
Abstract
The structure-function gradient along the hippocampal long axis is recognized but poorly understood. Rodent recordings mainly sample the dorsal hippocampus (posterior), while surgical recordings in epilepsy patients typically sample from the ventral hippocampus (anterior), because of greater known involvement in seizure networks. These divergent recording practices imply that the mechanisms of rodent spatial navigation may not directly apply to human episodic memory. In general, the role of the posterior hippocampus in human memory and seizure networks is under-appreciated. We survey the basic, translational, and clinical research accounting for long-axis differentiation and outline future research directions.
Introduction
Because of rich, reciprocal connectivity with widespread cortical and subcortical regions, the hippocampus is readily recruited into pathological network dynamics, including seizures and interictal epileptiform discharges. Topographically organized cortical and subcortical inputs and outputs vary along the hippocampal long axis (LA), contributing to a graduated structure–function organization.1–4 Classic rodent lesion and physiology studies have demonstrated functional distinctions between dorsal (posterior/septal) and ventral (anterior/temporal) hippocampus. Differences in humans have also been observed. The translation from rodent to human functional neuroanatomy is non-trivialas evolution of the primate brain has involved expansion of the forebrain and temporal lobes and correspondingly, the anterior hippocampus. The posterior hippocampus has become involuted and diminutive.3–5
Differing recording practices between the rodent and primate hippocampus constrain cross-species translation. The dorsal (posterior) portion of the rodent hippocampus is largest and most surgically accessible, making it the most common target for experimental recordings. Conversely, due to the majority of focal epilepsies arising from the frontal (25%-30%) and temporal (50%) lobes, 6 surgical epilepsy recordings primarily sample from anterior hippocampus. Species-specific recording practices may contribute to inconsistent observations of theta presence and peak frequency across species, 7 especially in humans.8–11
The circuit architecture of the hippocampus varies across three principal axes: (1) transverse (ie, entorhinal–dentate–CA3–CA1/subiculum), (2) longitudinal (ie, long axis), and (3) radial (ie, layers and lamina, which will not be further discussed in this review). Each axis supports both gradients and domains.3,12 The transverse axis is highly conserved across mammalian species and referred to as the “tri-synaptic circuit.” 13 This cross-sectional circuit consists of the perforant path synapses from the entorhinal cortex to the dentate gyrus (DG), the mossy fiber synapses from the dentate gyrus to the CA3, and finally the Schaffer collateral synapses from the CA3 to the CA1 region, with the CA1 and the subiculum delivering hippocampal output.
Anatomical, Physiological, and Functional Variation Along the Rodent Longitudinal Axis
Molecular, Synaptic, and Connectivity Gradients
This basic tri-synaptic circuit is thought to be conserved across the curved longitudinal axis. 14 However, differences in intrinsic and extrinsic connectivity, molecular expression, and neuromodulatory receptor profiles have been described along the long axis.1–4,15 In rodents, dorsal hippocampus predominantly interacts with visuospatial, and sensorimotor networks responsible for spatial memory and navigation, whereas ventral hippocampus is coupled to affective, autonomic, and associative networks supporting emotion, salience, and context.2,3 Lesions in rat dorsal hippocampus produce spatial memory deficits, especially in detailed spatial representations. On the other hand, damage to ventral hippocampus inhibits fear-driven behaviors16,17 (Figure 1, BOTTOM).
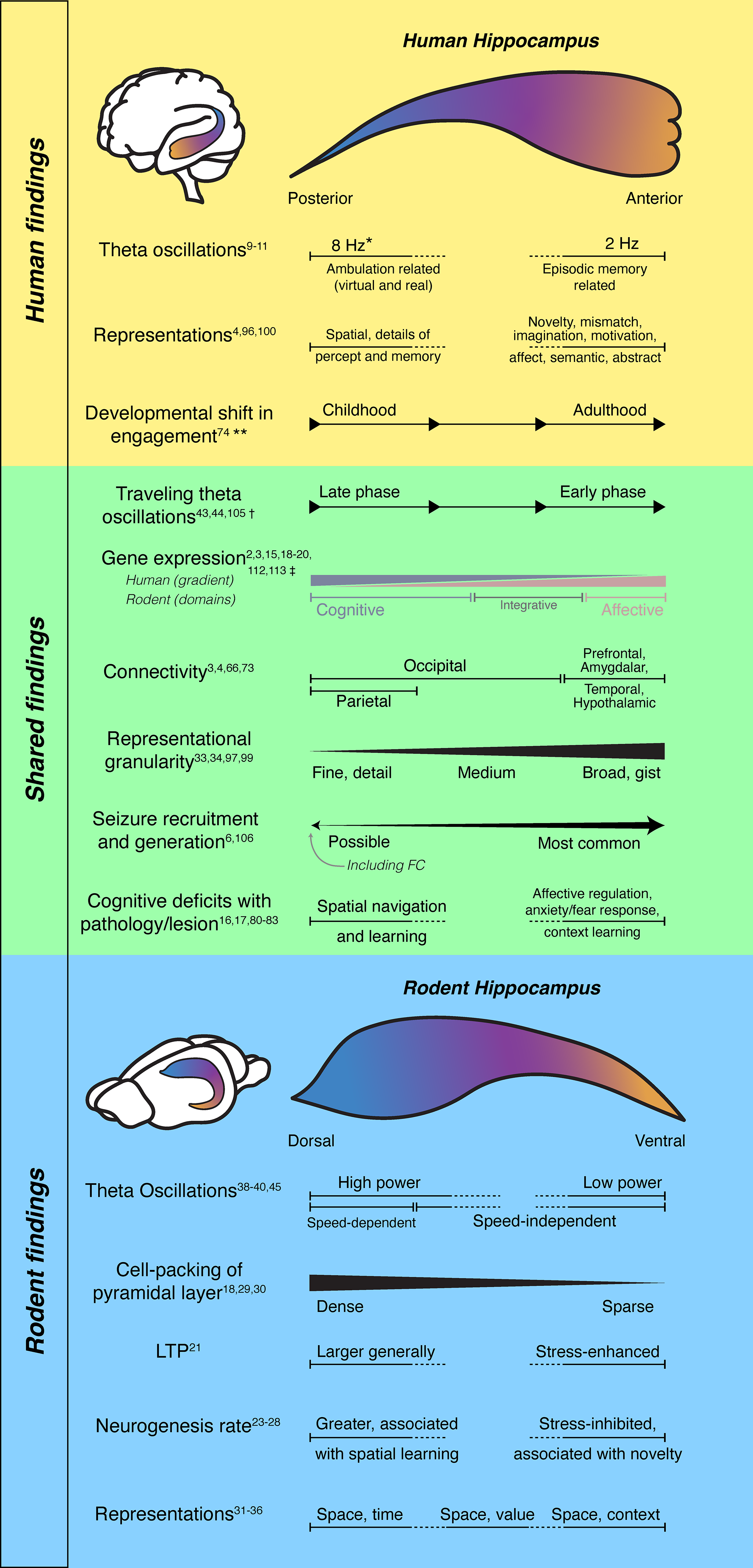
Long-axis organization in human and rodent hippocampus. the hippocampus is schematized within the human and rodent brains and unfolded along its long axis. in each schematic, color gradients run from the septal/dorsal/posterior pole (blue) to the temporal/ventral/anterior pole (orange). Functional, physiological, and anatomical features (listed on the left) are shown as varying along the hippocampal long axis. Findings derived primarily from human studies are summarized in the TOP panel, rodent studies in the BOTTOM panel, and features reported across both species in the MIDDLE panel. Horizontal bars indicate the structure and extent of variation along the long axis; graded slopes denote continuous, monotonic changes, arrows indicate direction of movement, capped, solid bars indicate discrete domains, and dotted segments indicate unclear boundaries and/or intervals that have not been tested for a given feature. *Theta frequencies are highly modulated by specific task demands. 18 ** Functional connectivity and functional association with episodic recall shift from posterior to anterior hippocampus over development.19,20 † In humans, the direction and magnitude of propagation may vary with behavior and cognitive demands. 21 ‡ Enriched expression of genes associated with cognition and affect has been shown in rodents and humans, however smooth gradients of their expression levels are present along the human long axis,22,23 while discrete gene expression domains have been identified in rodents, including a distinct integrative domain in the intermediate hippocampus. 2 .
Longitudinally organized projections from entorhinal, septal, thalamic, and hypothalamic sources, together with topographic outputs to retrosplenial and prefrontal cortex, as well as the amygdala, nucleus accumbens and other subcortical targets, establish distinct dorsal-to-ventral network affiliations within the hippocampus.3,12 Gene-expression profiling reveals molecularly distinct domains along CA1's LA,2,24 with graded differences in receptors and signaling pathways.3,15,25,26 These receptors are implicated in long-term plasticity, particularly in fear-learning contexts. In general, synapses in dorsal hippocampus undergo greater potentiation than those in ventral hippocampus in response to the same stimulation, though this effect reverses following acute stress. 27 Ventral principal cells also exhibit longer and more complex dendritic arborization. 28 Neurogenesis, which is restricted to the hippocampal DG in adult mammals, 29 occurs across the LA, typically at a higher rate in the dorsal than ventral regions. 30 Axis-specific differences in neurogenesis have been associated with distinct behavioral tendencies 31 ; for example, increased neurogenesis in dorsal and ventral DG have been linked to continued use of outdated spatial rules in reversal tasks, and reduced exploration of novel environments, respectively.32,33 Neurogenesis in ventral DG is also differentially inhibited by stress. 34 At a histological level, dorsal hippocampal pyramidal CA1 cell layers are more tightly packed compared to the more dispersed ventral counterpart.24,35,36 Although basic circuit architecture is conserved along the LA, numerous cellular, anatomical, and synaptic properties vary along this axis—some as smooth gradients and others as more discrete domains, motivating consideration of how hippocampal functions and dynamics are organized along the LA.
Representational Variation
Despite the surgical challenges of recording from rodent intermediate and ventral hippocampus, a few studies suggest a longitudinal gradient in how neural activity encodes spatial information and other variables. Compared with canonical place cells in dorsal hippocampus, intermediate CA1 contains fewer place cells, with broader place fields and reduced spatial selectivity37–40—consistent with a coarser spatial code at the population level.40,41 Intermediate CA1 neurons were shown to increase remapping in response to changes in reward location. 38 In ventral hippocampus, representations are even coarser: in ventral CA3, one study has reported neurons with very large place fields on a long linear track, 39 while another has demonstrated more abstract, spatially discontinuous representations on an 8-arm maze (eg, inward vs outward trajectories). 42 Beyond spatial coding, an odor-reward association task has shown that dorsal CA1 neurons encoded odor identities, while ventral CA1 neurons combined odor identity with predicted reward following learning. 43 Imprecise definitions of “dorsal,” “intermediate,” and “ventral” hippocampus, subfield recording sites (CA1 vs CA3), and task designs preclude a unitary model of long axis organization. Nonetheless, there is general consensus that spatial coding becomes coarser from dorsal to ventral poles.39,40 Yet, the nature of spatial representation change (eg, scale vs context), and whether these changes are gradual or abrupt (Figure 1, MIDDLE), remain unclear.
Spatiotemporal Organization in Oscillatory Dynamics
Hippocampal theta oscillations are low-frequency oscillations (8-10 Hz in rodents) seen during active behavior that increase in frequency and power with locomotion speed.44–46 Theta oscillations phase-modulate gamma oscillations and unit spiking to organize item, sequence, and content associations. 47 Theta waves travel from the dorsal to ventral pole with phase lags approaching a half-cycle (180 degrees) across the full length,48,49 although an earlier report suggests a larger full cycle shift. 50 The mechanism underlying this traveling wave is not fully understood. Theta generation depends on reciprocal connections between CA1 and the medial septum (cholinergic, glutamatergic and GABAergic septal projections to CA1, with GABAergic feedback 51 ), which are topographically organized along the LA. A phase delay across medial septal inputs could hypothetically drive theta propagation along LA. Additionally, local circuits and neuronal resonance properties at theta frequency may contribute to traveling waves. Likely, a combination of long-range drive and local resonance produces the observed propagation.51,52 Theta power and phase coherence have also been observed to decrease from dorsal to ventral poles, although these features may vary by region or behavioral context.3,49
Sharp-wave ripples (SPW-Rs) also demonstrate heterogeneous features (eg, slow-wave phase preference) along the LA.53–57 SPW-Rs are high-frequency oscillations (100-200 Hz) seen during awake rest and NREM sleep. SPW-Rs appear in the local field potential as a negative sharp-wave in the CA1 stratum radiatum followed by a burst of high-frequency oscillations in the CA1 pyramidal layer. 58 In rodents, place cell sequences acquired during exploratory behavior are replayed in SPW-Rs during quiet rest and NREM sleep.59,60 SPW-Rs may be local or global; they may propagate toward either pole, split, or remain confined locally.61,62 Together, these findings raise the question of whether the rodent hippocampus operates as a single computational module with graded parameters or as multiple, partially segregated modules that route distinct computations to different downstream targets.3,4
Translational Challenges From Rodent to Human Brain
Species differences in neuroanatomy, behavior, and recording practices complicate translation. The lissencephalic rodent brain features relatively enlarged olfactory and sensorimotor cortices. The enlarged dorsal hippocampus is surgically accessible and thus heavily studied in tasks emphasizing locomotion and spatial navigation. 63 By contrast, the human brain is over 1000 times larger than the rodent brain. The human cortex is expanded and folded, especially in association cortex (prefrontal, temporal, and parietal).64,65 These anatomical differences and their inputs to entorhinal cortex and hippocampus likely explain why the anterior hippocampus is significantly enlarged compared to the posterior end and the rodent ventral axis. Enlarged frontal and temporal cortex and their involvement in over 90% of adult focal epilepsies also explain why anterior mesial temporal structures are frequently sampled in epilepsy surgery.3,4,66 These divergent sampling strategies probably contribute to reported differences in theta-band properties, sharp-wave ripple prevalence and coupling, and place-like coding across species. Finally, a unique feature of the human brain is hemispheric specialization. Thus, human hippocampal LA functional differentiation may differ by laterality. This functional specialization is mainly driven by speech and language functions localizing to the dominant hemisphere (or the left hemisphere in 96% of right-handers 67 ).
Hippocampal Long Axis Maturation in Typically Developing Children and Children With Epilepsy
During human development, the hippocampus enlarges.68,69 Typically developing (TD) adults show a topographically preserved hippocampal gradient,70,71 with frontal and temporal inputs into the anterior hippocampus and occipital and parietal input into the posterior hippocampus. 72 In TD children, preservation of this typical pattern is associated with memory ability. 19 Functional magnetic resonance imaging (fMRI) studies have revealed that the anterior hippocampus is preferentially connected to an anterior temporal (AT) pathway 71 ; responsible for object-based memory and semantic processing. The AT network includes perirhinal cortex, entorhinal cortex, amygdala, temporal pole, orbitofrontal cortex, and anterior inferior and middle temporal gyri.71,73–78 The posterior hippocampus, the hub of the posterior medial pathway (PM), 71 is thought to be responsible for spatial and contextual memory. The PM network includes parahippocampal cortex, retrosplenial cortex, posterior cingulate, precuneus, angular gyrus, and ventral medial prefrontal cortex. 78 Between early childhood to early adulthood (4-25 years), a whole-brain connectivity shift occurs from the posterior to the anterior hippocampus. Functionally, episodic recall tends to involve posterior hippocampus in TD children, and anterior hippocampus in TD adults, 20 a difference which may be explained by an immature anterior hippocampus in the pediatric brain.
Hippocampal function is impaired in epilepsy due to primary or secondary involvement in seizure generation, interictal discharges,79,80 and neuronal dysfunction and death. Similar to adults, 70% of children with epilepsy report learning and memory deficits,81,82 which affects academic and adaptive functioning. 83 The material specificity theory posits that verbal memory is supported by the left hippocampus84,85 while visuospatial function is supported by the right hippocampus. 84 Some have argued that this functional division is overly simplistic, overlooks the complex features of everyday memory constructs, and is dependent on specific task demands. 86 Nonetheless, observational studies have generally supported that left temporal lobe epilepsy (TLE) is associated with lower verbal memory scores and right TLE with lower visual memory.87,88 However, in a meta-analysis of postsurgical outcomes, Sherman et al report that 44% of patients with a left temporal lobe resection experienced verbal memory decline, but 22% of patients with right temporal lobe epilepsy also demonstrated a verbal memory decline. Furthermore, visual memory performance declines after both left- and right-sided surgeries at equal rates (21% and 23%, respectively 89 ). These observations suggest that verbal and visual function—especially verbal function—is specialized between hemispheres, but not completely dichotomized.90,91
In children with epilepsy, normal hippocampal, and neocortical development may be disrupted—and reorganized—by ongoing seizures. Epileptogenesis and ongoing network-level dysfunction often cause functional reorganization of language, motor function, 92 and visual function. 93 Because of greater bilateral cortical representation of verbal and visuospatial functions, the left and right hippocampi may be less specialized than in adults. 71 This could explain why children do not suffer from the degree of postoperative impairment as adults. 71 Pre- and postsurgical material-specific deficits occur in pediatric TLE,94–96 but most observational studies do not show these lateralizing impairments presurgically87,97,98 or postsurgically. 99
Theta Heterogeneity Across the Long Axis in Humans
In humans, the anterior hippocampus is expanded and preferentially connected with anterior temporal and frontal association cortices, while posterior hippocampus interfaces with occipital and parietal visuospatial systems—an organization reminiscent of a coarse-to-fine representational gradient with differential roles in gist versus detailed memory and navigation.4,5,72,73,78,100–104
Human hippocampal theta has been debated because of observed variability in frequency, coherence, and task dependence across studies and, critically, across recording sites along the LA (Figure 1 TOP). Early intracranial work during virtual navigation demonstrated hippocampal theta near 8 Hz. 8 Subsequent direct recordings across a wide range of tasks and locations indicated that human hippocampal theta is often slower than in rodents, with prominent 2-4 Hz activity that can dissociate functionally and anatomically from faster 6-10 Hz components.9,11,18,105 It is unclear whether slower and faster oscillations reflect true longitudinal gradients, as intracranial electroencephalogram sampling has primarily been in hippocampal head and body, under varying task conditions. However, there is some evidence that faster ambulation is associated with higher frequency theta activity (8 Hz), akin to rodent theta activity. 18
Real-world ambulatory recordings have further shown theta bouts elicited by movement and speed. 10 Similar to rodents, theta appears to be modulated at decision points 106 and boundaries. 107 Theta bouts appear to be modulated in real-world, observed, and imagined ambulation.106,107 Consistent with rodents, human hippocampal theta can manifest as traveling waves with systematic phase gradients along the LA, although the direction and magnitude of propagation can vary with behavior and cognitive demands. 21 Reports of reduced coherence between hippocampus and surrounding medial temporal structures in some contexts may reflect heterogeneity in sampling along the LA, task-related state differences, and analytic choices.11,105 Accumulating evidence thus supports the view that distinct slow and fast theta components differentially engage the hippocampus and map onto nonspatial/associative versus spatial/precise mnemonic operations, respectively.4,18,101
Hippocampal Tail Onset as an Underrecognized Source of Seizures
Focal onset seizures arise primarily from the frontal (30%) or temporal (65%) lobes, 6 with a majority involving the anterior hippocampus. However, emerging evidence suggests that posterior hippocampal structures, including the hippocampal tail and the fasciola cinereum (FC), play an underrecognized role in seizure generation and propagation. 108 In rodents, the FC, a small strip-like region at the dorsal/posterior extreme of the hippocampal formation, shows context- and space-related coding and can be recruited during interictal and ictal events; manipulating FC excitability with modern activity-integrator tagging and optogenetic tools can modulate seizure dynamics in experimental epilepsy109–111 (Figure 1 MIDDLE). In clinical populations, case-based SEEG evidence indicates that posterior hippocampal contacts can show early ictal involvement in a subset of patients—including those with persistent seizures after anterior mesial temporal interventions—supporting the rationale for more systematic posterior sampling and, in select cases, tailored posterior ablations.66,108,112,113 Given the posterior hippocampus’ strong coupling to occipital and medial parietal networks in humans, seizures arising from or rapidly engaging the tail could present with atypical semiology and elude detection when posterior contacts are absent.100,102 Systematic anatomical targeting that includes the hippocampal tail/FC in SEEG hypotheses—guided by imaging, semiology, and network models—may improve localization and outcomes in otherwise “nonlesional” or surgically refractory cases.
Summary
Though we only partially understand the LA structure of the hippocampus in rodents and humans, cross-species similarities have emerged. Our understanding is limited due to disparate recording practices in rodents and humans (tail vs head) and differences in tasks. An important future step towards addressing this gap is designing cross-species tasks and sampling schemes that allow us to directly compare anterior/posterior in the human or primate with ventral/dorsal in the rodent and better capture LA diversity.
As emergent features of population activity, oscillatory dynamics are particularly important to understand from an LA perspective. How theta, gamma and sharp-wave ripple properties vary along the LA may provide computationally relevant gradients that relate to scale and task demands. The patterns of travel and synchrony across the LA may play an important role in hippocampal-cortical communication that occurs during memory and navigation.
Our model of hippocampal LA organization is incomplete. Some features tend toward gradual changes across the LA, such as representational granularity or cell-packing density in rodents, while others suggest distinct modules bounded by abrupt changes, such as connectivity and gene expression. For several other rodent features and many human features, sampling across the LA may provide enough data to identify variation, but insufficient coverage to determine continuous versus discrete differences. Altogether, it is not clear whether a gradient structure, a modular structure, or some combination of the two, is most accurate, nor how the structure might reconfigure with state, task, development, or disease. How the LA structure emerges through development, the adaptations induced by experience, stress and injury, as well as the underlying molecular programs that give rise to these changes are all open questions.
Basic scientific understanding of the hippocampus benefits from cross-species translation. Such translation is currently hindered by asymmetric LA sampling in humans and rodents. An understanding of rodent LA differentiation, while appreciating the nonequivalence of recording practice between species, raises exciting new translational questions. Furthermore, understanding the functional and physiological structure of the LA hippocampus will generate novel insights into disease mechanisms. The recent identification of posterior/FC-onset seizures is a clear example of how awareness of longitudinal variation can lead to meaningful clinical and behavioral hypotheses.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Esther A. and Joseph Klingenstein Fund, National Institute of Neurological Disorders and Stroke, National Institute on Deafness and Other Communication Disorders (grant number Fellowship Award in Neuroscience, K23NS093152 (LS), NS131728 (IS), NS136355 (IS), R00NS121399 (QN), R01NS127954 (AL), R61DC022830 (AL).