1021 nt) (Figure 7(E)). Furthermore, a luciferase assay revealed that over-expression of RBM25 itself increased the luciferase activity of RBM25 promoter, and knockdown of Grhl2 simultaneously offset this effect (Figure 7(F)). Taken together, our combined results showed that Grhl2 promotes RBM25 transcription and forms a positive feedback regulation mode in PE progression.
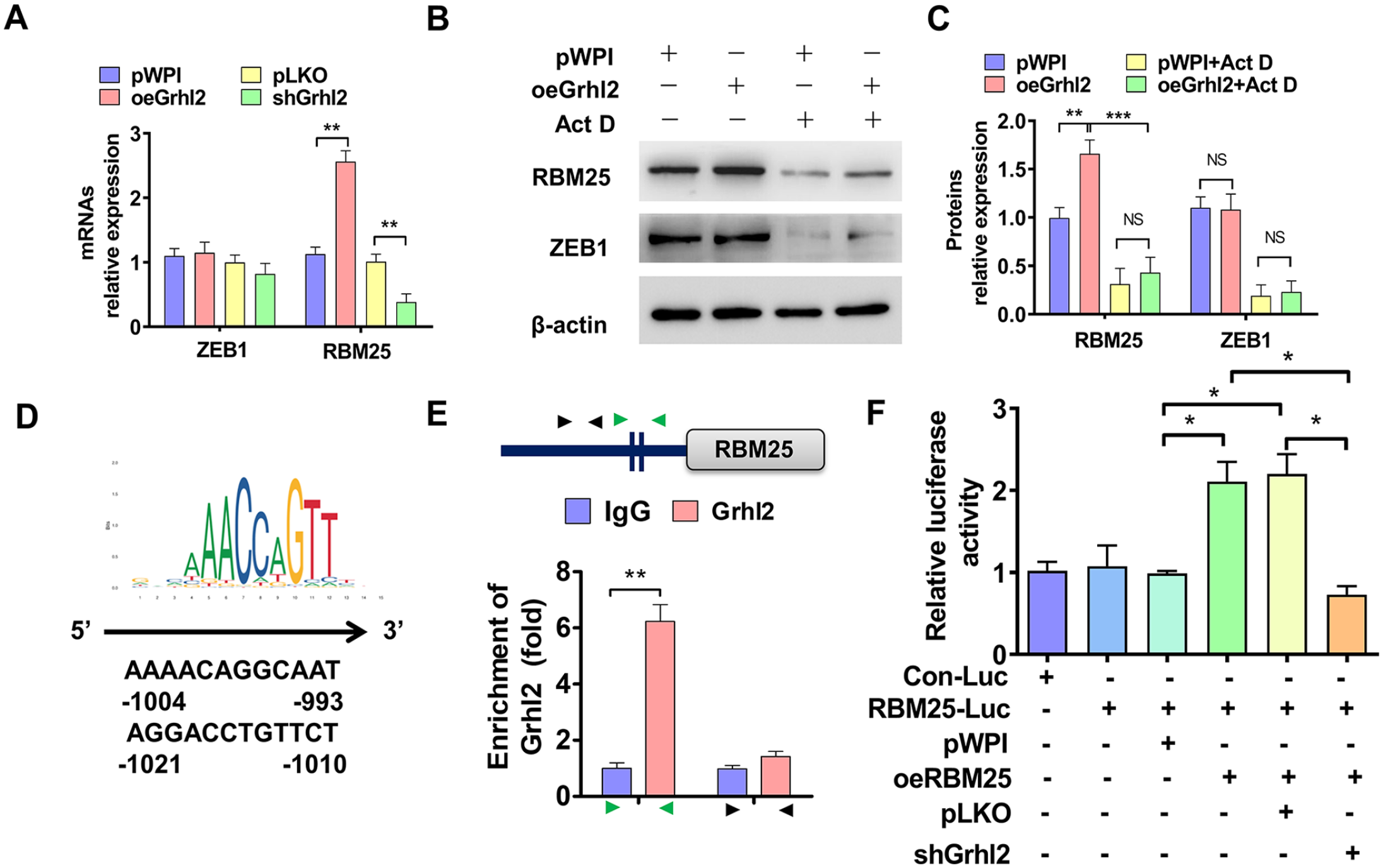
Grhl2 promotes RBM25 expression by transcription. (A) HTR-8/SVneo cells were transfected with oeGrhl2 or shGrhl2 or their control vectors, and then RT-qPCR was used to detect ZEB1 and RBM25 mRNAs expression. (B) HTR-8/SVneo cells were transfected with oeGrhl2 or pwpI, and then treated with Act D. Western blot was used to examine RBM25 and ZEB1 expression. (C) Quantitative analysis of Western blot results in (B). (D) The potential Grhl2-binding motif on the promoter of RBM25 gene. (E) ChIP-PCR was used to identify the binding site of Grhl2 on the RBM25 promoter. (F) Dual luciferase reporter gene examined the role of Grhl2 and RBM25 in regulating RBM25 promoter activity by co-transfection with indicated constructs. All data were collected from three independent experiments and were presented as a mean ± standard error.
Discussion
It is generally accepted that superficial and limited invasion of the spiral artery of the uterus by trophoblasts is a key link in the onset of PE. 26 In the early stages of blastocyst implantation, ectoderm cells differentiate into cytotrophoblast cells and syncytiotrophoblast cells. 27 The former can differentiate into more invasive EVT cells, some of which invade the deeper layers of the endometrium up to the inner third of the myometrium; The other part invades and reshapes the spiral artery of the uterus to provide adequate blood flow to the fetal placenta. 9 When trophoblast invasion is impaired, the spiral artery invades too superficially, and placental blood flow is reduced. These changes of trophoblast cells not only lead to intrauterine growth restriction of the fetus, but also secrete some cytokines or hormones, which trigger systemic vascular endothelial dysfunction and inflammatory response, leading to the occurrence of PE. 5 The invasion process of trophoblasts is similar to that of tumor cells, except that the invasion of trophoblasts is finely regulated, 28 but the exact molecular mechanism is not fully understood. Therefore, the study of the defect of the invasion ability of trophoblast cells may become one of the effective ways to solve PE.
Epithelial-mesenchymal transition (EMT) is a dynamic and reversible process in which epithelial cells acquire mesenchymal cell characteristics. 29 After a series of gene expression regulation, the highly expressed cell junction proteins such as E-cadherin and ZO-1 in epithelial cells were inhibited, and then the interstitial cell marker proteins such as Vimentin and N-cadherin were highly expressed. Epithelial cells lost their typical features and gained the ability to migrate and invade surrounding tissues. 30 Grhl2 plays an important role in physiological processes such as epidermal integrity, embryonic neural tube closure, and wound healing. 31 Emerging studies have shown that Grhl2 is a negative regulator of cellular EMT process. 32 Yang et al. 25 reported that Grhl2 depresses the progression and metastasis of colorectal cancer by inhibiting epithelial-mesenchymal transition. Grhl2 inhibits bladder cancer cell proliferation and inhibits EMT process by downregulating ZEB1 expression. 32 Our group previous study revealed that Grhl2 was highly expressed in PE placenta, and circTNRC18-induced miR-762 inhibited upregulation of Grhl2, leading to trophoblast migration and EMT. 16 In this study, we found that Grhl2 was involved in RBM25-regulated trophoblast cells migration, invasion, and EMT. In addition, Grhl2 promotes RBM25 expression through transcription, and positive feedback regulation between them promotes the occurrence and development of PE. However, how Grhl2 regulates cellular EMT during PE remains to be further studied. Previous studies have reported that Grhl2 is involved in the EMT process of tumor cells by binding to ZEB1 promoter to inhibit its transcription. 32 However, this study also found that ZEB1 was not involved in Grhl2-/RBM25-regulated cell migration and EMT. Next, we will further investigate how Grhl2 regulates the EMT of trophoblasts during PE.
It is widely believed that RBM25 is an RBP with splicing factor function. Using proteomic analysis, Carlson et al. 33 found that RBM25 interacts with components of early spliceosomes and regulators of alternative splicing. Transcriptome scoping analysis of splicing events confirmed that RBM25 regulates a large number of alternate splicing exons throughout the human genome 33 Hypoxia induced the increased binding of RBM25 and SCN5A mRNA, and promoted the decrease of Na (+) current by upregulating the abundance of SCN5A splicing variants and reducing the full-length SCN5A protein. 34 But as an RBP, RBM25 can do much more than that. Recent studies have shown that YY1-dependent activities, including chromatin binding, DNA loop, and transcription, were blocked after RBM25 was knocked out in cells. Furthermore, it is proposed that RBP, such as RBM25, may regulate RNA to control transcription and enhance molecular network interaction. 35 In addition, RBM25 directly binds to circRNA and induces circAMOTL1L biogenesis, thereby regulating cell EMT and migration. 14 In this study, we found that the expression of RBM25 was upregulated in placental tissue of PE. Upregulation of RBM25 inhibits trophoblast cell migration and invasion by regulating the expression of EMT marker genes. On further exploration, we found for the first time that RBM25 promoted the expression of EMT-related gene Grhl2 by stabilizing mRNA. However, it is not clear how RBM25 regulates mRNA stabilization. Some studies have reported that RBM25 can bind to the CGGGCA sequence of pre-RNA to regulate splicing. 12 We suspect that RBM25 may directly bind to mature mRNA, extending the half-life of RNA. Next, we will further investigate how RBM25, as an RBP, promotes mRNA stabilization.
In conclusion, this study revealed that the expression of RBM25 in PE placental tissue was significantly increased. Knockdown of RBM25 promotes and over-expression of RBM25 inhibits trophoblast migration and invasion. Further studies showed that RBM25 inhibited the EMT process by regulating the EMT marker gene. Our results reveal for the first time that Grhl2 is involved in RBM25-regulated trophoblast migration, invasion, and EMT through RBM25-promoted mRNA stabilization. As a transcription factor, Grhl2 promotes the expression of RBM25 through transcription, and the positive feedback regulation between the two molecules promotes the occurrence and development of PE.
Supplemental Material
sj-pdf-1-ebm-10.1177_15353702231191199 – Supplemental material for RBM25 induces trophoblast epithelial-mesenchymal transition and preeclampsia disorder by enhancing the positive feedback loop between Grhl2 and RBM25
Supplemental material, sj-pdf-1-ebm-10.1177_15353702231191199 for RBM25 induces trophoblast epithelial-mesenchymal transition and preeclampsia disorder by enhancing the positive feedback loop between Grhl2 and RBM25 by Jing Zhang, Wen-qi Chen, Kai Yang, Zhao-xi Wang, Dong-lan Sun, Yuan-yuan Peng, Mei Yu, Shao-xiong Wang and Qing Guo in Experimental Biology and Medicine
Footnotes
Authors’ contributions
JZ contributed to conceptualization and project administration; WQC contributed to data curation and writing—original draft; KY contributed to writing—review & editing; ZXW contributed to data curation; DLS contributed to formal analysis; YYP contributed to software; MY contributed to validation; SXW contributed to visualization; and QG contributed to methodology, supervision, and funding acquisition.
Data availability statement
The data that support the findings of this study are uploaded to figshare repository. The links of the data in figshare repository is 10.6084/m9.figshare.21993035.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Hebei Natural Science Foundation Precision Medicine Joint Fund Cultivation Project (No. H2021106001).
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.