
Abstract
Recent reports have suggested functional interactions between the masticatory and head motor systems. However, the neurophysiological mechanisms behind these phenomena have not been thoroughly investigated. In this study, the motor unit activity was recorded bilaterally from the dorsal neck splenius muscles (SPL) while the cell bodies of trigeminal afferent fibers were stimulated at either the trigeminal mesencephalic nucleus (Me5) or the Gasserian ganglion (5Gn) in 13 barbiturate-anesthetized Wistar rats. After Me5 electrical stimulation, SPL motor unit responses were recorded ipsilaterally (n = 24), with an onset latency distribution between 1.3 and 1.8 ms. After 5Gn stimulation, SPL responses were recorded bilaterally from 18 single units, with latencies ranging from 2.2 to 2.4 ms. Our results suggest that there are at least two distinct pathways from the trigeminal afferents to the neck muscles, with significant functional differences.
INTRODUCTION
The coordination of rhythmic movements of jaw and head motor systems has been reported in both animal models (Igarashi et al., 2000; Martaresche et al., 2000; Wilga and Motta, 2000) and humans (Ferrario et al., 1997; Eriksson et al., 2000; Matsubara et al., 2002). Recently, it was also observed that reflex activities and tonic responses in neck muscles could be elicited by periodontal mechanical stimulation (Zeredo et al., 2002). These reports suggest that trigeminal sensory inputs can produce head movements or modify the neck muscle tonus in addition to the modulation of jaw movements during mastication.
Previous studies have indicated that electrical stimulation of trigeminal nerve branches, such as the infra-orbital and alveolar nerves, can produce excitation of dorsal neck motoneurons in cats (Abrahams and Richmond, 1977; Sumino and Nozaki, 1977). Although this stimulation was not necessarily noxious, the reflex responses were regarded to be of a protective nature. Later studies have reinforced the hypothesis that such a reflex mechanism might be associated with more complex behaviors like grooming, feeding, or aggression (Alstermark et al., 1992; Abrahams et al., 1993; Bermejo and Zeigler, 1999).
From neurophysiological studies, the trigemino-cervical reflex evoked by Gasserian ganglion (5Gn) stimulation has been described in a cat model (Alstermark et al., 1992). This reflex is thought to be transmitted through at least two synapses from the ipsi- and contralateral trigeminal spinal subnucleus oralis to the dorsal neck motoneurons. Recent evidence from retrograde and intracellular labeling studies in rats, nevertheless, indicates that some primary trigeminal afferents with somata in the mesencephalic trigeminal nucleus (Me5) may project monosynaptically to neck motoneurons in the cervical spinal cord (Dessem and Luo, 1999).
The presence of two distinct populations of primary afferent neurons within the trigeminal sensory system, i.e., cells with origin in the 5Gn or Me5, suggests significant functional differences despite the redundant representation of some of the innervated structures (reviewed by Lazarov, 2002). Therefore, the purpose of the present study was to compare the nature of the trigemino-cervical reflexes evoked through the trigeminal mesencephalic and spinal trigeminal sensory nuclei.
MATERIALS & METHODS
Animal Preparation
Experiments were carried out in 13 female Wistar albino rats, weighing from 250 to 350 g each. The animals were anesthetized with 60 mg/kg (i.p.) thiamylal sodium (Isozol®, Yoshitomi Pharmaceutical, Osaka, Japan). Supplemental injection of 5 mg/kg (i.p.) was administered when necessary. We monitored the level of anesthesia continuously by checking the pupil size, withdrawal, and corneal reflexes as well as heart rate. The body temperature was maintained at 37°C by means of a heating pad. The trachea was cannulated to establish artificial ventilation (3–4 mL/cycle, 70 rpm). The animals were then transferred in a prone position to a stereotaxic frame (RA-4 and EB-4, Narishige Scientific Instruments, Tokyo, Japan).
After the experiments, the brain was removed while each animal was under deep anesthesia for histological sectioning (50-μm frozen sections, cresyl violet staining), and the electrode position was confirmed by electrolytical markings (50 μA negative current for 10 sec) and signs of electrode penetration. The experimental procedure was in agreement with the Animal Care Standards of Tokyo Medical and Dental University and Nagasaki University, and had approval of the respective Animal Welfare Committees.
Stimulation and Recording
To allow for the introduction of the stimulating electrode, we incised the scalp at the midline and prepared a small aperture about 3 mm wide in the skull with a stereotaxic microengine. Monopolar tungsten microelectrodes were used (A-M Systems, Inc., Carlsborg, WA, USA; 250-μm diameter shaft with 8° tapered tip, 5 MΩ of AC impedance). The electrode was inserted into the caudal third of the Me5 by means of a stereotaxic apparatus calibrated at the cranial point bregma. Electrode coordinates were: anterior/posterior, -9.1 mm; medial/ lateral, +1.4 mm; and ventral, +6.8 mm (Paxinos and Watson, 1998). For 5Gn stimulation, the stereotaxic coordinates were: anterior/posterior, -5.7 mm; medial/lateral, +3.3 mm; and ventral, +9.3 mm (Paxinos and Watson, 1998). After the electrode was positioned, the exposed brain was covered with a thin layer of liquid paraffin so that tissue dehydration would be avoided. Rectangular negative electrical stimulation was performed at 100-μs pulse duration. Double or triple pulses were given at 300 Hz.
Recording electrodes were placed in the dorsal neck splenius muscles (SPL) on both sides. Bipolar stainless steel wires (type E-2, Narishige Scientific Instruments, Tokyo, Japan), enamel-coated except for the tips, with 100-μm diameter and 1- to 3-mm interpolar distance, were used as recording electrodes. Spike signals were amplified by a differential amplifier (DAM-80, WPI, Sarasota, FL, USA; x1000 gain, 300 Hz and 3 KHz for low and high filters, respectively).
Data Analysis
Single-unit responses were recognized according to the all-or-none principle, where spikes showed full response after above-threshold stimulus and no response at all from a sub-threshold stimulus. All data were captured by means of a CED 1401 interface (Cambridge Electronic Design, Cambridge, UK) and stored in a computer hard disc. The data were later analyzed off-line with the Spike2® software for Windows (Version 4.02a, Cambridge Electronic Design, Cambridge, UK).
RESULTS
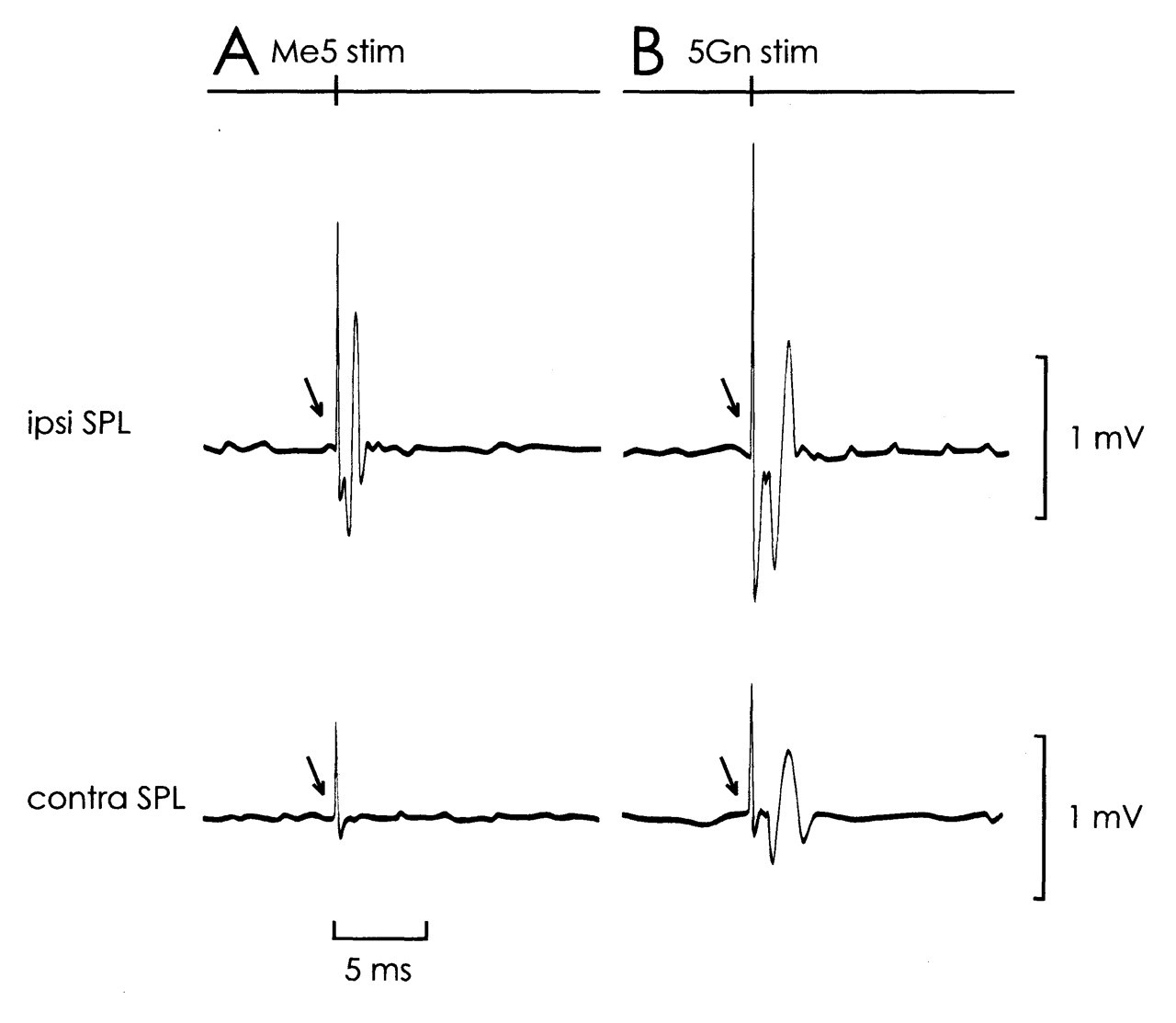
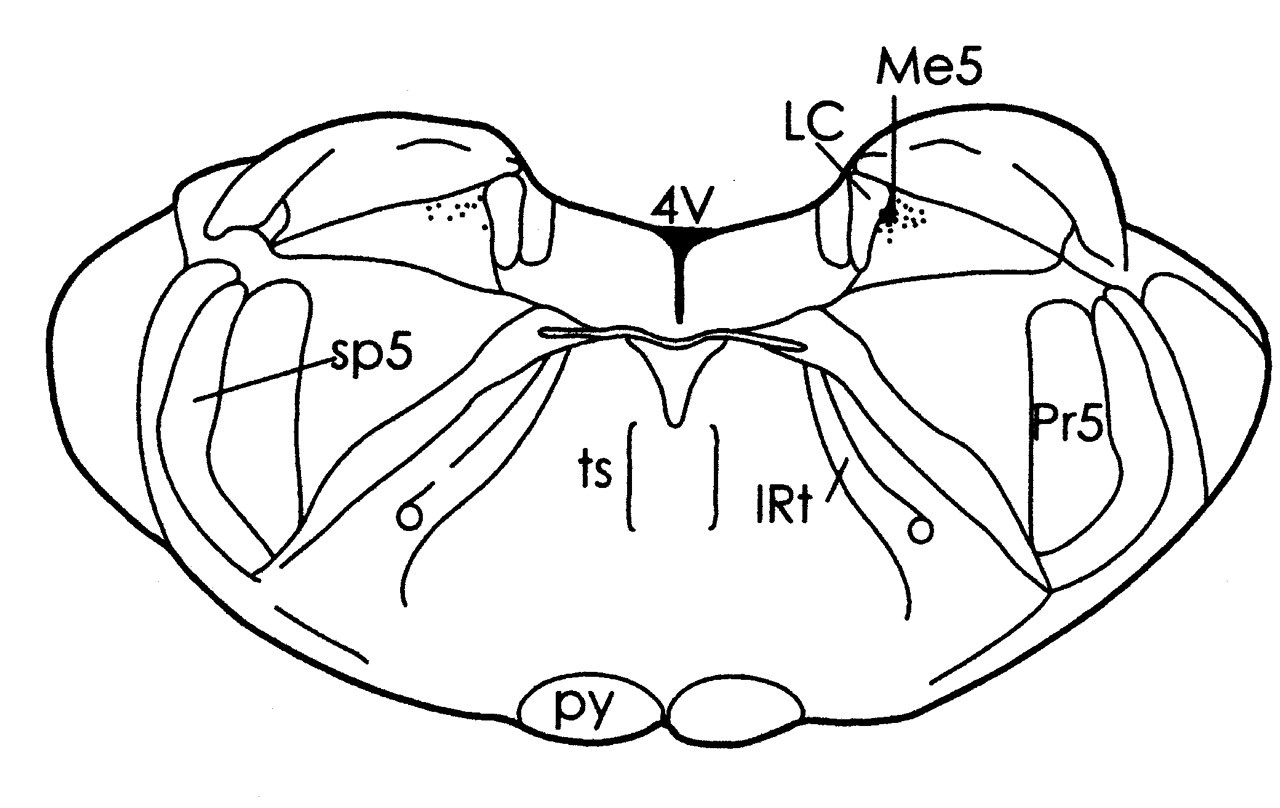
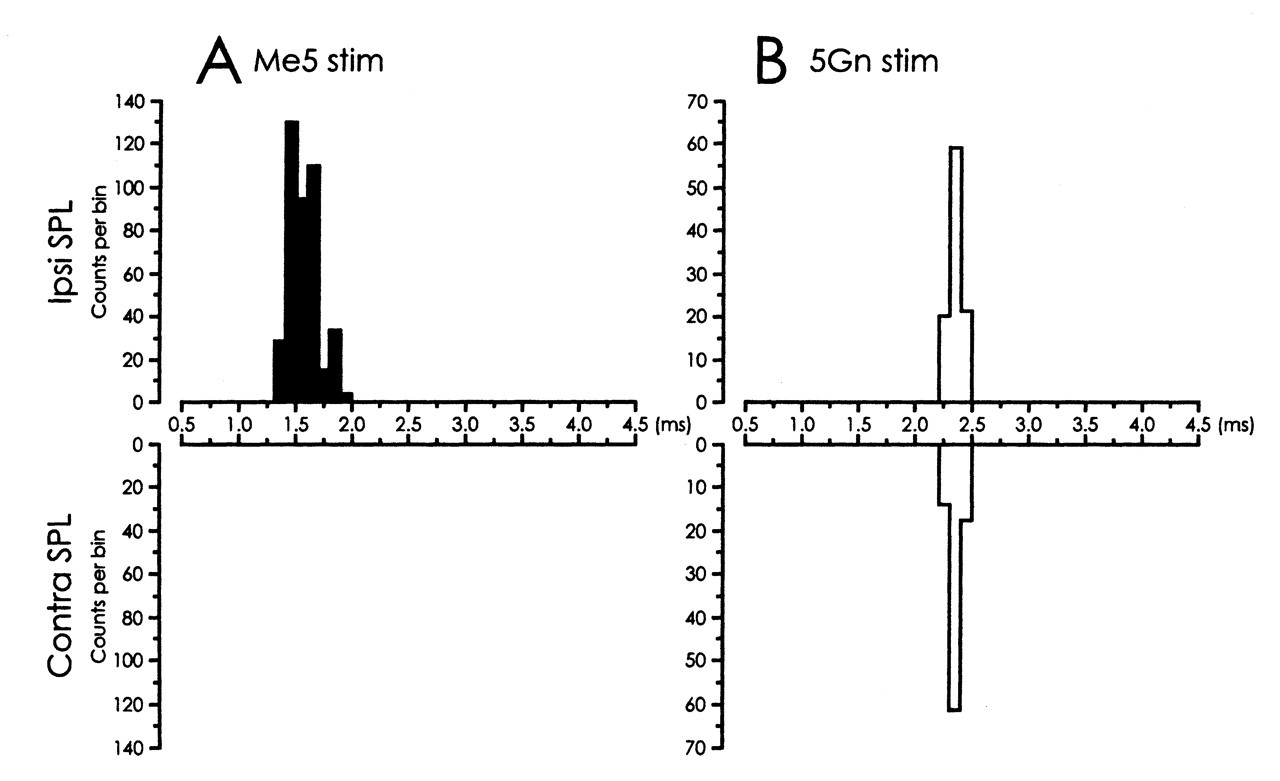
Clear SPL motor unit responses to Me5 electrical stimulation were observed ipsilaterally (Fig. 1A). Single-motor-unit spikes were recorded with an onset latency of 1.3 to 1.9 ms (1.5 ms ± 0.13 SD, 390 responses were totaled from 24 single motor units). The threshold for the ipsilateral SPL response ranged from 16 to 20 μA. For means of comparison, the ipsilateral masseter (MAS) motor unit activity was recorded following Me5 stimulation (MAS recording technique similar to that used in SPL). MAS motor units responded at about 0.5 to 0.8 times the threshold of the ipsilateral SPL, with an onset latency of 0.9 to 1.1 ms (1.0 ± 0.02 SD, 100 responses totaled from 5 single motor units). A schematic representation of the stimulated site is shown in Fig. 2.
Electrical stimulation of 5Gn elicited bilateral SPL responses. In 18 single motor units, a similar onset latency distribution was found for the ipsi- and contralateral SPL responses between 2.2 and 2.4 ms (ipsi- 2.3 ms ± 0.05 SD, 100 responses; contralateral 2.3 ms ± 0.05 SD, 53 responses). Clear ipsi- and contralateral SPL spikes were evoked at similar stimulus strengths, which varied from 30 to 50 μA (Fig. 1B). Fig. 3 shows the latency histograms for ipsi- and contralateral SPL responses after Me5 (3A) and 5Gn (3B) electrical stimulation.
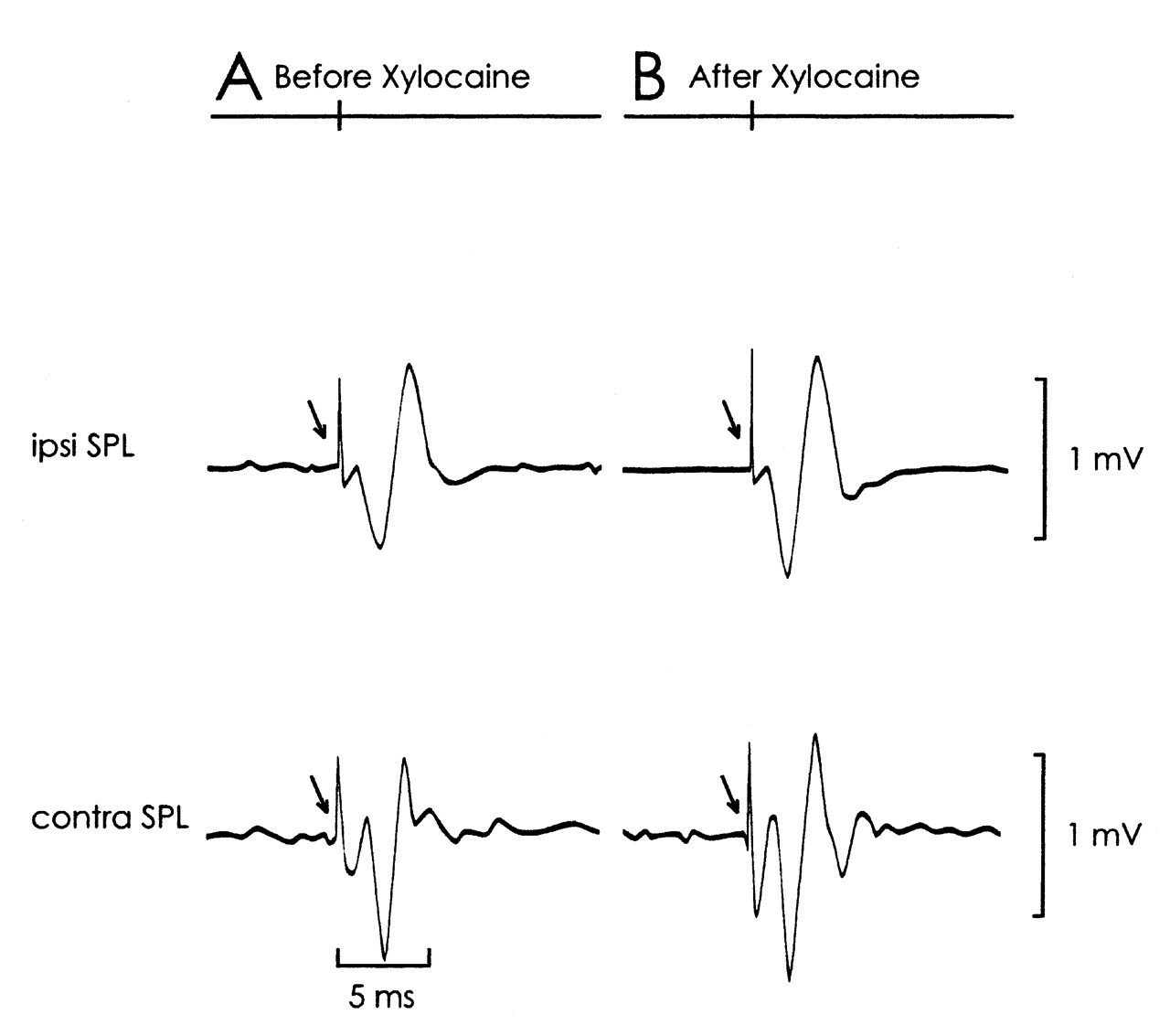
We performed a supplementary experiment to evaluate the relationships between Me5- and 5Gn-evoked trigemino-cervical reflexes. In 2 rats, 1 μL of Xylocaine 2% (mixed with histological marker Fast Green) was injected into the ipsilateral Me5 with a stereotaxically placed microsyringe (anterior/posterior -9.1 mm, medial/lateral +1.4 mm, ventral +6.8 mm). No difference in the properties of 5Gn-evoked SPL responses could be distinguished before or after Xylocaine administration (Fig. 4).
Me5 electrical stimulation failed to evoke contralateral SPL responses at low intensity levels (< 50 μA). Splenius single units failed to follow high-frequency stimulation, which implied that the responses were trans-synaptic. There was no temporal facilitation of ipsilateral SPL or masseteric responses after double or triple pulse stimulation of Me5.
DISCUSSION
The somata of most trigeminal afferent fibers lie in the 5Gn, except for afferents from muscle spindles of masticatory muscles and half the periodontal mechanoreceptors, which have their somata in the Me5 (Cody et al., 1972). Afferents from periodontal mechanoreceptors are present only in the caudal, triangular part of the nucleus (Minkels et al., 1991), where 60% of the Me5 cells are located (Rakhawy et al., 1972). In this study, we stimulated the 5Gn and the Me5 and compared the motor unit activities of the dorsal neck splenius muscles.
The latency values obtained after Me5 stimulation suggest a direct connection to the cervical spinal cord. The minimal latency of 1.3 ms for the ipsilateral SPL response would include only the conduction time to the C2/C3 level (about 14 mm), transmission in the synapses at the motoneurons, and transmission from the end plates to the muscle fibers. Of particular interest was the relative lack of contralateral response after Me5 stimulation. Given the intimate connections between Me5 afferents and the trigeminal spinal nuclei (Rokx et al., 1986; Shigenaga et al., 1988a,b, 1989; Luo et al., 2001), and the abundance of bilateral projections from these nuclei to the cervical spinal cord (Ruggiero et al., 1981; Matsushita et al., 1982; Phelan and Falls, 1991; Dessem and Luo, 1999), one would expect contralateral reflex responses in an oligosynaptic range.
Results from the 5Gn-stimulation were similar to those previously reported in cats (Alstermark et al., 1992). The average latency after 5Gn stimulation was the same for ipsi- and contralateral SPL responses, with values suggesting a disynaptic transmission. It would be unlikely to have a monosynaptic transmission, due to the lack of anatomically defined crossing projections from primary trigeminal afferents reaching the spinal cord (Matsushita et al., 1981; Dessem and Luo, 1999). The rather shorter ipsilateral latencies obtained after Me5 stimulation, however, suggest that trigeminal primary afferents may excite SPL motor units ipsilaterally through a monosynaptic pathway.
Our supplementary experiment indicated minimum interaction between the Me5- and 5Gn-evoked SPL responses, with likely independent pathways for both reflexes. Stimulation of 5Gn alone failed to evoke monosynaptic ipsilateral SPL action potentials at low intensity values, although such stimulation might have activated some of the passing fibers of Me5 afferents as well. This observation, together with those of other studies (Sumino and Nozaki, 1977; Alstermark et al., 1992; Dessem and Luo, 1999), suggests that trigeminal afferent feedback reaches the spinal cord predominantly via relays in the spinal trigeminal nuclei. Functionally, the short-latency, phasic SPL response after 5Gn stimulation might most likely be related to the head withdrawal reflex, while more complex orofacial behaviors would rely on oligo- or polysynaptic pathways according to different modalities of somatosensation.
It has been reported that neurons of the Me5 project ipsilaterally to the upper cervical segments as far as C2 or C3 (Matsushita et al., 1981). Later studies of retrograde and intracellular neuronal labeling indicated the presence of spinal-projecting collaterals of Me5 cells supplying jaw muscle spindles to the cervical spinal cord (Shigenaga et al., 1988a; Raappana and Arvidsson, 1993; Dessem and Luo, 1999).
It has been proposed that Me5 would play a role in the process of generating the central masticatory pattern (Taylor et al., 1981). Inputs from Me5, presumably jaw muscle spindles, may act as interneurons during mastication and provide positive feedback, thus increasing the cycle duration and electromyographic activity of jaw-closing muscles (Luschei and Goodwin, 1974). The Me5-evoked trigemino-cervical reflex observed in this study, and the previously reported concomitant jaw and head movements observed during fictive mastication (Igarashi et al., 2000) support the idea that the head motor system may be involved in the basic patterns of mastication generated in the brainstem.
Short-latency, phasic SPL motor unit responses evoked by Schematic representation of the Me5 drawn from coronal sections of the
brainstem, -9.1 mm posterior to bregma. An asterisk indicates the stimulated
site. 4V, fourth ventricle; IRt, intermediate reticular nucleus; LC, locus
coeruleus; Me5, trigeminal mesencephalic nucleus; Pr5, principal sensory
trigeminal nucleus; py, pyramidal tract; sp5, spinal trigeminal tract; ts,
tectospinal tract. Latency histograms of spike responses generated from SPL motor units (30 bins
of 0.1 ms each). SPL motor unit responses were evoked by 5Gn electrical stimulation
Footnotes
Acknowledgements
The authors thank Dr. E.S. Suhr for assistance in brain histology. This study was financially supported by a Grant-in-Aid for Scientific Research (no. 1387735, 2001–2002) from the Ministry of Education, Culture, Sports, Science, and Technology of Japan.