
Abstract
Arboviral diseases comprise a group of important infectious diseases imposing a heavy burden to public health in many locations of the world. Orthoflaviviruses are viruses belonging to the genus Orthoflavivirus; this genus includes some of the most relevant arboviruses to human health. Orthoflaviviruses can infect several different hosts, with some species being transmitted in cycles involving birds and anthropophilic mosquitoes and others transmitted between mammals and mostly Aedes sp. mosquitoes. Some of the most important sylvatic reservoirs of orthoflaviviruses are non-human primates (NHPs). Many flaviviruses that infect NHPs in nature have the potential to cause epidemics in humans, as has been observed in the cases of Orthoflavivirus denguei (dengue virus – DENV), Orthoflavivirus flavi (yellow fever virus – YFV), and Orthoflavivirus zikaense (Zika virus – ZIKV). In this minireview, we discuss important aspects regarding history, ecology involving NHP, distribution, disease outcome, and pathogenesis of these three major orthoflaviviruses that affect humans and NHP and relate this information to the potential of using NHP as experimental models. In addition, we suggest some orthoflaviviruses that could be better investigated, both in nature and in experimental studies, in light of the recent revolution in molecular biology.
Impact Statement
Arboviral diseases are among the major infectious diseases with potential to emerge and spread to novel locations in the future. The Orthoflavivirus genus comprises some of the most impacting viruses to human and animal health worldwide, and primates are susceptible to some of the most important orthoflaviviruses causing human diseases, such as dengue virus (DENV), yellow fever virus (YFV), and Zika virus (ZIKV). In this minireview, we included the ecological, phylogenetic, and pathogenic aspects regarding these viruses and non-human primates (NHPs) focusing in neotropical primates’ susceptibility and potential to be used as animal models. This article summarizes and highlights the most relevant fields of active research to advance in Orthoflavivirus ecology and pathogenesis involving neotropical primates.
Introduction
Diseases caused by arthropod-borne viruses represent a 17% of all infectious diseases in the World and, as are most emerging infectious diseases, arbovirus diseases are zoonotic.1–3 Most of the zoonotic emerging infectious diseases are acquired from mammals. Urbanization and agricultural expansion in regions where there are potential host populations can cause spillover events. 2
The genus Orthoflavivirus comprises approximately 70 viral species with transmission cycles involving both vertebrates and invertebrates that cause neurotropic or multisystemic diseases in vertebrate hosts. Several orthoflaviviruses that can infect humans originated in sylvatic environments, being transmitted between vertebrate hosts, such as NHP and arthropods. Repeated spillover events in interface areas led to sporadic human infections, and possibly during these events, some of these viruses became adapted to anthropophilic mosquitoes and humans, sustaining endemic cycles in small communities. Ultimately, some of these viruses adapted perfectly to urban environments, such DENV in tropical regions of the world, while others caused important pandemic events with catastrophic consequences, such as ZIKV in the Americas 4 and others are responsible for several human deaths and diseases in areas that were sylvatic cycles still occur and/or vaccination is challenging, as is observed with YFV.
Orthoflaviviruses have a single-stranded, positive-sense RNA genome with approximately 11 kb in an enveloped icosahedral viral particle of approximately 50 nm. 5 The genus Orthoflavivirus can be divided according to the hosts involved in each virus transmission cycle, with species that are transmitted between vertebrate hosts by Culex sp., or Aedes sp. mosquitoes, species that are transmitted by ticks, species of viruses that are insect-specific and species of viruses with unknown invertebrate vectors. 6 The phylogeny of the members of the Orthoflavivirus genus reflects their invertebrate vector’s ecological niches. Culex-transmitted orthoflaviviruses evolved from Aedes-transmitted orthoflaviviruses and more often circulate among birds. Still, some mammals can act as amplifiers or reservoirs for some of these viruses, since Culex mosquitoes tend to feed in a broader range of hosts. Orthoflaviviruses transmitted by Aedes mosquitoes are maintained in nature in cycles involving mostly primates or other mammalian species as reservoirs, depending on the feeding behavior and vertebrate host availability. Birds can be infected by some of these viruses and serve as dead-end hosts.6,7
Amino acid sequence analysis of non-structural protein 5 (NS5), envelope, and polyprotein from orthoflaviviruses, based on Bayesian inference under a relaxed molecular clock model, demonstrates that these viruses can be separated into clades according to the genus of their invertebrate vector. The use of amino acid sequence analysis allows comparison of phenotype similarities regardless of codon usage bias and avoids potentially saturated nucleotides within a genus. The inference used in the study considers different mutation rates along the genome, and is, therefore, robust, as arbovirus genomes usually present variable mutation rates along the genome as a result from the strong pressures imposed by the interaction with immune systems from completely different hosts. 8 In orthoflaviviruses, RNA sequences from the untranslated regions (UTR) form secondary structures that are important for genome replication, therefore, tend to be well conserved. This mosquito vector-based division explains part of the evolutionary history of some orthoflaviviruses; however, ZIKV has also recombinant regions derived from interaction with culex-borne flaviviruses within the vertebrate host and its immune system, during co-infections that strengthen their phylogenetic relationships with culex-borne orthoflaviviruses. 9
Different methods of direct and serological detection that indicate previous infection have been employed in several studies involving wildlife,10–24 as summarized in Table 1. Orthoflaviviruses involved in past and present human diseases have been detected in several animal species. Of these, mammals from the order primates in Africa, Asia, and South and Central Americas have been shown to harbor evidence of flavivirus infection and, therefore, deserve attention in efforts to investigate these viruses.11–14,16–18,22,24
Studies performed after 1990 including NHP and other animal species detecting evidence of contact between animals and orthoflaviviruses of human health relevance. Primate species are highlighted in bold.
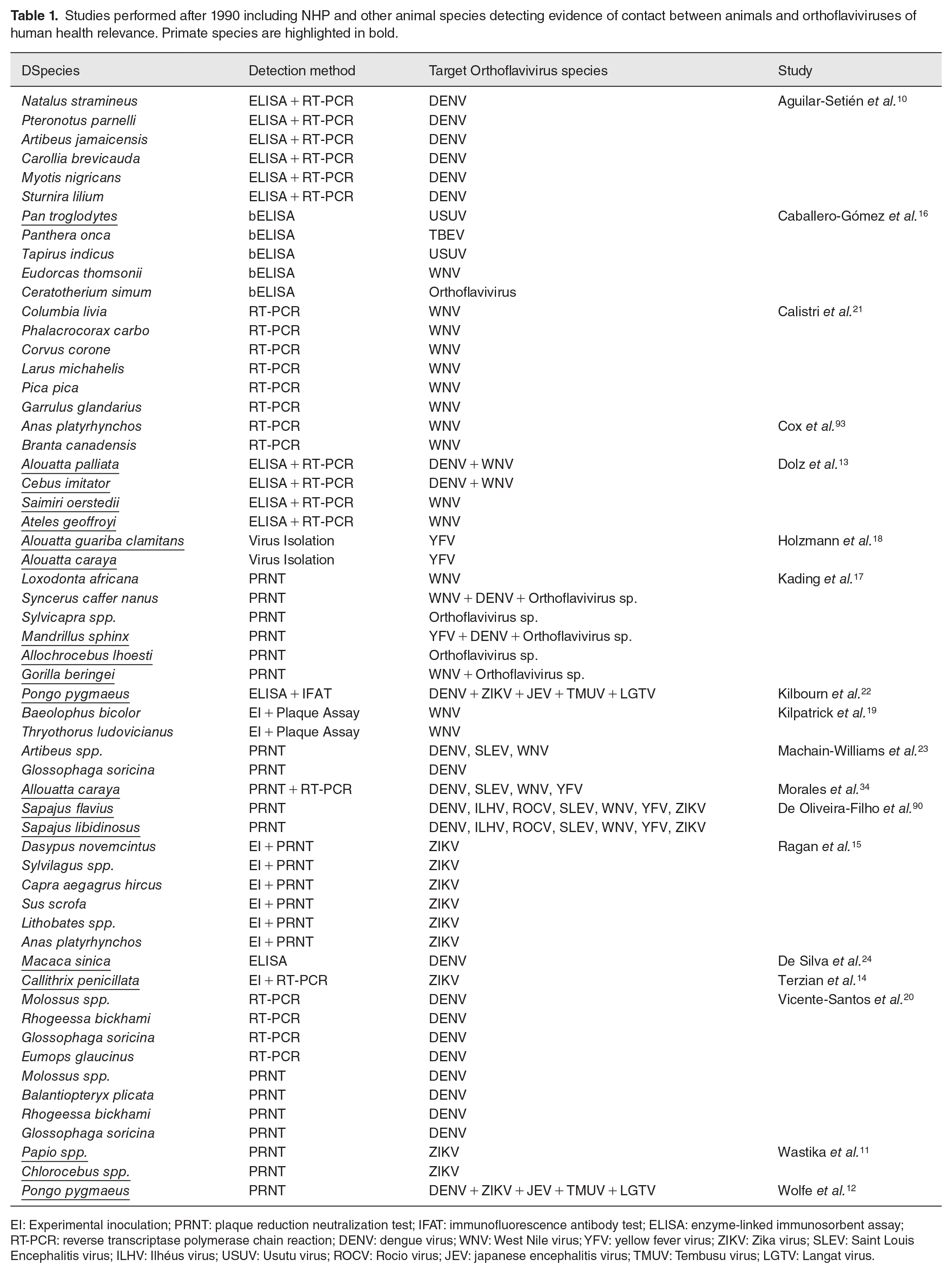
EI: Experimental inoculation; PRNT: plaque reduction neutralization test; IFAT: immunofluorescence antibody test; ELISA: enzyme-linked immunosorbent assay; RT-PCR: reverse transcriptase polymerase chain reaction; DENV: dengue virus; WNV: West Nile virus; YFV: yellow fever virus; ZIKV: Zika virus; SLEV: Saint Louis Encephalitis virus; ILHV: Ilhéus virus; USUV: Usutu virus; ROCV: Rocio virus; JEV: japanese encephalitis virus; TMUV: Tembusu virus; LGTV: Langat virus.
NHPs host orthoflaviviruses causing human health emergencies such as DENV, YFV, and ZIKV and other viruses with a high potential to cause global emergencies in human health. 25 This is expected, since many viruses co-evolve with hosts and throughout this process, many host factors associated with viral pathogenesis may be similar among members of taxonomically close taxa. The history of primate evolution involves spread to several continents, ultimately giving rise to several novel primate species belonging to the clade Platyrrhini in the Americas.26,27 The taxonomic proximity between NHP from Africa, which is the probable place where the genus Orthoflavivirus arose, 28 and NHP from America makes NHPs from the New World obvious candidates to act as new hosts for orthoflavivirus infections. In this minireview, we will focus on the sylvatic cycle of DENV, YFV, and ZIKV involving NHP, including biogeography, phylogeny, clinical and pathological findings, and NHP potential to be used in research for the most relevant orthoflaviviruses that are transmitted to and from these primates.
DENV
Dengue is the arboviral disease that causes the greatest number of cases and deaths worldwide. It is considered hyperendemic in tropical regions and is fully adapted to humans and anthropophilic mosquitoes from Aedes aegypti and Aedes albopictus species. 29 DENV is divided in four serotypes, all of which have already been detected in NHP. Many studies show evidence of DENV in sylvatic cycles involving NHP in Asia,28,30 and phylogenetic studies show that all four serotypes of DENV affecting humans evolved from serotype-specific ancestors in sylvatic cycles involving forest-dwelling mosquitoes and NHP. 31 Today, all DENV serotypes include genotypes that are highly adapted to humans.
The probable origin of urban DENV cycles is in some interface area between forests and human settlements in Southeast Asia. Malaysia harbors the higher diversity in sylvatic DENV variants. All four serotypes of DENV have been evidenced there in sylvatic cycles: specifically, serotypes 1, 2, and 4 were isolated from sentinel monkeys, and serotype 3 has been inferred from seroconversion of sentinel NHPs.32,33
There are some serological evidences implying that DENV can infect NHP from urban and wild places in several countries from Asia and Africa, and in the Americas, DENV from urban cycles has been evidenced in free-ranging NHPs.13,25,34,35 There has been serological evidence of orthoflavivirus infections in free-ranging NHP and Sloths from Brazil 36 and free-ranging NHPs from Argentina. 34 Nonetheless, many of these studies are based solely in the presence of antibodies that may lack specificity; hence, those findings could still be attributed to seroconversion from contact with other flaviviruses. In our laboratory, we have detected DENV-1 through virome analysis of mosquito abdominal samples from interface areas containing several NHP packs in Rio Grande do Sul, Brazil (unpublished data). Interestingly, the sequence diverged from DENV-1 sequences detected in an outbreak affecting humans in the same state. This finding is still being further analyzed; however, this highlights the importance of xenosurveillance in forest areas with NHP.
DENV from serotypes 1, 2, and 4 has been identified as belonging to variants involved in urban and in sylvatic cycles.37,38 Serotypes 1, 2, and 4 have been detected in sylvatic cycles involving Asian NHPs from Macaca and Presbytis genus and Aedes niveus mosquitoes.35,39,40 In Africa, sylvatic DENV-2 detection has been achieved in Erythrocebus patas monkeys and several arboreal Aedes sp. mosquitoes in Senegal. 41 Even though there is no sequence information about the sylvatic DENV-3 serotype, due to the history of DENV evolution it is likely that this serotype has variants adapted to sylvatic cycles as well. 37 Exposure to DENV has been detected in orangutans and toco monkeys from Southeast Asia.22,24 Large epizootics have been evidenced in the past involving DENV-2 and free-ranging primates, with sustained antibody titers to the virus and passive antibody transfer from previously infected mothers to their offspring. 24 Baboons (Papio anubis), African green monkeys (Chlorocebus aethiops sabaeus), rhesus (Macaca mulatta), and cynomolgus (Macaca fascicularis) experimentally infected with human epidemic lineages of DENV produce sustained viremia and mild clinical signs.42,43 Human adapted isolates of DENV-1, DENV-2, and DENV-4 produce longer viremia in NHPs when compared with DENV-3. 42 Sylvatic lineages of DENV-2 tend to produce shorter viremia, rashes, and elevation of AST levels in serum. 44 Callithrix penicillata NHPs subcutaneously infected with DENV-3 demonstrated early monocytosis, slight neutrophilia and lymphocytosis, persistent leucopenia starting four days pot infection, increased levels of alanine transaminase (ALT) and aspartate aminotransferase (AST), anemia, and thrombocytopenia. Viremia peaked in these marmosets between days 3 and 6 postinfection. 45 Lesions caused by DENV in NHPs vary, and some NHPs remain asymptomatic while in others, such as Macaca mulata, intravenous innoculation yields cutaneous rashes and hemorrhages. These hemorrhages range from mild petechiae to severe coagulopathy, resembling human dengue hemorrhagic syndrome. 46
Emergencies of DENV transmission have occurred in both ways: emerging from sylvatic environments to areas interfacing with human settlements and from urban regions to those interface areas. In both cases, there is much concern about the rise of mutations in the genome of Orthoflavivirus species adapted to sylvatic cycles and momentary disappearances of epidemic strains to sylvatic environments that can re-emerge in the future after herd immunity vanishes from human populations.37,38,47
Regarding DENV pathogenesis, there are many molecules involved in DENV entry into host cells through interaction with the envelope protein of the virus. 48 Heparan sulfate receptors are implicated in the pathogenesis of several orthoflaviviruses enhancing viral entry efficiency into host cells, 49 while C-type lectin receptors (CLR) such as mannose receptors Dendritic Cell-Specific Intercellular adhesion molecule-3-Grabbing Non-integrin (DC-SIGN), and its homolog, DC-SIGN-related (L-SIGN), and C-type lectin domain family 5 member A (CLE5A) have been associated with DENV entry and propagation under in vitro studies, with strong correlation between polymorphisms in DC-SIGN receptors and occurrence of dengue hemorrhagic syndrome with different patterns of inheritance in different regions of the world. 50 Moreover, laminin receptors are involved in the DENV-1 cell entry in the liver, 51 and phosphatidylserine receptors are able to interact with this component in the viral membrane and mediate internalization of DENV.48,52 Other molecules involved in DENV entrance in host cells include integrins, Claudin-1, and scavenger receptor class b type I. 48 Apes and macaques present different expression patterns during the development of immune responses to infectious pathogens; these differences tend to converge to a very similar response after 24 h. 53 However, DC-SIGN coding regions from NHPs are highly similar to those from humans and suffer from the same pressures of selection originating from host–pathogen interactions. 54 The taxonomic proximity of NHPs and humans is greater than that of any other animal models, and therefore, these animals are important to understanding the pathogenesis and immune responses to DENV infection. 55
Yellow fever
Yellow fever was the first formally identified and documented as a flaviviral disease affecting humans.56,57 The disease caused by YFV is a severe hepatic disease with potential to cause a multisystemic hemorrhagic disease with fatality rates between 30% and 50%. 58 The burden of this disease affects primarily countries in Latin America and Africa. 57 This disease occurs in sporadic outbreaks of epizootics with increased fatality in susceptible NHPs, followed by epidemics with cases of disease and disease in humans. 58 Sylvatic cycles occur in forests in Africa and South America involving forest-dwelling mosquitoes and NHPs. 59
YFV originated in East Africa is currently divided into three different lineages: one from South America, one from West Africa, and one from East Africa. 59 These lineages are divided into seven genotypes, five from Africa and two from South America. 57 The South American lineage derived from a common ancestor with the West African lineage during slave trade period. 60
YFV was transmitted in urban cycles until Aedes sp. mosquito eradication during the middle of the twentieth century in several countries effectively eliminated these urban cycles.59,61 With the development of the YFV 17D vaccine and elimination of the main urban invertebrate vector, the major concern regarding YFV is now the extent and the consequences of sylvatic cycles involving this virus.58,62,63 YFV is transmitted exclusively in sylvatic cycles in some locations of the world and in sylvatic and an intermediate cycle in rural areas in other locations, depending on the invertebrate vector competence of transmitting the virus and the susceptibility of the vertebrate hosts (Figure 1). Currently, YFV is transmitted among NHPs in sylvatic cycles involving forest-dwelling Aedes sp. mosquitoes in Africa and Haemagogus sp. and Sabethes sp. mosquitoes in South America, and spill over events result in epidemics that usually follow epizootics. 61 In addition, in Africa, some landscapes favor the occurrence of transmission in intermediate locations that lead to small-scale epidemics.58,61 In some situations, these can cause larger outbreaks, such as the epidemics that started in Angola and Democratic Republic of Congo in 2015, and spread to urban environments, raising concerns about the risk of transmission by Aedes aegypti mosquitoes. 62 Low vaccine coverage is implicated as one of the most important factors related to re-emergence of YFV in humans today,58,63 and this can lead to several consequences, including the emergence of novel strains of YFV and/or of the encounter of this virus with lineages of Aedes aegypti with vectorial competence to transmit YFV in urban cycles. 62
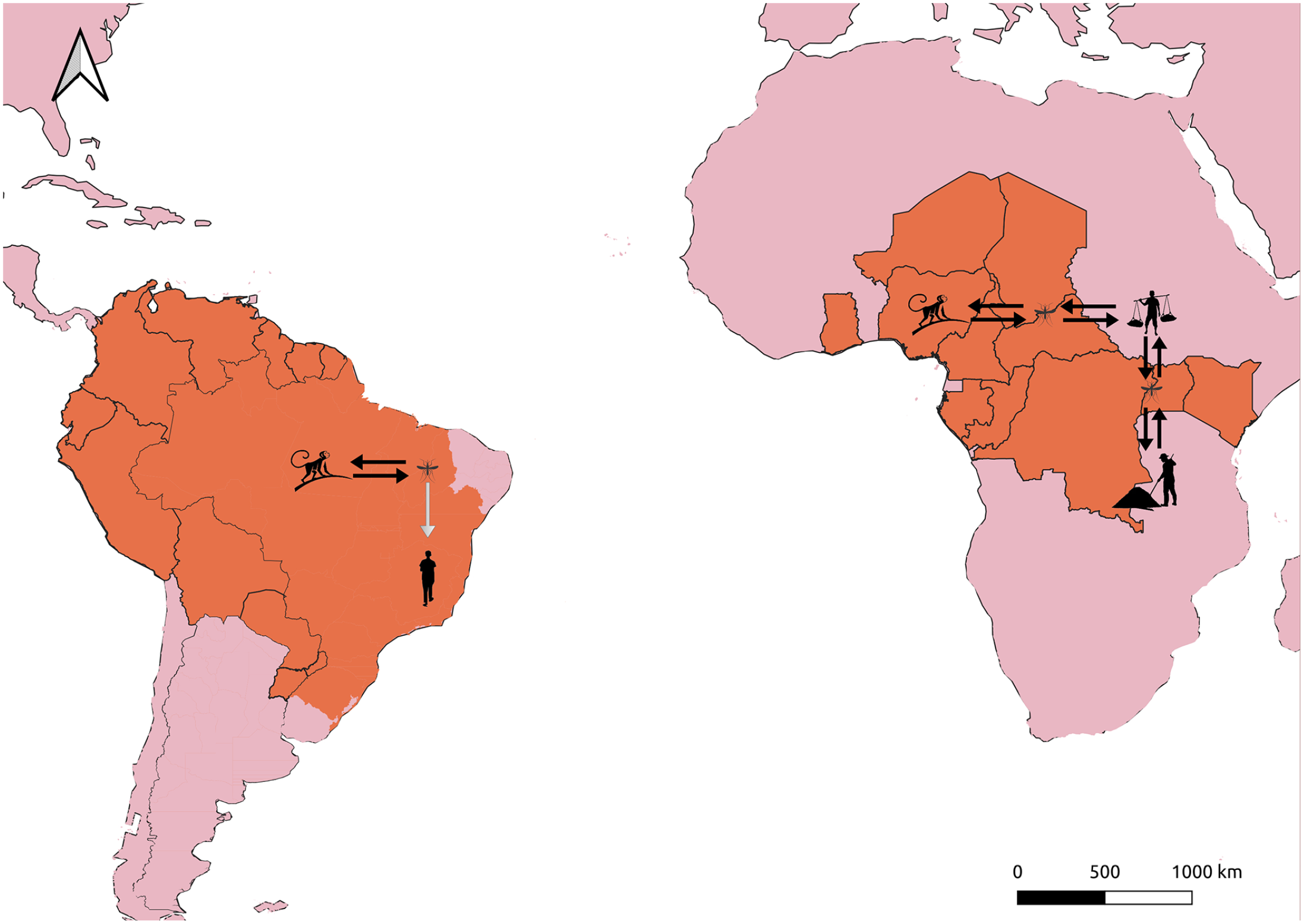
YFV patterns of transmission in Africa and in South America. Areas of YFV major concern are depicted in orange. While in South America there is currently only a sylvatic cycle involving NHPs and Haemagogus sp. and Sabethes sp. mosquitoes, in Africa there is a prevailing sylvatic cycle and emergencies that tend to spread to rural areas and YFV is sustainably transmitted by Aedes spp. mosquitoes between humans in these areas. Black arrow: sustained transmission; Gray arrow: spill over transmission.
YFV evolution has a slow rate when compared with other orthoflaviviruses, and variants are more consistently related geographically than by time scale. 61 The YFV strains that have been circulating in Brazil in the past two decades belong to the sublineage 1E, and the last epidemics that occurred in Brazil between 2016 and 2019 presented signature non-synonymous mutations that differentiate this strain from the previous prevailing strain related to the outbreak from 2008. 64 Still, these differences seem to have little impact on phenotypic features. 65 The most recent phylogenetic and biological data collected from YFV in Brazil were obtained from NHP samples. The surveillance of this virus in these animals is paramount to control and prevent YFV epidemics in humans. 66
The susceptibility to developing a severe disease after YFV infection varies among primate species. While some are highly susceptible to the virus, others display varying degrees of resistance. Among Old World primates, baboons (Papio spp.), colobus monkeys (Colobus spp.), green and vervet monkeys (Cercopithecus spp.), mangabeys (Cercocebus spp.), and chimpanzees (Pan troglodytes) produce viremia without overt clinical signs.25,67 Bush babies (Galago spp.), which are prosimians, are susceptible to YFV infection, developing fatal hepatitis. 67 In the Americas, some species of howler monkeys (Alouatta spp.), squirrel monkeys (Saimiri spp.), spider monkeys (Ateles spp.), and owl monkeys (Aotus spp.) have been found to be highly susceptible to develop yellow fever, while others such as marmosets (Callithrix spp.) and capuchin monkeys (Sapajus spp.) appear to be less affected by the virus.67,68
NHPs are susceptible to YFV infection, and severely affected neotropical primates develop marked hepatic disease with bridging to massive necrosis, hepatic lipidosis, Councilman bodies, and other lesions similar to those observed in human cases leading to severe hepatic failure and death in many cases, with Alouatta sp. individuals in particular are highly susceptible to this infection. 69 YFV has been detected in deceased NHPs from Rio Grande do Sul, the southernmost state of Brazil, with a temperate climate that prevents vectors to reproduce throughout the year, at least since 2002 70 and in more recent outbreaks as well, 65 mostly in Alouatta guariba individuals. We have recently detected YFV in 3 out of 30 NHP lungs from the most recent outbreak in southern Brazil, from samples of deceased Alouatta sp. during the years 2020 and 2021 submitted for severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) investigation (unpublished data). In some situations, NHP sampled for the official surveillance program in Brazil is in an advanced state of autolysis, and this can harm molecular tests, especially from tissues that tend to degrade quickly, such as the liver. This finding suggests that lungs provide an option to YFV detection in cases where liver is in severe autolysis. Nonetheless, crossed contamination of tissues cannot be ruled out.
Pathogenic YFV enters the host cells using clathrin-dependent endocytosis, while vaccine strains of YFV use dinamyn-2-dependent endocytosis. 49 The candidate receptors to YFV entry into host cells are phosphatidylserine receptors and heparan sulfate receptors. 48 The cytokine and chemokine profile released by macrophages and dendritic cells in infection with vaccine strains is different from that released in infections caused by wild-type strains of YFV. The immune response to YFV is similar between NHPs and humans: the release in NHP and human antigen-presenting cells is similar to pathogenic strains of YFV such as the Asibi strain. 71 Interestingly, even though NHPs are the best animal models for studying YFV, there is still a lack of information about YFV pathogenesis in NHP. The fact that some members of this taxon develop a disease similar to severe cases in humans could be better explored to understand pathogenetic mechanisms leading to severe forms of YFV in humans.
ZIKV
ZIKV was first isolated in 1947 in the Zika Forest of Uganda. Based on NS5 and envelope sequence analysis using Markov Chain Monte Carlo (MCMC) Bayesian approach with molecular clock, the lineage of ZIKV emerged in Uganda sometime between 1892 and 1943, probably around 1920. After its emergence, three distinct ZIKV lineages spread throughout Africa: the MR766 cluster, the Nigerian cluster, and a third lineage that was driven to Micronesia and Malaysia to the East, in the decade of 1960, to form the Asian Cluster. 72 ZIKV is highly adapted to sylvatic environments and is transmitted among NHPs, mostly monkeys, by forest-dwelling mosquitoes. 73 The discovery of ZIKV circulation in Asia occurred at the same time that studies of the virus were conducted in Africa, and the existence of a sylvatic cycle in Asia was suspected since seropositivity was detected in semi-captive orangutans in Malaysia. 74 African lineages of ZIKV were only isolated from mosquito pools and NHP samples, while naturally occurring ZIKV from the Asian lineage was mostly isolated from human samples and mosquitoes. 75 ZIKV emerged as a serious public health concern in 2007–2008 in Micronesia. It was the first time that ZIKV was associated with a major epidemic in humans. 76 More recently, this virus raised the concerns about its pathogenic potential, as it emerged as an important pathogen related to severe congenital malformations, first observed in northeastern Brazil.77,78
The phylogenetic proximity between humans and NHPs elevates the risk of spillover events of infectious diseases between these two taxa, and thus, forest regions of Brazil are among those with higher risk for these events. 2 Consequently, the concern about ZIKV infection and establishment in a sylvatic cycle involving free-ranging NHPs from a sylvatic cycle in the Americas is expected, and there are still questions regarding the range of susceptible animals living in forest–village interface areas of Brazil that could maintain a sylvatic ZIKV cycle, resembling YFV sylvatic cycle. 74
NHPs present varying levels of susceptibility to ZIKV infection. Certain primate species, such as rhesus macaques (Macaca mulatta) and marmosets (Callithrix spp.), can be experimentally infected with ZIKV and show clinical signs similar to those observed in humans.14,73 These signs include fever, rash, and neurological complications. 14 However, susceptibility to ZIKV can differ among primate species. Some primates, including certain species of baboons (Papio sp.) and African green monkeys (Chlorocebus sp.), have shown high seroprevalence of ZIKV antibodies in nature; however, no clinical signs have been observed in these species. 79
The recent ZIKV emergence in Yap Island causing Guillain–Barré Syndrome and in Brazil causing thousands of microcephaly cases have highlighted the variety of possible clinical and pathological outcomes caused by this virus.76,78,80 The clinical presentation of ZIKV infection varies, and can manifest as a Dengue-like disease, with mild fever and rash, a life-threatening Guillain–Barré, and also several congenital neurological malformations, including microcephaly.76,78 ZIKV strains from African and Asian/American lineages are able to reproduce microcephaly experimentally in rodents. 81
Pathological findings in human fetuses infected with ZIKV include thickening of leptomeninges, subcortical calcifications with cortical displacement, ventricular dearrangement, astrocytosis, macrophagic infiltrate, and microglial proliferation.78,82 ZIKV infection in mice causes apoptosis, lymphocytic infiltrate, and persistent infection in glomerular and tubular cells; moreover, ZIKV lineage MR 766 produces higher viremia and cytopathic effects in kidney cells than in the Asian lineage. 83
NHPs are important models of ZIKV infections, and the use of these animals in studies of ZIKV pathogenesis has provided important information, helping the scientific community to understand the virus after its emergence in 2015. Inoculation of Asian ZIKV in rhesus monkeys spreads to several tissues including kidney, bladder, uterus, with persistence in central nervous system (CNS), joints, and lymphnodes. Histologically, in animals experimentally infected with ZIKV, inflammatory infiltrates were observed in multiple tissues, and viral persistence in experimentally infected NHP was detected in liver, spleen, kidney, uterus, CNS, bladder, testes, heart, lymph nodes, joints, and bladder.83,84 ZIKV infection in pregnant NHP women resulted in several fetal alterations compatible with congenital Zika syndrome. 85 We have recently detected an African lineage-like ZIKV in free-ranging howler monkeys. All the outbreaks occurred in the summer/fall season, and two of these cases resulted in congenital Zika syndrome in fetuses. 86 Our findings imply that there must be adaptive restrains to the circulation of this strain in humans that limit its circulation to NHP species.
ZIKV has tropism to neuron progenitor cells, causing neurodegeneration; in addition, keratinocytes, skin fibroblasts, and skin dendritic cells are permissive to ZIKV infection. Furthermore, ZIKV is capable of entering fetal endothelial cells, a unique feature among flaviviruses.49,87 ZIKV also uses clathrin-mediated endocytosis, 88 and DC-SIGN, T-cell immunoglobulin and mucin domain 1 (TIM-1), and TAM (especially AXL) receptors are involved in ZIKV entrance into host cells.49,88 It has been shown that natural killer cells, monocytes, macrophages, and myeloid DC cell populations are activated after 5 dpi during ZIKV infection in NHPs and can remain activated until 85 dpi in some cases. Furthermore, there is important antibody development in infection of pregnant macaques. 85
Conclusions and perspectives
As orthoflaviviruses and primates evolved, they interacted on several occasions, originating many diseases that emerged in human populations and are today some of our major concerns in public health. The recognition of sylvatic cycles of orthoflaviviruses is a powerful strategy that can help prevent or accelerate responses to future threats related to these viruses. 89 Some of the main targets of these efforts should be investigation of forests with NHPs, given the taxonomic proximity and the history of infectious disease emergencies. 2
There is still a lack of information about the biology, pathogenesis, and ecology of many orthoflaviviruses that have been evidenced in free-ranging NHPs. 28 Active surveillance using non-invasive methods and xenosurveillance combined with novel virome discovery approaches should be employed in these environments to gather high-quality information about these pathogens. 89
Orthoflaviviruses from the species Spondweni, Uganda S, and Banzi are some examples of NHP-related orthoflaviviruses that require further study. 89 Moreover, orthoflaviviruses transmitted between birds and ornithophilic mosquitoes should be investigated in NHP, since seroprevalence has been evidenced in capuchin monkeys from northeastern Brazil to Saint Louis Encephalitis virus (SLEV) and Ilhéus virus (ILHV). 90 Furthermore, some studies conducted in Southern Brazil indicated seroprevalence to unknown orthoflaviviruses in passeriformes, raptors, and horses.91,92 Hence, strategies to sequence and reveal novel orthoflaviviruses should be implemented in NHP samples as well.
In addition to being important hosts of important orthoflaviviruses, NHPs are important animal models to help us understand the pathogenesis and outcomes of orthoflavivirus infections, particularly because many of them develop clinical disease and pathological findings similar to what is observed in our species. NHPs are essential to maintain balanced ecosystems around the world and are still the best alternative to help us understand the relationships between ourselves and these shared pathogens.
Footnotes
Authors’ Contributions
PRdA, MNW, LS, and FRS planned the conception of the review presented in the manuscript. PRdA wrote the initial text. MNW, LS, and FRS reviewed the text and added information to the final version of the text.
Declaration Of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: P.R.d.A. acknowledges funding from Coordenação de Aperfeiçoamento Pessoal de Nível Superior, the National Research Council, and Ministry of Health (Process number: 88887.131359/2016-00, MCTIC/FNDTC-CNPq/MEC-CAPES/MS-Decit/No 14/2016 – Prevenção e Combate ao vírus Zika). F.R.S. is a fellow of National Council for Scientific and Technological Development (CNPq) (grant number 302668/2018-7).